Abstract
Available research on invertebrates in Corallina officinalis settlements shows a high level of biodiversity due to a complex habitat structure. Our aim was to examine seasonal changes in the invertebrate population, considering the algae’s growth patterns. Nine locations with over 90% algal coverage were selected in southern Istria, where quantitative sampling was performed using six replicates of 5 × 5 cm quadrats in each location. Results showed that 29,711 invertebrates were found during winter (maximum algae growth) and 22,292 during summer (minimum algae growth), with an extrapolated average density of 220,000 and 165,200 individuals per square meter, respectively. The total number of individuals showed a linear increase as the algae biomass increased. The highest density, 586,000 individuals, was recorded in the Premantura area during winter. Dominant groups such as amphipods, polychaetes, bivalves and gastropods made up over 80% of the invertebrates. Our study confirms high invertebrate richness in the C. officinalis settlements, with the maximum density being the highest when compared to previously published data.
1. Introduction
The algae Corallina officinalis Linnaeus is the most commonly found species of its genus in temperate regions worldwide, forming dense settlements in the intertidal zone [1]. Previous mapping of macroalgae communities in the eastern Adriatic area using the CARLIT method showed that communities dominated by species of the genus Corallina are located in 13% of the investigated coast [2]. The unique structure of this alga modifies its surroundings, serving as an example of how algae can shape the composition and relationships of associated organisms. Its structure provides shelter for both vagile and sessile organisms during adverse weather [3,4,5,6,7,8]. Many benthic invertebrates found in algal forests have a larval stage that floats or swims in the plankton, making the macroalgae significant as larval collectors [9]. Macroalgae are crucial in supporting marine invertebrate communities, attracting diverse organisms [10,11,12,13,14]. These inhabitants form intricate associations, relying on macroalgae for various purposes, creating behavioral patterns that keep them within the algae [8]. Algal epibionts adapt to abiotic factors and exhibit varying degrees of dependence on algae resources [15,16,17].
According to previous research, several main groups of invertebrates have been recognized as the most abundant within C. officinalis settlements. These are mollusks (classes Gastropoda and Bivalvia), polychaetes and higher crustaceans (orders Amphipoda, Isopoda and Tanaidacea) [18,19,20,21,22,23,24,25,26]. Gastropods that usually live associated with macroalgae, whether they are species from the genus Corallina or some other macroalgae (such as fucalean species), are mainly herbivores and feed on the algae itself or on diatoms and epiphytic algae. Other species are detritivores, which feed on sediment trapped inside the algal thallus and use organic matter and microorganisms from that sediment [27,28]. The bivalves that live within C. officinalis settlements are diverse and can survive at various depths, temperatures and salinity [29]. In addition, there are also species that can survive in conditions in which the quality of sea water is lower [30]. Polychaetes are mainly marine invertebrates that inhabit a wide range of habitats, from the intertidal zone down to deep-sea sediments. They are one of the most diverse groups of invertebrates that constitute a significant percentage of the total number of organisms living associated with C. officinalis [24]. Peracarids are the most dominant crustaceans found within coralline algae [31]. Their feeding habits have been studied, and it has been found that, while detritus is one of their primary food sources, carnivorous species often feed on other types of crustaceans, and herbivores feed on macroalgae tissue [32]. This explains why they are a frequent group found in C. officinalis habitats. Other organisms that can be found in higher abundances in this type of habitat include pycnogonids, mites and nematodes [24,25,26,33,34,35].
As C. officinalis is a turf-forming algae, the effect of its biomass on the richness of invertebrate species has proven to be an important factor. Previous research has shown a positive correlation between the number of invertebrate species and algae biomass [36]. In the same research, another positive effect was recorded, namely the fact that turf-forming alga retain a larger amount of sediment that serves as a secondary habitat for many macrofauna, meiofauna and microfauna species [37]. Based on this observation, our research aimed to investigate how seasonal variations in C. officinalis algae, specifically its minimum and maximum growth phases, influence the dynamics of invertebrate structure and composition within its habitat. We hypothesized that changes in algal biomass during its maximum and minimum growth phases would result in differences in the number of individuals within the algae settlements.
2. Materials and Methods
2.1. Research Area and Sampling Methods
The research was conducted in the coastal area of southern Istria and the Brijuni National Park (Croatia, Adriatic Sea). Based on preliminary research by Buršić et al., 2019 [24] and essential coastal features (percentage of Corallina officinalis cover, slope of the coast and wind exposure), four areas were selected for seasonal sampling. In three of these areas (Pula, Banjole and Premantura), two locations were selected, while in NP Brijuni there were three locations (Figure 1). Within each location, two sites were chosen (minimum distance of 100 m between them) from which three subsamples were collected. Sampling was done within a 5 × 5 cm quadrat, as this was determined, after our preliminary research [24], to be the most appropriate size to provide a more accurate description of the research area, and also considering the number of invertebrates collected and further laboratory analyses to be conducted.

Figure 1.
Corallina officinalis sampling locations (black dots) within four investigated areas in south Istria, namely Pula, Banjole, Premantura and NP Brijuni—Brijuni National Park; arrows indicate initial and final mapping points (a). C. officinalis habitat in Javorika Bay in NP Brijuni area (b).
A total of 108 quadrats were sampled in the four areas during two seasons, 54 quadrats during the algae’s maximum growth (from November 2017 to April 2018, the “winter” samples) and 54 quadrats during the algae’s minimum growth (from June to August 2018, the “summer” samples). According to Croatian Meteorological and Hydrological Service data (https://meteo.hr/index_en.php, accessed on 12 October 2023), winters in Croatia’s northern Adriatic have average temperatures of 5 °C to 10 °C, occasionally dropping below freezing, with frequent rainfall ranging from 20 mm to 130 mm. Snowfall is rare and usually light, not leading to lasting snow cover. In contrast, summers in Istria are warm to hot, with temperatures averaging from 22 °C to 25 °C and the possibility of heatwaves. Summers are typically dry, with low monthly rainfall between 20 mm and 60 mm, occasionally interrupted by thunderstorms. Quadrats were sampled in areas where algae coverage was evenly distributed and exceeded 90% along the entire coastline. The coverage was evaluated in a coastal belt with a width of approximately 50 cm. Samples were taken during low tide when C. officinalis was completely out of the water to minimize the loss of associated organisms. Collection was performed with a hammer and chisel in order to gather the entire algal thallus. Immediately after collection, samples were stored in plastic bottles and fixed with alcohol. The next day, samples were washed in the laboratory through a sieve with a mesh size of 0.5 mm and stored in alcohol for further processing.
Invertebrates were separated manually from algae samples under a stereomicroscope with 10× to 60× magnification. Each algal thallus was examined in detail to isolate all the invertebrates present. After the separation of all invertebrates, the algae were dried at 80 °C for 24 h and weighed. By using dry weight, the water component is removed, leading to a more consistent and comparable measurement. Organisms were determined to the lowest accessible taxonomic group according to various resources available in the literature [38,39,40,41,42,43,44,45,46].
2.2. Analysis of the Invertebrate Structure and Composition
The total number of individuals (N) per investigated area was recorded and expressed per unit area of 150 cm2 (two sets of three replicate quadrats 5 × 5 cm in size). In addition, relative abundance and frequency of occurrence were calculated. These values were calculated separately for all taxonomic groups depending on sampling season, i.e., for 54 sampled quadrats during the algae’s maximum growth (“winter”) and for 54 sampled quadrates during the algae’s minimum growth (“summer”). Based on the relative abundance and frequency of occurrence, the taxonomic groups found can be classified into several different categories. To assess the dominance structure, the relative abundance was employed using the formula A = (ni/N) × 100, where ni represents the count of individual representatives for each taxonomic group and N indicates the total count of all individuals. The classification system introduced by Tischler, 1949 [47] for invertebrates and used by Travizi, 2010 [48] for marine invertebrates was adopted, comprising five categories as follows: eudominant (>10% of all individuals); dominant (5 to 10%); subdominant (2 to 5%); recedent (1 to 2%) and subrecedent (<1%). Frequency of occurrence was calculated using the formula Fa = Na/N × 100, where Na represents the number of samples in which taxonomic group a occurs and N represents the total number of samples. Taxonomic groups were classified into 4 categories: very frequent—present in 75–100% of samples, frequent—present in 50–75% of samples, widespread—present in 25–50% of samples and rare—present in 0–25% of samples [48,49].
The linear dependence of the total number of individuals of higher taxonomic groups against the dry weight of C. officinalis was verified by linear regression analysis in Microsoft Office Excel 2016. The linear regression analysis was used to analyze the relationship between two variables: the number of individuals and the dry weight (biomass) of C. officinalis. The seasonal variation in invertebrates within the Corallina officinalis turf was analyzed using non-metric multidimensional scaling (nMDS) based on square root transformation and Bray–Curtis similarity.
3. Results
The southern Istrian coast was mapped from Pula in the north-west to Ližnjan in the south-east. A total of 29,711 individuals of invertebrates were isolated in the winter samples of Corallina officinalis, and 22,292 individuals in the summer samples. In the non-metric multidimensional scaling (nMDS) analysis, clear seasonal variations were observed between periods of minimum and maximum algae growth within the invertebrate taxonomic groups associated with C. officinalis settlements (Figure 2). The average number of individuals per sampled quadrat was 550 for winter samples and 413 for summer samples, which, calculated per square meter, amounts to 220,000 individuals (ind m−2) for “winter” and 165,200 ind m−2 for “summer”. As in the preliminary sampling [24], the density of individuals associated with C. officinalis settlements varied between different sampling locations. Thus, locations Kamenjak and Stupice in Premantura had the highest recorded abundance of invertebrates. The maximum recorded number of individuals within a 5 × 5 cm quadrat was 1465 (amounting to 586,000 ind m−2) and was sampled during the period of maximum algae growth at the Stupice location (Figure 3).

Figure 2.
Non-metric MDS ordination based on square root transformation and Bray-Curtis similarity of the total number of individuals during both sampling seasons in nine locations (“winter”—samples taken during the algae’s maximum growth; “summer”—samples taken during the algae’s minimum growth).
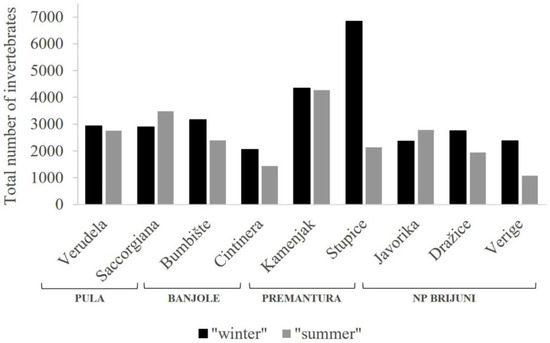
Figure 3.
Seasonal variations in the total number of invertebrates, expressed per sampling location described by six replicates, i.e., area of 150 cm2 (“winter” and “summer” having the same definitions as in Figure 2).
The recorded dry weight of algae from 108 samples was compared for both seasons (Figure 4). During the period of maximum growth, the average dry weight per sampling quadrat was 9.18 g, while during the minimum growth, it was 7.35 g.
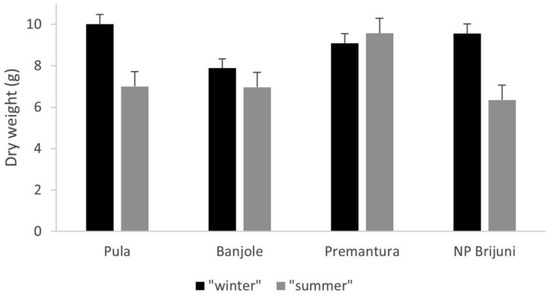
Figure 4.
Comparison of C. officinalis average dry weight (g/25 cm2) during both sampling seasons (“winter” and “summer” having the same definitions as in Figure 2).
Isolated invertebrates were classified within 20 different taxonomic groups, whose relative abundance and frequency of occurrence varied depending on the sampling season. During the algae’s maximum growth, relative abundance ranged from 0.01% to 31.72%, and frequency of occurrence from 7.41% to 100%, while during algae’s minimum growth, relative abundance ranged from 0.01% to 55.24%, and frequency of occurrence from 1.85% to 100% (Table 1).

Table 1.
Relative abundance (A) and frequency of occurrence (F) of the taxonomic groups recorded in this investigation (“winter”—samples taken during the algae’s maximum growth; “summer”—samples taken during the algae’s minimum growth).
The calculation for the relative abundance of taxonomic groups during both sampling seasons showed that the category of subrecedent was the most represented, including 11 taxons out of 20 (Table 2). Regarding frequency of occurrence, the category of euconstant groups was the most represented, and 50% of the recorded taxonomic groups in both sampling seasons belonged to this category (Table 3).
Table 2.
Relative abundance of taxonomic groups in investigated areas (%—percentage of groups belonging to a certain category of relative abundance).
Table 3.
Frequency of occurrence of taxonomic groups in investigated areas (%—percentage of groups belonging to a certain category of frequency of occurrence).
The taxonomic groups of invertebrates that dominated in samples were amphipod crustaceans, polychaetes, bivalves and gastropods, which constituted 80% of all isolated invertebrates in “winter” samples and 84% in “summer” samples (Figure 5). Certain taxonomic groups were represented only by a few individuals, and their relative abundance was less than 0.1%. These were flatworms, decapods, sea urchins in “winter” samples and cumaceans in “summer” samples.
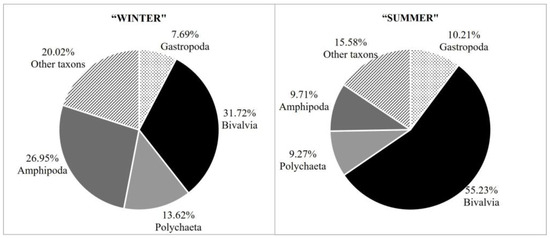
Figure 5.
Dominant taxonomic groups within C. officinalis settlements in all sampling areas during both seasons (“winter” and “summer” having the same definition as in Figure 2).
The four previously mentioned dominant taxonomic groups had the highest value of 100% for both sampling seasons in terms of frequency of occurrence. The remaining euconstant taxonomic groups were almost always the same during both sampling seasons. The exceptions were the polyplacophorans and pantopods, whose frequency of occurrence was classified into two different categories depending on the sampling season. The Polyplacophora group had a frequency of occurrence of 62.96% in the winter samples, which placed it in the lower category, while the Pantopoda group had a frequency of occurrence of 33.33% during the summer (Table 1 and Table 3).
The detailed qualitative and quantitative composition of invertebrates for each separate sampling area during both seasons is shown in Table 4. In addition, the frequency of occurrence and relative abundance of the taxonomic groups were calculated for each area to see the seasonal fluctuation in organisms with respect to different sampling areas. Detailed species lists for mollusks and pycnogonids are available in previously published papers [25,26,33]. In terms of relative abundance, the same four taxonomic groups were again the most dominant. Brijuni National Park is the only area where a higher relative abundance of amphipods over bivalves was recorded. Although in all other areas, regardless of the sampling season, bivalves always dominated, an increase in the relative abundance of this group can be observed if both sampling seasons are compared. During summer, no taxonomic group had a relative abundance higher than 50%, while in winter, bivalves were present in all locations with a relative abundance higher than 50%, i.e., in the range from 54% to 68%.
Table 4.
Number of individuals in each taxonomic group during “winter” (a) and “summer” (b) in the areas of Pula, Banjole, Premantura and Brijuni National Park. Locations are indicated as follows: A—Verudela, B—Saccorgiana, C—Bumbište, D—Cintinera, E—Kamenjak, F—Stupice, G—Javorika, H—Dražice, I—Verige. Each location is described with three replicate quadrats 5 × 5 cm in size.
The relationship between the number of individuals and the dry weight of C. officinalis was studied for the taxonomic groups with the highest number of individuals (gastropods, bivalves, amphipods, tanaidaceans, isopods, mites, polychaetes and nematodes). No correlation was found between the number of bivalves and gastropods and the dry weight of C. officinalis, with low coefficients of determination (R2 = 0.011 for Bivalvia and R2 = 0.005 for Gastropoda) and high probabilities that the regression coefficients were not different from zero (P = 0.318 for Bivalvia and P = 0.511 for Gastropoda) (Figure 6). In contrast, other taxonomic groups showed a significant correlation with the dry weight of C. officinalis, with regression lines explaining up to 12.1% of the total variability in the number of individuals for amphipods (P = 0.001) and 7.9% for nematodes (P = 0.005) (Figure 7). The total number of individuals across all taxonomic groups increased linearly with the algae biomass (P = 0.002), explaining 9.3% of the variability in the number of fauna individuals (Figure 8).
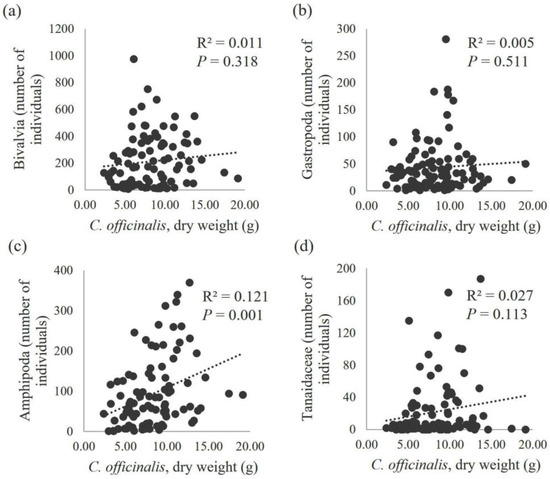
Figure 6.
Scatter diagram of the total number of bivalves (a), gastropods (b), amphipods (c) and tanaidaceans (d) in relation to the dry weight of C. officinalis (g/25 cm2). With the corresponding regression lines (n = 96), the values of the coefficient of determination (R2) and the statistical significance of the slope coefficient (P) are displayed.
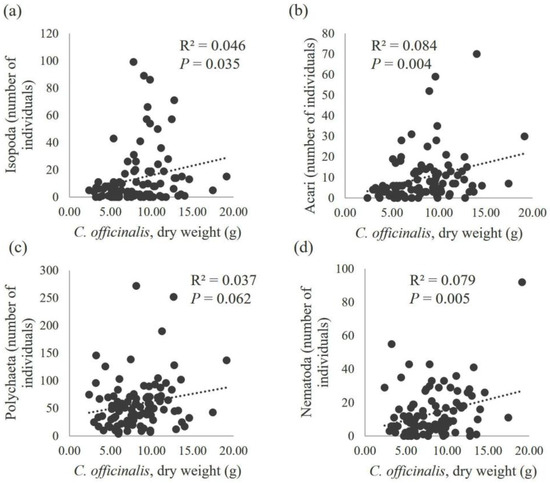
Figure 7.
Scatter diagram of the total number of isopods (a), mites (b), polychaetes (c) and nematodes (d) in relation to the dry weight of C. officinalis (g/25 cm2). With the corresponding regression lines (n = 96), the values of the coefficient of determination (R2) and the statistical significance of the slope coefficient (P) are displayed.
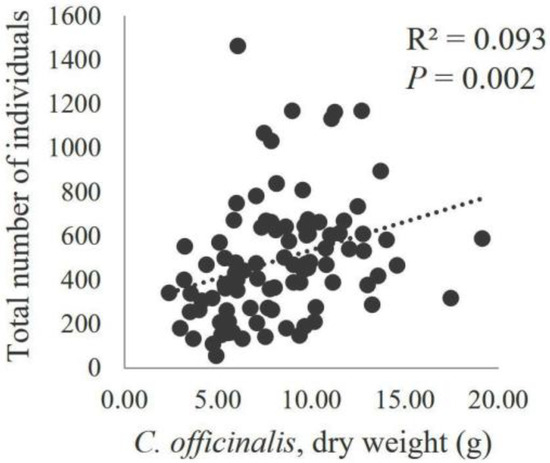
Figure 8.
Scatter diagram of the total number of all recorded invertebrates in relation to the dry weight of C. officinalis (g/25 cm2). With the corresponding regression lines (n = 96), the values of the coefficient of determination (R2) and the statistical significance of the slope coefficient (P) are displayed.
4. Discussion
Previous research on the influence of turf-forming algal biomass on invertebrate species richness shows a positive correlation between the number of invertebrate species and the algal biomass [36]. The greatest diversity and abundance of fauna is always associated with thalli of higher density, which have the most complex structure [50]. Comparing the recorded dry weight of algae in this research with the total number of invertebrates, these numbers follow the same positive trend. Another positive effect of such a habitat has been recorded, which is the fact that algae with this specific structure retain a larger amount of sediment that serves as a secondary habitat for many macrofauna and meiofauna species [37]. In addition, the effect of habitat loss and changes in habitat structure on the number and distribution of species has previously been investigated. It was concluded that the reduction in habitat diversity has a negative effect on the diversity of taxonomic groups, considering that the habitat structure itself greatly affects the spatial distribution of species [8,51]. Research was conducted in an artificial habitat that simulates the appearance of algae to determine the extent to which these morphological changes affect the composition of the macrofauna [22,52,53]. As part of this current research, sampling was performed twice a year, i.e., during the minimum and maximum biomass of C. officinalis, and thus the results from this research confirm the previous findings. Regarding the total number of isolated individuals, 30% more individuals were recorded in samples taken during the algae’s maximum biomass.
Seasonal variability in the composition and structure of the fauna within the algal community is influenced by the structure of the alga itself, its growth rate and its substrate complexity. It also depends on the availability of food, i.e., the amount of detritus and the abundance of epiphytic algae, as well as seasonal changes in population density due to migration of species [54]. The importance of detritus in benthic communities has often been mentioned in the literature, because much more energy and matter are exchanged in detritus than in the food web of organisms that feed on macroalgae. Many organisms can use detritus directly as food, because shortly after microorganisms colonize that detritus, its nutritional value increases significantly [55]. It was observed that differences in density, branching and overall compactness of the genus Corallina have different effects on the accumulation of detritus, and thus on the species living in the intertidal sediments [56]. Small changes in such morphological characteristics, as well as changes in water currents, also affect the abundance of fauna. With its structure and densely branched thallus, Corallina is a trap for sediment and food. Microgastropods that feed on microalgae in the sediment can obtain sufficient food from the sediment located within the algal turf. Sediment in exposed coastal areas is coarser than densely packed sediment in sheltered locations. This makes it susceptible to disturbance caused by seawater movement, which can limit the primary production of microalgae. Grain size can be a limiting factor for primary production, because microphytobenthos bind more strongly to coarser sediment. All this can prevent the survival of organisms that feed on microalgae on such sediment [57] and affect the overall abundance and composition of invertebrates.
Various factors, including diverse reproductive strategies, can also explain fluctuations in species abundance [58]. Species without pelagic larvae, like amphipods and isopods, experience significant abundance variations during recruitment. Additionally, species with larvae that prefer substrate settlement, such as Spirorbis (Spirorbis) corallinae, contribute to increased abundance during settlement periods [59]. The timing of reproduction also plays a role, especially when it is seasonal or intensified during certain times of the year. Many of these species exhibit seasonal reproductive timing, notably gastropods, like those within the Littorina genus [59]. Reproduction may take place throughout the year, with potential reductions in fecundity during the summer [60]. Variations in the timing of reproduction among these direct-developing species can significantly influence their abundance and occurrence, creating distinct seasonal patterns. Another example is the gastropod Rissoella, which develops directly from eggs in the spring and summer, when it can reach high abundance. Eggs are then laid at the end of the summer, which over-winter within the turf and hatch the following year [60,61].
The seasonal dynamics of invertebrates within this research show that the distribution of dominant groups is more uniform during the algae’s maximum growth than during its minimum growth. To be more specific, during summer, bivalves become dominant and, considering the total number of individuals recorded at all research locations, constitute over 55% of the total. Given that the species Mytilus galloprovincialis was by far the most dominant bivalve, and that previous studies recorded the highest density of juvenile individuals of that species in May [16], it is understandable that in the summer period the competition for space within C. officinalis settlements will be on the side of bivalves. They can also eliminate larval/juvenile stages of other potential “settlers” through filtration. A study in southern Italy documented a high occurrence of juvenile M. galloprovincialis in algal settlements in coastal rocky areas, accounting for 96.6% of the total abundance within three fucalean species [28]. The research found adult forms of M. galloprovincialis located under or inside the algal branches, attached to the rocky bottom, where they compete for space with the algae [28].
Analysis of the linear dependence of the total number of individuals of higher taxonomic groups against the dry weight of C. officinalis indicates that while there are certain noteworthy correlations between the abundances of invertebrates and the algal biomass, the explained variance, as reflected by the R2 values, remains relatively low. Consequently, approximately 90% of the variance remains unaccounted for by the regression models. This implies that factors other than the algal biomass are likely to contribute significantly to the observed patterns. One plausible explanation for this unexplained variance is the presence of patchiness in the distribution of algae, both within individual sampling locations and among different sampling locations. It is conceivable that variations in environmental conditions, habitat characteristics or other factors lead to heterogeneity in the distribution and abundance of both invertebrates and algae. The relative scatter of data points within and between sampling locations further supports this notion, offering indications of the potential influence of algal patchiness on the relationships with invertebrate abundances. These findings underscore the need for further investigation into the specific factors driving the unexplained variance and the role of algal patchiness in shaping the relationships between invertebrates and algal biomass.
The comprehensive analysis of invertebrates in this study not only provides valuable insights into the community structure and composition within C. officinalis turfs but also sheds light on the overall biodiversity of the coastal zone of southern Istria, where this alga occupies a significant area. This research recorded that the presence of the algae C. officinalis was almost three times higher than in a previously recorded study conducted in the eastern Adriatic Sea [2], and this indicates that the area of southern Istria is a suitable habitat for this species. The fact that the habitat is so widespread in the coastal area of southern Istria, and that the abundance of invertebrates is high therein, contributed to the need to conduct this research. Previous studies worldwide have documented the presence of numerous invertebrates within C. officinalis settlements, with reported values of up to 329,000 individuals per square meter [21]. Comparing those findings to the maximum (586,000 ind m−2) and average (192,600 ind m−2) number of individuals recorded in our research, it becomes evident that this habitat holds immense importance due to its high abundance and diversity of invertebrates.
Moreover, by considering clustering techniques to assess similarity in invertebrate assemblages among the research locations, further insights into the factors influencing the variation in invertebrate community composition can be gained. Exploring potential relationships between environmental variables, algal biomass, patchiness and other relevant habitat characteristics, as well as the observed clustering patterns, valuable information on invertebrate assemblage variations can be provided. These analyses may reveal the role of localized factors, spatial dynamics and ecological interactions in shaping the community structure within C. officinalis turfs and contribute to a deeper understanding of the ecological processes governing coastal ecosystems.
This study not only enhances our understanding of the community structure and composition within C. officinalis turfs but also highlights the significant biodiversity and the abundance of invertebrates in this habitat.
Author Contributions
Conceptualization, M.B. and L.I.; data curation, M.B., L.I., A.J. and M.A.P.; formal analysis, M.B. and L.I.; funding acquisition, E.P.; investigation, M.B., L.I., A.J., M.A.P., N.I. and P.P.; methodology, M.B. and L.I.; resources, M.B., L.I., A.J., M.A.P. and E.P.; writing—original draft preparation, M.B., L.I., A.J., M.A.P., B.B.M., L.N., E.P., P.B., N.I. and P.P.; writing—review and editing, M.B., L.I., A.J., B.B.M., P.B. and L.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The authors can provide the data if needed.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Williamson, C.J.; Perkins, R.; Voller, M.; Yallop, M.L.; Brodie, J. The regulation of coralline algal physiology, an in situ study of Corallina officinalis (Corallinales, Rhodophyta). Biogeosciences 2017, 14, 4485–4498. [Google Scholar] [CrossRef]
- Nikolić, V.; Žuljević, A.; Mangialajo, L.; Antolić, B.; Kušpilić, G.; Ballesteros, E. Cartography of littoral rocky-shore communities (CARLIT) as a tool for ecological quality assessment of coastal waters in the Eastern Adriatic Sea. Ecol. Indic. 2013, 34, 87–93. [Google Scholar] [CrossRef]
- Bertness, M.D.; Crain, C.M.; Silliman, B.R.; Bazterrica, M.C.; Reyna, M.V.; Hildago, F.; Farina, J.K. The community structure of western Atlantic Patagonian rocky shores. Ecol. Monogr. 2006, 76, 439–460. [Google Scholar] [CrossRef]
- Liuzzi, M.; Gappa, J.L. Macrofaunal assemblages associated with coralline turf: Species turnover and changes in structure at different spatial scales. Mar. Ecol. Prog. Ser. 2008, 363, 147–156. [Google Scholar] [CrossRef]
- Nelson, W.A. Calcified macroalgae—Critical to coastal ecosystems and vulnerable to change: A review. Mar. Freshw. Res. 2009, 60, 787–801. [Google Scholar] [CrossRef]
- Bracken, M.E.S.; Gonzalez-Dorantes, C.A.; Stachowicz, J.J. Whole-community mutualism: Associated invertebrates facilitate a dominant habitat-forming seaweed. Ecology 2007, 88, 2211–2219. [Google Scholar] [CrossRef] [PubMed]
- Heck, K., Jr.; Crowder, L.B. habitat structure and predator—Prey interactions in vegetated aquatic systems. In Habitat Structure: The Physical Arrangement of Objects in Space; Springer: Dordrecht, The Netherlands, 1991; pp. 281–299. [Google Scholar]
- Gallardo, D.; Oliva, F.; Ballesteros, M. Marine invertebrate epibionts on photophilic seaweeds: Importance of algal architecture. Mar. Biodivers. 2021, 51, 16. [Google Scholar] [CrossRef]
- Orlando-Bonaca, M.; Trkov, D.; Klun, K.; Pitacco, V. Diversity of Molluscan Assemblage in Relation to Biotic and Abiotic Variables in Brown Algal Forests. Plants 2022, 11, 2131. [Google Scholar] [CrossRef]
- Bégin, C.; Johnson, L.; Himmelman, J. Macroalgal canopies: Distribution and diversity of associated invertebrates and effects on the recruitment and growth of mussels. Mar. Ecol. Prog. Ser. 2004, 271, 121–132. [Google Scholar] [CrossRef]
- Costa-Lotufo, L.V.; Colepicolo, P.; Pupo, M.T.; Palma, M.S. Bioprospecting macroalgae, marine and terrestrial invertebrates & their associated microbiota. Biota Neotropica 2022, 22, 1345. [Google Scholar] [CrossRef]
- Choi, J.-Y.; Kim, S.-K.; Jeong, K.-S.; Joo, G.-J. Distribution pattern of epiphytic microcrustaceans in relation to different macrophyte microhabitats in a shallow wetland (Upo wetlands, South Korea). Oceanol. Hydrobiol. Stud. 2015, 44, 151–163. [Google Scholar] [CrossRef]
- Guerra García, J.M.; Cabezas Rodríguez, M.D.P.; Baeza-Rojano Pageo, E.; Izquierdo, D.; Corzo, J.; Ros Clemente, M.; Sánchez, J.A.; Dugo Cota, Á.; Flores León, A.M.; Soler Hurtado, M.D.M. Abundance patterns of macrofauna associated to marine macroalgae along the Iberian Peninsula. Zool. Baetica 2011, 22, 3–17. [Google Scholar]
- Schmidt, A.L.; Scheibling, R.E. Effects of native and invasive macroalgal canopies on composition and abundance of mobile benthic macrofauna and turf-forming algae. J. Exp. Mar. Biol. Ecol. 2007, 341, 110–130. [Google Scholar] [CrossRef]
- Koehl, M.; Daniel, T.L. Hydrodynamic Interactions Between Macroalgae and Their Epibionts. Front. Mar. Sci. 2022, 9, 872960. [Google Scholar] [CrossRef]
- Bulleri, F.; Pardi, G.; Tamburello, L.; Ravaglioli, C. Nutrient enrichment stimulates herbivory and alters epibiont assemblages at the edge but not inside subtidal macroalgal forests. Mar. Biol. 2020, 167, 1–15. [Google Scholar] [CrossRef]
- Burnett, N.P.; Koehl, M.A.R. Ecological biomechanics of damage to macroalgae. Front. Plant Sci. 2022, 13, 981904. [Google Scholar] [CrossRef]
- George, J.D. The polychaetes of Lewis and Harris with notes on other marine invertebrates. Proc. R. Soc. Edinb. Sect. B Biol. Sci. 1979, 77, 189–216. [Google Scholar] [CrossRef]
- Johnson, S.B.; Attramadal, Y.G. Reproductive behaviour and larval development of Tanais cavolinii (Crustacea: Tanaidacea). Mar. Biol. 1982, 71, 11–16. [Google Scholar] [CrossRef]
- López, C.A.; Stotz, W.B. Description of the fauna associated with Corallina officinalis L. in the intertidal of the rocky shore of Palo Colorado (Los Vilos, IV-region, Chile). Oceanogr. Lit. Rev. 1998, 3, 512. [Google Scholar]
- Bussell, J.A.; Lucas, I.A.; Seed, R. Patterns in the invertebrate assemblage associated with Corallina officinalis in tide pools. J. Mar. Biol. Assoc. United Kingd. 2007, 87, 383–388. [Google Scholar] [CrossRef]
- Kelaher, B.P.; Castilla, J.C.; Prado, L.; York, P.; Schwindt, E.; Bortolus, A. Spatial variation in molluscan assemblages from coralline turfs of Argentinean Patagonia. J. Molluscan Stud. 2007, 73, 139–146. [Google Scholar] [CrossRef]
- Magill, C.L.; Maggs, C.A.; Johnson, M.P.; O’connor, N. Sustainable Harvesting of the Ecosystem Engineer Corallina officinalis for Biomaterials. Front. Mar. Sci. 2019, 6, 285. [Google Scholar] [CrossRef]
- Buršić, M.; Iveša, L.; Jaklin, A.; Pijevac, M.A. A preliminary study on the diversity of invertebrates associated with Corallina officinalis Linnaeus in southern Istrian peninsula. Acta Adriat. 2019, 60, 127–136. [Google Scholar] [CrossRef]
- Buršić, M.; Iveša, L.; Jaklin, A.; Pijevac, M.A.; Kučinić, M.; Štifanić, M.; Neal, L.; Mađarić, B.B. DNA Barcoding of Marine Mollusks Associated with Corallina officinalis Turfs in Southern Istria (Adriatic Sea). Diversity 2021, 13, 196. [Google Scholar] [CrossRef]
- Buršić, M.; Iveša, L.; Jaklin, A.; Pijevac, M.A.; Mađarić, B.B.; Neal, L.; Pustijanac, E.; Burić, P.; Iveša, N.; Paliaga, P. Changes in Composition of Mollusks within Corallina officinalis Turfs in South Istria, Adriatic Sea, as a Response to Anthropogenic Impact. Diversity 2023, 15, 939. [Google Scholar] [CrossRef]
- Terlizzi, A.; Scuderi, D.; Fraschetti, S.; Guidetti, P.; Boero, F. Molluscs on subtidal cliffs: Patterns of spatial distribution. J. Mar. Biol. Assoc. United Kingd. 2003, 83, 165–172. [Google Scholar] [CrossRef]
- Chiarore, A.; Fioretti, S.; Meccariello, A.; Saccone, G.; Patti, F.P. Molluscs community associated with the brown algae of the genus Cystoseira in the Gulf of Naples (South Tyrrhenian Sea). BioRxiv 2017, 160200. [Google Scholar] [CrossRef]
- Laakkonen, H.M.; Strelkov, P.; Väinölä, R. Molecular lineage diversity and inter-oceanic biogeographical history in Hiatella (Mollusca, Bivalvia). Zool. Scr. 2015, 44, 383–402. [Google Scholar] [CrossRef]
- Sánchez-Moyano, J.; Estacio, F.; García-Adiego, E.; García-Gómez, J. The molluscan epifauna of the alga halopteris scoparia in southern Spain as a bioindicator of coastal environmental conditions. J. Molluscan Stud. 2000, 66, 431–448. [Google Scholar] [CrossRef]
- Izquierdo, D.; Guerra-García, J.M. Distribution patterns of the peracarid crustaceans associated with the alga Corallina elongata along the intertidal rocky shores of the Iberian Peninsula. Helgol. Mar. Res. 2011, 65, 233–243. [Google Scholar] [CrossRef]
- Guerra-García, J.M.; de Figueroa, J.M.T.; Barranco, C.N.; Ros, M.; Sanchez-Moyano, J.E.; Moreira, J. Dietary analysis of the marine Amphipoda (Crustacea: Peracarida) from the Iberian Peninsula. J. Sea Res. 2014, 85, 508–517. [Google Scholar] [CrossRef]
- Lehmann, T.; Spelda, J.; Melzer, R.; Buršić, M. Pycnogonida (Arthropoda) from Northern Adriatic Corallina officinalis Linnaeus, 1758 belts. Mediterr. Mar. Sci. 2020, 22, 102–107. [Google Scholar] [CrossRef]
- Esquete, P.; Rubal, M.; Veiga, P.; Troncoso, J. New records of Sea Spiders (Arthropoda: Pycnogonida) for continental Portugal and notes on species distribution. Mar. Biodivers. Rec. 2016, 9, 1–5. [Google Scholar] [CrossRef]
- Irwin, S.; Davenport, J. Oxygen microenvironment of coralline algal tufts and their associated epiphytic animals. In Biology and Environment: Proceedings of the Royal Irish Academy; Royal Irish Academy: Dublin, Ireland, 2010; pp. 185–193. [Google Scholar]
- Matias, M.G.; Arenas, F.; Rubal, M.; Pinto, I.S. Macroalgal Composition Determines the Structure of Benthic Assemblages Colonizing Fragmented Habitats. PLoS ONE 2015, 10, e0142289. [Google Scholar] [CrossRef]
- Airoldi, L.; Cinelli, F. Effects of sedimentation on subtidal macroalgal assemblages: An experimental study from a mediterranean rocky shore. J. Exp. Mar. Biol. Ecol. 1997, 215, 269–288. [Google Scholar] [CrossRef]
- Nordsieck, F. Die Europäischen Meeres-Gehäuseschnecken (Prosobranchia); Gustav Fischer Verlag: Stuttgart, Germany, 1968; pp. 1–273. [Google Scholar]
- Nordsieck, F. Die Europäischen Meeresmuscheln (Bivalvia); Gustav Fisher Verlag: Stuttgart, Germany, 1969; pp. 1–256. [Google Scholar]
- Parenzan, P. Gasteropodi. In Carta d’Identità delle Conchiglie del Mediterraneo; Bios Taras: Taranto, Italy, 1970; Volume 1, pp. 1–283. [Google Scholar]
- Parenzan, P. Bivalves. In Carta d’Identità delle Conchiglie del Mediterraneo; Bios Taras: Taranto, Italy, 1974; Volume 2, pp. 1–277. [Google Scholar]
- Sabelli, B.; Gianuzzi-Savelli, R.; Bedulli, D. Catalogo Annotato dei Molluschi Marini del Mediterraneo; Libreria Naturalistica Bolognese: Bologna, Italy, 1990; Volume 1, pp. 1–348. [Google Scholar]
- Poppe, G.T.; Goto, Y. Scaphopoda, Bivalvia, Cephalopoda. In European Seashells; Verlag Christa Hemmen: Wiesbaden, Germany, 1993; Volume 2, pp. 1–221. [Google Scholar]
- Gianuzzi-Savelli, R.; Pusateri, F.; Palmeri, A.; Ebreo, C. Atlante delle Conchiglie Marine del Mediterraneo; La Conchiglia: Roma, Italy, 1996; pp. 1–258. [Google Scholar]
- Gofas, S.; Moreno, D.; Salas, C. (Eds.) Moluscos marinos de Andalucía; Universidad de Málaga: Málaga, Spain, 2011; Volume 1, pp. 1–342. [Google Scholar]
- Gofas, S.; Moreno, D.; Salas, C. (Eds.) Moluscos marinos de Andalucía; Universidad de Málaga: Málaga, Spain, 2011; Volume 2, pp. 343–798. [Google Scholar]
- Tischler, W. Grundzüge der Terrestrischen Tierökologie; Friedrich Vieweg und Sohn: Braunschweig, Germany, 1949; 219p. [Google Scholar] [CrossRef]
- Travizi, A. The nematode fauna of the northern Adriatic offshore sediments: Community structure and biodiversity. Acta Adriat. 2010, 51, 169–180. [Google Scholar]
- Năstase, A.; Honț, S.; Iani, M.; Paraschiv, M.; Cernișencu, I.; Năvodaru, I. Ecological status of fish fauna from Razim Lake and the adjacent area, the Danube Delta Biosphere Reserve, Romania. Acta Ichthyol. Piscat. 2022, 52, 43–52. [Google Scholar] [CrossRef]
- Kelaher, B.P. Changes in habitat complexity negatively affect diverse gastropod assemblages in coralline algal turf. Oecologia 2003, 135, 431–441. [Google Scholar] [CrossRef]
- Matias, M.; Underwood, T.; Hochuli, D.; Coleman, R. Habitat identity influences species—Area relationships in heterogeneous habitats. Mar. Ecol. Prog. Ser. 2011, 437, 135–145. [Google Scholar] [CrossRef]
- Kelaher, B. Influence of physical characteristics of coralline turf on associated macrofaunal assemblages. Mar. Ecol. Prog. Ser. 2002, 232, 141–148. [Google Scholar] [CrossRef]
- Lavender, J.T.; Dafforn, K.A.; Bishop, M.J.; Johnston, E.L. Small-scale habitat complexity of artificial turf influences the development of associated invertebrate assemblages. J. Exp. Mar. Biol. Ecol. 2017, 492, 105–112. [Google Scholar] [CrossRef]
- Urra, J.; Rueda, J.; Ramírez, M.; Marina, P.; Tirado, C.; Salas, C.; Gofas, S. Seasonal variation of molluscan assemblages in different strata of photophilous algae in the Alboran Sea (western Mediterranean). J. Sea Res. 2013, 83, 83–93. [Google Scholar] [CrossRef]
- Mann, K.H. Production and use of detritus in various freshwater, estuarine, and coastal marine ecosystems. Limnol. Oceanogr. 1988, 33, 910–930. [Google Scholar] [CrossRef]
- Olabarria, C.; Chapman, M. Comparison of patterns of spatial variation of microgastropods between 2 contrasting intertidal habitats. Mar. Ecol. Prog. Ser. 2001, 220, 201–211. [Google Scholar] [CrossRef]
- Olabarria, C.; Chapman, M. Habitat-associated variability in survival and growth of three species of microgastropods. J. Mar. Biol. Assoc. United Kingd. 2001, 81, 961–966. [Google Scholar] [CrossRef]
- Hicks, G.R.F. Meiofauna associated with rocky shore algae. In The Ecology of Rocky Coasts; Moore, P.G., Seed, R., Eds.; Hodder and Stoughton: London, UK, 1985; pp. 36–56. [Google Scholar]
- Bussell, J.A. Biodiversity of the Invertebrate Community Associated with the Turf-Forming Red Alga Corallina officinalis in Tide Pools; Bangor University: London, UK, 2003; pp. 1–248. [Google Scholar]
- Graham, A. Molluscs: Prosobranch and Pyramidellid Gastropods: Keys and Notes for the Identification of the Species; E.J. Brill/Dr. W. BackhuYs: Leiden, Germany, 1988; pp. 1–662. [Google Scholar]
- Hayward, P.J.; Ryland, J.S. Handbook of the Marine Fauna of North-West Europe; Oxford University Press: Oxford, UK, 1995; pp. 1–799. [Google Scholar]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).