
Wild Apples Are Not That Wild: Conservation Status and Potential Threats of Malus sieversii in the Mountains of Central Asia Biodiversity Hotspot

 ,
, 
 and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Area
2.2. Data Source
2.2.1. Species Occurrence Records for Wild Apple
2.2.2. Current and Future Climate Data
2.2.3. Current and Future Land-Use Data
2.2.4. Topographic and Edaphic Variables
2.3. Species Distribution Models
2.4. Data Analysis
3. Results
3.1. Range Shifts of Wild Apple
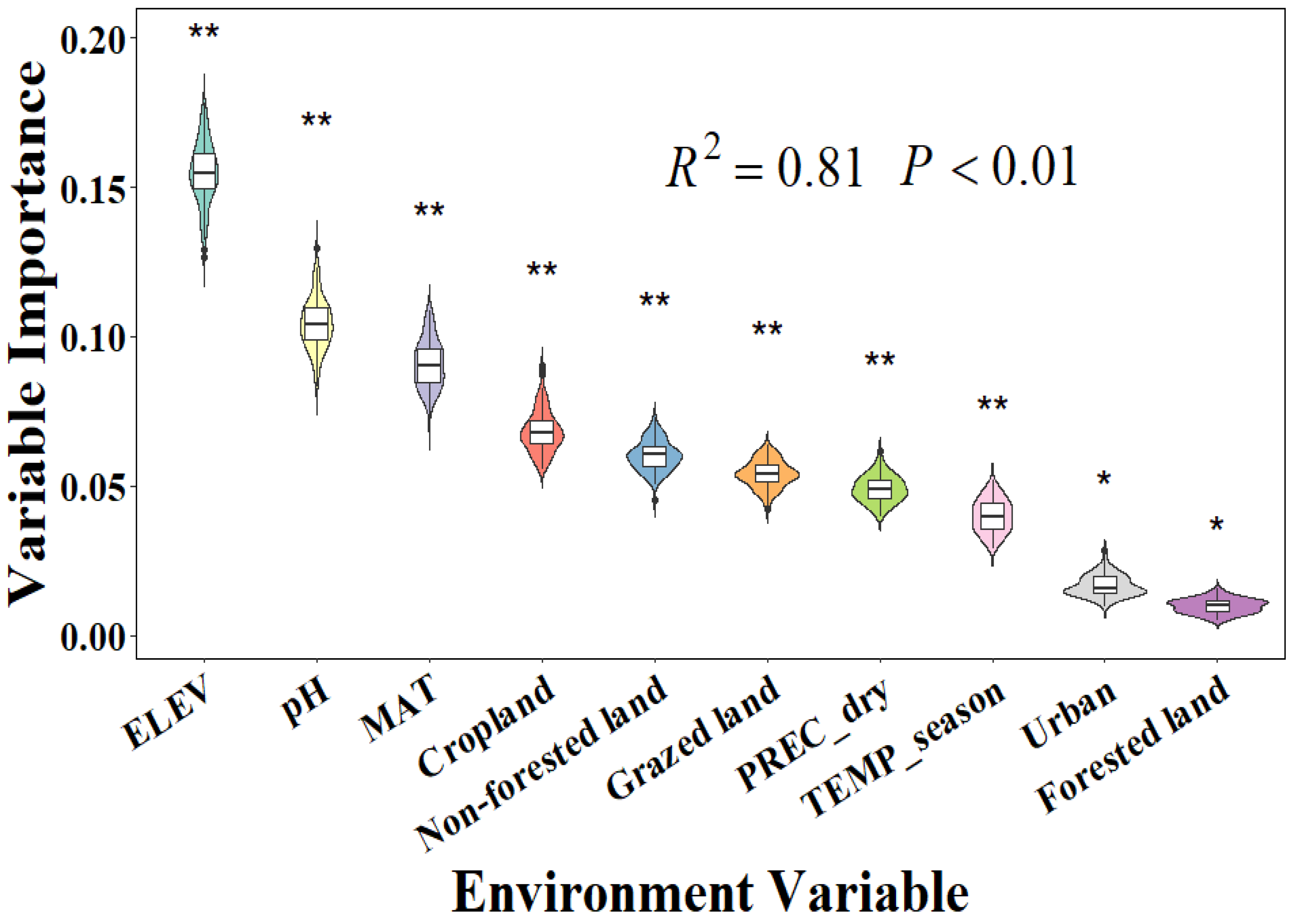
3.2. The Importance of Environmental Factors
3.3. The Effectiveness of Current Protected Areas
4. Discussion
4.1. Dramatic Range Shifts of Wild Apple
4.2. The Influence of Climate Change on Wild Apple and Wild Fruit Forests
4.3. The Influence of Land-Use Changes on Wild Apple and Wild Fruit Forests
4.4. Implications for the Protection of Wild Apple and Wild Fruit Forests
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maxted, N.; Ford-Lloyd, B.V.; Jury, S.; Kell, S.; Scholten, M. Towards a definition of a crop wild relative. Biodivers. Conserv. 2006, 15, 2673–2685. [Google Scholar] [CrossRef]
- Myrans, H.; Diaz, M.V.; Khoury, C.K.; Carver, D.; Henry, R.J.; Gleadow, R. Modelled distributions and conservation priorities of wild sorghums (Sorghum Moench). Divers. Distrib. 2020, 26, 1727–1740. [Google Scholar] [CrossRef]
- Castañeda-Álvarez, N.P.; Khoury, C.K.; Achicanoy, H.A.; Bernau, V.; Dempewolf, H.; Eastwood, R.J.; Guarino, L.; Harker, R.H.; Jarvis, A.; Maxted, N.; et al. Global conservation priorities for crop wild relatives. Nat. Plants 2016, 2, 16022. [Google Scholar] [CrossRef]
- van Treuren, R.; Hoekstra, R.; van Hintum, T.J. Inventory and prioritization for the conservation of crop wild relatives in The Netherlands under climate change. Biol. Conserv. 2017, 216, 123–139. [Google Scholar] [CrossRef]
- Vincent, H.; Amri, A.; Castañeda-Álvarez, N.P.; Dempewolf, H.; Dulloo, E.; Guarino, L.; Hole, D.; Mba, C.; Toledo, A.; Maxted, N. Modeling of crop wild relative species identifies areas globally for in situ conservation. Commun. Biol. 2019, 2, 136. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Geng, X.; Hao, Z.; Zheng, J. Changes in Climate Extremes in Central Asia under 1.5 and 2 °C Global Warming and their Impacts on Agricultural Productions. Atmosphere 2020, 11, 1076. [Google Scholar] [CrossRef]
- Kell, S.; Qin, H.; Chen, B.; Ford-Lloyd, B.; Wei, W.; Kang, D.; Maxted, N. China’s crop wild relatives: Diversity for agriculture and food security. Agric. Ecosyst. Environ. 2015, 209, 138–154. [Google Scholar] [CrossRef]
- Aguirre-Gutiérrez, J.; van Treuren, R.; Hoekstra, R.; van Hintum, T.J. Crop wild relatives range shifts and conservation in Europe under climate change. Divers. Distrib. 2017, 23, 739–750. [Google Scholar] [CrossRef]
- Greene, S.L.; Williams, K.A.; Khoury, C.K.; Kantar, M.B.; Marek, L.F. (Eds.) North American Crop Wild Relatives, Volume 1: Conservation Strategies; Springer Nature: Cham, Switzerland, 2018. [Google Scholar]
- Lapeña, I.; Turdieva, M.; López Noriega, I.; Ayad, W.G. Conservation of Fruit Tree Fiversity in Central Asia: Policy Options and Challenges; Bioversity International: Rome, Italy, 2014. [Google Scholar]
- Newbold, T.; Hudson, L.N.; Hill, S.L.L.; Contu, S.; Lysenko, I.; Senior, R.A.; Börger, L.; Bennett, D.J.; Choimes, A.; Collen, B.; et al. Global effects of land use on local terrestrial biodiversity. Nature 2015, 520, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Maxted, N.; Vincent, H. Review of congruence between global crop wild relative hotspots and centres of crop origin/diversity. Genet. Resour. Crop Evol. 2021, 68, 1283–1297. [Google Scholar] [CrossRef]
- Labokas, J.; Maxted, N.; Kell, S.; Brehm, J.M.; Iriondo, J.M. Development of national crop wild relative conservation strategies in European countries. Genet. Resour. Crop Evol. 2018, 65, 1385–1403. [Google Scholar] [CrossRef]
- FAO. The State of the World’s Biodiversity for Food and Agriculture; Food and Agriculture Organization of the United Nation: Rome, Italy, 2019. [Google Scholar]
- Rodríguez, A.; Pérez-López, D.; Centeno, A.; Ruiz-Ramos, M. Viability of temperate fruit tree varieties in Spain under climate change according to chilling accumulation. Agric. Syst. 2021, 186, 102961. [Google Scholar] [CrossRef]
- Sthapit, B.R.; Ramanatha Rao, V.; Sthapit, S. Tropical Fruit Tree Species and Climate Change; Bioversity International: New Delhi, India, 2012. [Google Scholar]
- Paull, R.E.; Duarte, O. Tropical Fruits, 2nd ed.; CAB International: London, UK, 2011. [Google Scholar]
- Maxted, N.; Kell, S. Establishment of a Global Network for the In Situ Conservation of Crop Wild Relatives Status and Needs; Commission on Genetic Resources for Food and Agriculture, Food and Agriculture Organization of the United Nations: Rome, Italy, 2009. [Google Scholar]
- Conservation International. Biodiversity Hotspots. 2016. Available online: https://www.conservation.org (accessed on 1 March 2022).
- Sitpayeva, G.T.; Kudabayeva, G.M.; Dimeyeva, L.A.; Gemejiyeva, N.G.; Vesselova, P.V. Crop wild relatives of Kazakhstani Tien Shan: Flora, vegetation, resources. Plant Divers. 2020, 42, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Dzhangaliev, A.D.; Salova, T.N.; Turekhanova, P.M. The Wild Fruit and Nut Plants of Kazakhstan. Hortic. Rev. 2003, 29, 305–371. [Google Scholar] [CrossRef]
- Panyushkina, I.; Mukhamadiev, N.; Lynch, A.; Ashikbaev, N.; Arizpe, A.; O’Connor, C.; Abjanbaev, D.; Mengdibayeva, G.; Sagitov, A. Wild Apple Growth and Climate Change in Southeast Kazakhstan. Forests 2017, 8, 406. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.; Zhang, C.; Hu, Q.; Tian, H. Temperature Changes in Central Asia from 1979 to 2011 Based on Multiple Datasets. J. Clim. 2014, 27, 1143–1167. [Google Scholar] [CrossRef]
- Hu, Y.; Hu, Y. Land Cover Changes and Their Driving Mechanisms in Central Asia from 2001 to 2017 Supported by Google Earth Engine. Remote Sens. 2019, 11, 554. [Google Scholar] [CrossRef] [Green Version]
- Eastwood, A.; Lazkov, G.; Newton, A. The Red List of Trees of Central Asia; Fauna & Flora International: Cambridge, UK, 2009. [Google Scholar]
- Hamidov, A.; Helming, K.; Balla, D. Impact of agricultural land use in Central Asia: A review. Agron. Sustain. Dev. 2016, 36, 6. [Google Scholar] [CrossRef] [Green Version]
- Velasco, R.; Zharkikh, A.; Affourtit, J.; Dhingra, A.; Cestaro, A.; Kalyanaraman, A.; Fontana, P.; Bhatnagar, S.K.; Troggio, M.; Pruss, D.; et al. The genome of the domesticated apple (Malus × domestica Borkh.). Nat. Genet. 2010, 42, 833–839. [Google Scholar] [CrossRef]
- Zhang, X.S. On the eco-geographical characters and the problems of classification of the wild fruit-tree forest in the Ili Valley of Sinkiang. Chin. Acta Bot. Sinica 1973, 15, 239–253. [Google Scholar]
- Dzhangaliev, A. The Wild Apple Tree of Kazakhstan. Hortic. Rev. 2003, 29, 63–304. [Google Scholar]
- Ding, Y.; Zang, R.G.; Zhang, Z. Population Structure and Spatial Pattern of Forest Plant in Northern Xinjiang. In Ecological Characteristics of Forest Vegetation in Northern Xinjiang; Zang, R.G., Jin, X.H., Liu, H., Eds.; Modern Education Press: Beijing, China, 2011; pp. 156–170. [Google Scholar]
- IUCN. Malus Sieversii. The IUCN Red List of Threatened Species 2007: E.T32363A9693009. 2007. Available online: https://doi.org/10.2305/IUCN.UK.2007.RLTS.T32363A9693009.en (accessed on 1 March 2022).
- Bothe, O.; Fraedrich, K.; Zhu, X. Precipitation climate of Central Asia and the large-scale atmospheric circulation. Theor. Appl. Climatol. 2012, 108, 345–354. [Google Scholar] [CrossRef]
- de Beurs, K.M.; Henebry, G.M. Land surface phenology, climatic variation, and institutional change: Analyzing agricultural land cover change in Kazakhstan. Remote Sens. Environ. 2004, 89, 497–509. [Google Scholar] [CrossRef]
- GBIF.org. GBIF Occurrence Download. 2021. Available online: https://www.gbif.org/dl.x6ngcf (accessed on 6 October 2021).
- CVH. Chinese Virtual Herbarium. 2020. Available online: https://www.cvh.ac.cn (accessed on 10 October 2021).
- Aiello-Lammens, M.E.; Boria, R.A.; Radosavljevic, A.; Vilela, B.; Anderson, R.P. spThin: An R package for spatial thinning of species occurrence records for use in ecological niche models. Ecography 2015, 38, 541–545. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Eyring, V.; Bony, S.; Meehl, G.A.; Senior, C.A.; Stevens, B.; Stouffer, R.J.; Taylor, K.E. Overview of the Coupled Model Intercomparison Project Phase 6 (CMIP6) experimental design and organization. Geosci. Model Dev. 2016, 9, 1937–1958. [Google Scholar] [CrossRef] [Green Version]
- Hurtt, G.C.; Chini, L.; Sahajpal, R.; Frolking, S.; Bodirsky, B.L.; Calvin, K.; Doelman, J.C.; Fisk, J.; Fujimori, S.; Goldewijk, K.K.; et al. Harmonization of Global Land-Use Change and Management for the Period 850–2100 (LUH2) for CMIP6. Geosci. Model Dev. Discuss. 2020, 13, 5425–5464. [Google Scholar] [CrossRef]
- Song, H.; Ordonez, A.; Svenning, J.-C.; Qian, H.; Yin, X.; Mao, L.; Deng, T.; Zhang, J. Regional disparity in extinction risk: Comparison of disjunct plant genera between eastern Asia and eastern North America. Glob. Chang. Biol. 2021, 27, 1904–1914. [Google Scholar] [CrossRef]
- Hoskins, A.J.; Bush, A.; Gilmore, J.; Harwood, T.; Hudson, L.N.; Ware, C.; Williams, K.J.; Ferrier, S. Downscaling land-use data to provide global 30″ estimates of five land-use classes. Ecol. Evol. 2016, 6, 3040–3055. [Google Scholar] [CrossRef] [Green Version]
- Hengl, T.; Mendes de Jesus, J.; Heuvelink, G.B.M.; Ruiperez Gonzalez, M.; Kilibarda, M.; Blagotić, A.; Shangguan, W.; Wright, M.N.; Geng, X.; Bauer-Marschallinger, B.; et al. SoilGrids250m: Global gridded soil information based on machine learning. PLoS ONE 2017, 12, e0169748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araújo, M.B.; New, M. Ensemble forecasting of species distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Valavi, R.; Guillera-Arroita, G.; Lahoz-Monfort, J.J.; Elith, J. Predictive performance of presence-only species distribution models: A benchmark study with reproducible code. Ecol. Monogr. 2022, 92, e01486. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kammon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Hao, T.; Elith, J.; Lahoz-Monfort, J.J.; Guillera-Arroita, G. Testing whether ensemble modelling is advantageous for maximising predictive performance of species distribution models. Ecography 2020, 43, 549–558. [Google Scholar] [CrossRef] [Green Version]
- Roberts, D.R.; Bahn, V.; Ciuti, S.; Boyce, M.S.; Elith, J.; Guillera-Arroita, G.; Hauenstein, S.; Lahoz-Monfort, J.J.; Schröder, B.; Thuiller, W.; et al. Cross-validation strategies for data with temporal, spatial, hierarchical, or phylogenetic structure. Ecography 2017, 40, 913–929. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD-a platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Gallien, L.; Münkemüller, T.; Albert, C.H.; Boulangeat, I.; Thuiller, W. Predicting potential distributions of invasive species: Where to go from here? Divers. Distrib. 2010, 16, 331–342. [Google Scholar] [CrossRef]
- Choe, H.; Thorne, J.H.; Hijmans, R.; Kim, J.; Kwon, H.; Seo, C. Meta-corridor solutions for climate-vulnerable plant species groups in South Korea. J. Appl. Ecol. 2017, 54, 1742–1754. [Google Scholar] [CrossRef] [Green Version]
- Jiao, S.; Chen, W.; Wang, J.; Du, N.; Li, Q.; Wei, G. Soil microbiomes with distinct assemblies through vertical soil profiles drive the cycling of multiple nutrients in reforested ecosystems. Microbiome 2018, 6, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warren, D.L.; Matzke, N.J.; Cardillo, M.; Baumgartner, J.B.; Beaumont, L.J.; Turelli, M.; Glor, R.E.; Huron, N.A.; Simões, M.; Iglesias, T.L.; et al. ENMTools 1.0: An R package for comparative ecological biogeography. Ecography 2021, 44, 504–511. [Google Scholar] [CrossRef]
- UNEP-WCMC. Protected Areas Map of the World. 2021. Available online: http://www.protectedplanet.net (accessed on 1 March 2022).
- R Core Team. R: A Language and Environment for Statistical Computing, v4.2.0. R Foundation for Statistical Computing: Vienna, Austria, 2022. Available online: https://www.R-project.org (accessed on 22 April 2022).
- Zhang, H.X.; Li, X.; Wang, J.; Zhang, D. Insights into the aridification history of Central Asian Mountains and international conservation strategy from the endangered wild apple tree. J. Biogeogr. 2021, 48, 332–344. [Google Scholar] [CrossRef]
- Shan, Q.; Wang, Z.; Ling, H.; Zhang, G.; Yan, J.; Han, F. Unreasonable human disturbance shifts the positive effect of climate change on tree-ring growth of Malus sieversii in the origin area of world cultivated apples. J. Clean. Prod. 2021, 287, 125008. [Google Scholar] [CrossRef]
- Lehmann, P.; Ammunét, T.; Barton, M.; Battisti, A.; Eigenbrode, S.D.; Jepsen, J.U.; Kalinkat, G.; Neuvonen, S.; Niemelä, P.; Terblanche, J.S.; et al. Complex responses of global insect pests to climate warming. Front. Ecol. Environ. 2020, 18, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Jiang, T.; Wang, Y.; Su, B.; Tao, H.; Qin, J.; Zhai, J. Spatiotemporal variations of aridity index over the Belt and Road region under the 1.5 °C and 2.0 °C warming scenarios. J. Geogr. Sci. 2020, 30, 37–52. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Jiang, D.; Lang, X. Future changes in Aridity Index at two and four degrees of global warming above preindustrial levels. Int. J. Climatol. 2021, 41, 278–294. [Google Scholar] [CrossRef]
- Li, J.; Chen, H.; Zhang, C.; Pan, T. Variations in ecosystem service value in response to land use/land cover changes in Central Asia from 1995–2035. PeerJ 2019, 7, e7665. [Google Scholar] [CrossRef]
- Zhang, C.; Ren, W. Complex climatic and CO2 controls on net primary productivity of temperate dryland ecosystems over central Asia during 1980–2014. J. Geophy. Res.-Biogeo. 2017, 122, 2356–2374. [Google Scholar] [CrossRef]
- Luedeling, E.; Zhang, M.; Girvetz, E.H. Climatic changes lead to declining winter chill for fruit and nut trees in California during 1950–2099. PLoS ONE 2009, 4, e6166. [Google Scholar] [CrossRef]
- Salama, A.-M.; Ezzat, A.; El-Ramady, H.; Alam-Eldein, S.M.; Okba, S.K.; Elmenofy, H.M.; Hassan, I.F.; Illés, A.; Holb, I.J. Temperate Fruit Trees under Climate Change: Challenges for Dormancy and Chilling Requirements in Warm Winter Regions. Horticulturae 2021, 7, 86. [Google Scholar] [CrossRef]
- Ju, H.; van der Velde, M.; Lin, E.; Xiong, W.; Li, Y. The impacts of climate change on agricultural production systems in China. Clim. Chang. 2013, 120, 313–324. [Google Scholar] [CrossRef]
- Feeley, K.J.; Silman, M.R. Land-use and climate change effects on population size and extinction risk of Andean plants. Glob. Chang. Biol. 2010, 16, 3215–3222. [Google Scholar] [CrossRef]
- Titeux, N.; Henle, K.; Mihoub, J.-B.; Regos, A.; Geijzendorffer, I.R.; Cramer, W.; Verburg, P.H.; Brotons, L. Global scenarios for biodiversity need to better integrate climate and land use change. Divers. Distrib. 2017, 23, 1231–1234. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Likens, G.E. Adaptive monitoring: A new paradigm for long-term research and monitoring. Trends Ecol. Evol. 2009, 24, 482–486. [Google Scholar] [CrossRef]
- Nichols, J.D.; Williams, B.K. Monitoring for conservation. Trends Ecol. Evol. 2006, 21, 668–673. [Google Scholar] [CrossRef]
- Wolf, C.; Levi, T.; Ripple, W.J.; Zárrate-Charry, D.A.; Betts, M.G. A forest loss report card for the world’s protected areas. Nat. Ecol. Evol. 2021, 5, 520–529. [Google Scholar] [CrossRef]
- Beale, C.M.; Baker, N.E.; Brewer, M.J.; Lennon, J.J. Protected area networks and savannah bird biodiversity in the face of climate change and land degradation. Ecol. Lett. 2013, 16, 1061–1068. [Google Scholar] [CrossRef]
- Wessely, J.; Hülber, K.; Gattringer, A.; Kuttner, M.; Moser, D.; Rabitsch, W.; Schindler, S.; Dullinger, S.; Essl, F. Habitat-based conservation strategies cannot compensate for climate-change-induced range loss. Nat. Clim. Chang. 2017, 7, 823–827. [Google Scholar] [CrossRef]
- Alagador, D.; Cerdeira, J.O.; Araújo, M.B. Shifting protected areas: Scheduling spatial priorities under climate change. J. Appl. Ecol. 2014, 51, 703–713. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | Region | Range Size (km2) and Proportion (%) | ||||
|---|---|---|---|---|---|---|
| Current | 2050s (SSP126) | 2050s (SSP585) | 2090s (SSP126) | 2090s (SSP585) | ||
| China | Xinjiang | 693 (4.93%) | 966 (5.76%) | 903(7.19%) | 966(5.74%) | 609(7.29%) |
| Kazakhstan | East Kazakhstan | 588 (19.05%) | 2961 (19%) | 4536(21.82%) | 2730(18.08%) | 7497(26.27%) |
| Almaty | 3171 (36.39%) | 3759 (48.77%) | 3591(52.13%) | 3780(50.42%) | 3087(49.49%) | |
| Zhambyl | 21 (0.74%) | 0 | 0 | 0 | 0 | |
| South Kazakhstan | 903 (27.04%) | 504 (33.33%) | 315(46.88%) | 483(31.08%) | 294(36.84%) | |
| Kyrgyzstan | / | 483 (3.55%) | 294 (4.86%) | 168(4.37%) | 294(4.88%) | 105(8.47%) |
| Tajikistan | / | 84 (7.27%) | 63 (17.65%) | 0 | 42(15.38%) | 0 |
| Uzbekistan | / | 42 (25%) | 21 (100%) | 21(100%) | 0 | 0 |
| Total | 5985 (12.75%) | 9534 (17.27%) | 9534(20.72%) | 8295(16.96%) | 11,592 (25.1%) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, Z.; Song, H.; Wang, Y.; Li, J.; Maimaiti, M.; Liu, Z.; Zhang, H.; Zhang, J. Wild Apples Are Not That Wild: Conservation Status and Potential Threats of Malus sieversii in the Mountains of Central Asia Biodiversity Hotspot. Diversity 2022, 14, 489. https://doi.org/10.3390/d14060489
Tian Z, Song H, Wang Y, Li J, Maimaiti M, Liu Z, Zhang H, Zhang J. Wild Apples Are Not That Wild: Conservation Status and Potential Threats of Malus sieversii in the Mountains of Central Asia Biodiversity Hotspot. Diversity. 2022; 14(6):489. https://doi.org/10.3390/d14060489
Chicago/Turabian StyleTian, Zhongping, Houjuan Song, Yuzhuo Wang, Jin Li, Mierkamili Maimaiti, Zhongquan Liu, Hongxiang Zhang, and Jian Zhang. 2022. "Wild Apples Are Not That Wild: Conservation Status and Potential Threats of Malus sieversii in the Mountains of Central Asia Biodiversity Hotspot" Diversity 14, no. 6: 489. https://doi.org/10.3390/d14060489