
Exploring the Genetic Diversity and Population Structure of Daphnia cucullata Sars, 1862 in Boreal Lakes (Latvian Lakeland) Based on Microsatellites


Abstract
1. Introduction
2. Material and Methods
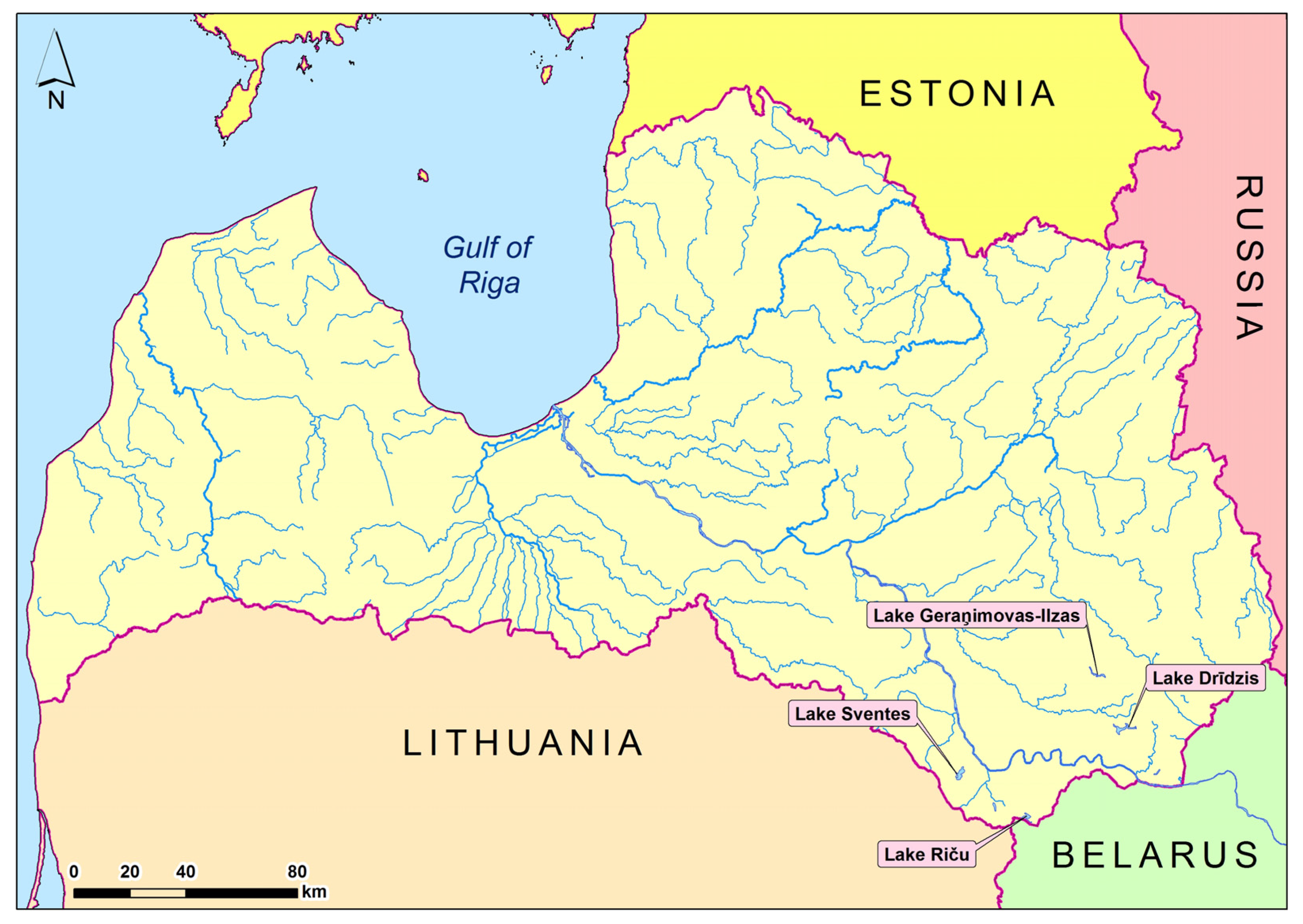
2.1. Sampling Sites and Material Collection of Daphnia cucullata
2.2. Genetic Analysis
2.2.1. DNA Extraction
2.2.2. Determination of the Quantity and Quality of Isolated DNA
2.2.3. Microsatellites Analysis
2.2.4. Statistical Processing and Analysis of the Obtained Data
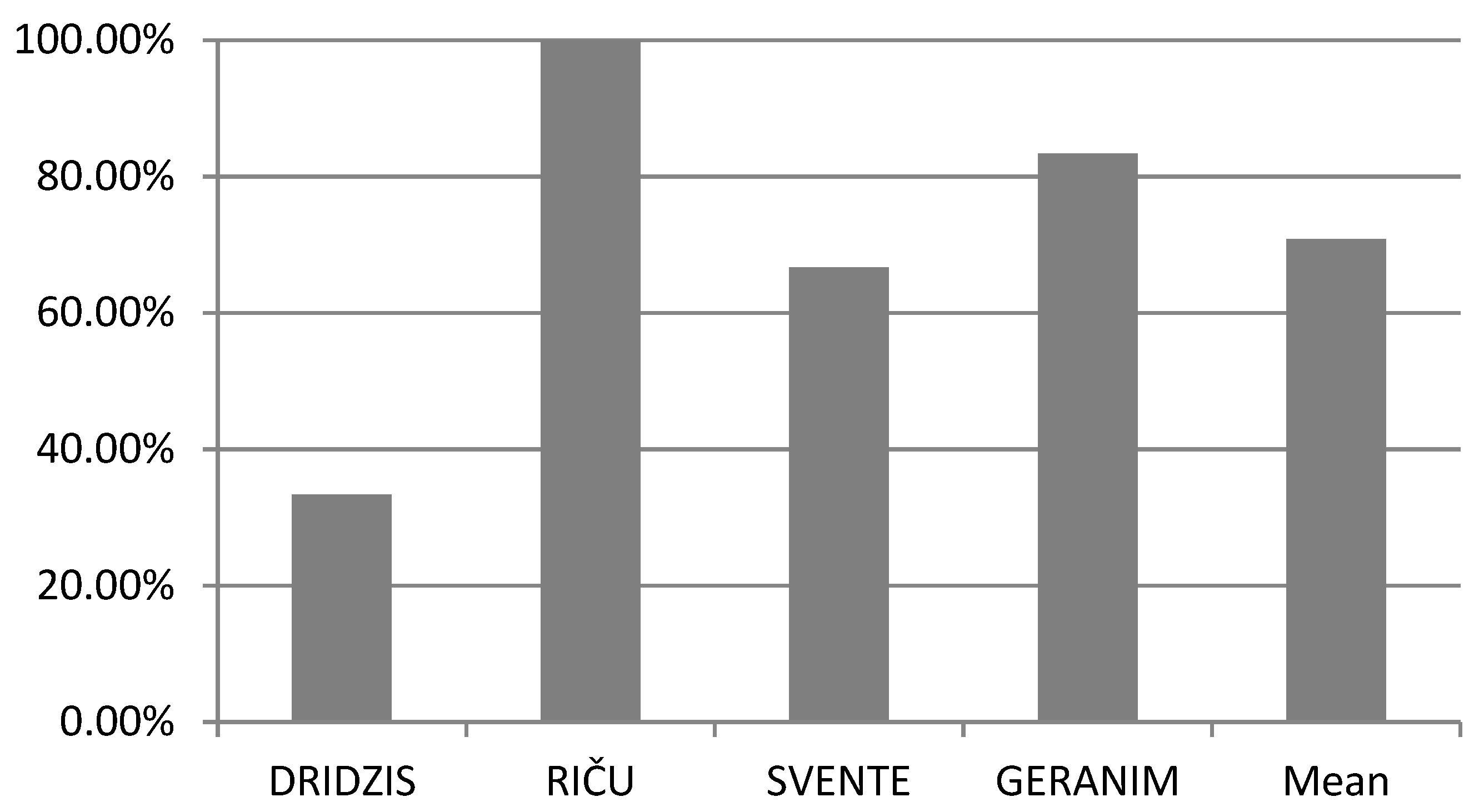
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hebert, P.D.N. Competition in zooplankton communities. Ann. Zool. Fenn. 1982, 19, 349–356. [Google Scholar]
- Malone, B.J.; McQueen, D.J. Horizontal patchiness in zooplankton populations in two Ontario kettle lakes. Hydrobiologia 1983, 99, 101–124. [Google Scholar] [CrossRef]
- Pinel-Alloul, B. Spatial heterogeneity as a multiscale characteristic of zooplankton community. Hydrobiologia 1995, 300, 17–42. [Google Scholar] [CrossRef]
- Wetzel, R.G. Limnology. In Lake and River Ecosystems, 3rd ed.; Academic Press: Cambridge, MA, USA, 2001; p. 1006. [Google Scholar]
- Karpowicz, M.; Sługock, Ł.; Kozłowska, J.; Ochocka, A.; López, C. Body size of Daphnia cucullata as an indicator of the ecological status of temperate lakes. Ecol. Indic. 2020, 117, 1–10. [Google Scholar] [CrossRef]
- Adamowicz, S.J.; Hebert, P.D.N.; Marinone, M.C. Species diversity and endemism in the Daphnia of Argentina: A genetic investigation. Zool. J. Linn. Soc. Lond. 2004, 140, 171–205. [Google Scholar] [CrossRef]
- Colbourne, J.K.; Hebert, P.D.N. The systematics of North American Daphnia (Crustacea: Anomopoda): A molecularphylogenetic approach. Philos. Trans. R. Soc. Lond. Ser. B 1996, 351, 349–360. [Google Scholar]
- Colbourne, J.K.; Wilson, C.C.; Hebert, P.D.N. The systematics of Australian Daphnia and Daphniopsis (Crustacea: Cladocera): A shared phylogenetic history transformed by habitat-specific rates of evolution. Biol. J. Linn. Soc. 2006, 89, 469–488. [Google Scholar] [CrossRef][Green Version]
- Dussart, B.H.; Defaye, D. Introduction to the Copepoda. Guides to the Identification of the Microinvertebrates of the Continental Waters of the World, 2nd ed.; Backhuys Publishers: Leiden, The Netherlands, 2001; p. 344. [Google Scholar]
- Harris, K.D.M.; Bartlett, N.J.; Lloyd, V.K. Daphnia as an emerging epigenetic model organism. Genet. Res. Int. 2012, 2012, 147892. [Google Scholar] [CrossRef]
- Lampert, W. Daphnia: Model herbivore, predator and prey. Pol. J. Ecol. 2006, 54, 607–620. [Google Scholar]
- Colbourne, J.K.; Hebert, P.D.N.; Taylor, D.J. Evolutionary origins of phenotypic diversity in Daphnia. In Molecular Evolution and Adaptive Radiation; Givnish, T.J., Sytsma, K.J., Eds.; Cambridge University Press: London, UK, 1997; pp. 163–188. [Google Scholar]
- Dodson, S.I.; Hanazato, T. Commentary on effects of anthropogenic and natural organic-chemicals on development, swimming behavior and reproduction of Daphnia, a key member of aquatic ecosystems. Environ. Health Perspect. 1995, 103, 7–11. [Google Scholar]
- Brede, N.; Thielsch, A.; Sandrock, C.; Spaak, P.; Keller, B.; Streit, B.; Schwenk, K. Microsatellite markers for European Daphnia. Mol. Ecol. Notes 2006, 6, 536–539. [Google Scholar] [CrossRef]
- Cousyn, C.; De Meester, L.; Colbourne, J.K.; Brendonck, L.; Verschuren, D.; Volckaert, F. Rapid, local adaptation of zooplankton behavior to changes in predation pressure in the absence of neutral genetic changes. Proc. Natl. Acad. Sci. USA 2001, 98, 6256–6260. [Google Scholar] [CrossRef] [PubMed]
- De Meester, L.; Weider, L.J.; Tollrian, R. Alternative antipredator defences and genetic polymorphism in a pelagic predator-prey system. Nature 1995, 378, 483–485. [Google Scholar] [CrossRef]
- Harris, R.P.; Wiebe, P.H.; Lenz, J.; Skjoldal, H.R.; Huntley, M. Zooplankton Methodology Manual; Elsevier Academic Press: Cambridge, MA, USA, 2005; pp. 533–570. [Google Scholar]
- Hellsten, M.E.; Sundberg, P. Genetic variation in two sympatric European populations of Bosmina spp. (Cladocera) tested with RAPD markers. Hydrobiologia 2000, 421, 157–164. [Google Scholar] [CrossRef]
- Brakovska, A. Daphnia cucullata Sars, 1862 (CRUSTACEA: CLADOCERA) distribution and location in composition of zooplankton cenosis in Lake Dridzis. Acta Biol. Univ. Daugavp. 2014, 14, 1–19. [Google Scholar]
- Brakovska, A.; Škute, R. Ecological characterization of zooplankton groups in the deepest lakes of East Latvia. Acta Biol. Univ. Daugavp. 2007, 7, 165–174. [Google Scholar]
- Brakovska, A.; Škute, R. Ecological evaluation of zooplankton groups in Lake Geranimovas-Ilzas and Lake Garais. Proc. 7th Int. Sci. Pract. Conf. Environment. Technol. Resour. 2009, 2, 43–50. [Google Scholar] [CrossRef][Green Version]
- Brakovska, A.; Škute, R.; Škute, A. Heterogeneity of distribution and community composition of zooplankton in upper layers of Lake Svente. Zool. Ecol. 2012, 22, 172–180. [Google Scholar] [CrossRef]
- Brakovska, A.; Škute, N. Optimisation of DNA extraction and RAPD-PCR amplification for population genetic analysis of Daphnia cucullata Sars, 1862 (Crustacea: Cladocera). Acta Biol. Univ. Daugavp. 2013, 13, 11–20. [Google Scholar]
- Brakovska, A.; Paidere, J.; Škute, A. Dynamics and factors influencing zooplankton in the lakes Svente, Riča, Dridzis and Geraņimovas-Ilzas (Eastern Latvia). Acta Biol. Univ. Daugavp. 2020, 20, 71–94. [Google Scholar]
- Selkoe, K.A.; Toonen, R.J. Microsatellites for ecologists: A practical guide to using and evaluating microsatellite markers. Ecol. Lett. 2006, 9, 615–629. [Google Scholar] [CrossRef] [PubMed]
- Haag, T.; Santos, A.S.; Sana, D.A.; Morato, R.G.; Cullen, L., Jr.; Crawshaw, P.G., Jr.; De Angelo, C.; Di Bitetti, M.S.; Salzano, F.M.; Eizirik, E. The effect of habitat fragmentation on the genetic structure of a top predator: Loss of diversity and high differentiation among remnant populations of Atlantic Forest jaguars (Panthera onca). Mol. Ecol. 2010, 22, 4906–4921. [Google Scholar] [CrossRef] [PubMed]
- Colbourne, J.K.; Robison, B.; Bogart, K.; Lynch, M. Five hundred and twenty-eight microsatellite markers for ecological genomic investigations using Daphnia. Mol. Ecol. Notes 2004, 4, 485–490. [Google Scholar] [CrossRef]
- Frisch, D.; Morton, P.K.; Roy Chowdhury, P.; Culver, B.W.; Colbourne, J.K.; Weider, L.J.; Jeyasingh, P.D. A millennial-scale chronicle of evolutionary responses to cultural eutrophication in Daphnia. Ecol. Lett. 2014, 17, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Forest, F.; Chase, M.W.; Persson, C.; Crane, P.R.; Hawkins, J.A. The role of biotic and abiotic factors in evolution of ant dispersal in the milkwort family (polygalaceae). Evolution 2007, 61, 1675–1694. [Google Scholar] [CrossRef] [PubMed]
- Pálsson, S. Microsatellite variation in Daphnia pulex from both sides of the Baltic Sea. Mol. Ecol. 2000, 9, 1075–1088. [Google Scholar] [CrossRef]
- Ender, A.; Schwenk, K.; Stadler, T.; Streit, B.; Schierwater, B. RAPD identification of microsatellites in Daphnia. Mol. Ecol. 1996, 5, 437–441. [Google Scholar] [CrossRef]
- Fox, J.A. New microsatellite primers for Daphnia galeata mendotae. Mol. Ecol. Notes 2004, 4, 544–546. [Google Scholar] [CrossRef]
- Lynch, M.; Spitze, K. Evolutionary genetics of Daphnia. In Ecological Genetics; Real, L., Ed.; Princeton University Press: Princeton, NJ, USA, 1994; pp. 109–128. [Google Scholar]
- Schwenk, K.; Spaak, P. Evolutionary and ecological consequences of interspecific hybridization in cladocerans. Experientia 1995, 51, 465–481. [Google Scholar] [CrossRef]
- Lynch, M.; Pfrender, M.; Spitze, K.; Lehman, N.; Hicks, J.; Allen, D.; Latta, L.; Ottene, M.; Bogue, F.; Colbourne, J. The quantitative and molecular genetic architecture of a subdivided species. Evolution 1999, 53, 100–110. [Google Scholar] [CrossRef]
- Mergeay, J.; Verschuren, D.; De Meester, L. Cryptic invasion and dispersal of an American Daphnia in East Africa. Limnol. Ocean. 2005, 50, 1278–1283. [Google Scholar] [CrossRef]
- Silva, R.M.; Pereira, F.; Carneiro, J.; Sobral, O.; Ribeiro, R.; Amorim, A.; Soares, A.M.V.M.; Lopes, I. Microevolution in a Natural Population of Daphnia longispina Exposed to Acid Mine Drainage. In Interdisciplinary Studies on Environmental Chemistry—Biological Responses to Contaminants; Hamamura, N., Suzuki, S., Mendo, S., Barroso, C.M., Iwata, H., Tanabe, S., Eds.; Terrapub: Pozzuoli, Italy, 2010; pp. 213–218. [Google Scholar]
- Reid, V.A.; Carvalho, G.R.; George, D.G. Molecular genetic analysis of Daphnia in the English Lake District: Species identity, hybridisation and resting egg banks. Freshw. Biol. 2020, 44, 247–253. [Google Scholar] [CrossRef]
- Urtāne, L. Cladocera as types and trophic status indicators of Latvian Lakes. Thesis 1998, 168. Not published (In Latvian) [Google Scholar]
- Available online: https://www.ezeri.lv/database (accessed on 24 January 2023).
- Eipurs, I. Lake Geraņimovas-Ilzas. The Nature of Latvia. Riga Encycl. Latv. 1995, 2, 106. (In Latvian) [Google Scholar]
- Tidriķis, A. Lake Svente. In The Nature of Latvia; Riga, Publishing House: Riga, Latvia, 1998; Volume 5, p. 180. (In Latvian) [Google Scholar]
- Tidriķis, A. Lake Riču. In The Nature of Latvia; Riga, Publishing House: Riga, Latvia, 1997; Volume 4, pp. 243–244. (In Latvian) [Google Scholar]
- APHA. Standart Methods for the Examination of Waters and Wastewater, 21st ed.; American Public Health Association: Washington, DC, USA, 2005. [Google Scholar]
- Wetzel, R.G.; Likens, G.E. Limnological Analyses; Springer Science, Business Media: New York, NY, USA, 2000; p. 429. [Google Scholar]
- Fitzsimmons, J.M.; Innes, D.M. No evidence of Wolbachia among Great Lakes area populations of Daphnia pulex (Crustacea: Cladocera). J. Plankton Res. 2005, 27, 121–124. [Google Scholar] [CrossRef]
- Brakovska, A.; Paidere, J.; Škute, R.; Škute, N.; Škute, A. Occurrence of Cladocera and genetic diversity of Daphnia cucullata in pelagic zone of the Latvian salmonid lakes. Est. J. Ecol. 2013, 62, 244–264. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Yeh, F.C.; Yang, R.C.; Boyle, T. Population Genetics Software, POPGENE 32-version 1.31: Microsoft Window-Based Freeware for Population Genetics Analysis; University of Alberta: Edmonton, AB, Canada, 1999.
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987; p. 512. [Google Scholar]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef]
- Nei, M.; Tajima, F.; Tateno, Y. Accurancy of estimated phylogenetic trees from molecular data. 2 Genefrequency data. J. Mol. Evol. 1983, 19, 153–170. [Google Scholar] [CrossRef]
- Hubisz, M.J.; Falush, D.; Stephens, M.; Pritchard, A.J. Interring weak population structure with the assistance of sample group information. Mol. Ecol. Resour. 2009, 9, 1322–1332. [Google Scholar] [CrossRef]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Nei, M. Estimation of Average Heterozygosity and Genetic Distance from a small number of Individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Biogeographical Regions in Europe—European Environment Agency. Available online: europa.eu (accessed on 24 January 2023).
- De Meester, L.; Vanoverbeke, J.; De Gelas, K.; Ortells, R.; Spaak, P. Genetic structure of cyclic parthenogenetic zooplankton populations—A conceptual framework. Arch. Für Hydrobiol. 2006, 167, 217–244. [Google Scholar] [CrossRef]
- De Meester, L.; Gómez, A.; Simon, J.C. Evolutionary and ecological genetics of cyclical parthenogens. In Evolution from Molecules to Ecosystems; Moya, A., Font, E., Eds.; Oxford University Press: Oxford, UK, 2004; pp. 122–134. [Google Scholar]
- Kalff, J. Limnology: Inland Water Ecosystems; Prentice-Hall: Upper Saddle River, NJ, USA, 2002; p. 592. [Google Scholar]
- Lampert, W.; Sommer, U. Limnoecology: The Ecology of Lakes and Streams; Oxford University Press: New York, NY, USA; Oxford, UK, 1997; p. 382. [Google Scholar]
- Haag, C.R.; Riek, M.; Hottinger, J.W.; Pajunen, V.I.; Ebert, D. Genetic diversity and genetic differentiation in Daphnia metapopulations with subpopulations of known age. Genetics 2005, 170, 1809–1820. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.J.; Hebert, P.D.N. Habitat dependent hybrid parentage and differential introgression between neighboringly sympatric Daphnia species. Proc. Natl. Acad. Sci. USA 1993, 90, 7079–7083. [Google Scholar] [CrossRef]
- Taylor, D.J.; Hebert, P.D.N. A reappraisal of phenotypic variation in Daphnia galeata mendotae: The role of interspecific hybridization. Can. J. Fish. Aquat. Sci. 1993, 50, 2137–2146. [Google Scholar] [CrossRef]
- Green, A.J.; Figuerola, J. Recent advances in the study of longdistance dispersal of aquatic invertebrates via birds. Divers. Distrib. 2005, 11, 149–156. [Google Scholar] [CrossRef]
- Figuerola, J.; Green, A.J.; Santamaria, L. Passive internal transport of aquatic organisms by waterfowl in Doñana, south-west Spain. Glob. Ecol. Biogeogr. 2003, 12, 427–436. [Google Scholar] [CrossRef]
- Figuerola, J.; Green, A.J.; Michot, T.C. Invertebrate eggs can fly: Evidence of waterfowl-mediated gene flow in aquatic invertebrates. Am. Nat. 2005, 165, 274–280. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Moran, C. Enzyme variability in natural populations of Daphnia carinata King. Heredity 1980, 45, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Crease, T.J.; Lynch, M.; Spitze, K. Hierarchical analysis of population genetic variation in mitochondrial and nuclear genes of Daphnia pulex. Mol. Biol. Evol. 1990, 7, 444–458. [Google Scholar] [PubMed]
- Gomez, A.; Carvalho, G.R. Sex, parthenogenesis and genetic structure of rotifers: Microsatellite analysis of contemporary and resting egg bank populations. Mol. Ecol. 2000, 9, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Vanoverbeke, J.; De Meester, L. Among-populational genetic differentiation in the cyclical parthenogen Daphnia magna (Crustacea: Anomopoda) and its relation to geographic distance and clonal diversity. Hydrobiologia 1997, 126, 135–142. [Google Scholar] [CrossRef]
- Decaestecker, E.; De Meester, L.; Mergeay, J. Cyclical Parthenogenesis in Daphnia: Sexual Versus Asexual Reproduction. Lost Sex 2009, 295–316. [Google Scholar]
- Hughes, R.N. A Functional Biology of Clonal Animals; Chapman & Hall: London, UK; New York, NY, USA, 1989. [Google Scholar]
- Hobæk, A.; Larsson, P. Sex determination in Daphnia magna. Ecology 1990, 71, 2255–2268. [Google Scholar] [CrossRef]
- Hebert, P.D.N. Genetics of Daphnia. In Daphnia; Peters, R.H., De Bernardi, R., Eds.; Mem Inst Ital Idrobiol: Verbania Pallanza, Italia, 1987; Volume 45, pp. 439–460. [Google Scholar]
- De Meester, L. Local genetic differentiation and adaptation in freshwater zooplankton populations: Patterns and processes. Ecoscience 1996, 3, 385–399. [Google Scholar] [CrossRef]
- Vanoverbeke, J.; De Gelas, K.; De Meester, L. Habitat size and the genetic structure of a cyclical parthenogen, Daphnia magna. Heredity 2007, 98, 419–426. [Google Scholar] [CrossRef]
- Deng, H.W.; Lynch, M. Change of genetic architecture in response to sex. Genetics 1996, 143, 203–212. [Google Scholar] [CrossRef]
- Hairston, N.G.; Holtmeier, C.L.; Lampert, W.; Weider, L.J.; Post, D.M.; Fischer, J.M.; Cáceres, C.E.; Fox, J.A.; Gaedke, U. Natural selection for grazer resistance to toxic cyanobacteria: Evolution of phenotypic plasticity? Evolution 2001, 55, 2203–2214. [Google Scholar] [CrossRef]
- Lynch, M.; Deng, H.W. Genetic slippage in response to sex. Am. Nat. 1994, 144, 242–261. [Google Scholar] [CrossRef]
- Michels, E.; Audenaert, E.; Ortells, R.; De Meester, L. Population genetic structure of three pond-inhabiting Daphnia species on a regional scale (Flanders, Belgium). Freshw. Biol. 2003, 48, 1825–1839. [Google Scholar] [CrossRef]
- Spaak, P.; Denk, A.; Boersma, M.; Weider, L.J. Spatial and temporal patterns of sexual reproduction in a hybrid Daphnia species complex. J. Plankton Res. 2004, 26, 625–635. [Google Scholar] [CrossRef]
- Wolf, H.G. Interspecific hybridization between Daphnia hyalina, D. galeata and D. cucullata and seasonal abundances of these species and their hybrids. Hydrobiologia 1987, 145, 213–217. [Google Scholar] [CrossRef]
- Nybom, H. Comparison of different nuclear DNA markers for estimating intraspecific genetic diversity in plants. Mol. Ecol. 2004, 13, 1143–1155. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lakes | Coordinates X/Y | Elevation of Lakes above Sea Level, m | Surface Area with Island, km2 | Surface Area without Island, km2 | Max. Depth, m * | Mean Depth, m * | Catchment Basin, km2 | Shore Length, km |
|---|---|---|---|---|---|---|---|---|
| Dridzis | 705,390.852/ 208,462.077 | 159.8 | 7.72 | 7.56 | 64 | 12.8 | 46 | 42 |
| Riča | 670,715.594/ 175,721.067 | 145.8 | 13.12 | 13.07 | 39 | 9.7 | 123 */130 ** | 34 |
| Svente | 647,412.511/ 192,388.091 | 136.9 | 7.06 | 7.03 | 38 | 7.8 | 20 | 26 |
| Geraņimovas-Ilzas | 696,251.015/ 228,167.042 | 150.7 | 3.17 | 3.17 | 46 | 9.8 | 66 | 24 |
| Locus | Primer Sequences (5′−3′) | Repeat Unit | Label Dye | Size Range (bp) (Our Data) | Size Range (bp) (Data after Brede et al.) | T (°C) |
|---|---|---|---|---|---|---|
| SwiD1 | F:GCCGTGTTCGAAAGCTAGTC R: AGCCGAACGAAAAACATGC | (TG)18 | 5’TAM | 122–127 | 116–142 | 59.4 |
| Dgm105 | F:ATGTGAGCGCGCGAGCATTT R:GTCCAGCCGGCCCATTTCAGTT | (CAG)8AG | 5’FAM | 165–240 | 172–197 | 59.4 |
| Dgm101 | F: TCTTGCTCGAATTCTCTCC R: CCTGTCTCACACGGAGC | (GA)10AGA | 5’HEX | 165–180 | 162–177 | 54.5 |
| DaB17/17 | F:GAGAACCTTTTATCAGCTTCG R:ACTCATCTGGTGAGATGGATC | T9 | 5’TAM | 100–106 | 100–109 | 55.9 |
| Dgm109 | F: CCAGCTGTTGACCACCTG R: TGCGCGAGGATTTCCAACAC | (ACC)7AC | 5’FAM | 250–303 | 247–266 | 58.2 |
| Dp519 | F:AGTCGCGACGACATAAAGC R:GTGGTAGTTGTGGAATCCG | (TG)6(GA)7 | 5’HEX | 140–142 | 144–160 | 56.7 |
| DaB10/15 | F:AGAGAAGTGTTTGCGTTTC R:TGTTTCCTATATCCCTCGG | TC6 | 5’TAM | No result | 75–89 | 52.4 |
| Dp512 | F:TTTCGTTCTACCCAGGGAAG R:TTTGCTCGTCTGTGATAGGC | (TG)4…(GT)8 | 5’HEX | No result | 125–141 | 57.3 |
| DaB17/16 | F: AGGGAACGAGCGGCGATAAG R:TCTTTGGCAGGCCACTGCCAAGG | GA10 | 5’FAM | No result | 189–195 | 61.4 |
| Locus | Total Number of Alleles in the Locus | Number of Private Alleles in the Locus | Proportion of Private Alleles (%) | Number of Populations in Which Private Alleles Have Been Detected |
|---|---|---|---|---|
| SwiD1 | 5 | 1 | 20 | 1 |
| Dgm105 | 8 | 5 | 62 | 3 |
| Dgm101 | 7 | 4 | 57 | 3 |
| DaB17/17 | 4 | 2 | 50 | 2 |
| Dgm109 | 5 | 3 | 60 | 2 |
| Dp519 | 2 | 0 | 0 | 0 |
| Sample | SwiD1 | Dgm105 | Dgm101 | DaB17/17 | Dgm109 | Dp519 | |
|---|---|---|---|---|---|---|---|
| Dridzis | N | 4 | 4 | 1 | 14 | 4 | 14 |
| Na | 4 | 1 | 1 | 2 | 1 | 1 | |
| No | 0 | 0 | 1 | 0 | 0 | 0 | |
| Ho | 0 | 0 | 0 | 0 | 0 | 0 | |
| He | 0.75 | 0 | 0 | 0.13 | 0 | 0 | |
| Riča | N | 13 | 12 | 12 | 19 | 11 | 19 |
| Na | 2 | 4 | 5 | 2 | 3 | 2 | |
| No | 1 | 1 | 2 | 0 | 1 | 0 | |
| Ho | 0 | 0.25 | 0 | 0 | 0 | 0 | |
| He | 0.14 | 0.51 | 0.68 | 0.46 | 0.31 | 0.1 | |
| Svente | N | 4 | 8 | 0 | 15 | 0 | 16 |
| Na | 3 | 4 | 0 | 2 | 0 | 2 | |
| No | 0 | 2 | 0 | 1 | 0 | 0 | |
| Ho | 0 | 0.25 | 0 | 0 | 0 | 0 | |
| He | 0.62 | 0.33 | 0 | 0.12 | 0 | 0.37 | |
| Geranimovas-Ilzas | N | 7 | 6 | 7 | 14 | 6 | 8 |
| Na | 2 | 5 | 4 | 3 | 4 | 1 | |
| No | 0 | 2 | 1 | 1 | 2 | 0 | |
| Ho | 0 | 0.17 | 0 | 0 | 0.17 | 0 | |
| He | 0.24 | 0.74 | 0.73 | 0.36 | 0.68 | 0 |
| Population/Microsatellite Loci | SwiD1 | Dgm105 | Dgm101 | DaB17/17 | Dgm109 | Dp519 |
|---|---|---|---|---|---|---|
| Dridzis | ns | M | M | *** | M | M |
| Riča | *** | ns | *** | *** | *** | *** |
| Svente | * | * | M | *** | M | *** |
| Geraņimovas-Ilzas | ** | ns | ** | *** | ns | M |
| Population | Dridzis | Riča | Svente | Geraņimovas-Ilzas |
|---|---|---|---|---|
| Dridzis | 0.29 | 0.45 | 0.37 | |
| Riča | 0.56 | 0.50 | 0.08 | |
| Svente | 0.50 | 1.14 | 0.49 | |
| Geraņimovas-Ilzas | 0.70 | 0.16 | 1.35 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brakovska, A.; Škute, N. Exploring the Genetic Diversity and Population Structure of Daphnia cucullata Sars, 1862 in Boreal Lakes (Latvian Lakeland) Based on Microsatellites. Diversity 2023, 15, 1128. https://doi.org/10.3390/d15111128
Brakovska A, Škute N. Exploring the Genetic Diversity and Population Structure of Daphnia cucullata Sars, 1862 in Boreal Lakes (Latvian Lakeland) Based on Microsatellites. Diversity. 2023; 15(11):1128. https://doi.org/10.3390/d15111128
Chicago/Turabian StyleBrakovska, Aija, and Nataļja Škute. 2023. "Exploring the Genetic Diversity and Population Structure of Daphnia cucullata Sars, 1862 in Boreal Lakes (Latvian Lakeland) Based on Microsatellites" Diversity 15, no. 11: 1128. https://doi.org/10.3390/d15111128
APA StyleBrakovska, A., & Škute, N. (2023). Exploring the Genetic Diversity and Population Structure of Daphnia cucullata Sars, 1862 in Boreal Lakes (Latvian Lakeland) Based on Microsatellites. Diversity, 15(11), 1128. https://doi.org/10.3390/d15111128