
Structure and Characteristics of Plant-Frugivore Network in an Urban Park: A Case Study in Nanjing Botanical Garden Mem. Sun Yat-Sen


Abstract
:1. Introduction
2. Materials and Methods
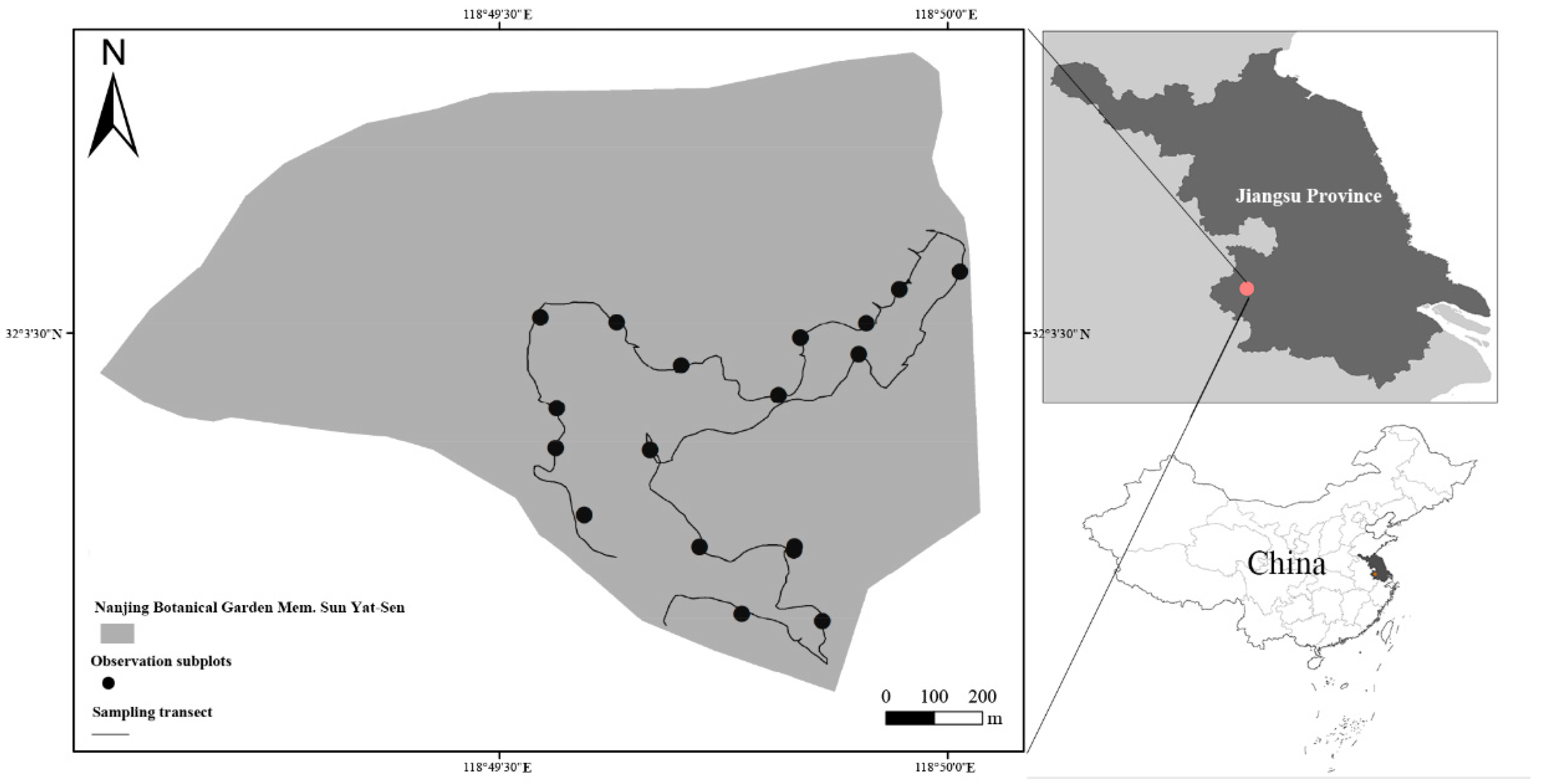
2.1. Study Site and Species
2.2. Experimental Design
2.3. Field Methods
2.3.1. Phenological Observation of Fruit Ripening
2.3.2. Plant–Frugivore Bird Network
2.4. Ecological Traits
2.5. Data Analysis
2.5.1. Network Structure
2.5.2. Network Roles of Fruit Plants and Frugivore Birds
2.5.3. Relationships between Ecological Traits and the Network Roles of Species
2.5.4. Important Species of Fruit Plants and Frugivore Birds
3. Results
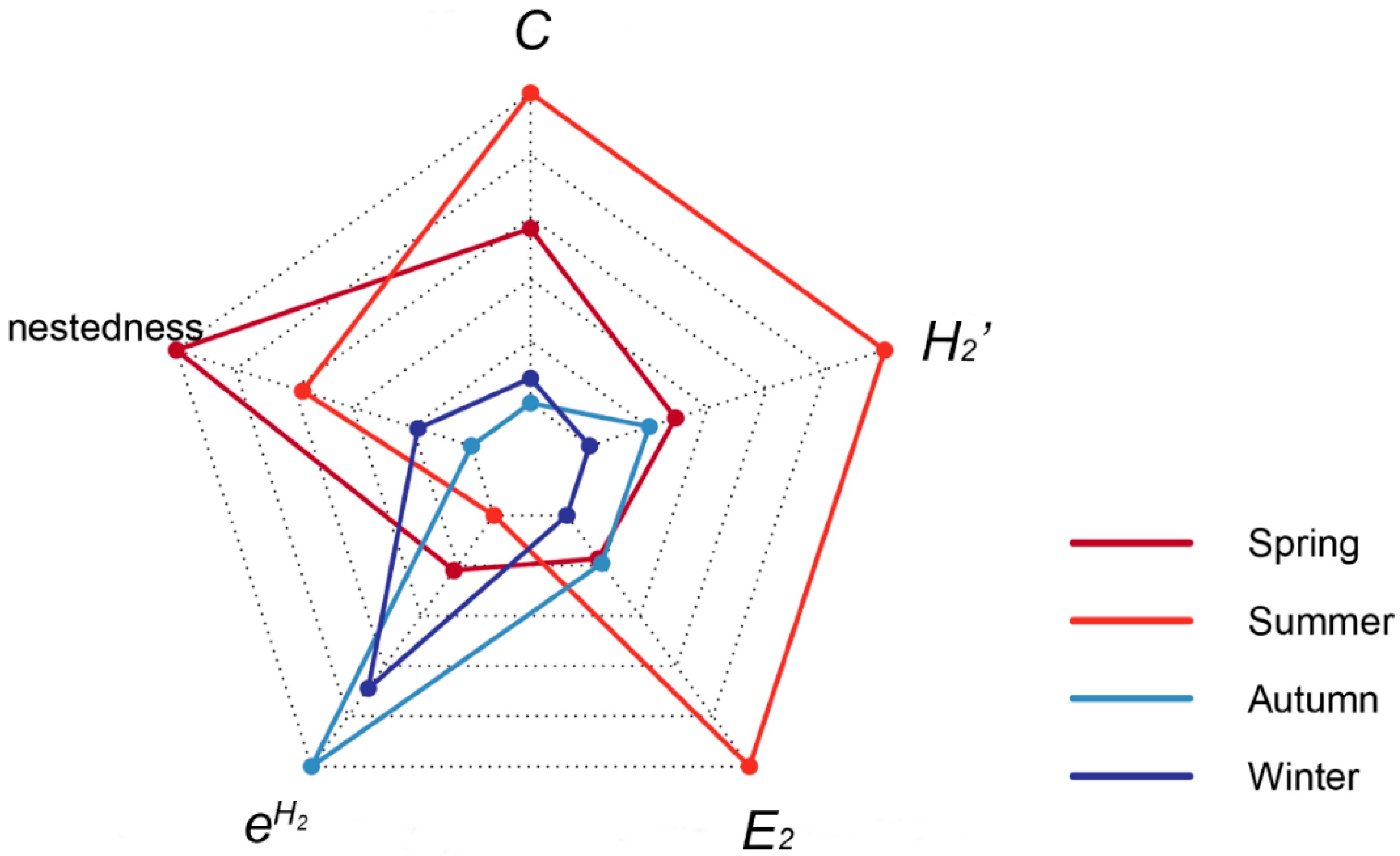
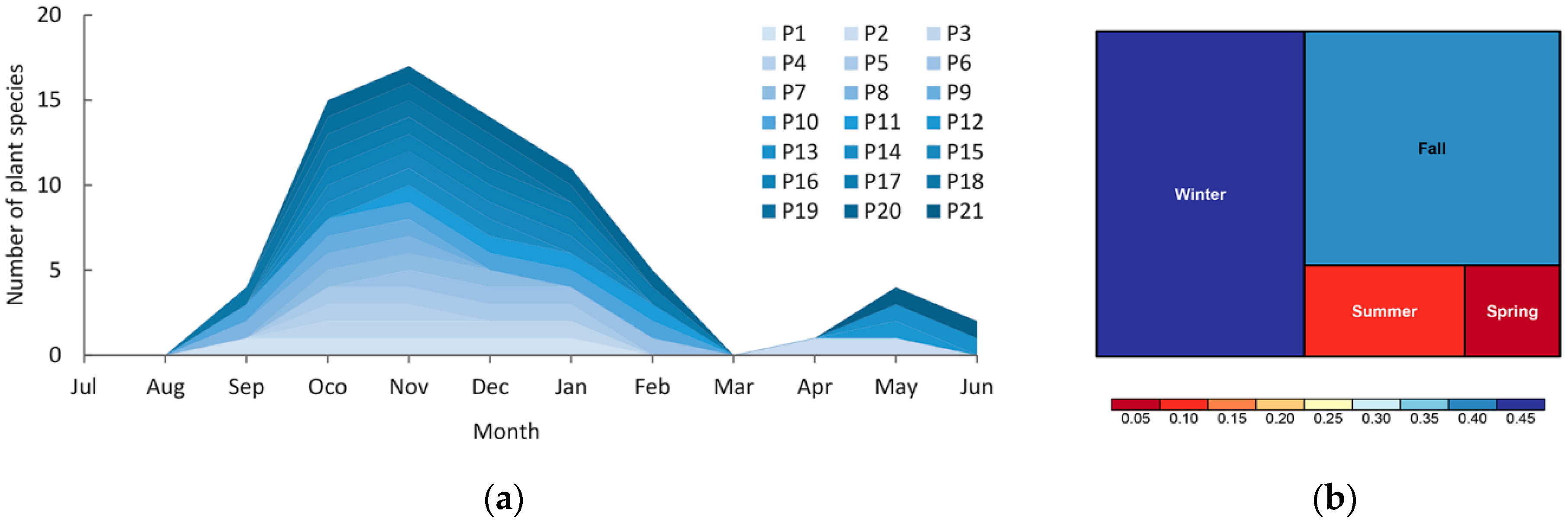
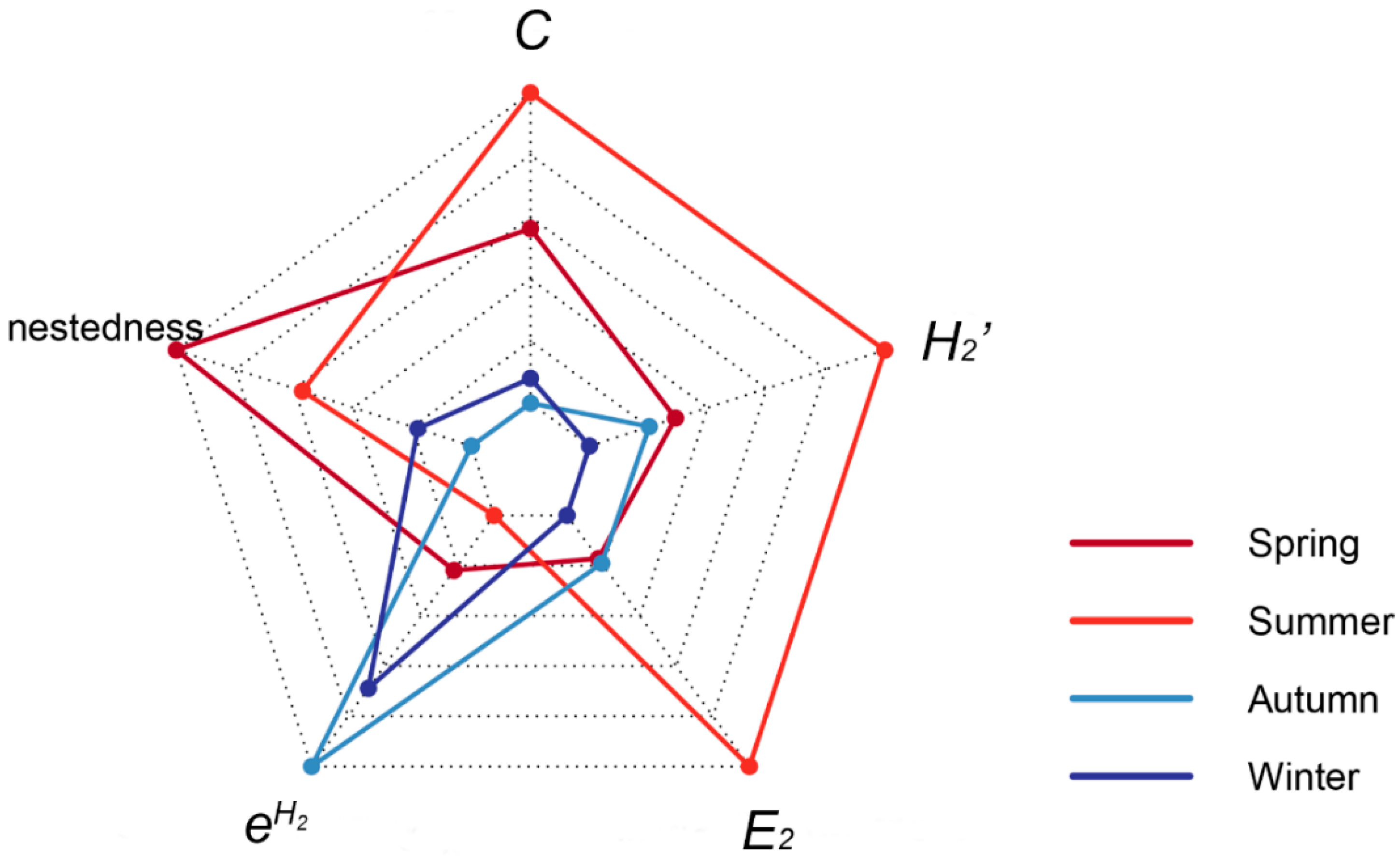
3.1. Fruit Ripening Phenology and Seasonal Plant–Frugivore Network
3.2. Year-Round Plant–Frugivore Network
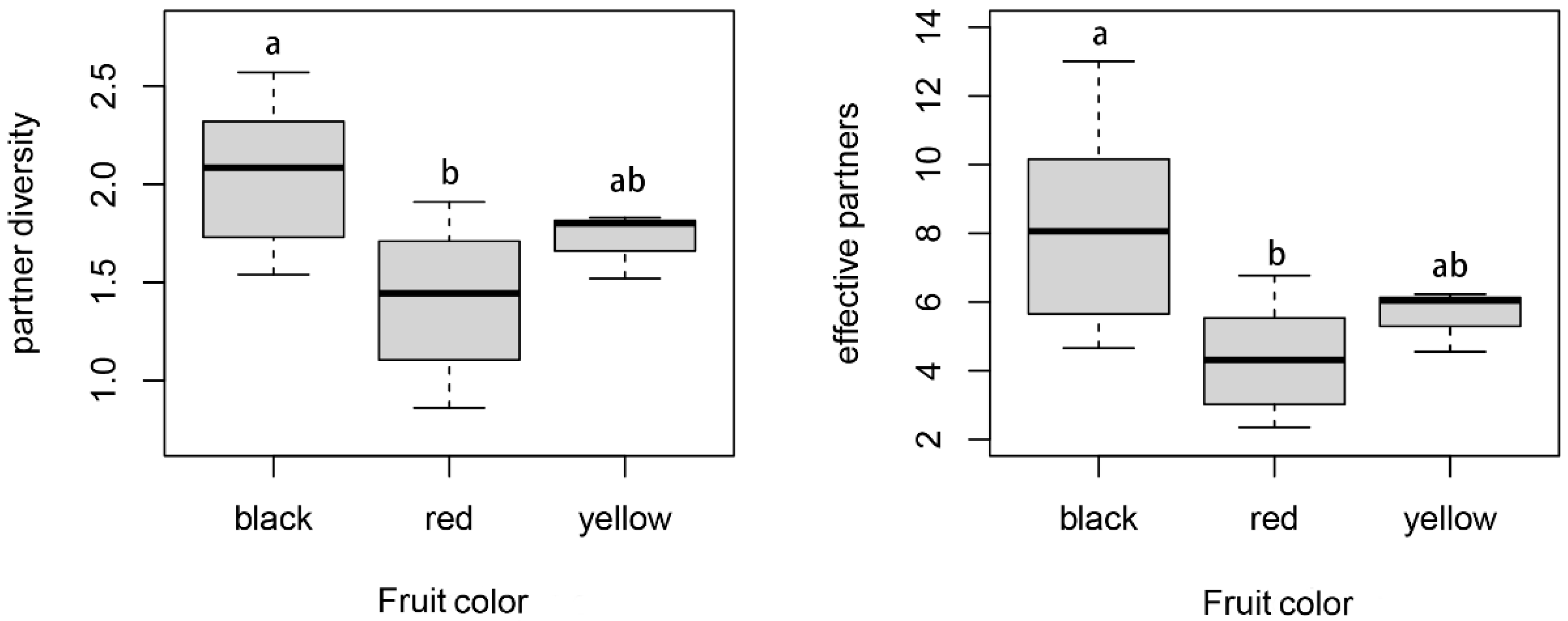
3.3. Relationship between Species’ Network Roles and Their Ecological Traits
3.4. Important Fruit Plants and Frugivore Birds in the Plant–Frugivore Network
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Palacio, F.X. Seed dispersal effectiveness by frugivorous birds: Identifying functional equivalent species in bird assemblages. Avian Biol. Res. 2019, 12, 103–108. [Google Scholar] [CrossRef]
- Valdovinos, F.S. Mutualistic networks: Moving closer to a predictive theory. Ecol. Lett. 2019, 22, 1517–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, J.M.; Murrell, D.J. The community-level consequences of seed dispersal patterns. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 549–574. [Google Scholar] [CrossRef] [Green Version]
- Bleher, B.; Bohning-Gaese, K. Consequences of frugivore diversity for seed dispersal, seedling establishment and the spatial pattern of seedlings and trees. Oecologia 2001, 129, 385–394. [Google Scholar] [CrossRef]
- Heleno, R.; Garcia, C.; Jordano, P.; Traveset, A.; Gómez, J.M.; Blüthgen, N.; Memmott, J.; Moora, M.; Cerdeira, J.; Rodríguez-Echeverría, S.; et al. Ecological networks: Delving into the architecture of biodiversity. Biol. Lett. 2014, 10, 20131000. [Google Scholar] [CrossRef] [Green Version]
- Bascompte, J.; Jordano, P. Plant-animal mutualistic networks: The architecture of biodiversity. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 567–593. [Google Scholar] [CrossRef] [Green Version]
- Garcia, D. Birds in ecological networks: Insights from bird-plant mutualistic interactions. Ardeola 2016, 63, 151–180. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Juricic, E. Bird community composition patterns in urban parks of Madrid: The role of age, size and isolation. Ecol. Res. 2000, 15, 373–383. [Google Scholar] [CrossRef]
- Mckinney, M.L. Urbanization, biodiversity, and conservation. BioScience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Kühn, I.; Klotz, S. Urbanization and homogenization—Comparing the floras of urban and rural areas in Germany. Biol. Conserv. 2006, 127, 292–300. [Google Scholar] [CrossRef]
- Rigacci, E.D.B.; Paes, N.D.; Felix, G.M.; Silva, W.R. The resilient frugivorous fauna of an urban forest fragment and its potential role in vegetation enrichment. Urban Ecosyst. 2021, 24, 943–958. [Google Scholar] [CrossRef] [PubMed]
- Bonier, F.; Martin, P.R.; Wingfield, J.C. Urban birds have broader environmental tolerance. Biol. Lett. 2007, 3, 670–673. [Google Scholar] [CrossRef] [PubMed]
- Lai, S.-M.; Liu, W.-C.; Chen, H.-W. Exploring trophic role similarity and phylogenetic relatedness between species in food webs. Community Ecol. 2021, 22, 1–14. [Google Scholar] [CrossRef]
- Schneiberg, I.; Boscolo, D.; Devoto, M.; Marcilio-Silva, V.; Dalmaso, C.A.; Ribeiro, J.W.; Ribeiro, M.C.; de Camargo Guaraldo, A.; Niebuhr, B.B.; Varassin, I.G. Urbanization homogenizes the interactions of plant-frugivore bird networks. Urban Ecosyst. 2020, 23, 457–470. [Google Scholar] [CrossRef]
- Blum, J. Urban Forests Ecosystem Services and Management; Apple Academic Press: New Jersey, NJ, USA, 2017. [Google Scholar]
- Mccleery, R.A.; Moorman, C.E.; Peterson, M.N. Urban Wildllife Conservation: Theory and Practice; Springer: New York, NY, USA, 2014. [Google Scholar]
- Beninde, J.; Veith, M.; Hochkirch, A. Biodiversity in cities needs space: A meta-analysis of factors determining intra-urban biodiversity variation. Ecol. Lett. 2015, 18, 581–592. [Google Scholar] [CrossRef]
- Farinha-Marques, P.; Lameiras, J.M.; Fernandes, C.; Silva, S.; Guilherme, F. Urban biodiversity: A review of current concepts and contributions to multidisciplinary approaches. Innov. Eur. J. Soc. Sci. Res. 2011, 24, 247–271. [Google Scholar] [CrossRef]
- Garrard, G.E.; Williams, N.S.G.; Mata, L.; Thomas, J.; Bekessy, S.A. Biodiversity Sensitive Urban Design. Conserv. Lett. 2018, 11, e12411. [Google Scholar] [CrossRef]
- Fuller, R.A.; Irvine, K.N.; Davies, Z.G.; Armsworth, P.R.; Gaston, K.J. Interactions between People and Birds in Urban Landscapes. In Urban Bird Ecology and Conservation; University of California Press: Berkeley, CA, USA, 2012; pp. 249–266. [Google Scholar]
- Niemelä, J. Ecology of urban green spaces: The way forward in answering major research questions. Landsc. Urban Plan. 2014, 125, 298–303. [Google Scholar] [CrossRef]
- Zhu, Z.-X.; Roeder, M.; Xie, J.; Nizamani, M.M.; Friedman, C.R.; Wang, H.-F. Plant taxonomic richness and phylogenetic diversity across different cities in China. Urban For. Urban Green. 2019, 39, 55–66. [Google Scholar] [CrossRef]
- Dalsgaard, B.; Schleuning, M.; Maruyama, P.K.; Dehling, D.M.; Sonne, J.; Vizentin-Bugoni, J.; Zanata, T.B.; Fjeldså, J.; Böhning-Gaese, K.; Rahbek, C. Opposed latitudinal patterns of network-derived and dietary specialization in avian plant-frugivore interaction systems. Ecography 2017, 40, 1395–1401. [Google Scholar] [CrossRef] [Green Version]
- Donoso, I.; Stefanescu, C.; Martínez-Abraín, A.; Traveset, A. Phenological asynchrony in plant-butterfly interactions associated with climate: A community-wide perspective. Oikos 2016, 125, 1434–1444. [Google Scholar] [CrossRef] [Green Version]
- Quitián, M.; Santillán, V.; Espinosa, C.I.; Homeier, J.; Böhning-Gaese, K.; Schleuning, M.; Neuschulz, E.L. Elevation-dependent effects of forest fragmentation on plant-bird interaction networks in the tropical Andes. Ecography 2018, 41, 1497–1506. [Google Scholar] [CrossRef] [Green Version]
- Gallinat, A.S.; Primack, R.B.; Lloyd-Evans, T.L. Can invasive species replace native species as a resource for birds under climate change? A case study on bird-fruit interactions. Biol. Conserv. 2020, 241, 108268. [Google Scholar] [CrossRef]
- Hagen, M.; Kissling, W.D.; Rasmussen, C.; De Aguiar, M.A.; Brown, L.E.; Carstensen, D.W.; Alves-Dos-Santos, I.; Dupont, Y.L.; Edwards, F.K.; Genini, J.; et al. Biodiversity, Species Interactions and Ecological Networks in a Fragmented World. Adv. Ecol. Res. 2012, 46, 89–210. [Google Scholar] [CrossRef] [Green Version]
- Lai, X.; Guo, C.; Xiao, Z.S. Trait-mediated seed predation, dispersal and survival among frugivore-dispersed plants in a fragmented subtropical forest, Southwest China. Integr. Zoöl. 2014, 9, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Robles, M.; Dáttilo, W.; Díaz-Castelazo, C.; Andresen, E. Fruit traits and temporal abundance shape plant-frugivore interaction networks in a seasonal tropical forest. Sci. Nat. 2018, 105, 29. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, M.C.; Schaefer, H.M.; Böhning-Gaese, K.; Schleuning, M. Importance of animal and plant traits for fruit removal and seedling recruitment in a tropical forest. Oikos 2016, 126, 823–832. [Google Scholar] [CrossRef]
- Jin, C.; Jiang, B.; Ding, Y.; Yang, S.; Xu, Y.; Jiao, J.; Huang, J.; Yuan, W.; Wu, C. Functional traits change but species diversity is not influenced by edge effects in an urban forest of Eastern China. Urban For. Urban Green. 2021, 64, 127245. [Google Scholar] [CrossRef]
- Endrédi, A.; Patonai, K.; Podani, J.; Libralato, S.; Jordán, F. Who is where in marine food webs? A trait-based analysis of network positions. Front. Mar. Sci. 2021, 8, 8. [Google Scholar] [CrossRef]
- Schleuning, M.; Neuschulz, E.L.; Albrecht, J.; Bender, I.M.; Bowler, D.E.; Dehling, D.M.; Fritz, S.; Hof, C.; Mueller, T.; Nowak, L.; et al. Trait-Based Assessments of Climate-Change Impacts on Interacting Species. Trends Ecol. Evol. 2020, 35, 319–328. [Google Scholar] [CrossRef] [PubMed]
- González, A.M.M.; Ornelas, J.F.; Dalsgaard, B.; Luna, U.M.; Lara, C. Meta-networks for the study of biogeographical traits in ecological networks: The Mexican hummingbird-plant assemblage. Sci. Nat. 2018, 105, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, F.; Xun, Y.; Cai, H.; Jin, G. Functional traits can improve our understanding of niche- and dispersal-based processes. Oecologia 2018, 186, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Bender, I.M.A.; Kissling, W.D.; Blendinger, P.G.; Böhning-Gaese, K.; Hensen, I.; Kühn, I.; Muñoz, M.C.; Neuschulz, E.L.; Nowak, L.; Quitián, M.; et al. Morphological trait matching shapes plant-frugivore networks across the Andes. Ecography 2018, 41, 1910–1919. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.; Xu, H.; Ju, F.; Liu, S.; Wan, Z. Plant diversity and its conservation strategies in Zijin Mountain National Forest Park in Nanjing. J. Jiangsu For. Sci. Technol. 2011, 38, 30–35. [Google Scholar]
- Dong, L.N.; Xu, H.B.; Liu, S.W.; Song, H.L.; Chen, X.; Ju, F. Investigation and analysis on seed plant flora at zijinshan national forest park in nanjing. J. Zhejiang For. Sci. Technol. 2010, 30, 41–47. [Google Scholar]
- Li, X.; Yin, X. Seed dispersal by birds in Nanjing Botanical Garden Mem. Sun Yat.-Sen in spering and summer. Acta Ecol. Sin. 2004, 7, 1452–1458. [Google Scholar]
- Zou, F.; Jones, H.; Colorado Z., G.J.; Jiang, D.; Lee, T.-M.; Martínez, A.; Sieving, K.; Zhang, M.; Zhang, Q.; Goodale, E. The conservation implications of mixed-species flocking in terrestrial birds, a globally-distributed species interaction network. Biol. Conserv. 2018, 224, 267–276. [Google Scholar] [CrossRef]
- Cortés-Flores, J.; Cornejo-Tenorio, G.; Urrea-Galeano, L.A.; Andresen, E.; González-Rodríguez, A.; Ibarra-Manríquez, G. Phylogeny, fruit traits, and ecological correlates of fruiting phenology in a Neotropical dry forest. Oecologia 2019, 189, 159–169. [Google Scholar] [CrossRef]
- Kolarova, E.; Nekovar, J.; Adamik, P. Long-term temporal changes in central European tree phenology (1946–2010) confirm the recent extension of growing seasons. Int. J. Biometeorol. 2014, 58, 1739–1748. [Google Scholar] [CrossRef]
- Lediuk, K.D.; Damascos, M.A.; Puntieri, J.G.; Svriz, M. Differences in phenology and fruit characteristic between invasive and native woody species favor exotic species invasiveness. Plant Ecol. 2014, 215, 1455–1467. [Google Scholar] [CrossRef]
- Neuschulz, E.L.; Botzat, A.; Farwig, N. Effects of forest modification on bird community composition and seed removal in a heterogeneous landscape in South Africa. Oikos 2011, 120, 1371–1379. [Google Scholar] [CrossRef]
- Albrecht, J.; Neuschulz, E.L.; Farwig, N. Impact of habitat structure and fruit abundance on avian seed dispersal and fruit predation. Basic Appl. Ecol. 2012, 13, 347–354. [Google Scholar] [CrossRef]
- Jordano, P. Sampling networks of ecological interactions. Funct. Ecol. 2016, 30, 1883–1893. [Google Scholar] [CrossRef]
- Zheng, G.M. A Checklist on the Classification and Distribution of the Birds of China, 3rd ed.; Science Press: Beijing, China, 2017. [Google Scholar]
- Mackinnon, J.; Phillipps, K. A Field Guide to the Birds of China; Hunan Education Press: Changsha, China, 2000. [Google Scholar]
- Zhao, Z.J. A Handbook of the Birds of China; Science and Technology Press: Jilin, China, 2001. [Google Scholar]
- Albrecht, J.; Calen, A.; Vollstädt, M.G.R.; Mayr, A.; Mollel, N.P.; Costa, D.S.; Dulle, H.I.; Fischer, M.; Hemp, A.; Howell, K.M.; et al. Plant and animal functional diversity drive mutualistic network assembly across an elevational gradient. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pigot, A.L.; Bregman, T.; Sheard, C.; Daly, B.; Etienne, R.S.; Tobias, J.A. Quantifying species contributions to ecosystem processes: A global assessment of functional trait and phylogenetic metrics across avian seed-dispersal networks. Proc. R. Soc. B Biol. Sci. 2016, 283, 1844. [Google Scholar] [CrossRef] [Green Version]
- Saavedra, F.; Hensen, I.; Beck, S.G.; Böhning-Gaese, K.; Lippok, D.; Töpfer, T.; Schleuning, M. Functional importance of avian seed dispersers changes in response to human-induced forest edges in tropical seed-dispersal networks. Oecologia 2014, 176, 837–848. [Google Scholar] [CrossRef]
- Fernández, V.P.; Fontúrbel, F.E. Temporal variation of daily activity on pollinator and frugivorous birds simultaneously interacting with a specialized mistletoe. Community Ecol. 2021, 22, 217–223. [Google Scholar] [CrossRef]
- Almeida-Neto, M.; Ulrich, W. A straightforward computational approach for measuring nestedness using quantitative matrices. Environ. Model. Softw. 2011, 26, 173–178. [Google Scholar] [CrossRef]
- Blüthgen, N.; Menzel, F.; Hovestadt, T.; Fiala, B.; Blüthgen, N. Specialization, Constraints, and Conflicting Interests in Mutualistic Networks. Curr. Biol. 2007, 17, 341–346. [Google Scholar] [CrossRef] [Green Version]
- Blüthgen, N. Why network analysis is often disconnected from community ecology: A critique and an ecologist’s guide. Basic Appl. Ecol. 2010, 11, 185–195. [Google Scholar] [CrossRef]
- Vaughan, I.P.; Gotelli, N.J.; Memmott, J.; Pearson, C.E.; Woodward, G.; Symondson, W.O.C. Econullnetr: An R package using null models to analyse the structure of ecological networks and identify resource selection. Methods Ecol. Evol. 2018, 9, 728–733. [Google Scholar] [CrossRef] [Green Version]
- Vazquez, D.P.; Aizen, M.A. Null model analyses of specialization in plant-pollinator interactions. Ecology 2003, 84, 2493–2501. [Google Scholar] [CrossRef] [Green Version]
- Bascompte, J.; Jordano, P.; Olesen, J.M. Asymmetric coevolutionary networks facilitate biodiversity maintenance. Science 2006, 312, 431–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bersier, L.F.; Banasek-Richter, C.; Cattin, M.F. Quantitative descriptors of food-web matrices. Ecology 2002, 83, 2394–2407. [Google Scholar] [CrossRef]
- Bluthgen, N.; Menzel, F.; Bluthgen, N. Measuring specialization in species interaction networks. BMC Ecol. 2006, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Tang, L.; Jia, C.; Holyoak, M.; Fründ, J.; Huang, X.; Xiao, Z. The functional roles of species in metacommunities, as revealed by metanetwork analyses of bird–plant frugivory networks. Ecol. Lett. 2020, 23, 1252–1262. [Google Scholar] [CrossRef]
- Vidal, M.M.; Hasui, É.; Pizo, M.A.; Tamashiro, J.Y.; Silva, W.R.; Guimaraes, P. Frugivores at higher risk of extinction are the key elements of a mutualistic network. Ecology 2014, 95, 3440–3447. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Albert, R.; Carlo, T.A. Transience and constancy of interactions in a plant-frugivore network. Ecosphere 2013, 4, 1–25. [Google Scholar] [CrossRef]
- Plein, M.; Längsfeld, L.; Neuschulz, E.L.; Schulthei, C.; Ingmann, L.; Töpfer, T.; Böhning-Gaese, K.; Schleuning, M. Constant properties of plant–frugivore networks despite fluctuations in fruit and bird communities in space and time. Ecology 2013, 94, 1296–1306. [Google Scholar] [CrossRef] [PubMed]
- Marjakangas, E.; Abrego, N.; Grotan, V.; de Lima, R.A.F.; Bello, C.; Bovendorp, R.S.; Culot, L.; Hasui, É.; Lima, F.; Muylaert, R.L.; et al. Fragmented tropical forests lose mutualistic plant–animal interactions. Divers. Distrib. 2020, 26, 154–168. [Google Scholar] [CrossRef] [Green Version]
- Coux, C.; Rader, R.; Bartomeus, I.; Tylianakis, J.M. Linking species functional roles to their network roles. Ecol. Lett. 2016, 19, 762–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wall, S.B.V.; Beck, M.J. A comparison of frugivory and scatter-hoarding seed-dispersal syndromes. Bot. Rev. 2012, 78, 10–31. [Google Scholar] [CrossRef]
- Yan, P.; Yang, J. Species diversity of urban forests in China. Urban For. Urban Green. 2017, 28, 160–166. [Google Scholar] [CrossRef]
- Jha, R.K.; Nölke, N.; Diwakara, B.; Tewari, V.; Kleinn, C. Differences in tree species diversity along the rural-urban gradient in Bengaluru, India. Urban For. Urban Green. 2019, 46, 126464. [Google Scholar] [CrossRef]
- Van Heezik, Y.M.; Freeman, C.; Porter, S.; Dickinson, K.J.M. Native and exotic woody vegetation communities in domestic gardens in relation to social and environmental factors. Ecol. Soc. 2014, 19, 17. [Google Scholar] [CrossRef] [Green Version]
- Aronson, M.F.J.; La Sorte, F.A.; Nilon, C.H.; Katti, M.; Goddard, M.; Lepczyk, C.A.; Warren, P.S.; Williams, N.; Cilliers, S.; Clarkson, B.; et al. A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc. R. Soc. B Boil. Sci. 2014, 281, 20133330. [Google Scholar] [CrossRef] [PubMed]
- Palacio, R.D.; Valderrama-Ardila, C.; Kattan, G.H. Generalist species have a central role in a highly diverse plant-frugivore network. Biotropica 2016, 48, 349–355. [Google Scholar] [CrossRef]
- Cristaldi, M.A.; Giraudo, A.R.; Arzamendia, V.; Bellini, G.P.; Claus, J. Urbanization impacts on the trophic guild composition of bird communities. J. Nat. Hist. 2017, 51, 2385–2404. [Google Scholar] [CrossRef]
- Shochat, E.; Warren, P.S.; Faeth, S.H.; McIntyre, N.E.; Hope, D. From patterns to emerging processes in mechanistic urban ecology. Trends Ecol. Evol. 2006, 21, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Evans, K.L.; Newson, S.E.; Gaston, K.J. Habitat influences on urban avian assemblages. Ibis 2009, 151, 19–39. [Google Scholar] [CrossRef]
- Ramos-Robles, M.; Andresen, E.; Diaz-Castelazo, C. Modularity and robustness of a plant-frugivore interaction network in a disturbed tropical forest. Ecoscience 2018, 25, 209–222. [Google Scholar] [CrossRef]
- Albrecht, J.; Berens, D.G.; Blüthgen, N.; Jaroszewicz, B.; Selva, N.; Farwig, N. Logging and forest edges reduce redundancy in plant-frugivore networks in an old-growth European forest. J. Ecol. 2013, 101, 990–999. [Google Scholar] [CrossRef]
- Buitron-Jurado, G.; Sanz, V. Specialization increases in a frugivorous bird-plant network from an isolated montane forest remnant. Community Ecol. 2020, 22, 261–274. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Species | Ratio (%) | PC1 | Species Strength | Effective Partners | Partner Diversity |
|---|---|---|---|---|---|---|
| P6 | Cinnamomum camphora | 39.12 | 26.33 | 13.11 | 10.16 | 2.32 |
| P11 | Photinia davidsoniae | 11.91 | 10.32 | 3.83 | 6.04 | 1.80 |
| P3 | Taxus chinensis | 6.12 | 6.71 | 0.70 | 2.45 | 0.90 |
| P7 | Phoebe chekiangensis | 6.80 | 5.49 | 0.84 | 5.03 | 1.61 |
| P20 | Melia azedarach | 3.75 | 3.29 | 0.23 | 3.09 | 1.13 |
| P16 | Fatsia japonica | 4.07 | 3.16 | 1.14 | 2.35 | 0.86 |
| B12 | Pycnonotus sinensis | 34.98 | 24.73 | 4.79 | 6.65 | 1.90 |
| B4 | Cyanopica cyanus | 16.29 | 10.03 | 4.09 | 8.01 | 2.08 |
| B29 | Turdus mandarinus | 16.28 | 14.48 | 2.42 | 4.67 | 1.54 |
| Fruit Plants | Group 1 (N = 6) | Group 2 (N = 15) | Differences |
| PC1 | 9.22 ± 8.79 | 1.23 ± 3.51 | U = 4, Z = −3.192, p < 0.01 |
| Species strength | 2.13 ± 2.45 | 1.75 ± 1.34 | U = 44, Z = −0.08, p = 0.97 |
| Log(Effective partners) | 0.90 ± 0.14 | 0.63 ± 0.15 | F = 0.19, T = 3.68, df = 19, p = 0.02 |
| Partner diversity | 2.32 ± 0.59 | 2.19 ± 0.48 | F = 0.23, T = 5.6, df = 19, p = 0.58 |
| Specialization (d′) | 0.09 ± 0.05 | 0.14 ± 0.10 | F = 0.01, T = −0.08, df = 19, p = 0.94 |
| Frugivore Birds | Group 1 (N = 3) | Group 2 (N = 36) | Differences |
| PC1 | 10.09 ± 10.17 | −1.48 ± 0.95 | U = 0, Z = −2.85, p < 0.001 |
| Log(Species strength) | 0.56 ± 0.15 | −1.01 ± 0.64 | F = 3.76, T = 4.367, df = 37, p < 0.01 |
| Effective partners | 8.07 ± 1.74 | 2.57 ± 1.74 | U = 7, Z = −2.49, p < 0.01 |
| Partner diversity | 2.07 ± 0.23 | 0.75 ± 0.62 | U = 7, Z = −2.49, p < 0.01 |
| Specialization (d′) | 0.12 ± 0.03 | 0.28 ± 0.15 | U = 98, Z = 2.32, p = 0.015 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Lu, C.; Han, Q.; Lu, C. Structure and Characteristics of Plant-Frugivore Network in an Urban Park: A Case Study in Nanjing Botanical Garden Mem. Sun Yat-Sen. Diversity 2022, 14, 71. https://doi.org/10.3390/d14020071
Zhang M, Lu C, Han Q, Lu C. Structure and Characteristics of Plant-Frugivore Network in an Urban Park: A Case Study in Nanjing Botanical Garden Mem. Sun Yat-Sen. Diversity. 2022; 14(2):71. https://doi.org/10.3390/d14020071
Chicago/Turabian StyleZhang, Manyu, Caihong Lu, Qian Han, and Changhu Lu. 2022. "Structure and Characteristics of Plant-Frugivore Network in an Urban Park: A Case Study in Nanjing Botanical Garden Mem. Sun Yat-Sen" Diversity 14, no. 2: 71. https://doi.org/10.3390/d14020071
APA StyleZhang, M., Lu, C., Han, Q., & Lu, C. (2022). Structure and Characteristics of Plant-Frugivore Network in an Urban Park: A Case Study in Nanjing Botanical Garden Mem. Sun Yat-Sen. Diversity, 14(2), 71. https://doi.org/10.3390/d14020071