
Insights into Virus–Prokaryote Relationships in a Subtropical Danshui River Estuary of Northern Taiwan in Summer



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
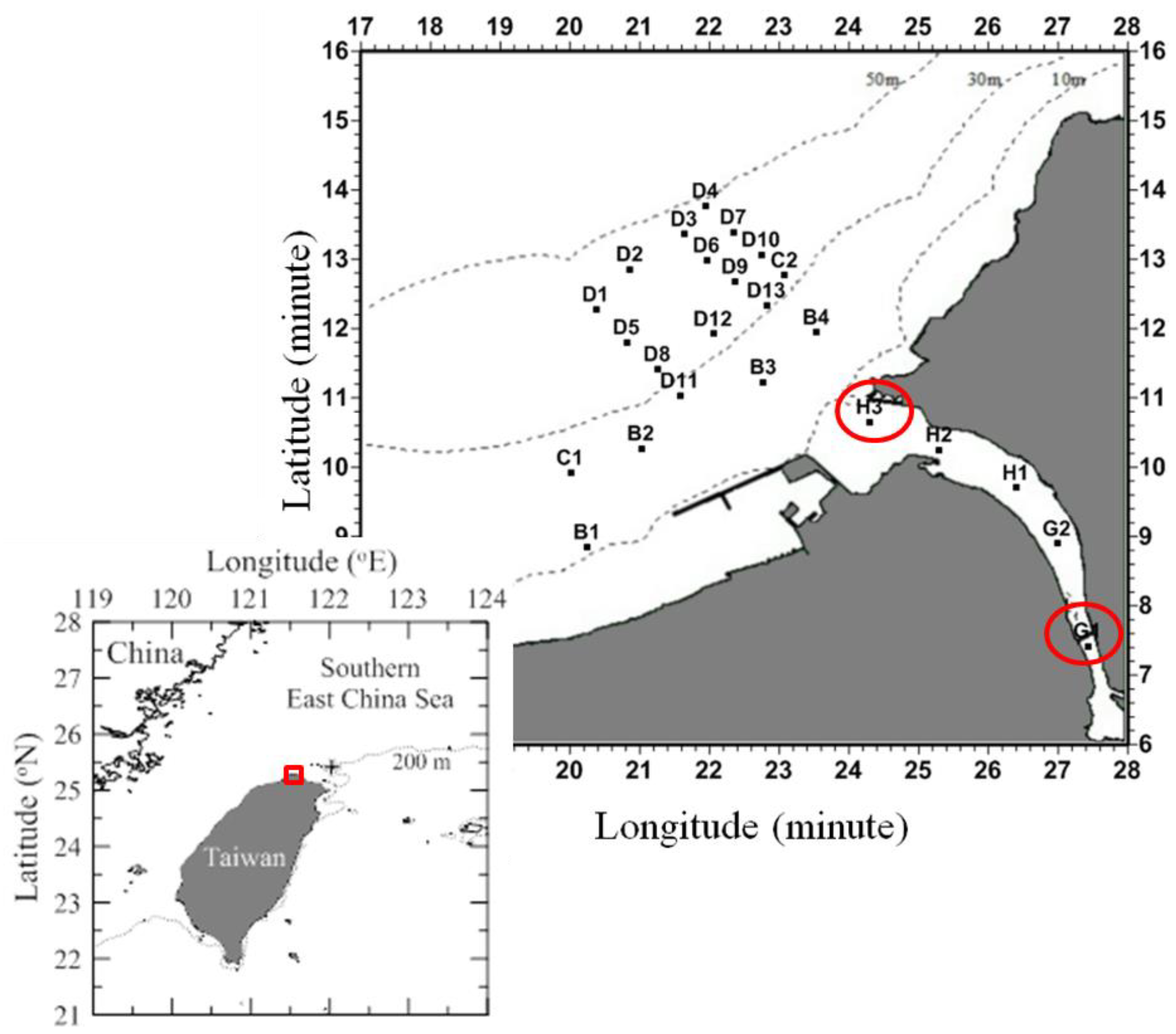
2.1. Sampling
2.2. Viral Production and Viral Decay Rate Experiments
2.3. Determination of Prokaryotic Growth Rates
2.4. Fluorescence Cytometry (FCM) for the Estimation of Viral and Prokaryotic Abundance
2.5. Statistical Analysis
3. Results
3.1. Environmental Parameters
3.2. Prokaryotic and Viral Abundance in the Surface Water
3.3. Prokaryotic Growth Rate, Viral Production, and Decay Rates
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Wommack, K.E.; Colwell, R.R. Virioplankton: Viruses in aquatic ecosystems. Microbiol. Mol. Biol. Rev. 2000, 64, 69–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinbauer, M.G. Ecology of prokaryotic viruses. FEMS Microbiol. Rev. 2004, 28, 127–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corinaldesi, C.; Rastelli, E.; Canensi, S.; Tangherlini, M.; Danovaro, R.; Dell’Anno, A. High rates of viral lysis stimulate prokaryotic turnover and C recycling in bathypelagic waters of a Ligurian canyon (Mediterranean Sea). Prog. Oceanogr. 2019, 171, 70–75. [Google Scholar] [CrossRef]
- Mathias, C.B.; Kirschner, K.T.; Velimirov, B. Seasonal variations of virus abundance and viral control of the bacterial population in backwater system of the Danube River. Appl. Environ. Microbiol. 1995, 61, 3734–3740. [Google Scholar] [CrossRef] [Green Version]
- Weinbauer, M.G.; Fuks, D.; Puskaric, S.; Peduzzi, P. Diel, seasonal and depth-related variability of viruses and dissolved DNA in the northern Adriatic Sea. Microb. Ecol. 1995, 30, 25–41. [Google Scholar] [CrossRef]
- Ogunseitan, O.A.; Sayler, G.S.; Miller, R.V. Dynamic interactions of Pseudomonas aeruginosa and bacteriophages in lake water. Microb. Ecol. 1990, 19, 171–185. [Google Scholar] [CrossRef]
- Wigington, C.H.; Sonderegger, D.; Brussaard, C.P.D.; Buchan, A.; Finke, J.F.; Fuhrman, J.A.; Lennon, J.T.; Middelboe, M.; Suttle, C.A.; Stock, C.; et al. Re-examination of the relationship between marine virus and microbial cell abundance. Nat. Microbiol. 2016, 1, 15024. [Google Scholar] [CrossRef] [Green Version]
- Parikka, K.J.; Romancer, M.L.; Wauters, N.; Jacquet, S. Deciphering the virus-to-prokaryote ratio (VPR): Insights into virus–host relationships in a variety of ecosystems. Biol. Rev. 2017, 92, 1081–1100. [Google Scholar] [CrossRef]
- Proctor, L.M.; Fuhrman, J.A. Viral mortality of marine bacteria and cyanobacteria. Nature 1990, 343, 60–62. [Google Scholar] [CrossRef]
- Clasen, J.L.; Brigden, J.S.; Payet, J.P.; Suttle, C. Evidence that viral abundance across oceans and lakes is driven by different biological factors. Freshw. Biol. 2008, 53, 1090–1100. [Google Scholar] [CrossRef]
- Jacquet, S.; Miki, T.; Noble, R.T.; Peduzzi, P.; Wilhelm, S. Viruses in aquatic ecosystems: Important advancements of the last 20 years and prospects for the future in the field of microbial oceanography and limnology. Adv. Oceanogra. Limnol. 2010, 1, 71–101. [Google Scholar] [CrossRef]
- Lin, H.J.; Shao, K.J.; Chiu, W.L.; Ma, C.J.W.; Hsieh, H.L.; Wu, W.L.; Severinhaus, L.L.; Wang, Y.T.; Chang, W.L. Biotic communities of freshwater marshes and mangroves in relation to salinity: Implications for wetland regulation. Biodiver. Conser. 2003, 12, 647–665. [Google Scholar] [CrossRef] [Green Version]
- Gong, G.C.; Shiah, F.-K.; Liu, K.-K.; Wen, Y.-H.; Liang, M.-H. Spatial and temporal variation of chlorophyll a, primary productivity and chemical hydrography in the southern East China Sea. Cont. Shelf. Res. 2000, 20, 411–436. [Google Scholar] [CrossRef]
- Noble, R.T.; Fuhrman, J.A. Viral decay and its causes in coastal waters. Appl. Environ. Microbiol. 1997, 63, 77–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mei, M.L.; Danovaro, R. Virus production and life strategies in aquatic sediments. Limnol. Oceanogr. 2004, 49, 459–470. [Google Scholar] [CrossRef]
- Brussaard, C.P.D. Optimization of procedures for counting viruses by flow cytometry. Appl. Environ. Microbiol. 2004, 70, 1506–1513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammes, F.; Egli, T. Cytometric methods for measuring bacteria in water: Advantages, pitfalls, and applications. Anal. Bioanal. Chem. 2010, 397, 1083–1095. [Google Scholar] [CrossRef]
- Day, J.W.; Hall, C.A.S.; Kemp, W.M.; Yanez-Arancibia, A. Estuarine Ecology; John Wiley and Sons: New York, NY, USA, 1989. [Google Scholar]
- Kan, J.; Crump, B.C.; Wang, K.; Chen, F. Bacterioplankton community in Chesapeake Bay: Predictable or random assemblages. Limnol. Oceanogr. 2006, 51, 2157–2169. [Google Scholar] [CrossRef] [Green Version]
- Schapira, M.; Buscot, M.J.; Leterme, S.C.; Pollet, T.; Chapperon, C.; Seuront, L. Distribution of heterotrophic bacteria and virus-like particles along a salinity gradient in a hypersaline coastal lagoon. Aquat. Microb. Ecol. 2009, 54, 171–183. [Google Scholar] [CrossRef]
- Wang, C.F.; Hsu, M.H.; Kuo, A.Y. Residence time of the Danshuei River estuary, Taiwan. Estuar. Coast. Shelf. Sci. 2004, 60, 381–393. [Google Scholar] [CrossRef]
- Shiah, F.K.; Ducklow, H.W. Temperature regulation of heterotrophic bacterioplankton abundance, production, and specific growth rate in Chesapeake Bay. Limnol. Oceanogr. 1994, 39, 1243–1258. [Google Scholar] [CrossRef]
- Fuhrman, J.A.; Noble, R.T. Viruses and protists cause similar bacterial mortality in coastal seawater. Limnol. Oceanogr. 1995, 40, 1236–1243. [Google Scholar] [CrossRef]
- Weinbauer, M.G.; Suttle, C.A. Lysogeny and prophage induction in coastal and offshore bacterial communities. Aquat. Microb. Ecol. 1999, 18, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Christaki, U.; Giannakourou, A.; Van Wambeke, F.; Gregori, G. Nanoflagellate predation on auto- and heterotrophic picoplankton in the oligotrophic Mediterranean Sea. J. Plankton. Res. 2001, 23, 1297–1310. [Google Scholar] [CrossRef] [Green Version]
- Cochlan, W.P.; Wikner, J.; Steward, G.R.; Smith, D.C.; Azam, F. Spatial distribution of viruses, bacteria and chlorophyll a in nentic, oceanic and estuarine environments. Mar. Ecol. Prog. Ser. 1993, 92, 77–87. [Google Scholar] [CrossRef]
- Bettarel, Y.; Sime-Ngando, T.; Amblard, C.; Dolan, J. Viral activity in two contrasting lake ecosystems. Appl. Environ. Microbiol. 2004, 70, 2941–2951. [Google Scholar] [CrossRef] [Green Version]
- Tsai, A.Y.; Gong, G.C.; Hung, J. Seasonal variations of virus- and nanoflagellate-mediated mortality of heterotrophic bacteria in the coastal ecosystem of subtropical western Pacific. Biogeosciences 2013, 10, 3055–3065. [Google Scholar] [CrossRef] [Green Version]
- Tsai, A.Y.; Gong, G.C.; Liu, H. Seasonal variations in virioplankton and picoplankton in semi-enclosed and open coastal waters. Terr. Atmos. Ocean. Sci. 2018, 29, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Parada, V.; Herndl, G.J.; Weinbauer, M.G. Viral burst size of heterotrophic prokaryotes in aquatic systems. J. Mar. Biol. Assoc. UK 2006, 86, 613–621. [Google Scholar] [CrossRef]
- Wilhelm, S.W.; Brigden, S.M.; Suttle, C.A. A dilution technique for the direct measurement of viral production: A comparison in stratified and tidally mixed coastal waters. Microb. Ecol. 2002, 43, 168–173. [Google Scholar] [CrossRef]
- Bettarel, Y.; Sime-Ngando, T.; Amblard, C.; Carrias, J.F.; Portelli, C. Virioplankton and microbial communities in aquatic systems: A seasonal study in two lakes of differing trophy. Freshw. Biol. 2003, 48, 810–822. [Google Scholar] [CrossRef]
- González, J.M.; Suttle, C.A. Grazing by marine nanoflagellates on viruses and virus-sized particles: Ingestion and digestion. Mar. Ecol. Prog. Ser. 1993, 94, 1–10. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, A.-Y.; Gong, G.-C.; Mukhanov, V.; Chen, P.W.-Y. Insights into Virus–Prokaryote Relationships in a Subtropical Danshui River Estuary of Northern Taiwan in Summer. Diversity 2022, 14, 237. https://doi.org/10.3390/d14040237
Tsai A-Y, Gong G-C, Mukhanov V, Chen PW-Y. Insights into Virus–Prokaryote Relationships in a Subtropical Danshui River Estuary of Northern Taiwan in Summer. Diversity. 2022; 14(4):237. https://doi.org/10.3390/d14040237
Chicago/Turabian StyleTsai, An-Yi, Gwo-Ching Gong, Vladimir Mukhanov, and Patrichka Wei-Yi Chen. 2022. "Insights into Virus–Prokaryote Relationships in a Subtropical Danshui River Estuary of Northern Taiwan in Summer" Diversity 14, no. 4: 237. https://doi.org/10.3390/d14040237
APA StyleTsai, A.-Y., Gong, G.-C., Mukhanov, V., & Chen, P. W.-Y. (2022). Insights into Virus–Prokaryote Relationships in a Subtropical Danshui River Estuary of Northern Taiwan in Summer. Diversity, 14(4), 237. https://doi.org/10.3390/d14040237