Abstract
This is the first report of the early development for the endemic Krbava minnow Delminichthys krbavensis from a karstic field in Croatia, and the first report of any species of the genus Delminichthys. The description is based on basic morphometric measures and meristic counts during a 48-day observation of embryos and larvae acquired from successful spawning and rearing in an aquarium. The mean total length of larvae at the time of hatching was 3.12 mm. Mouth opening and independent feeding and intestinal function were observed after day 11 post-hatching (PH) at which time the yolk sac was completely absorbed in all individuals. Flexion of the urostyle began after day 18 PH and was completed by day 23 PH. Pectoral fins developed on day 7 PH, pelvic fins begin to develop by day 44 PH and the formation of caudal, dorsal and anal fin was complete by day 48 PH, when individuals presented the full set of soft rays. Understanding its early development is particularly valuable in assessing the environmental impacts on this endangered species and is essential for its effective monitoring and conservation.
1. Introduction
The geographical position of Croatia encompasses the Adriatic and Black Sea (or Danube) basins, both of which have a complex geological, biological and hydrological history, resulting in one of the most diverse freshwater fish faunas in Europe, with a particularly high proportion of endemic species. As a result, Croatia is one of Europe’s most species-rich countries in fish, with 137 species that are exclusively freshwater or use freshwater periodically [1]. Cypriniformes is most species-rich order in Croatia, while the Leuciscidae has the largest species richness at the family level, and interestingly, includes the highest number of endemic taxa (21 species or 15% of all recorded taxa; [1,2]). Though Croatia possesses 52 endemic freshwater fish species, only 12 endemic taxa belong to the Danube Basin [2]. Delminichthys krbavensis (Zupančič & Bogutskaya 2002) is a stenoendemic species distributed only in Krbava Polje field [2,3] situated in the Lika region within the Danube Basin. Located in the Alpine biogeographic region and Dinaric karst area of Croatia, this region is a transitional area of the divide between the Danube and Adriatic Sea Basins, and is characterised by specific karstic landscape composed of wide fields with alluvial deposits or limestone plateaus, divided by mountain chains [1]. Hydrologically, rivers flowing through karst terrain often have springs with high discharge and eventually disappear in sinkholes on the opposite side of the field, although due to varied porosity, some areas are frequently flooded in winter, generating seasonal lakes [3].
The Krbava Polje field is a large karstic field (25 km long, 3–7 km wide, total area 120 km2) located in the central part of Lika region at an average elevation of 724 m [4]. Numerous freshwater streams and karstic wells intersect this area, and some, particularly the Krbava and Krbavica streams, are notable for their flooding nature and underground flow [5]. Presently, the Krbava Polje field is an important Natura 2000 site in Croatia (HR2000632, Krbavsko polje), with a diverse and protected biodiversity including several target species of bats, amphibians, and fish [6]. The Vukova Pećina cave in the Krbava Polje field is the type locality for the endemic D. krbavensis, demonstrating once again the natural uniqueness and global faunistic importance of this field [7,8,9].
Delminichthys krbavensis (Leuciscidae) has an extremely limited distribution range and inhabits only the small karstic springs, streams, temporary lakes and subterranean waters in the Krbava Polje field, with an area of occupancy of less than 10 km2 [2,7,8,9,10]. It prefers stagnant and clear karstic freshwaters, but when ecological conditions are unfavourable (during droughts in winter or summer), individuals of D. krbavensis retract underground [7,8,11]. Morphologically, the genus Delminichthys Freyhof, Lieckfeldt, Bogutskaya, Pitra & Ludwig 2006 is distinguished from the rest of the Leuciscidae according to the very thin, non-overlapping body scales, incomplete or interrupted lateral line, and a well-developed genital papilla in females [8,12]. Delminichthys krbavensis achieves a smaller body size than other Delminichthys spp. (less than 100 mm standard length), and can be distinguished interspecifically by the highly interrupted lateral line composed from roundish and poorly ossified small scales (usually 20–40 pored scales), irregular colour pattern with dark spots of irregular shape and size, short distance between pelvic and anal origins (usually 57% of the pectoral to pelvic origin distance) and a humped back with an elongated, shallow caudal peduncle [7,8,12]. In addition, body scales are reduced and are deeply implanted in the epidermis (i.e., they are not visible without dissection or staining), which is regarded to be an adaptation for subterranean life [11]. The mouth is subterminal while the snout is rounded and stout [7]. From the conservation standpoint, D. krbavensis is classified as a critically endangered taxa on the IUCN Red List (CR, B1ab(ii,iii,iv) + 2ab(ii,iii,iv) ver. 3.1.), both globally and locally [10,11], while it is legally protected as a strictly endangered species in Croatia [13]. Therefore, it is obvious that D. krbavensis is a highly vulnerable species, while water manipulation (extraction and melioration), pollution, road construction, and the introduction of alien fish species (such as Salmo trutta Linnaeus 1758 or Oncorhynchus mykiss (Walbaum 1792)) are all significant threats to the survival of this species in its natural habitat [8,10,11,14]. Finally, little is known about the dietary characteristics of D. krbavensis, but it is believed that this species either feeds on small aquatic invertebrates or experiences longer periods of starvation [2,11].
Although there is little information on the reproductive characteristics of D. krbavensis, it is believed that weather conditions, particularly water level fluctuations, have a significant impact on its spawning habits, with most individuals not spawning until late spring (May or July) when water levels rise due to intense floods and the ripe individuals resurface from the underground [2,7,8]. Sexual dimorphism between individuals is evident during the spawning season, especially due to breeding tubercles on the head and the pectoral region in ripe males and the large, protruding genital papilla in females [7]. Delminichthys krbavensis is a benthic spawner, with females likely laying eggs on coarse (rocky or gravel) bottoms [7,8]. Other information on life history traits or ontogeny of this fish, is currently unavailable and lacking.
The goal of this study was to give the first description of the ontogeny of aquarium-reared stenoendemic individuals of the Krbava minnow D. krbavensis, observed during the first 48 days of the post-hatching period, and to highlight the major external morphological characteristics (together with morphometric measurements and meristic counts) between the developmental stages of larvae. These findings, presented in the context of comparative biology with other endemic species, might be beneficial in the future for conservation, evolutionary, or ecological research.
2. Materials and Methods
Individuals used in this study were hatched from aquarium spawning individuals that were brought to laboratory as young of the year from the wild and were successfully raised to fertile age. There were eighth breeding individuals (three females and five males) of 50–60 mm TL. Spawning occurred on 14 March 2014 in the aquarium at a water temperature of 17 °C. The photoperiod at the time of spawning was set to 14 h daylight and 10 h night. Eggs were found attached to the sandy substrate and aquarium filter. Immediately after the eggs were observed, adult individuals were removed to a separate tank and the filter was turned off. A total of 25 eggs were extracted from the aquarium and the egg diameter was measured under a dissecting scope with a micrometre ocular lens. Upon hatching, the larva were fed decapsulated cysts of Artemia salina. The first individual hatched on 18 March and early development was observed until 4 May. Unfortunately, on that day the remaining individuals of the batch died and observation of development was not possible after 48 days post-hatching (PH). A total of 78 individuals were examined. Three random individuals were sampled on day 1 PH and five individuals were samples every three to four days for the remainder of the period (n = 75). Sampled individuals were over-anaesthetised in 300 mg/L of MS-222 solution (Sigma-Aldrich, St. Louis, Missouri, USA) and preserved in 4% formaldehyde solution. Later they were examined and measured with the ocular micrometre of the Zeiss Stemi 2000-C dissecting scope (precision to 0.01 mm) and photographed with an attached Olympus E-330 digital camera. Fourteen morphometric measures were taken (according to [8]): total length (TL), notochordal length (NL), standard length (SL), predorsal length (PreD), postdorsal length (PostD), snout length (PreO), eye diameter (ED), postorbital space (PostO), head length (HL), preventral length (PreV), preanal length (PreA), maximum body depth (H), depth of caudal peduncle (h), length of yolk sack (YSL). The examined meristic features included the number of fin rays in the dorsal, anal and caudal fins. In addition, phenotype characteristics of individuals, such as body form, pigmentation and development and disappearance of organs were also recorded.
This study was conducted entirely in accordance with the valid ethical standards and Croatian legislation.
3. Results
The fertilized eggs of D. krbavensis had a diameter of 1.65 ± 0.08 mm. Eggs were spherical, pale yellow and adhesive. Morphometric data of D. krbavensis are given in Table 1.

Table 1.
Mean values and standard deviation for TL (in mm) and percentages of morphometric measurements (%) of Delminichthys krbavensis. Percentages of morphometric measurements were calculated as a ratio of TL to NL, SL, H, h, PreA, PreD, PostD, PreV, YLS and as HL to PreO, ED and PostO.
Table 1. Mean values and standard deviation for TL and percentages of morphometric measurements (%) of Delminichthys krbavensis. Percentages of morphometric measurements were calculated as a ratio of TL to NL, SL, H, h, PreA, PreD, PostD, PreV, YLS and as HL to PreO, ED and PostO.
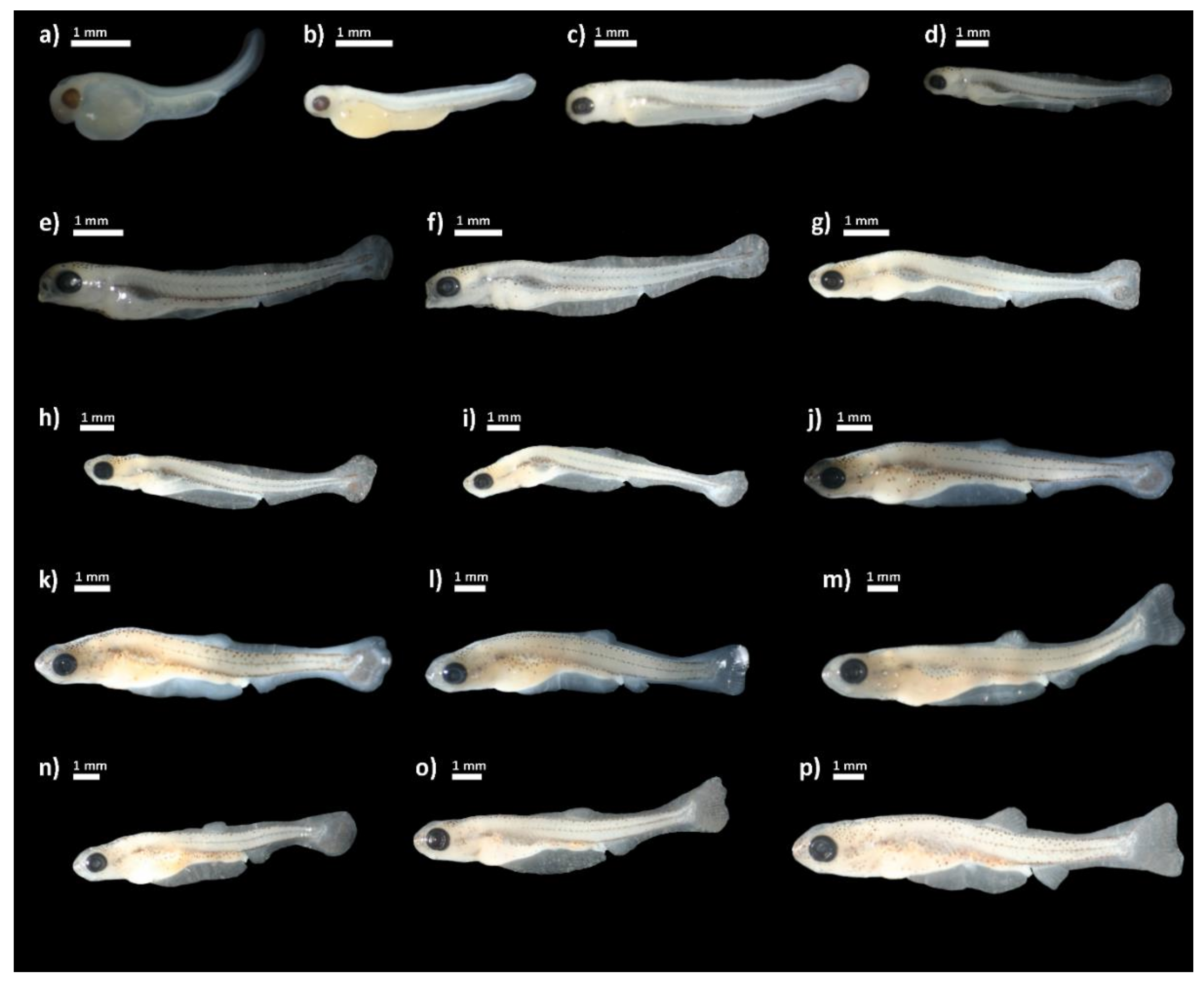
The development of individuals after hatching was examined over 48 days, during which we measured certain morphometric measures (Figure 1). The total length of newly hatched individuals (Figure 2a) was 3.12 ± 0.67 mm. Upon hatching, the mouth was not open. The yolk sack was large, extending from the head covering over two thirds of the body and measuring 2.18 ± 0.16 mm in length. Eyes were rounded with a visible lens. The post-anal finfold was present on the tail. Pigmentation was scarce in form of star shaped melanophores scattered along lateral sides of the body. On day 4 post-hatching (PH) (Figure 2b), the total length of hatched individuals was 4.67 ± 0.10 mm and the median finfold was extended further on the dorsal side reaching well over the centre of the body. Pigmentation increased and melanophores were visible laterally along the notochord and ventrally delineated the yolk sack. The yolk sack was more extended but reduced in vertical diameter. The mouth and anus were still closed. The head was nearly aligned with the horizontal axis of the body. On day 7 PH (Figure 2c), the mean total length further increased (6.27 ± 0.13 mm) and the head was fully aligned with horizontal axis of the body.
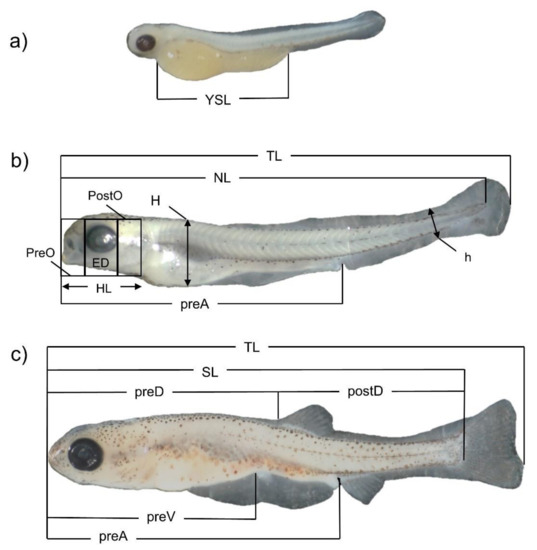
Figure 1.
Fourtheen morphometric measures taken for Delminichthys krbavensis: total length (TL), notochordal length (NL), standard length (SL), predorsal length (PreD), postdorsal length (PostD), snout length (PreO), eye diameter (ED), postorbital space (PostO), head length (HL), preventral length (PreV), pre-anal length (PreA), maximum body depth (H), depth of caudal peduncle (h), length of yolk sack (YSL). Distinct lower case letters denote different stages of development: (a) day 4 PH, (b) day 11 PH and (c) day 48 PH.
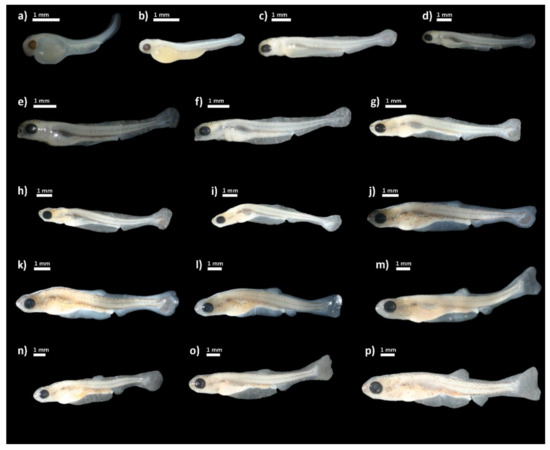
Figure 2.
Delminichthys krbavensis: (a) upon hatching; (b) day 4 post-hatching (PH); (c) day 7 PH; (d) day 9 PH; (e) day 11 PH; (f) day 14 PH; (g) day 18 PH; (h) day 21 PH; (i) day 23 PH; (j) day 25 PH; (k) day 28 PH; (l) day 32 PH; (m) day 36 PH; (n) day 39 PH; (o) day 44 PH; (p) day 48 PH.
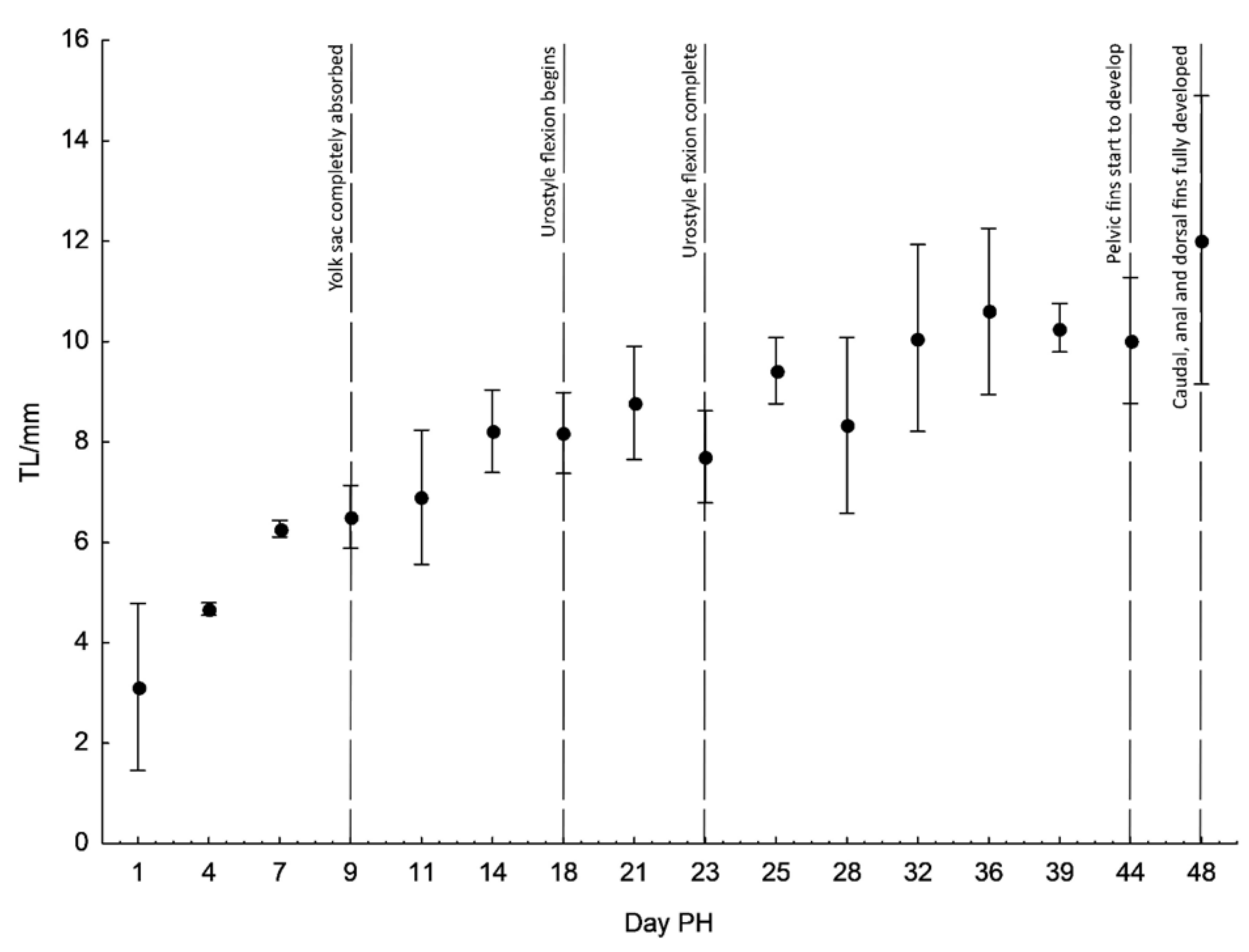
The finfold was differentiated into the caudal, ventral, dorsal and anal part. The mouth began to open but the alimentary canal was still not operational. The swim bladder appeared but was still not clearly visible. Volume of the yolk sack was drastically reduced, but its length remained almost the same. The pectoral fins were present. On day 9 PH (Figure 2d), the body was more elongated with a mean total length of 6.51 ± 0.13 mm. Pigmentation was increased on the head and ventral, lateral and dorsal surfaces. The snout became more pointed. The yolk sack was nearly completely absorbed and the swim bladder was clearly visible and filled with air. The anus was visible, but the alimentary canal was still empty with no visible organic matter. On day 11 PH (Figure 2e), the mouth was completely open and alimentary canal was filled with organic matter. The yolk sack was completely absorbed. The caudal peduncle and finfold were further differentiated. The body became more laterally compressed and the mean TL of larvae was 6.90 ± 1.08 mm. On day 14 PH (Figure 2f), the mean total length was 8.21 ± 0.66 mm with further lateral compression. Pectoral fins were very prominent. On day 18 PH, the mean total length was 8.18 ± 0.65 mm. Flexion of the urostyle began and rays in the caudal fin started to form (Figure 2g). On day 21 PH (TL = 8.78 ± 0.91 mm) (Figure 2h), rays in the caudal fin were even more visible and covered with pigmentation. The change of total length throughout the duration of the observation is shown in Figure 3.
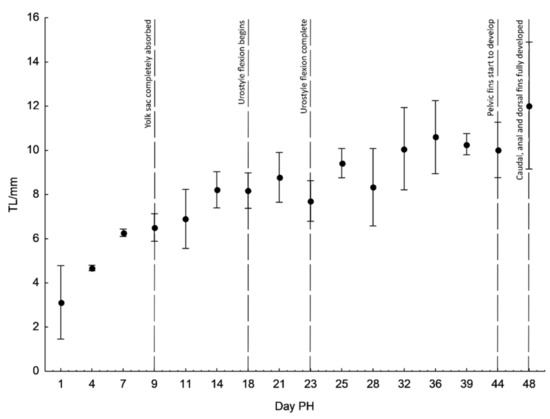
Figure 3.
Changes in total length (TL) of D. krbavensis during the early stages of development (from hatching to day 48 post-hatching (PH)) at 17 °C. Bars indicated SD (standard deviation).
On day 23 PH (Figure 2i), there were no prominent changes except the enlarged volume of the swim bladder, which was visible as a lateral bulge when viewed from above. The urostyle was completely flexed. On day 25 PH (Figure 2j), the dorsal and anal fins started to develop. The swim bladder was no longer visible due to skin thickness and coloration. On day 28 PH (Figure 2k), the subterminal mouth was formed. From days 32 to 39 PH (Figure 2l–n), all the rays on caudal, dorsal and anal fins were clearly visible and countable. The maximum observed number of fin rays in the caudal, dorsal and anal fins examined in the larvae achieved the full set as in adult specimens. The larvae continued to increase total length and the pigmentation levels. On day 44 PH (Figure 2o), the rays on pelvic fins began to develop. On day 48 PH (Figure 2p), the pelvic fins were fairly developed and all other fins were fully developed. The following phases were noted in terms of the full description of pigmentation evolution on the body: the beginning phase (from the 4th to the 11th day), the intermediate phase (14th–32nd day), and the final phase (36th–48th day). Several melanophores were found on the dorsal surface of the body in a dispersed pattern throughout the beginning phase, with the more strongly colored region located dorsally just behind the eyes and before the dorsal finfold. A pale midlateral line of slender melanophores stretched from the posterior edge of the eye to the caudal peduncle on the lateral sides of the body. Several small melanophores were also noticeable on the belly region. Ventrally, the individual melanophores were present in the area between the gill covers and from the anus to the caudal peduncle. The snout became colored at the conclusion of the first phase (dorsal and ventral lines were noticable), and tiny pigments emerged on the ventral side of the head. The dorsal (from snout to peduncle) and midlateral line of melanophores was more prominent in the intermediate phase, when the melanophores grew bigger in diameter. The presence of small but numerous melanophores had intensified the coloring of the caudal peduncle and caudal fin, whilst a higher number of wider melanophores had covered the snout and head region. The abdomen was coated with a small number of thin melanophores. Finally, two distinct, intensely colored parallel lines of melanophores were found on the dorsal side of the body, running from the eyes to the peduncle, in the final phase. Small and abundant melanophores coloured the pectoral, dorsal, anal, and caudal fins, while the dorsal and ventral sides of the skull developed a black speckled continuous pattern. The midlateral line of melanophores was now plainly visible. Numerous melanophores covered the cheek area as well.
4. Discussion
The knowledge of early life stages of fishes is critical for designing effective conservation measures since extinction risk is directly connected with the life history of the species [15] and early life stages directly influence the stability of all populations. Energy available in the environment and its allocation to different biological activities shape life histories [16]. Karstic ecosystems have distinct characteristics, and hence the animals living in karstic watersheds have distinctive life histories. Unfortunately, there is a severe lack of data on the life histories of such fish species, hindering their effective safeguarding.
This is the first description of the early development of D. krbavenis and the first report for the genus Delminichthys. Delminichthys krbavenis develops according to the embryonic development timetable proposed by [17] for other cyprinids, from larval to juvenile stages. Despite the fact that the group has a homogeneous pattern of early development, the size of free embryos and larvae, as well as the duration of yolk sac absorption, which is dependent on egg size and amount of yolk, distinguishes the diverse species in this group. According to [18], larger eggs are found in coldwater fishes, where larger eggs produce larger larvae, but the size of D. krbavensis eggs is comparable to other lithophilic cyprinid fishes with a mean egg diameter of 1.64 ± 0.08 mm, similar to species in the genera Barbus, Chondrostoma, Leuciscus, Telestes [17,19,20]. The size of the newly hatched D. krbavensis (3.12 ± 0.67 mm) is much smaller than of its former congeneric species T. karsticus (6.52 ± 0.27 mm) that spawns in colder water [21], T. pleurobipunctatus (6.00 mm; [20]) and other comparable cyprinids that spawn in warmer water, such as Leucisus aspius (Linnaeus, 1758) (7.43 mm; [22]), Leuciscus leuciscus (Linnaeus, 1758) (7.47 mm; [23]) and Leuciscus idus (Linnaeus, 1758) (7.74 mm; [24]). The newly hatched larvae are even smaller even than the phytophilic cyprinids that spawn at even higher temperature, such as Tinca tinca (Linnaeus, 1758) (3.49 mm; [25]) and Scardinius erythrophthalmus (Linnaeus, 1758) (4.7 mm; [26]).
The succession of morphological characteristics follows that of its close relative T. karsticus, with main difference in that D. krbavensis develops even slower, i.e., urostyle flexion begins on day 18 PH and finishes on day 23 PH, whereas in T. karsticus it started on day 9 PH and finished on day 23 PH; pelvic fins started to develop on day 44 PH versus on day 21 PH in T. karsticus [21]. Ref. [27] stated that the level of ontogenetic development at hatching reflects an adaptation to the environmental parameters in the natural environment. The water temperature in the spawning period of D. krbvavensis in the wild is not known, but it is unlikely to be lower than the temperature at which T. karsticus spawns (from 7.3 to 13.5 °C) [19]. The slower rate of development could be the consequence of the lower water temperature in the aquarium, though the temperature difference between D. krbavensis and T. karsticus was not significant (17 °C vs. 18 °C in T. karsticus), but this is most likely linked with the higher water levels that lingers longer in the Krbava Polje field than in the Sušik field. The flooding and high-water levels in the Krbava Polje field persist due to the limited reception capacities of the sinkholes in the field [28].
The description of the early development of D. krbavensis can be useful for both comparative studies of ontogeny in other species of this genus and for contributing to the conservation of this endangered species by providing a better understanding of this vulnerable part of its life history, allowing decision-makers to avoid any environmental disturbances from the time of spawning to the end of early development.
Author Contributions
Conceptualization, Z.M., A.J. and P.M.; methodology, Z.M., A.J., S.H. and P.M.; validation, Z.M., A.J., M.Ć, I.B., D.Z., S.H., P.M.; formal analysis, Z.M., A.J. and S.H.; investigation, Z.M., A.J. and S.H.; resources, Z.M. and P.M.; data curation, Z.M., A.J. and S.H.; writing—original draft preparation, Z.M., A.J. and S.H.; writing—review and editing, Z.M., A.J., M.Ć, I.B., D.Z., S.H., P.M.; visualization, Z.M., A.J. and S.H.; supervision, P.M.; funding acquisition, P.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ćaleta, M.; Marčić, Z.; Buj, I.; Zanella, D.; Mustafić, P.; Duplić, A.; Horvatić, S. A review of extant Croatian freshwater fish and lampreys. Annotated list and distribution. Croat. J. Fisheries 2019, 77, 136–232. [Google Scholar] [CrossRef] [Green Version]
- Ćaleta, M.; Buj, I.; Mrakovčić, M.; Mustafić, P.; Zanella, D.; Marčić, Z.; Duplić, A.; Mihinjač, T.; Katavić, I. Endemic Fishes of Croatia; Croatian Environmental Agency: Zagreb, Croatia, 2015. [Google Scholar]
- Martinić, I. (Ed.) An Overview of the State of Biological and Landscape Diversity of Croatia with the Protection Strategy and Action Plans; Ministry of Environmental Protection and Physical Planning: Zagreb, Croatia, 2000; p. 158. [Google Scholar]
- Bognar, A. Gorski Kotar i Ogulinsko-Plašćanska Udolina. U: PAVIĆ, R. (ur.) Geografija SR Hrvatske, Knjiga 4, Gorska Hrvatska; Školska Knjiga: Zagreb, Croatia, 1975. [Google Scholar]
- Vukelić, M. Laudonov gaj. Šumarski List 2001, 7/8, 425–436. [Google Scholar]
- Bioportal. HR2000632 Krbavsko Polje. Department of Nature Conservation of the Ministry of Economy and Sustainable Development. 2022. Available online: http://www.bioportal.hr (accessed on 1 March 2022).
- Zupančič, P.; Bogutskaya, N.G. Description of two new species, Phoxinellus krbavensis and P. jadovensis, re-description of P. fontinalis Karaman, 1972, and discussion of the distribution of Phoxinellus species (Teleostei: Cyprinidae) in Croatia and in Bosnia and Herzegovina. Nat. Croat. 2002, 11, 411–437. [Google Scholar]
- Kottelat, M.; Freyhof, J. Handbook of European Freshwater Fishes; Kottelat: Cornol, Switzerland; Freyhof: Berlin, Germany, 2007. [Google Scholar]
- Mihinjač, T.; Marčić, Z.; Buj, I.; Zanella, D.; Mustafić, P.; Mrakovčić, M.; Ćaleta, M. Threatened fishes of the world: Delminichthys krbavensis (Zupančič & Bogutskaya, 2002) (Cyprinidae). Croat. J. Fish. 2015, 73, 33–34. [Google Scholar] [CrossRef] [Green Version]
- Crivelli, A.J. Delminichthys krbavensis. IUCN Red List. Threat. Species 2006, 2006, eT60446A12367313. [Google Scholar] [CrossRef]
- Mrakovčić, M.; Brigić, A.; Buj, I.; Ćaleta, M.; Mustafić, P.; Zanella, D. Red Book of Freshwater Fish of Croatia; Ministry of Culture, State Institute for Nature Protection: Zagreb, Croatia, 2006; pp. 61–62. [Google Scholar]
- Mustafić, P.; Buj, I.; Opašić, M.; Zanella, D.; Marčić, Z.; Ćaleta, M.; Šanda, R.; Horvatić, S.; Mrakovčić, M. Morphological comparison of Delminichthys ghetaldii (Steindachner, 1882), D. adspersus (Heckel, 1843), D. jadovensis (Zupančič and Bogutskaya, 2002) and D. krbavensis (Zupančič & Bogutskaya, 2002), endemic species of the Dinaric karst, Croatia. J. Appl. Ichthyol. 2017, 33, 256–262. [Google Scholar] [CrossRef]
- Official Gazette of the Republic of Croatia. Ordinance on Strictly Protected Taxa (OG 144/13, 73/16). Available online: https://narodne-novine.nn.hr/clanci/sluzbeni/2013_12_144_3086.html (accessed on 16 February 2022.).
- Zupančič, P. Rare and Threatened Freshwater Fish of the Adriatic Basin of Croatia, Slovenia and Bosnia and Herzegovina; Narodna in Univerzitetna Knjižnica: Ljubljana, Slovenia, 2008. (In Slovenian) [Google Scholar]
- Hutchings, J.A. Handbook of fish biology and fisheries. In Fish Biology; Hart, P.J.B., Reynolds, J.D., Eds.; Blackwell Science: Oxford, UK, 2002; Volume 12002, pp. 149–174. [Google Scholar]
- García, V.B.; Lucifora, L.O.; Myers, R.A. The importance of habitat and life history to extinction risk in sharks, skates, rays, and chimaeras. Proc. R. Soc. B 2008, 275, 83–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peňáz, M. A general framework of fish ontogeny: A review of the ongoing debate. Folia Zool. 2001, 50, 241–256. [Google Scholar]
- Kamler, E. Parent-egg-progeny relationships in teleost fishes: An energetics perspective. Rev. Fish Biol. Fish. 2005, 15, 399–421. [Google Scholar] [CrossRef]
- Marčić, Z.; Milković, A.; Ćaleta, M.; Buj, I.; Zanella, D.; Mustafić, P. Reproductive biology of the endemic dace Telestes karsticus Marčić and Mrakovčić 2011 (Cyprinidae, Leuciscinae) endemic to Croatia. J. Appl. Ichthyol. 2017, 33, 203–208. [Google Scholar] [CrossRef]
- Barbieri, R.; Stoumboudi, M.; Kalogianni, E.; Leonardos, I. First report on the spawning migration and early life development of a cyprinid species of the genus Telestes. J. Appl. Ichthyol. 2020, 36, 817–824. [Google Scholar] [CrossRef]
- Marčić, Z.; Abramović, A.; Ćaleta, M.; Buj, I.; Zanella, D.; Horvatić, S.; Mustafić, P. Early development of the endemic dace Telestes karsticus (Leuciscidae, Cypriniformes) in a Dinaric karst stream in Croatia. J. Appl. Ichthyol. 2021, 37, 99–105. [Google Scholar] [CrossRef]
- Korzelecka-Orkisz, A.; Bonisławska, M.; Taǹski, A.; Smaruj, I.; Szulc, J.; Formicki, K. Embryonic development of Aspius aspius L. (Actinopterygii: Cypriniformes: Cyprinidae). Electron. J. Pol. Agric. Univ. 2013, 16, #09. Available online: http://www.ejpau.media.pl/volume16/issue3/art-09.html (accessed on 15 March 2022).
- Kupren, K.; Rams, I.; Żarski, D.; Kucharczyk, D. Early development and allometric growth patterns of rheophilic cyprinid common dace Leuciscus leuciscus (Cyprinidae: Leuciscinae). Ichthyol. Res. 2016, 63, 382–390. [Google Scholar] [CrossRef] [Green Version]
- Kupren, K.; Żarski, D.; Kucharczyk, D. Early development and allometric growth patterns in ide Leuciscus idus (Linnaeus 1758). J. Appl. Ichthyol. 2015, 31, 509–517. [Google Scholar] [CrossRef]
- Peňáz, M.; Prokeš, M.; Kouřil, J.; Hamačkowa, J. Early development of the carp, Cyprinus carpio. Acta Sci. Nat. Acad. Sci. Bohem. Brno 1983, 17, 1–39. [Google Scholar]
- Korzelecka-Orkisz, A.; Bonisławska, M.; Pawlos, D.; Szulc, J.; Winnicki, A.; Formicki, K. Morphophysiological aspects of the embryonic development of tench (Tinca tinca L.). Electron. J. Pol. Agric. Univ. 2009, 12, #21. Available online: http://www.ejpau.media.pl/volume12/issue4/art-21.html (accessed on 15 March 2022).
- Korwin-Kossakowski, M. The influence of temperature during the embryonic period on larval growth and development in carp, Cyprinus carpio L.; and grass carp, Ctenopharyngodon idella (Val.): Theoretical and practical aspects. Arch. Pol. Fish. 2008, 16, 231–314. [Google Scholar] [CrossRef]
- Biondić, R.; Rubinić, J.; Biondić, B.; Meaški, H.; Radišić, M. Defining of Trends and Assessment of Groundwater Status in the Karst Area in Croatia; Technical Report; Faculty of Geotechnical Engineering and Rijeka, University of Zagreb: Varaždin, Croatia; Faculty of Civil Engineering, University of Rijeka: Varaždin, Croatia, 2016. (In Croatian) [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).