
Plant Species Turnover on Forest Gaps after Natural Disturbances in the Dinaric Fir Beech Forests (Omphalodo-Fagetum sylvaticae)


Abstract
:1. Introduction
- − that plant species diversity would vary between specific forest gaps, depending on their age;
- − that mostly plant species from surrounding the forest will spread to the forest gaps;
- − there would be no significant appearance of invasive plants;
- − that beech would be the most common tree species in forest gaps.
2. Materials and Methods
2.1. Study Sites
2.2. Floristic Survey and Sampling
2.3. Data Analysis
3. Results
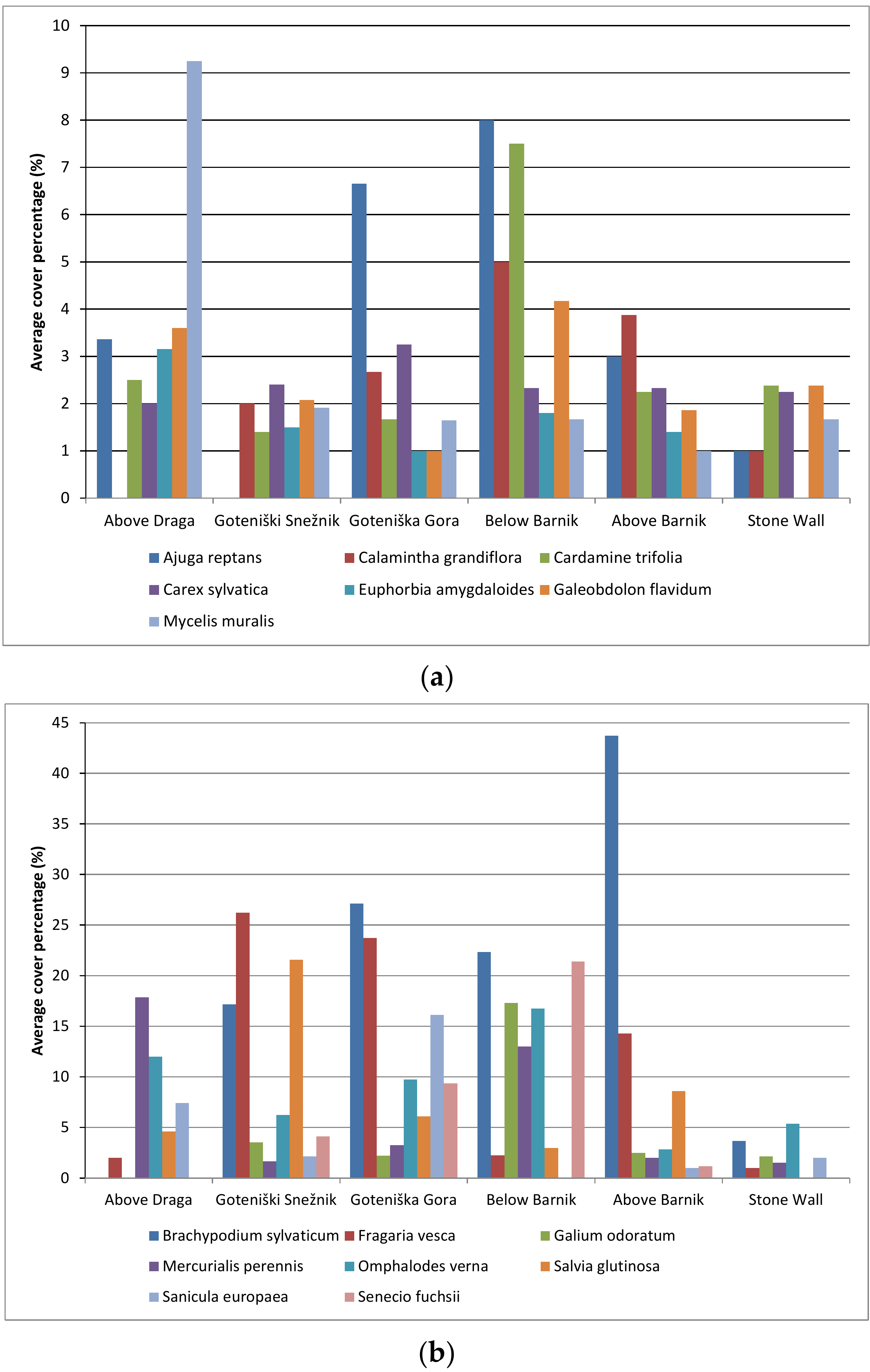
3.1. Species Richness
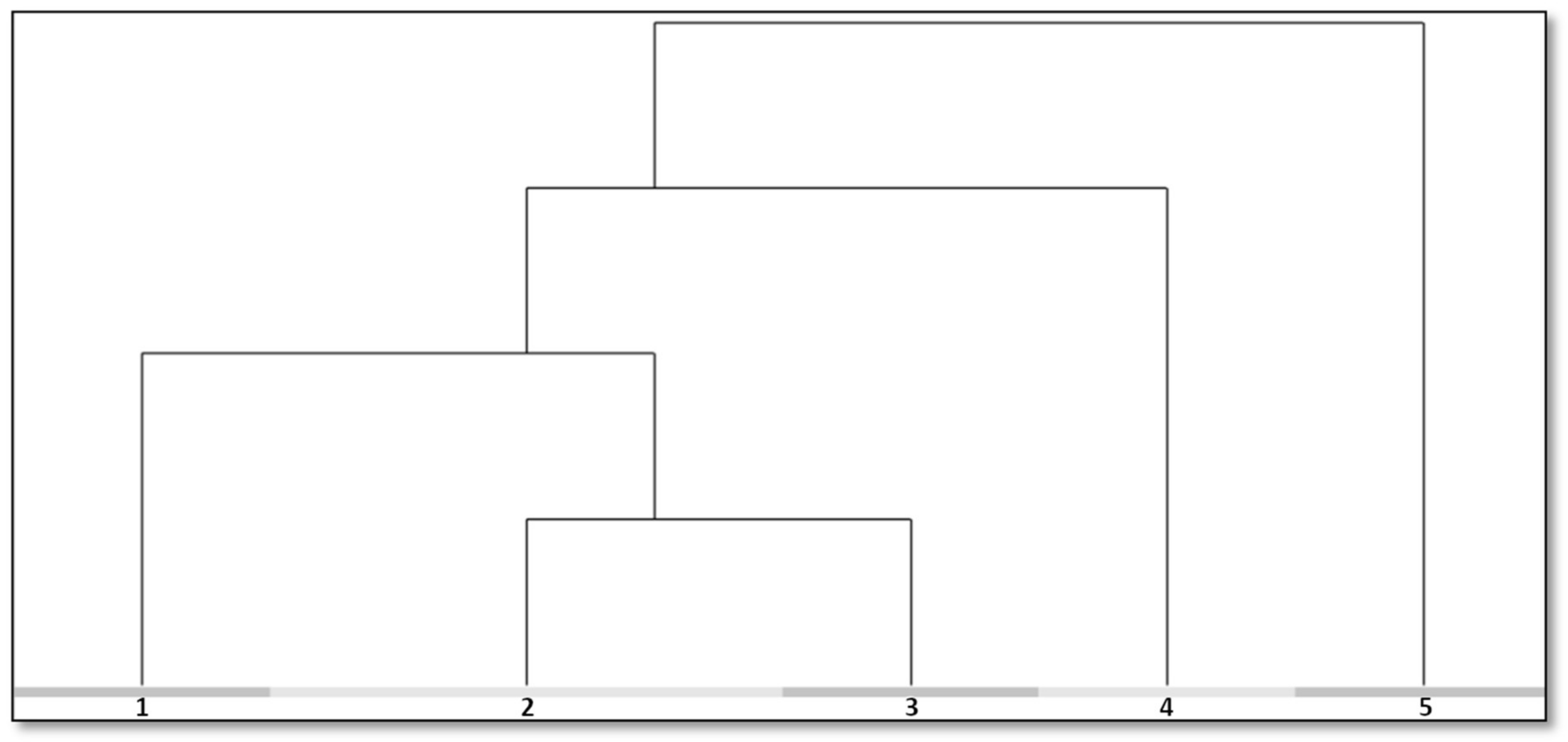
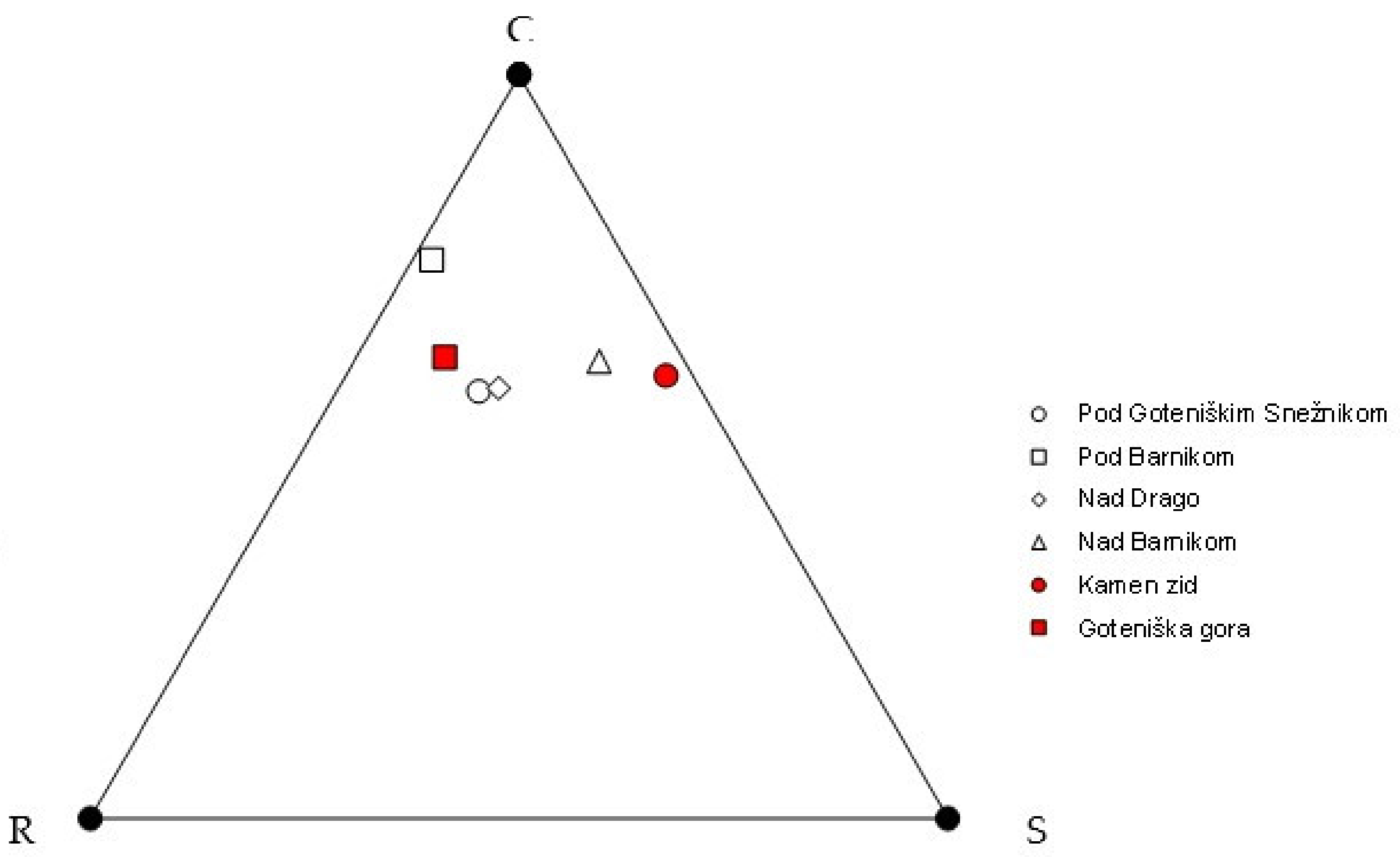
3.2. Analysis of Species Composition
3.3. Analysis of Environmental Factors and Ecological Strategy of Community
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Helms, J.A. The Dictionary of Forestry; Society of American Foresters and CABI Publishing: Bethesda, MD, USA, 1998; 210p. [Google Scholar]
- Frelich, L.E. Forest Dynamics and Disturbance Regimes; Cambridge University Press: Cambridge, UK, 2002; 90p. [Google Scholar]
- Gálhidy, L.; Mihok, B.; Hagyo, A.; Rajkai, K.; Standovár, T. Effects of gap size and associated changes in light and soil moisture on the understorey vegetation of a Hungarian beech forest. Plant Ecol. 2006, 183, 133–145. [Google Scholar] [CrossRef]
- Vilhar, U.; Roženbergar, D.; Simončič, P.; Diaci, J. Variation in irradiance, soil features and regeneration patterns in experimental forest canopy gaps. Ann. For. Sci. 2015, 72, 253–266. [Google Scholar] [CrossRef] [Green Version]
- Kermavnar, J.; Ferlan, M.; Marinšek, A.; Eler, K.; Kobler, A.; Kutnar, L. Effects of various cutting treatments and topographic factors on microclimatic conditions in Dinaric fir-beech forests. Agric. For. Meteorol. 2020, 295, 108–186. [Google Scholar] [CrossRef]
- Latif, Z.A.; Blackburn, G.A. The effects of gap size on some microclimate variables during late summer and autumn in a temperate broadleaved deciduous forest. Int. J. Biometeorolgy 2010, 54, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Seyednasrollah, B.; Kumar, M. Net radiation in a snow-covered discontinuous forest gap for a range of gap sizes and topographic configurations. J. Geophys. Res. Atmos. 2014, 10, 323–342. [Google Scholar] [CrossRef]
- O’Hara, K. Multiaged Silviculture: Managing for Complex Forest Stand Structures; Published to Oxford Scholarship Online: Oxford, UK, 2014; 262p. [Google Scholar]
- Duguid, M.C.; Frey, B.R.; Ellum, D.S.; Kelty, M.; Ashton, M.S. The influence of ground disturbance and gap position on understory plant diversity in upland forests of southern New England. For. Ecol. Manag. 2013, 303, 148–159. [Google Scholar] [CrossRef]
- Kollár, T. Light Conditions, Soil Moisture, and Vegetation Cover in Artificial Forest Gaps in Western Hungary. Acta Silv. Et. Lignaria Hung. 2017, 13, 25–40. [Google Scholar] [CrossRef] [Green Version]
- Kermavnar, J.; Eler, K.; Marinšek, A.; Kutnar, L. Post-harvest forest herb layer demography: General patterns are driven by pre-disturbance conditions. For. Ecol. Manag. 2021, 491, 119–121. [Google Scholar] [CrossRef]
- Feldmann, E.; Drößler, L.; Hauck, M.; Kucbel, S.; Pichler, V.; Leuschner, C. Canopy gap dynamics and tree understory release in a virgin beech forest, Slovakian Carpathians. For. Ecol. Manag. 2018, 415–416, 38–46. [Google Scholar] [CrossRef]
- Kermavnar, J.; Eler, K.; Marinšek, A.; Kutnar, L. Initial understorey vegetation responses following different forest management intensities in Illyrian beech forest. Appl. Veg. Sci. 2019, 22, 48–60. [Google Scholar] [CrossRef] [Green Version]
- Ravnjak, B.; Bavcon, J.; Osterc, G. Physiological response of local populations of species Cyclamen purpurascens Mill. To forest gaps. Appl. Ecol. Environ. Res. Int. Sci. J. 2019, 17, 11489–11508. [Google Scholar] [CrossRef]
- Fahey, R.T.; Puettmann, K.J. Ground-layer disturbance and initial conditions influence gap partitioning of understorey vegetation. J. Ecol. 2007, 95, 1098–1109. [Google Scholar] [CrossRef]
- Collins, B.S.; Dunne, K.P.; Pickett, S.T.A. Reponses of Forest Herbs to Canopy Gaps. In The Ecology of Natural Disturbance and patch Dynamics; Picket, S.T.A., White, P.S., Eds.; Academic Press: London, UK, 1985; pp. 217–234. [Google Scholar]
- Kelemen, K.; Mihók, B.; Gálhidy, L. Dynamic response of herbaceous vegetation to gap opening in a central European beech stand. Silva Fenn. 2012, 46, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Vukelić, J.; Mikac, S.; Baričević, D.; Šapić, I.; Bakšić, D. Vegetation and structural features of Norway spruce stands (Picea abies Karst.) in the virgin forest of Smrčeve doline in northern Velebit. Croat. J. For. Eng. J. Theory Appl. For. Eng. 2011, 32, 85–86. [Google Scholar]
- Schoennagel, T.; Smithwick, E.A.H.; Turner, M.G. Landscape heterogeneity following large fires: Insights from Yellowstone National Park, USA. Int. J. Wildland Fire 2008, 17, 742–753. [Google Scholar] [CrossRef]
- Bottero, A.; Garbarino, M.; Dukic, V.; Goveda, Z.; Lingu, E.; Nagel, T.A.; Motta, R. Gap-phase dynamics in the old-growth forest of Lom, Bosnia and Herzegovina. Silva Fenn. 2011, 45, 875–887. [Google Scholar] [CrossRef] [Green Version]
- Muscolo, M.; Bagnato, S.; Sidari, M.; Mercurio, R. A review of the roles of forest canopy gaps. J. For. Res. 2014, 25, 725–736. [Google Scholar] [CrossRef]
- Hilmers, T.; Friess, N.; Bässler, C.; Heurich, M.; Brandl, R.; Pretzsch, H.; Seidl, R.; Müller, J. Biodiversity along temperate forest succession. J. Appl. Ecol. 2018, 55, 2756–2766. [Google Scholar] [CrossRef] [Green Version]
- Grime, J.P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. Am. Nat. 1997, 3, 1169–1194. [Google Scholar] [CrossRef]
- Meterc, G.; Skudnik, M.; Jurc, M. Vpliv gospodarjenja na biotsko raznovrstnost saproksilnih hroščev [The impact of forest management to the biodiversity of saproxylic beetles]. Gozdarski Vestn. 2015, 73, 3–18. [Google Scholar]
- Honnay, O.; Hermy, M.; Coppin, P. Impact of habitat quality on forest plant species colonization. For. Ecol. Manag. 1999, 115, 157–170. [Google Scholar] [CrossRef]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flanningan, M.D.; Hanson, P.J.; Irland, L.C.; Lugo, A.E.; Peterson, C.J.; et al. Climate Change and Forest Disturbance: Climate change can effect forests by altering the frequency, intensity, duration, and timing of fire, drought, introduced species, insects and pathogen outbreaks, hurricanes, windstorms, ice storms or landslides. BioScience 2001, 51, 723–734. [Google Scholar] [CrossRef] [Green Version]
- Slovenian Environment Agency—ARSO. Podnebne Razmere v Sloveniji (Obdobje 1985–2010) [Weather Conditions in Slovenia (Period 1985–2010)]; Slovenian Environment Agency: Ljubljana, Slovenija, 2010; 27p. [Google Scholar]
- Kordiš, F. Dinarski Jelovo-Bukovi Gozdovi v Sloveniji [Dinaric Silver Fir-Beech Forests in Slovenia]; Biotechnical Faculty, Department of Forestry: Ljubljana, Slovenija, 1993; 139p. [Google Scholar]
- Nagel, T.A.; Mikac, S.; Dolinar, M.; Klopčič, M. The natural disturbance regime in forests of the Dinaric Mountains: A synthesis of evidence. For. Ecol. Manag. 2017, 388, 29–42. [Google Scholar] [CrossRef]
- Wraber, M. Pflanzengeographische Stellung und Gliederung Sloweniens. Vegetatio 1969, 17, 176–199. [Google Scholar] [CrossRef]
- Wraber, M. Vegetacija slovenskega bukovega gozda v luči ekologije in palinologije. Biološki Vestn. 1964, 12, 77–95. [Google Scholar]
- Praprotnik, N. Ilirski Florni Element v Sloveniji. Doktorska Disertacija; Univerza Edvarda Kardelja v Ljubljani, Biotehniška fakulteta, VTOZD za biologijo: Ljubljana, Slovenija, 1987; 234p. [Google Scholar]
- Marinček, L. Bukovi Gozdovi na Slovenskem [Beech Forests in Slovenia]; Delavska Enotnost: Ljubljana, Slovenija, 1987; 153p. [Google Scholar]
- Fraver, S.; Wagner, R.G.; Day, M. Dynamics od coarse woody debris following gap harvesting in the Acadian forests of central Maine, U.S.A. Can. J. For. Res. 2002, 32, 2094–2105. [Google Scholar] [CrossRef]
- Müller-Using, S.; Bartsch, N. Decay dynamic of coarse and fine woody debris of a beech (Fagus sylvatica L.) forest in Central Germany. Eur. J. For. Res. 2009, 128, 287–296. [Google Scholar] [CrossRef] [Green Version]
- Sefidi, K.; Marvie Mohadjer, M.R. 2010: Characteristics of coarse woody debris in successional stages of natural beech forests (Fagus orientalis) of Northern Iran. J. For. Sci. 2010, 56, 7–17. [Google Scholar] [CrossRef] [Green Version]
- Jakša, J. Natural disasters in Slovenian forest. Gozdraski Vestn. 2007, 65, 177–192. [Google Scholar]
- Jakša, J. Natural disasters in Slovenian forest. Gozdraski Vestn. 2007, 65, 241–256. [Google Scholar]
- Tarman, K. Osnove Ekologije in Ekologija Živali; Državna Založba Slovenije: Ljubljana, Slovenija, 1992; 547p. [Google Scholar]
- Braun-Blanquet, J. Pflanzesoziologie. Grundzüge der Vegetationskunde; Springer: Berlin/Heidelberg, Germany, 1964; 330p. [Google Scholar]
- Gilliam, F.S. The Ecological Significance of the Herbaceus Layer in Temperate Forest Ecosystems. BioScience 2007, 57, 845–858. [Google Scholar] [CrossRef]
- Tichý, L. JUICE, software for vegetation classification. J. Veg. Sci. 2002, 13, 451–453. [Google Scholar] [CrossRef]
- Zur, A.F.; Ieno, E.N.; Smith, G.M. Analysing Ecological Data; Springer: Berlin/Heidelberg, Germany, 2007; 672p. [Google Scholar]
- Pignatti, S.; Menegoni, P.; Pietrosanti, S. Valori di bioindicazione delle piante vascolari della flora d’Italia [Bioindicator values of vascular plants of the Flora of Italy]. Braun-Blanquetia 2005, 39, 1–97. [Google Scholar]
- Jarolímek, I.; Šibik, J. Diagnostic, Constant and Dominant Species of the Higher Vegetation Units in Slovakia; VEDA: Bratislava, Slovakia, 2008; 329p. [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. Biometry, 3rd ed.; W.H. Freeman: New York, NY, USA, 1995. [Google Scholar]
- Smith, R.L.; Smith, T.M. Ecology & Field Biology; Benjamin Cummings: San Francisco, CA, USA, 2001; 771p. [Google Scholar]
- Tichy, L.; Holt, J. JUICE Program for Management Analysis and Classification of Ecological Data; Vegetation Science Group, Masaryk University Brno: Brno, Czech Republic, 2006; 98p. [Google Scholar]
- Klotz, S.; Kühn, I.; Durka, W. BIOLFLOR—Eine Datenbank zu Biologisch-Ökologischen Merkmalen der Gefässpflanzen in Deutschland—Schriftenreihe für Vegetationskunde 38; Bundesamt für Naturschutz: Bonn, Germany, 2002. [Google Scholar]
- Hunt, R.; Hodgson, J.G.; Thompson, K.; Bungener, P.; Dunnett, N.P.; Askew, A.P. A new practical tool for deriving a functional signature for herbaceous vegetation. Appl. Veg. Sci. 2004, 7, 163–170. [Google Scholar] [CrossRef]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulissen, D. Zeigerwerte von Pflanzen in Mitteleuropa; Erich Goltze: Göttingen, Germany, 1992; 262p. [Google Scholar]
- Večeřa, M.; Divíšek, J.; Lenoir, J.; Jiménez-Alfaro, B.; Biurrun, I.; Knollova, I.; Agrillo, E.; Campos, J.A.; Čarni, A.; Crespo, G.; et al. Alpha diversity of vascular plants in European forests. J. Biogeogr. 2019, 46, 1919–1935. [Google Scholar] [CrossRef]
- Degen, T.; Devillez, F.; Jacquemart, A.-L. Gaps promote plant diversity in beech forests (Luzulo-Fagetum), North Vosges, France. Ann. For. Sci. 2005, 62, 429–440. [Google Scholar] [CrossRef] [Green Version]
- Šilić, Č. Šumske Zeljaste Biljke; Zavod za Udžbenike i Nastavna Sredstva: Beograd, Serbia, 1988; 271p. [Google Scholar]
- Taylor, K. Biological Flora of the British Isles: Urtica dioica L. J. Ecol. 2009, 97, 1436–1458. [Google Scholar] [CrossRef]
- Mayer, M.; Matthews, B.; Rosinger, C.; Douglas, H.S.; Godbold, L.; Katzensteiner, K. Tree regeneration retards decomposition in a temperate mountain soil after forest gap disturbance. Soil Biol. Biochem. 2017, 115, 490–498. [Google Scholar] [CrossRef]
- Čarni, A. Vegetation of trampled habitats in the Prekmurje region (NE Slovenia). Hacquetia 2005, 4, 151–159. [Google Scholar]
- Burton, J.I.; Mladenoff, D.J.; Clayton, M.K.; Jodi, A.; Forrester, J.A. The roles of environmental filtering and colonization in the fine-scale spatial patterning of ground-layer plant communities in north temperate deciduous forests. J. Ecol. 2011, 99, 764–776. [Google Scholar] [CrossRef]
- Willner, W.; Jimenez-Alfaro, B.; Agrillo, E.; Biurrun, I.; Campos, J.A.; Čarni, A.; Cassela, L.; Csiky, J.; Ćušterevska, R.; Didukh, Y.P.; et al. Classification of European beech forests: A Gordian Knot? Appl. Veg. Sci. 2017, 20, 494–512. [Google Scholar] [CrossRef]
- Zupančič, M.; Vreš, B. Phytogeographic analysis of Slovenia. Folia Biol. Geol. 2018, 59, 160–211. [Google Scholar]
- Mucina, L.; Bueltmann, H.; Dierßen, K.; Theurillat, J.P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; Gavilán, R.; et al. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- Kermavnar, J. Vplivi Gospodarjenja na Funkcionalne Lastnosti Gozdne Vegetacije in Ekološke Razmere v Dinarskih Jelovo-Bukovih Gozdovih [Impacts of Forest Management on Functional Properties of Vegetation and Ecological Conditions in the Dinaric Fir-Beech Forests]. Ph.D. Thesis, Biotechnical Faculty, University of Ljubljana, Ljubljana, Slovenia, 2021; 225p. [Google Scholar]
- Myerscough, P.J.; Whitehead, F.H. Comparative biology of Tussilago farfara L.; Chamaenerion angustifolium (L.) Scop., Epilobium montanum L. and Epilobium adenocaulon Hausskn. Gen. Biol. Germination 1965, 1, 192–210. [Google Scholar]
- Rahmonov, O.; Jędrzejko, K.; Majgier, L. The secondary succession in the area of abandoned cemeteries in northern Poland. In Landscape Ecology—Methods, Applications and Interdisciplinary Approach; Barančoková, M., Krajčí, J., Kollár, J., Belčáková, J., Eds.; Institute of Landscape Ecology, Slovac Academy of Sciences: Bratislava, Slovakia, 2010; pp. 647–657. [Google Scholar]
- Ulanova, N. The effects of windthrow on forest at different spatial scales: A review. For. Ecol. Manag. 2000, 135, 155–167. [Google Scholar] [CrossRef]
- Mollaei Darabi, S.; Kooch, Y.; Hosseini, S. Dynamic of Plant Composition and Regeneration following Windthrow in a Temperate Beech Forest. Int. Sch. Res. Not. 2014, 2014, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Marinšek, A.; Celarc, B.; Grah, A.; Kokalj, Ž.; Nagel, T.A.; Ogris, N.; Planinšek, Š.; Roženbergar, D.; Veljanovski, T.; Vochl, S.; et al. Žledolom in njegove posledice na razvoj gozdov—Pregled dosedanjih znanj [Impacts of Ice Storms on Forest Development—A Review]. Gozdarski Vestn. 2015, 73, 392–405. [Google Scholar]
- Kraigher, H.; Jurc, D.; Kalan, P.; Kutnar, L.; Levanič, T.; Rupel, M.; Smolej, I. Beech coarse woody debris characteristic in two virgin forests reserves in southern Slovenia. Zb. Gozdarstva Lesar. 2002, 69, 91–134. [Google Scholar]
- Spears, J.D.H.; Lajtha, K. The imprint of coarse woody debris on soil chemistry in the western Oregon Cascades. Biogeochemistry 2004, 71, 163–175. [Google Scholar] [CrossRef]
- Bavcon, J. Belo Cvetoče Različice v Slovenski Flori [White-Flowered Varieties in Slovenian Flora]; University Botanic Gardens Ljubljana: Ljubljana, Slovenia, 2014; 349p. [Google Scholar]
- Eler, K.; Kermavnar, J.; Marinšek, A.; Kutnar, L. 2018: Short-term changes in plant functional traits and understory functional diversity after logging of different intensities: A temperate fir-beech forest experiment. Ann. For. Res. 2018, 61, 223–242. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
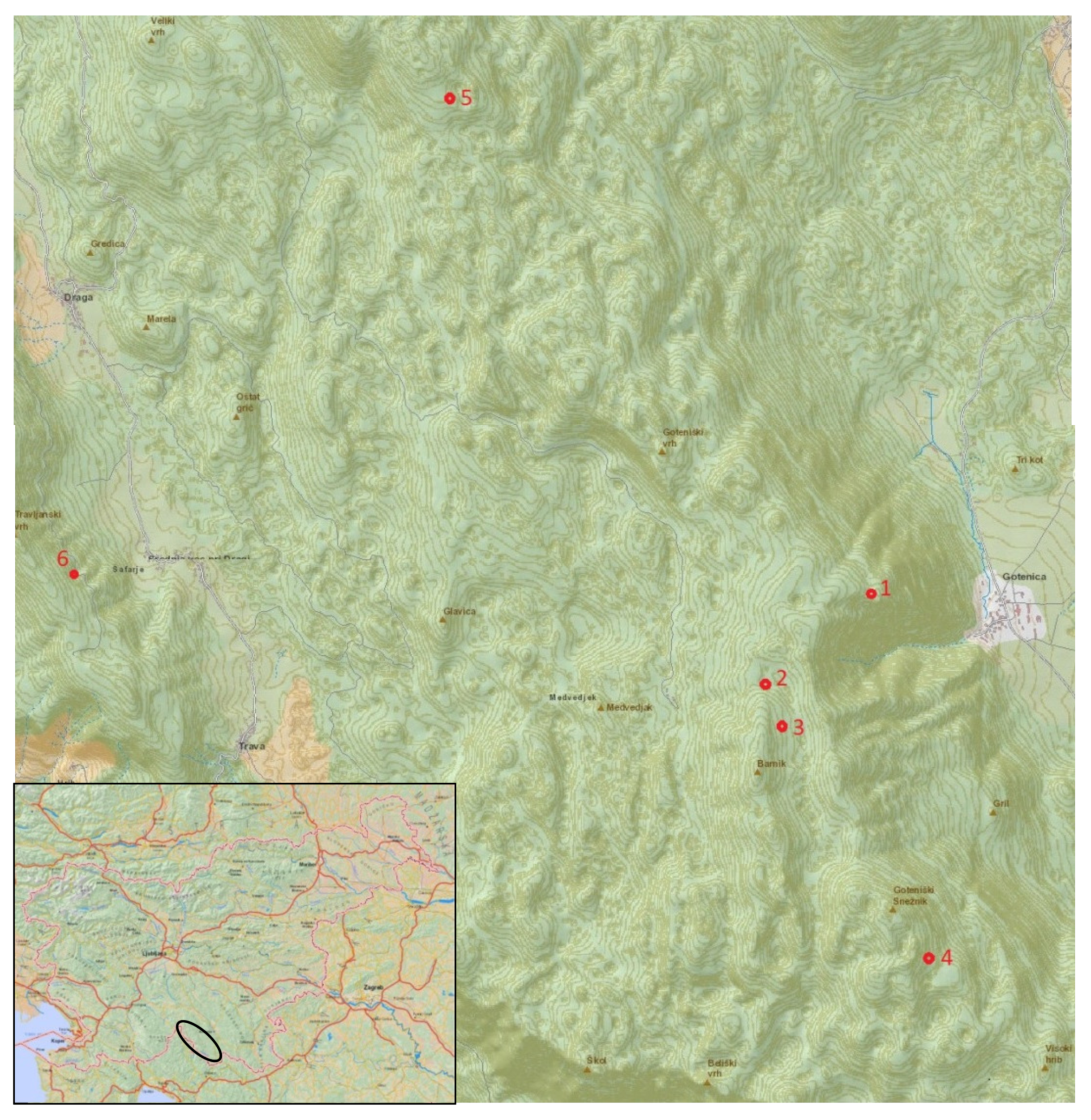
| Locality/Forest Gap | Coordinates | Altit. | Ex. | Surface | Substrate | Year |
|---|---|---|---|---|---|---|
| L1 (Stone wall) * | Y: 5479575 X: 5052223 | 1068 m | SE | 175 m2 | limestone | 2000 |
| L2 (Below Barnik) | Y: 5478774 X: 5051499 | 1132 m | SW | 600 m2 | dolomite | 2007 |
| L3 (Above Barnik) | Y: 5478904 X: 5051210 | 1161 m | SE | 1400 m2 | limestone, dolomite | 2007 |
| L4 (Goteniški Snežnik) | Y: 5480085 X: 5049403 | 1205 m | E | 1300 m2 | limestone, dolomite | 2013 |
| L5 (Goteniška gora) | X: 5055948 Y: 5476338 | 1100 m | SE | 3200 m2 | limestone, dolomite | 2011 |
| L6 (Above Draga) | Y: 5473400 X: 5052341 | 954 m | E | 1480 m2 | dolomite | 2014 |
| Locality | Floristic Survey Dates | Sampling Dates and No. of Sampling Plots |
|---|---|---|
| L1 (Stone wall) * | 24 August 2015 21 April, 7 June, 8 July, 3 August, 27 September 2016 29 March., 25 April, 24 May, 11 July 2017 | 11 July 2017/8 plots |
| L2 (Below Barnik) | 24 August 2015 21 April, 7 June, 8 July, 3 August, 27 September 2016 29 March, 25 April, 24 May, 11 July 2017 | 11 July 2017/8 plots |
| L3 (Above Barnik) | 24 August 2015 21 April, 7 June, 8 July, 3 August, 27 September 2016 29 March, 25 April, 24 May, 11 July 2017 | 11 July 2017/8 plots |
| L4 (Goteniški Snežnik) | 24 August 2015 21 April, 7 June, 8 July, 3 August, 27 September 2016 29 March, 25 April, 24 May, 20 July 2017 | 20 July 2017/14 plots |
| L5 (Goteniška gora) | 24 August 2015 21 April, 7 June, 8 July, 3 August, 27 September 2016 29 March, 25 April, 24 May,12 July 2017 | 12 July 2017/20 plots |
| L6 (Above Draga) | 24 August 2015 21 April, 7 June, 8 July, 3 August, 27 September 2016 29 March, 25 April, 24 May, 21 July 2017 | 21 July 2017/20 plots |
| L1 | L2 | L3 | L4 | L5 | L6 | |
|---|---|---|---|---|---|---|
| No. of all species per locality/forest gap | 58 | 84 | 92 | 96 | 106 | 93 |
| Only on individual locality | 2 | 9 | 9 | 8 | 13 | 23 |
| Only on sample plots | 0 | 3 | 4 | 10 | 7 | 14 |
| Only outside sample plots but at the locality | 27 | 27 | 26 | 28 | 32 | 20 |
| Species | Diagnostic | Constant | Dominant | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 1 | 2 | 3 | 4 | 5 | 6 | 1 | 2 | 3 | 4 | 5 | 6 | |
| Abies alba | * | |||||||||||||||||
| Acer pseudoplatanus | * | * | * | * | * | * | ||||||||||||
| Adenostyles glabra | * | |||||||||||||||||
| Aegopodium podagraria | * | |||||||||||||||||
| Agrimonia eupatoria | * | * | ||||||||||||||||
| Ajuga reptans | * | * | ||||||||||||||||
| Anemone nemorosa | * | |||||||||||||||||
| Asarum europaeum | * | |||||||||||||||||
| Athyrium filix-femina | * | |||||||||||||||||
| Brachypodium sylvaticum | * | * | * | * | * | * | * | * | ||||||||||
| Calamagrostis epigejos | * | * | ||||||||||||||||
| Calamintha grandiflora | * | |||||||||||||||||
| Cardamine trifolia | * | * | ||||||||||||||||
| Carex sylvatica | * | * | * | |||||||||||||||
| Circaea lutetiana | * | |||||||||||||||||
| Cyclamen purpurascens | * | |||||||||||||||||
| Digitalis ambigua | * | |||||||||||||||||
| Epimedium alpinum | * | |||||||||||||||||
| Euphorbia amygdaloides | * | * | * | |||||||||||||||
| Fagus sylvatica | * | * | * | * | * | * | * | * | * | * | * | * | ||||||
| Fragaria vesca | * | * | * | * | * | * | ||||||||||||
| Galeobdolon flavidum | * | * | * | * | ||||||||||||||
| Galium odoratum | * | * | * | * | * | * | ||||||||||||
| Hacquetia epipactis | * | * | ||||||||||||||||
| Hedera helix | * | |||||||||||||||||
| Helleborus niger | * | |||||||||||||||||
| Heracleum sphondylium | * | |||||||||||||||||
| Hypericum perforatum | * | |||||||||||||||||
| Laserpitium krapfii | * | |||||||||||||||||
| Maianthemum bifolium | * | |||||||||||||||||
| Mercurialis perennis | * | * | * | |||||||||||||||
| Mycelis muralis | * | * | * | * | ||||||||||||||
| Omphalodes verna | * | * | * | * | * | * | ||||||||||||
| Oxalis acetosella | * | |||||||||||||||||
| Petasites albus | * | * | ||||||||||||||||
| Picea abies | * | * | * | |||||||||||||||
| Polygonatum verticillatum | * | * | ||||||||||||||||
| Polystichum aculeatum | * | |||||||||||||||||
| Prunella vulgaris | * | |||||||||||||||||
| Ranunculus platanifolius | * | |||||||||||||||||
| Rosa pendulina | * | |||||||||||||||||
| Rubus idaeus | * | * | * | * | * | * | ||||||||||||
| Salvia glutinosa | * | * | * | |||||||||||||||
| Sanicula europaea | * | * | * | * | ||||||||||||||
| Scrophularia nodosa | * | * | ||||||||||||||||
| Senecio ovatus | * | * | * | * | * | * | ||||||||||||
| Tussilago farfara | * | * | ||||||||||||||||
| Urtica dioica | * | |||||||||||||||||
| Veronica officinalis | * | |||||||||||||||||
| Vicia oroboides | * | |||||||||||||||||
| Viola canina | * | |||||||||||||||||
| Average Cover in % | Maximum Cover in % |
|---|---|
| Fagus sylvatica (29%) | Fagus sylvatica (88%) |
| Brachypodium sylvaticum (24%) | Brachypodium sylvaticum (63%) |
| Tussilago farfara (20%) | Tusilago farfara (63%) |
| Epimedium alpinum (21%) | Fragaria vesca (63%) |
| Petasites albus (63%) | |
| Salvia glutinosa (63%) | |
| Senecio ovatus (63%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ravnjak, B.; Bavcon, J.; Čarni, A. Plant Species Turnover on Forest Gaps after Natural Disturbances in the Dinaric Fir Beech Forests (Omphalodo-Fagetum sylvaticae). Diversity 2022, 14, 209. https://doi.org/10.3390/d14030209
Ravnjak B, Bavcon J, Čarni A. Plant Species Turnover on Forest Gaps after Natural Disturbances in the Dinaric Fir Beech Forests (Omphalodo-Fagetum sylvaticae). Diversity. 2022; 14(3):209. https://doi.org/10.3390/d14030209
Chicago/Turabian StyleRavnjak, Blanka, Jože Bavcon, and Andraž Čarni. 2022. "Plant Species Turnover on Forest Gaps after Natural Disturbances in the Dinaric Fir Beech Forests (Omphalodo-Fagetum sylvaticae)" Diversity 14, no. 3: 209. https://doi.org/10.3390/d14030209
APA StyleRavnjak, B., Bavcon, J., & Čarni, A. (2022). Plant Species Turnover on Forest Gaps after Natural Disturbances in the Dinaric Fir Beech Forests (Omphalodo-Fagetum sylvaticae). Diversity, 14(3), 209. https://doi.org/10.3390/d14030209