Abstract
Thirty-one nominal species of the benthic harpacticoid family Zosimeidae Seifried, 2003 are known from several coastal systems less than 200 m depth and the deep sea. Several copepods of the family Zosimeidae were found in deep-sea sediment samples from the Gulf of Mexico and the Gulf of California (Mexico). The Mexican material represents new species of the genera Zosime Boeck, 1873, Peresime Dinet, 1974, Heterozosime Kim and Lee, 2021, and Acritozosime Kim and Lee, 2021. Standardized and complete descriptions for these new species are provided. Previous studies have shown that the genus Zosime can be subdivided into two species groups. We performed phylogenetic analyses (88 characters and 42 taxa) to test the naturalness of such subdivision, to subdivide these groupings further, and to shed some light on the phylogenetic relationships among the zosimeid genera and their species. Our results confirmed (i) the monophyly of previous subdivisions of the genus Zosime, and (ii) that the genus Zosime can be further subdivided into two major subgroups composed of several subordinate clades. Additionally, some comments on the phylogenetic hypotheses amongst the species of each genus reported here, are provided.
Keywords:
Crustacea; meiofauna; systematics; diversity; Zosimeidae; Gulf of California; Gulf of Mexico 1. Introduction
Zosimeidae Seifried, 2003 is a group of benthic harpacticoids with a wide geographical and eurybathic distribution. The family is currently composed of the genera Acritozosime Kim and Lee, 2021 (monotypic), Heterozosime Kim and Lee, 2021 (monotypic), Peresime Dinet, 1974 (three species), Pseudozosime Scott T., 1912 (monotypic), and Zosime Boeck, 1873 (25 species). The genera Acritozosime and Heterozosime were first described from the East Mariana Basin at depths ranging from 5078 to 5856 m, and from the Southern Sea (South Korea) at depths ranging from 78.7 to 116.3 m, respectively [1]. The genus Peresime is known from depths ranging from 1010 to 4100 m, and its species were described from the Gulf of Mexico [2], Walvis Ridge (south-western Africa) [3], and from the Iberian deep sea (north-eastern Atlantic) [4]. The monotypic genus Pseudozosime is known only from the South Orkney Islands (north-eastern Atlantic) [5]. Species of Zosime were first described from several coastal systems less than 200 m depth, and from the deep sea [2,6,7,8,9].
The harpacticoid fauna of the Mexican Pacific region has received some attention recently, and several shallow and deep-sea species have been described since 1998 (see [6,10] and references cited therein). On the other hand, despite the high diversity of harpacticoid copepods of the Gulf of Mexico (see [6] and references cited therein) only few species, viz., Smacigastes methanophilus Plum and Martínez Arbizu, 2009 and Pentacletopsyllus montagni Bang, Baguley and Moon, 2014, had been described from this region until 2021. More recently, Yeom et al. [2] described three species of the ubiquitous genus Zosime, and one species of Peresime from this critical area.
Some undescribed species of Zosime, Peresime, Heterozosime, and Acritozosime were found in sediment samples taken during several surveys in the Mexican Pacific and Gulf of Mexico. Zosime mexicana sp. nov., and Heterozosime klemie sp. nov. are the first species of the family Zosimeidae described from the Mexican Pacific. Zosime xiximi sp. nov., Pe. pryocta sp. nov., and Acritozosime sigsbeeiensis sp. nov. raised the number of zosimeid species from the Gulf of Mexico from four species distributed in two genera, to seven species distributed in three genera. Heterozosime klemie sp. nov. and A. sigsbeeiensis are the first records of these two genera from Mexico. Some comments on their relationships based upon the phylogenetic hypotheses herein presented, are given.
Seifried [11] diagnosed the family Zosimeidae upon several autapomorphies. However, recent evidence suggests that some amendments are necessary. Some comments on this are also provided.
Kim et al. [12] proposed subdividing the genus Zosime into three species groups (the anneae-, the typica-, and the incrassata-groups) in an attempt to clarify the relationships between the species of Zosime. The concept of Kim et al. [12] was later modified by Pointner [13] who incorporated Z. anneae (the only species of Kim et al.’s [12] anneae-group) into the typica-group. The monophyly of the groupings of Zosime is tested herein with maximum parsimony and Bayesian inference, and some comments on the hypothetical phylogenetic relationships amongst the species of the genus are provided.
2. Materials and Methods
2.1. Field and Laboratory Work
The new species presented here were obtained during cruises Xiximi-5 (Gulf of Mexico), and Talud IV and Talud XV (eastern Pacific Ocean). The sampling and sample processing methods followed during these surveys are detailed below.
Xiximi-5 cruise: Sediment samples were taken using a Soutar box corer at ten sampling stations distributed in the Gulf of Mexico, at depths ranging from 1242 to 3721 m. Two subsamples were taken at each station using acrylic cores (sampling surface = 78.5 cm2, length = 12 cm), and each core was subdivided horizontally into eight layers (0–1 cm, 1–2 cm, 2–3 cm, 3–4 cm, 4–6 cm, 6–8 cm, 8–10 cm, and 10–12 cm). The subsamples of one of the cores were fixed with a 4% formalin solution; the subsamples of the other core were preserved with DESS prepared following Yoder et al. [14]. All samples were frozen until further processing and only the surface 0–1-cm layer was considered for meiofauna analyses. Meio- and macrofauna in each 0–1-cm subsample were separated with 500 µm and 45 µ sieves. The material retained in the 45 µm sieve (meiofauna) was processed according to Somerfield and Warwick [15]. De Jonge and Bouwman [16] was followed for the separation of meiofauna using Ludox TM-50®.
Talud IV and Talud XV cruises: Sediment samples for meiofaunal analyses were taken in August 2000 at the southern Gulf of California from Carmen basin to off Nayarit State, and in July–August 2012 off the west coast of Baja California Sur, during Talud IV and Talud XV cruises, respectively, on board RV “El Puma” of the Universidad Nacional Autónoma de México (UNAM). Sediment samples were collected at depths ranging from 520 to 2120 m during the Talud IV cruise using a multiple sediment corer equipped with six cores (sampling surface = 3.9 cm2, length = 30 cm), and from 325 to 2000 m during the Talud XV cruise, using a box corer from which triplicate subsamples were taken with acrylic cores (sampling surface = 69 cm2, length = 20 cm). The upper 3 cm layer of sediment was preserved in 70% alcohol (Talud IV cruise) and in 96% alcohol (Talud XV cruise), and sieved through 500 and 38 μm sieves to separate macro- and meiofauna.
2.2. Taxonomic and Descriptive Work
The material under study was sorted using an Olympus SZX12 stereomicroscope equipped with DF PLAPO 1X objective and WHS10x eyepieces, and harpacticoid copepods were stored separately in 1 mL vials with 96% ethanol. Illustrations and figures were made from whole individuals and their dissected parts using a Leica DMLB microscope equipped with L PLAN 10X eyepieces, N PLAN 100X oil immersion objective, and a drawing tube. The dissected parts were mounted on separate slides using glycerin as mounting medium, and sealed with Neo-Mount®. Huys and Boxshall [17] and Seifried [11] were followed for general terminology.
Abbreviations used in the text and tables: A1, antennule; A2, antenna; ae, aesthetasc; apo, apophysis; BENP, baseoendopod; ENP, endopod; EXP, exopod; EXP (ENP)1 (2,3), first (second, third) exopodal (endopodal) segment; P1–P6, first to sixth legs.
The armature formulae of the amended diagnoses and of the new species presented here follow the format of Sewell [18]. Other armature formulae presented in other sections follow the format of Lang [19].
Primitive segments of the female antennule of Idyanthidimorpha sensu Seifried [11], Idyanthidae Lang, 1944 and Zosimeidae are in Roman numerals; actual segments are in Arabic numerals, and conform to the ground pattern as detailed by Seifried [11].
The type material was deposited in the Copepoda collection of the Instituto de Ciencias del Mar y Limnología, Unidad Académica Mazatlán (ICML-EMUCOP).
2.3. Phylogenetics
A phylogenetic analysis was performed with 88 characters (Table 1) and 42 taxa (Table S1). The taxa considered in this analysis were the genera Zosime (27 species), Peresime (four species), Pseudozosime (one species), Heterozosime (two species), and Acritozosime (two species) (Table S1). As outgroups, we used (i) the ground pattern of Idyanthidae and Zosimeidae as defined by Seifried [11]; (ii) Tachidiella minuta Sars G. O., 1909, T. kimi Lee and Huys, 1999, and T. reducta Lee and Huys, 1999; and (iii) Styracothorax gladiator Huys, 1993. Phylogenetic analyses were performed with two phylogenetic methods, the maximum parsimony and Bayesian inference. A Wagner parsimony method with unordered multistates, randomized input order species, and the weights option was carried out in Phylip ver. 3.697 [20]. A consensus tree was obtained from the majority rule and support values were derived from 10,000 bootstrap replicates. Bayesian inference was performed using MrBayes ver. 3.2.7a [21]. The default prior distribution of parameters was used for MCMCMC analyses, with one cold chain and three heated chains for 1,000,000 generations, and sampled every 100th. The resulting tree (Figure S1) showed Styracothorax nested within Heterozosime-Acritozosime. This could be due to the long branch attraction in the maximum parsimony by not considering other idyanthid taxa with two-segmented P1 exopods. Another edited tree is shown instead. The cladogram was visualized and edited using the Interactive Tree Of Life (iTOL, https://itol.embl.de, accessed on 22 January 2023) v6) [22].

Table 1.
List of characters and characters states used in the phylogenetic analysis.
Following Huys and Boxshall [17], oligomerization is regarded here as the main evolutionary trend in Copepoda.
3. Results
3.1. Systematics
Order Harpacticoida Sars G. O., 1903
Family Zosimeidae Seifried, 2003
3.1.1. Genus Zosime Boeck, 1873
Type species. Zosime typica Boeck, 1873, by original designation.
Other species. Zosime anneae Koller and George, 2011; Z. atlantica Bodin, 1968; Z. bathyalis Por, 1967; Z. bathybia Bodin, 1968; Z. bergensis Drzycimski, 1968; Z. carsteni Pointner, 2017; Z. changi Kim and Lee, 2019; Z. comata Kim and Lee, 2019; Z. destituta Kim, Jung and Yoon, 2016; Z. eliasi Pointner, 2017; Z. erythraea Por, 1967; Z. gisleni Lang, 1948; Z. gymnokosmosa Kim and Lee, 2019; Z. incrassata Sars G. O., 1910; Z. major Sars G. O., 1919; Z. mediterranea Lang, 1948; Z. mexicana sp. nov.; Z. montagnai Yeom, Rohal Lupher and Lee, 2022; Z. pacifica Fiers, 1991; Z. paramajor Bodin, 1968; Z. paratypica Becker and Schriever, 1979; Z. reyssi Dinet, 1974; Z. thistlei Yeom, Rohal Lupher and Lee, 2022; Z. tunnelli Yeom, Rohal Lupher and Lee, 2022; Z. valida Sars, G. O., 1919; Z. xiximi sp. nov.
- Zosime xiximi sp. nov.
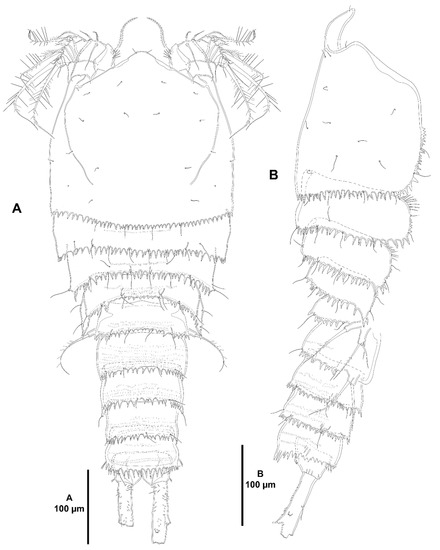
Figure 1.
Zosime xiximi sp. nov., intersexual holotype (ICML-EMUCOP-170616-01). (A), habitus, dorsal; (B), habitus, lateral.
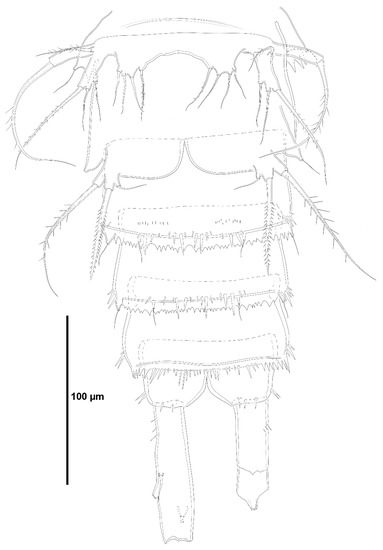
Figure 2.
Zosime xiximi sp. nov., intersexual holotype (ICML-EMUCOP-170616-01). Urosome, ventral, showing P5 and P6.
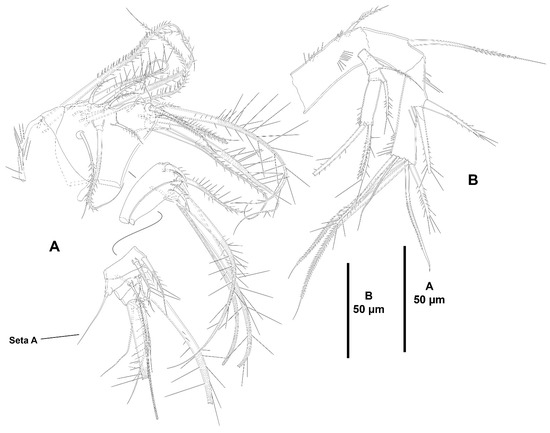
Figure 3.
Zosime xiximi sp. nov., intersexual holotype (ICML-EMUCOP-170616-01). (A), antennule, indicating position of seta A; (B), antenna.
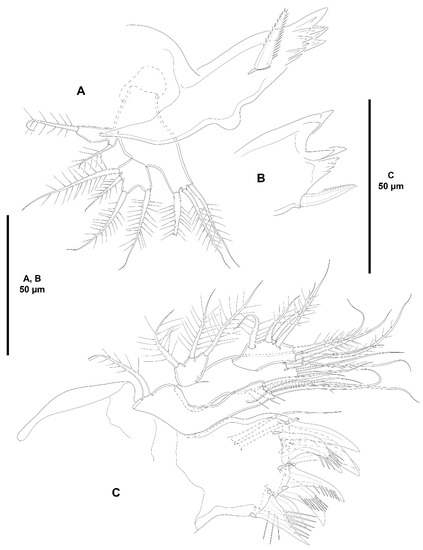
Figure 4.
Zosime xiximi sp. nov., intersexual holotype (ICML-EMUCOP-170616-01). (A), mandible; (B), distal part of mandibular gnathobase, another view; (C), maxillule.
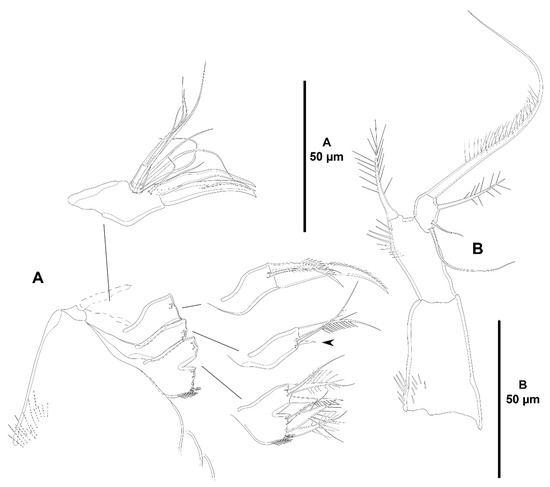
Figure 5.
Zosime xiximi sp. nov., intersexual holotype (ICML-EMUCOP-170616-01). (A), maxilla, indicating small element on first coxal endite; (B), maxilliped.
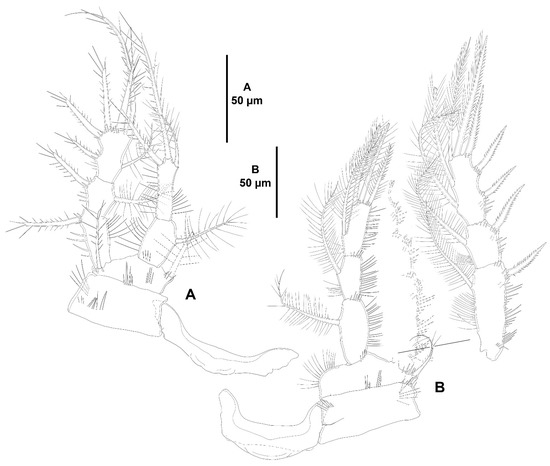
Figure 6.
Zosime xiximi sp. nov., intersexual holotype (ICML-EMUCOP-170616-01). (A), P1, anterior; (B), P2, anterior.
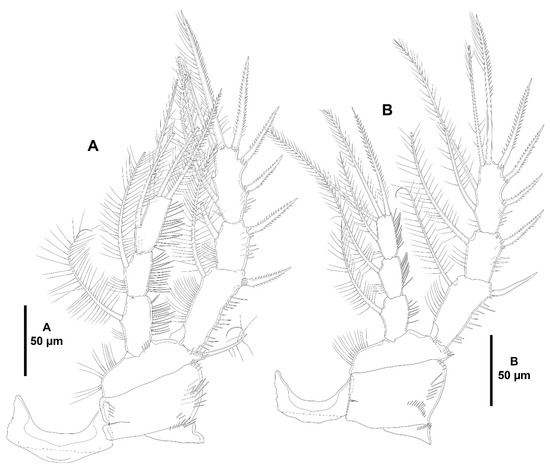
Figure 7.
Zosime xiximi sp. nov., intersexual holotype (ICML-EMUCOP-170616-01). (A), P3, anterior; (B), P4, anterior.
ZooBank Registration LSID
urn:lsid:zoobank.org:act:AD73021C-2217-4A9C-88AE-3764F652E575
Type locality. Xiximi-5 cruise; stn. A5, Gulf of Mexico, northern Sigsbee Deep, 25.1168° N, 92.0002° W; 3513 m depth; clay, 16%, silt, 66%, sand, 18%; 17 June 2016.
Material examined. Female holotype displaying female and male (P6) characteristics (ICML-EMUCOP-170616-01) dissected and mounted onto eight slides. Ivonne Martínez-Mendoza leg.
Etymology. The specific epithet refers the oceanographic campaigns Xiximi of the Gulf of Mexico Research Consortium (CIGoM). Xiximi also means “to spill” in the ancient Nahuatl or Aztec language, an American Indian language of the Uto-Aztecan family which is spoken in central and western Mexico. The specific epithet xiximi is to be treated as indeclinable and need not agree in gender with the generic name Zosime ([23] Art 31.2.3).
Description of female. Total body length, 659 µm measured from anterior margin of rostrum to posterior margin of caudal rami. Habitus (Figure 1A,B) subcylindrical, tapering posteriad, with constriction between pro- and urosome.
Prosome (Figure 1A,B) consisting of cephalothorax and free P2–P4-bearing somites. Rostrum fused to cephalothorax, well developed, broad, triangular, with pair of subdistal sensilla. Cephalothorax large; posterolateral margin coarsely serrate, anterolateral margin plain; with sensilla as shown. P2–P4-bearing somites with pattern of minute spinules as shown; posterior serrate margin of P2- and P3-bearing somites coarser than in cephalothorax, of P4-bearing somite visibly less coarse than in two previous somites; with dorsal and posterior sensilla as illustrated.
Urosome (Figure 1A,B and Figure 2) consisting of P5-bearing somite, genital somite with P6, three free somites, and anal somite with caudal rami. P5-bearing somite with minute spinules dorsally; posterolateral margin serrate as in previous segment; with posterior sensilla as shown; ventrally without spinular ornamentation. Genital somite (second urosomite) and third urosomite separated dorsally and ventrally. Genital somite covered with minute spinules dorsally, without ventral spinular ornamentation; posterodorsal serrate margin coarser than in previous somite; both P6 fused to somite, each a plate with three setae of which innermost shorter, medial spiniform, outer setiform. Third urosomite with posterior margin coarsely serrate dorsally and laterally, ventrally less coarsely serrate; dorsolateral surface covered with minute spinules, with few lateral stronger spinules; ventrally with few surface spinules, and with stronger and longer medial posterior spinules as shown; with few sensilla. Fourth urosomite largely as previous somite, but without surface minute spinules ventrally. Fifth urosomite with posterodorsal margin coarsely serrate, coarser medially forming a pseudoperculum; dorsolateral surface covered with minute spinules, with few lateral stronger spinules; ventrally without surface spinules, posterior margin non-serrate but with strong, long spinules; without sensilla. Anal somite short; operculum covered by pseudoperculum of previous somite not visible; with paired sensilla associated to anal operculum on each side; with few spinules laterally and ventrally as shown. Caudal rami damaged, as long as two preceding somites combined, parallel, cylindrical; most caudal setae broken off; lateral outer seta I short, arising midway outer margin; seta II posterior to seta I; dorsal seta VII issuing from long pedestal.
Antennule (Figure 3A) eight-segmented; short and robust. All segments smooth, except for first segment with short spinular row; with aesthetasc on third and eighth segments. With inner biarticulate seta issuing from long expansion of sixth segment (seta A in Figure 3A). Relative length of segments as shown. Armature formula: 1-[1], 2-[16], 3-[2(1+ae)], 4-[2], 5-[3], 6-[2], 7-[1], 8-[5+(1+ae)].
Antenna (Figure 3B) with basis ornamented with outer and medial spinules as shown, with abexopodal seta half as long as segment. Endopod two-segmented; first segment shorter than second, unornamented, with one long, pinnate, inner subdistal seta; second segment with spinules proximally, medially and subdistally, laterally with two outer spines and a small slender seta, and with six distal elements as shown. Exopod large, three-segmented; first segment with one seta; second segment shortest, with one seta; third segment longest, with three distal setae.
Mandible (Figure 4A,B) with cuspid teeth and one dorsal spiniform element on gnathobase. Palp three-segmented composed of basis, exopod and endopod. Basis elongate, with three setae. Exopod one-segmented, elongate, with two setae as shown. Endopod one-segmented, short, with four setae.
Maxillule (Figure 4C) with well-developed praecoxal arthrite armed with two surface setae, and distally with eight spines as shown. Coxa with epipodal seta; endite with four setae. Basis with some spinules, and five setae. Exopod one-segmented, small, with three setae. Endopod elongate, longer than exopod, with four lateral and two distal setae.
Maxilla (Figure 5A) with large syncoxa ornamented with few outer spinules; with two praecoxal endites fused basally, each with three elements, and two elongate coxal endites with three setae each (one seta of the first coxal endite very small, indicated in Figure 5A). Allobasis drawn out into strong claw accompanied by one unipinnate spine and three setae one of which close to insertion site of endopod. Endopod two-segmented; first segment with two, second segment with one proximal and three distal setae.
Maxilliped (Figure 5B) three-segmented. Syncoxa with proximal spinules as shown, unarmed. Basis with outer row of spinules and armed with one distal element. Endopod small, one-segmented; with two lateral, slender setae of which subdistal shorter, and two distal elements of different lengths.
P1 (Figure 6A) with bare intercoxal sclerite transversely prolonged as shown. Coxa rectangular, with some anterior and posterior outer spinules, and some medial distal ones. Basis with inner and outer spine; with spinules at the base of inner pinnate spine and close to insertion site of endopod. Exopod three-segmented, longer than endopod; all segments with spinules as shown; no pores detected; first segment longest, slightly longer than third, with outer spine, without inner armature; second segment shortest, with outer spine and inner seta; third segment with three outer spines, two distal elements of which outer spiniform and shorter, and one inner seta. Endopod two-segmented, reaching middle of EXP3; both segments with long spinules as shown; no pores detected; proximal segment shorter, with one inner seta; second segment with one inner, one distal, and one outer element as depicted.
P2–P4 (Figure 6B and Figure 7A,B) with bare, transversely prolonged intercoxal sclerites. Praecoxa of P2 lost during dissection, of P3 and P4 small, triangular, with transverse row of subdistal spinules. Coxa with spinules as shown. Basis with spinules at the base of outer seta and between rami, and with longer slender spinules along inner margin in P2 and P3, of P4 as in P2 and P3 but without spinules between rami. Exopod three-segmented, EXP1 longest, EXP2 shortest; exopodal segments ornamented with spinules as shown; no pores detected; EXP1 and EXP2 with outer spine and inner seta; EXP3 with three outer spines, two distal elements, and two inner setae. Endopod three-segmented, of P2 and P3 reaching insertion site of inner seta, of EXP4 barely beyond EXP2; each segment ornamented with spinules as shown; no pores detected; ENP1 and ENP2 with inner seta; ENP3 with one inner and three distal elements of which outer spiniform.
Armature formulae of P1–P4 as in Table 2.
Table 2.
Armature formulae of legs 1–4 of Zosime xiximi sp. nov.
Both P5 (Figure 2) fused; baseoendopod and exopod fused. Baseoendopod with outer basal seta arising from long setophore, and four elements on endopodal lobe of which outermost shortest. Exopod elongate; with three setae on segment as shown plus one surface seta arising at the base of ramus.
Description of male. Male unknown.
- Zosime mexicana sp. nov.
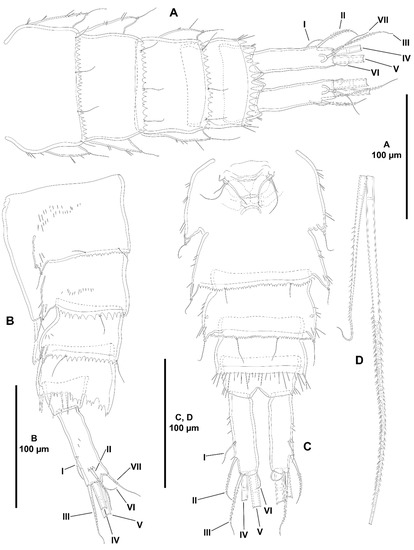
Figure 8.
Zosime mexicana sp. nov., female holotype (ICML-EMUCOP-230800-02). (A), urosome, dorsal, P5-bearing somite omitted; (B), urosome, lateral, P5-bearing somite omitted; (C), urosome, ventral, P5-bearing somite omitted; (D), caudal setae IV and V. Roman numerals indicate the different caudal setae.
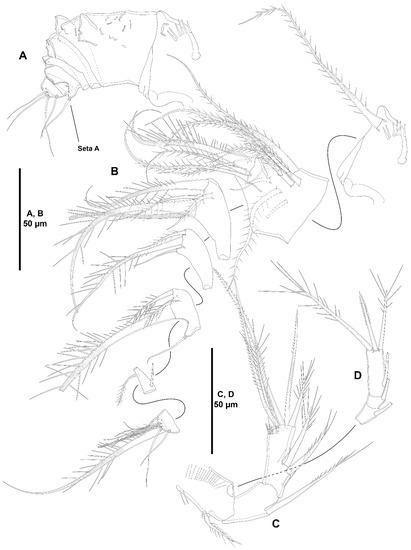
Figure 9.
Zosime mexicana sp. nov., female holotype (ICML-EMUCOP-230800-02). (A), antennule, armature complements omitted for clarity, seta A indicated; (B), antennule showing armature complements; (C), antenna, showing basis and endopod; (D), exopod of antenna.
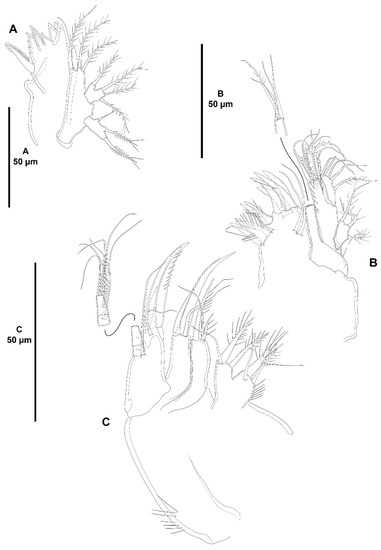
Figure 10.
Zosime mexicana sp. nov., female holotype (ICML-EMUCOP-230800-02). (A), mandible; (B), maxillule; (C), maxilla.
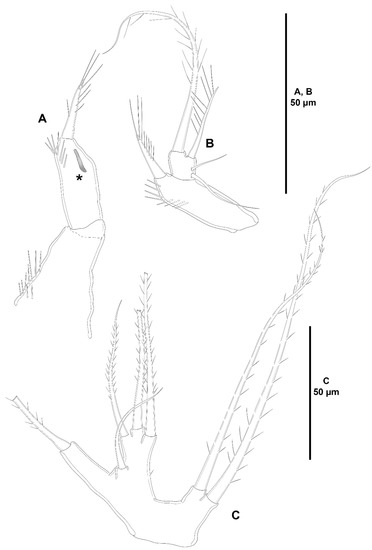
Figure 11.
Zosime mexicana sp. nov., female holotype (ICML-EMUCOP-230800-02). (A), maxilliped, asterisk indicates position of endopod; (B), maxilliped, another; (C), P5, anterior.
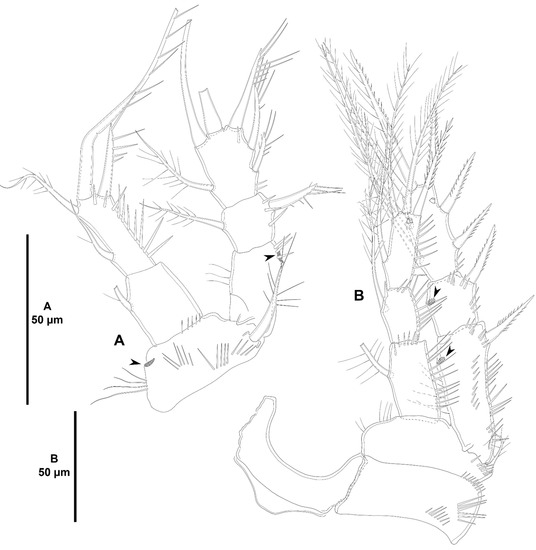
Figure 12.
Zosime mexicana sp. nov., female holotype (ICML-EMUCOP-230800-02). (A), P1, anterior, indicating the position of the inner basal element and outer spine of EXP1; (B), P2, anterior, indicating position of the inner setae on EXP1 and EXP2.
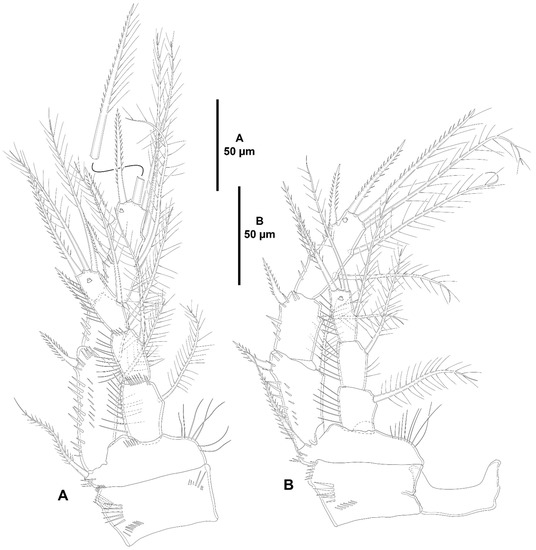
Figure 13.
Zosime mexicana sp. nov., female holotype (ICML-EMUCOP-230800-02). (A), P3, anterior; (B), P4, anterior.
ZooBank Registration LSID
urn:lsid:zoobank.org:act:95CAD21E-E1F9-4898-AC46-7BD224367810
Type locality. Talud IV cruise; stn. 4, eastern Pacific Ocean, off state of Nayarit, Mexico, 22.001389° N, 106.667222° W; 1228 m depth; organic matter, 4.06%, muddy sediment; 23 August 2000.
Other localities. Talud IV cruise; stn. 19, southern Gulf of California, off state of Sinaloa, Mexico, 24.266667° N, 108.401944° W; 1240 m depth; organic matter, 4.05%, muddy sediment; 25 August 2000.
Material examined. Female holotype from the type locality (stn. 4) (ICML-EMUCOP-230800-02) dissected and mounted onto seven slides; female paratype from stn. 19 (ICML-EMUCOP-250800-03) dissected and mounted onto seven slides. Samuel Gómez leg.
Etymology. The specific epithet comes from the Latin mexicāna, which is the inflection of the Latin mexicānus, from or pertaining to Mexico, and refers to the country where the new species was found. It is in the nominative singular, gender feminine.
Description of female. Length of urosome (P5-bearing somite omitted), 248 µm measured from anterior margin of genital double-somite to posterior margin of caudal rami. Urosome (P5-bearing somite omitted) subcylindrical, tapering posteriad (Figure 8A–C).
Urosome (Figure 8A–C) consisting of P5-bearing somite (omitted in figures), genital somite with P6, three free somites, and anal somite with caudal rami. Genital somite (second urosomite) and third urosomite separated dorsolaterally, completely fused ventrally forming genital double-somite. Genital somite (anterior half of genital double-somite) without spinules dorsally and ventrally, with spinules laterally, posterodorsal margin coarsely serrate, with posterior sensilla as shown, with paired ventrolateral sensillum-bearing pleural extensions ornamented with some spinules; P6 and genital field in the middle of somite; P6 with three setae of which outermost thickest, medial shortest. Third urosomite (posterior half of genital double-somite) with spinular ornamentation as in previous somite; posterodorsal margin coarsely serrate, posteroventral serrate margin less coarse, with sensilla as shown; with paired ventrolateral sensillum-bearing pleural extensions ornamented with some spinules. Fourth urosomite largely as posterior half of genital double-somite. Fifth urosomite without surface spinules; with lateroventral row of spinules along its posterior margin as shown; posterior dorsolateral margin coarsely serrate, with medial teeth comparatively longer forming pseudoperculum; without ventrolateral pleural extensions. Anal somite short, anal operculum and surface ornamentation (spinules and sensilla) covered by pseudoperculum of fifth somite, not visible, with few lateral spinules close to insertion site of caudal rami. The latter cylindrical, slightly convergent, about 3.5 times as long as wide; with seven setae as follows: lateral outer seta I short, arising at outer margin of distal third, slightly displaced ventrally; lateral seta II posterior to seta I; seta III issuing distally, slightly displaced ventrally, longer than seta II; seta IV and V pinnate, posterior half rat-tail like; seta VI short, arising at inner distal corner; dorsal seta VII issuing from pedestal, close to posterior margin, biarticulated as shown.
Antennule (Figure 9A,B) seven-segmented, short and robust; second segment longest. All segments smooth, except for first segment with spinules as shown; with aesthetasc on third and last segments. With inner biarticulate seta issuing from long expansion of sixth segment (seta A in Figure 9A). Relative length of segments as shown. Armature formula: 1-[1], 2-[14], 3-[3+(1+ae)], 4-[3], 5-[4], 6-[2], 7-[6+(1+ae)].
Antenna (Figure 9C,D) with basis ornamented with spinules as shown, with short abexopodal seta. Endopod two-segmented; first segment shorter, unornamented, with one long, pinnate, medial inner seta; second segment with inner spinules proximally, medially and subdistally, laterally with two strong outer spines and a small slender seta, distally with six elements as depicted. Exopod large, three-segmented; first segment with one seta; second segment shortest, with one seta; third segment longest, with one lateral and three distal elements of different lengths as shown.
Mandible (Figure 10A) with cuspid teeth and one dorsal, thick, pinnate seta on gnathobase. Palp three-segmented composed of basis, exopod and endopod. Basis elongate, with three setae. Exopod one-segmented, with three setae. Endopod one-segmented, with four setae.
Maxillule (Figure 10B) with well-developed praecoxal arthrite armed with two surface setae, and distally with eight strong spines and one setiform element as shown. Coxa with one epipodal seta; endite with four elements. Basis with some longitudinal spinules as shown; with two endites of which dorsal with five, ventral with two setae. Exopod one-segmented, with three setae. Endopod one-segmented, with six setae.
Maxilla (Figure 10C) with massive syncoxa ornamented with proximal outer spinules, with four endites as follows: two praecoxal endites fused basally of which proximal endite with spinular row, each endite with three elements; and two elongate coxal endites with three setae each. Allobasis drawn out into strong claw accompanied by one unipinnate spine and three setae one of which close to insertion site of endopod. Endopod two-segmented; first segment with two, second segment with one proximal and three distal setae.
Maxilliped (Figure 11A,B) three-segmented. Syncoxa with proximal outer spinules, unarmed. Basis with outer spinules subdistally, with one subdistal spinulose element. Endopod small, one-segmented, with two lateral, slender setae of which subdistal shorter and proximal very long, and two long distal elements of which inner shorter (the endopod of one maxilliped lost during dissection, but its insertion site indicated with an asterisk in Figure 11A).
P1 (Figure 12A) in bad condition and covered with detritus making spinular ornamentation difficult to see. Basis with spinules medially and at base of endopod, and with longer and slender inner spinules as shown; with inner and outer spines (inner spine lost during dissection, but its insertion site indicated with an arrowhead in Figure 12A). Exopod three-segmented, slightly longer than endopod, but reaching tip of ENP2 due to a lower insertion site on basis, all segments ornamented with spinules as shown; no pores detected; EXP1 as long as EXP3, with outer spine (the latter lost during dissection, but its insertion site indicated with an arrowhead in Figure 12A), without inner armature; EXP2 shortest, with outer spine and inner seta; EXP3 with three outer spines, two apical and one inner seta. Endopod two-segmented, distal segment longer than proximal; both segments with spinules as depicted; no pores detected; ENP1 with inner subdistal seta; second segment with two inner setae, and two distal elements of which outer spiniform.
P2–P4 (Figure 12B and Figure 13A,B) covered with detritus making spinular ornamentation difficult to see. Intercoxal sclerites unornamented (of P3 not shown). Coxa with proximal, medial, and distal outer spinules anteriorly; of P3 with, of P2 and P4 seemingly without anterior inner subdistal spinules, and posterior outer spinules. Basis of P2 with outer seta shorter than that of P3 and P4; basis of P2–P4 with spinules at the base of outer seta, and with longer and slender spinules along inner margin; of P2 and P4 seemingly without, of P3 with spinules between rami. Exopod three-segmented, longer than endopod; EXP1 longest, EXP2 shortest; exopodal segments ornamented with spinules as shown; with subdistal pore on P2–P4 EXP3; EXP1 with outer spine and inner seta (inner seta of P2 lost during dissection, but its insertion site indicated with an arrowhead in Figure 13B); EXP2 armed as in preceding segment (inner seta of P2 EXP2 lost during dissection, but its insertion site indicated with an arrowhead in Figure 13B); EXP3 with three outer spines, two distal elements of which outer spiniform, and two inner setae. Endopod three-segmented; of P2 reaching distal fourth of EXP3, of P3 reaching middle of EXP3, of P4 barely beyond distal margin of EXP2; all segments with spinules as depicted; with subdistal pore on P2–P4 ENP3; first segment longest (P2), as long (P3) or shorter (P4) than second segment; first and second segment with inner seta; P2 ENP3 and P3 ENP3 with three distal setae of which outer spiniform, P4 ENP3 with one inner seta, and two well-developed apical elements, between which lies a very reduced slender seta (tube-pore?).
Both P5 (Figure 11C) separated; baseoendopod and exopod fused but separated by deep notch. Baseoendopod with outer basal seta arising from long setophore, with two elements on endopodal lobe. Exopod with three distal setae plus a surface seta between base of exopodal lobe and setophore of basal seta.
Armature formulae of P1–P4 as in Table 3.
Table 3.
Armature formulae of legs 1–4 of Zosime mexicana sp. nov.
Description of male. Male unknown.
Remarks. Briefly, Z. xiximi sp. nov., attributed here to the typica-group, subordinate subgroup Ab (see below), is more closely related to Z. atlantica and Z. mediterranea. All these species conform to subordinate subgroup Ab. These three species share the armature formula of A2 EXP (1;1;111; without inner lateral seta and with three distal elements in EXP3). The loss of the lateral seta also occurred independently in the incrassata-group (Z. incrassata and Z. bathybia).
The males of Z. atlantica and Z. mediterranea remain unknown and nothing can be said regarding the male sexual dimorphism observed in the P6 of the intersexual specimen presented herein.
Following Bodin [24], Z. atlantica and Z. mediterranea differ in the shape of the pseudoperculum formed by the extension of the posterodorsal serrate margin of the fifth urosomite, being visibly coarser in Z. atlantica [24] (p. 37, plate XIX), and much less coarse in Z. mediterranea (Z. major in Monard [25] (p. 21, Figure 1e)). In this regard, Z. xiximi sp. nov. seems to be more closely related to Z. atlantica than to Z. mediterranea.
The mouth parts of Z. mediterranea have not been described. The antennary exopod and the maxilliped of Z. xiximi sp. nov. and Z. atlantica are similar, but these two species can be separated by the armature complement of the mandibular exopod (with four setae in Z. atlantica but reduced to two setae in Z. xiximi sp. nov.). The tetrasetose condition of the mandibular exopod of Z. atlantica is the most primitive within the genus and is present also in Pe. reducta, Pe. pryocta sp. nov., and Acritozosime (see below). The bisetose mandibular exopod of Z. xiximi sp. nov. is autapomorphic for the species within the genus and is the most derived condition within the typica-group (see below). The reduction in armature complement of the mandibular palp occurs also in the incrassata-group, i.e., with one seta only in the one-segmented mandibular exopod of Z. carsteni and represented by one seta in Z. incrassata and Z. bathybia. The one-segmented mandibular exopod with one seta is autapomorphic for Z. carsteni (see below). The exopod of Z. incrassata and Z. bathybia has been absorbed into the basis and is represented by one seta only. The mandibular exopod absorbed into the basis and represented by one seta is a synapomorphy for Z. incrassata and Z. bathybia (see below). The reduced one-segmented mandibular palp of Z. reyssi with one seta only (its basal, exopodal or endopodal origin is uncertain) is the most extreme reduction within the family and is autapomorphic for the species (see below).
In their study, Yeom et al. [2] argued that Z. montagnai from the southern Gulf of Mexico is morphologically similar to Z. paratypica from the Iberian deep sea in the north-eastern Atlantic. It should be highlighted that these researchers based their conclusions on the identification of their material using some identification keys. In our opinion, Z. mexicana sp. nov., attributed to the typica-group, subordinate subgroup Aa (see below), is more closely related to Z. montagnai and Z. pacifica instead. These species share the reduced medial seta of P4 ENP3 from a well-developed element to a small slender seta. The latter is regarded here as synapomorphic for Z. mexicana sp. nov., Z. montagnai, and Z. pacifica. A polytomy can be observed in the clade composed of Z. montagni, Z. paratypica, and Z. mexicana sp. nov.. Such polytomy is the result of missing and uncertain values for Z. paratypica (see Table S1). The—apomorphic—reduction in the medial seta of P4 ENP3 seems to have occurred independently in H. tenuis. The most derived reduction in the medial seta of P4 ENP3 within Zosime occurred independently in Z. tunnelli and Z. gymnokosmosa. The latter two species underwent complete loss of that seta. Similar apomorphic reductions also occurred independently in P4 ENP2 of Acritozosime and Pseudozosime. Peresime is unique within the family in the—autapomorphic—extreme reduction to one distal seta on the distal segment of the two-segmented endopod of P4.
Zosime mexicana sp. nov., Z. montagnai and Z. pacifica also share the bisetose endopodal lobe of the female P5. The bisetose endopodal lobe of the female P5 is also present in Z. valida, Z. destituta, and Z. changi (subordinate subgroup Ba), Table S1). Heterozosime tenuis also possesses a bisetose endopodal lobe of the female P5, but this reduction seems to have occurred independently, and is the most derived state—a potential apomorphy—within Heterozosime. Zosime montagnai and Z. pacifica also share the derived spiniform outer basal element of the male P6. The male of Z. mexicana sp. nov. remains unknown, but the spiniform outer basal element of the male P6 is regarded here as a potential synapomorphy for these three species (the outer basal element of the male P6 is also spiniform in H. tenuis and seems to have evolved independently, thus being a potential apomorphy for that species). The true relationship between Z. mexicana sp. nov., Z. montagnai, and Z. pacifica is uncertain and appears as a polytomy in our results. Zosime mexicana sp. nov. and Z. montagnai share (i) the segmentation and armature formula of the female antennule (seven-segmented in Z. mexicana sp. nov. and Z. montagnai but six-segmented in Z. pacifica; also note that Yeom et al. [2] gave the armature formula of the last segment as 5+(2+ae), but the aesthetasc of the last antennulary segment of the females of Zosime is commonly fused to a single seta, and Yeom et al.’s [2] armature formula of the seventh antennulary segment could be corrected to 6+(1+ae)); (ii) fusion of primitive female antennulary segments II and III resulting in the proximal aesthetasc on the third segment (without fusion of the proximal four segments in Z. pacifica, resulting in the position of the proximal aesthetasc on the fourth antennulary segment; see Table S2); and (iii) the number of epipodal setae on the maxillulary coxa (one seta in Z. mexicana sp. nov. and Z. montagnai, but two setae in Z. pacifica). On the other hand, Z. mexicana sp. nov. and Z. pacifica share (i) the armature complement of A2 EXP3 (with one lateral and three distal setae in Z. mexicana sp. nov. and Z. pacifica, but with one lateral and two distal setae in Z. montagnai), and (ii) the lateral armature of A2 ENP2 (with two spines and one small slender seta in Z. mexicana sp. nov. and Z. pacifica, but with two spines only in Z. montagnai). Zosime mexicana sp. nov. can be separated from the other two species by the number of segments of the maxillary endopod (two-segmented in Z. mexicana sp. nov., but one-segmented in Z. montagnai and Z. pacifica). The male of Z. mexicana sp. nov. remains undescribed and nothing can be said about the sexually dimorphic traits. Briefly, Z. montagnai and Z. pacifica differ in the male P5 (fused to the somite in Z. montagnai but separated in Z. pacifica; baseoendopods fused medially in Z. montagnai but separated in Z. pacifica; EXP and BENP fused in Z. montagnai but separated in Z. pacifica).
3.1.2. Genus Peresime Dinet, 1974
Type species. Peresime abyssalis Dinet, 1974, by original designation.
Other species. Peresime pryorae Yeom, Rohal Lupher and Lee, 2022; Pe. pryocta sp. nov.; Pe. reducta Becker and Schriever, 1979.
- Peresime pryocta sp. nov.
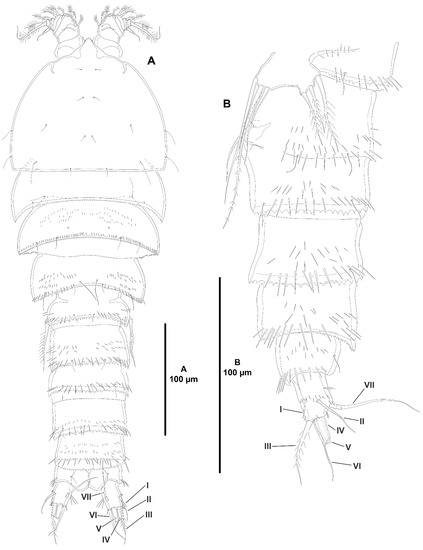
Figure 14.
Peresime pryocta sp. nov., female holotype (ICML-EMUCOP-061716-01). (A), habitus, dorsal; (B), urosome, lateral. Roman numerals indicate the different caudal setae.
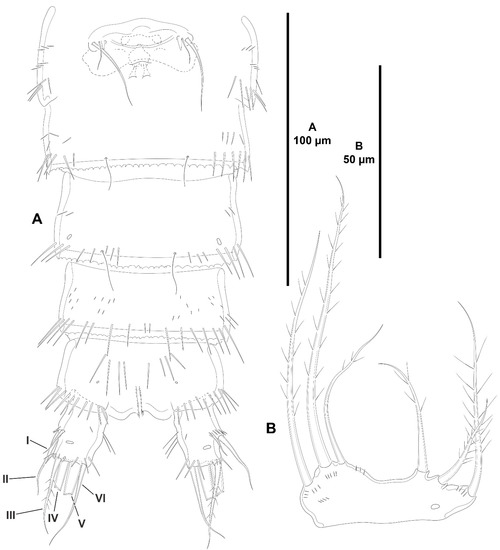
Figure 15.
Peresime pryocta sp. nov., female holotype (ICML-EMUCOP-061716-01). (A), urosome, ventral, P5-bearing somite omitted; (B), P5, anterior. Roman numerals indicate the different caudal setae.
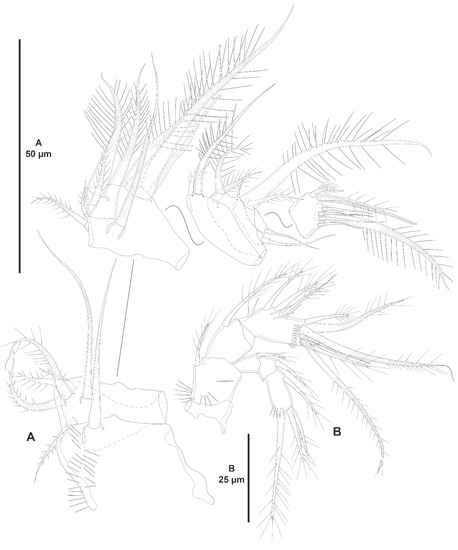
Figure 16.
Peresime pryocta sp. nov., female holotype (ICML-EMUCOP-061716-01). (A), antennule; (B), antenna.
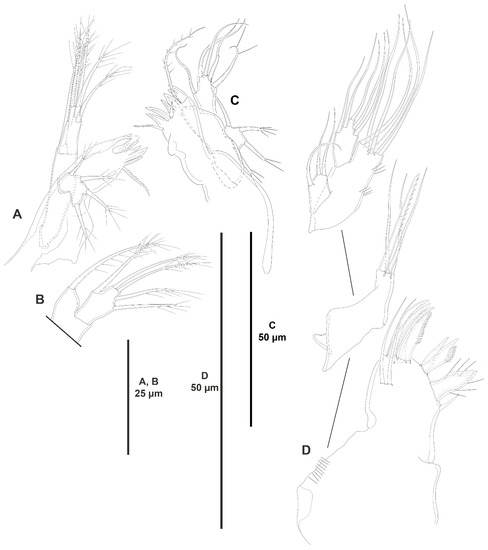
Figure 17.
Peresime pryocta sp. nov., female holotype (ICML-EMUCOP-061716-01): (A,B,D); female paratype (ICML-EMUCOP-061816-01): (C). (A), mandible; (B), distal part of mandibular palp; (C), mandible, another; (D), maxillule.
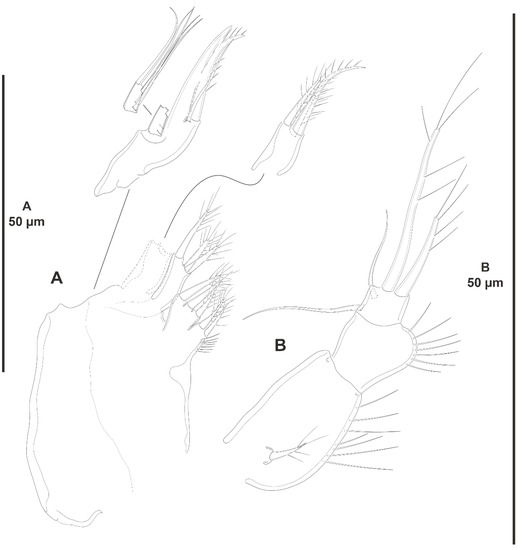
Figure 18.
Peresime pryocta sp. nov., female holotype (ICML-EMUCOP-061716-01). (A), maxilla; (B), maxilliped.
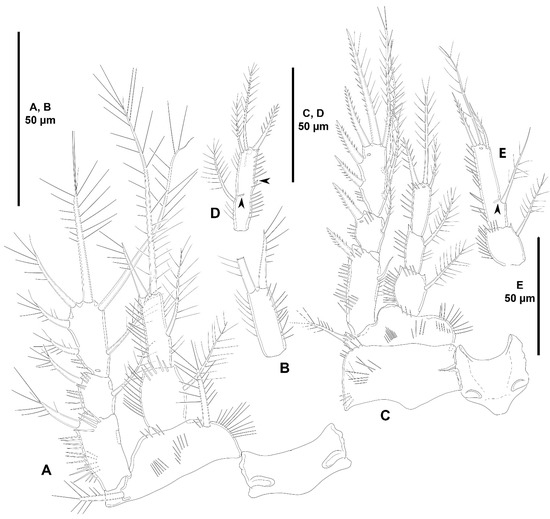
Figure 19.
Peresime pryocta sp. nov., female holotype (ICML-EMUCOP-061716-01): (A–D); female paratype (ICML-EMUCOP-061816-01): (E). (A), P1, anterior; (B), aberrant P1 ENP2 without inner seta; (C), P2, anterior; (D), distal segment of aberrant P2 ENP with ENP2 and ENP3 fused, remains of former division indicated; (E), aberrant P2 ENP with ENP2 and ENP3 fused, remains of former division indicated.
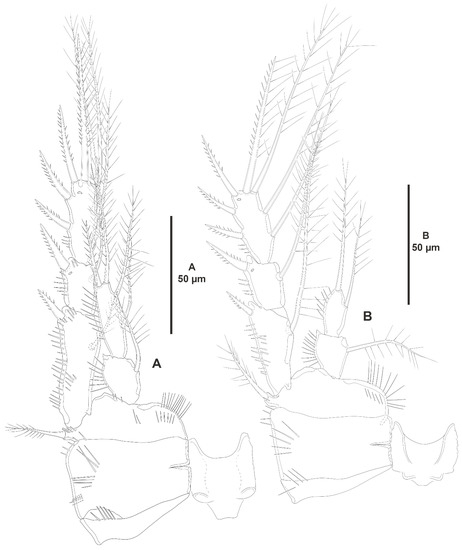
Figure 20.
Peresime pryocta sp. nov., female holotype (ICML-EMUCOP-061716-01). (A), P3, anterior; (B), P4, anterior.
ZooBank Registration LSID
urn:lsid:zoobank.org:act:7BB8B35C-03EF-447C-9E0E-7BC0145D464D
Type locality. Xiximi-5 cruise; stn. A5, Gulf of Mexico, northern Sigsbee Deep, 25.1168° N, 92.0002° W; 3513 m depth; clay, 16%, silt, 66%, sand, 18%; 17 June 2016.
Other localities. Xiximi-5 cruise; stn. A8, Gulf of Mexico, eastern Sigsbee Deep, 25.1168° N, 89.0500° W; 3477 m depth; clay, 55.5%, silt, 23.0%, sand, 21.5%; 18 June 2016.
Material examined. Female holotype (ICML-EMUCOP-061716-01) from the type locality dissected and mounted onto six slides; female paratype (ICML-EMUCOP-061816-01) from stn. A8 (see above) dissected and mounted onto seven slides. Ivonne Martínez-Mendoza leg.
Etymology. The specific name is an arbitrary combination of the specific epithets “pryorae” and “reducta” and refers to the close resemblance of the new species and Pe. pryorae and Pe. reducta. Gender feminine.
Description of female. Total body length, 417 µm measured from anterior margin of rostrum to posterior margin of caudal rami. Habitus (Figure 14A) subcylindrical, tapering posteriad, with constriction between pro- and urosome.
Prosome (Figure 14A) consisting of cephalothorax and free P2–P4-bearing somites. Rostrum fused to cephalothorax, well developed, triangular, with pair of subdistal sensilla. Cephalothorax large, with posterior margin minutely serrate; with sensilla as shown. Posterior serrate margin of P2–P4-bearing somites slightly coarser than in cephalothorax; P2-bearing somite seemingly without spinular ornamentation, P3–P4-bearing somites with dorsal spinular ornamentation and row of spinules along posterior margin (of P4-bearing somite coarser); with long and short sensilla along posterior margin as shown.
Urosome (Figure 14A,B and Figure 15A) consisting of P5-bearing somite, genital double-somite, two free somites, and anal somite with caudal rami. P5-bearing somite with transverse posterior spinular row and sensilla, and with some lateral spinules as shown; posterior serrate margin visibly coarser than in preceding somites. Genital somite (second urosomite) and third urosomite separated dorsally, fused ventrally forming genital double-somite. Genital somite (anterior half of genital double-somite) with dorsolateral posterior row of long spinules interrupted medially; with posterior margin coarsely serrate dorsally, plain laterally; ventrally without spinules; P6 and genital field in the middle of somite; P6 with a small, pointed projection and two setae of which inner very short. Third urosomite (posterior half of genital double-somite) without dorsal spinules, otherwise ornamented largely as previous somite; with posterior margin coarsely serrate dorsally and laterally, less coarse ventrally; with sensilla as shown. Fourth urosomite largely as previous somite, but with additional dorsal transverse spinular row close to posterior margin. Fifth urosomite largely as previous somite, but with more dorsal and ventral small spinules, and with ventral posterior spinules significantly longer; posterior serrate margin less coarse than in previous somite and extended dorsally forming a pseudoperculum; without sensilla. Anal somite short, with anal operculum covered with detritus and difficult to see, seemingly small and triangular, flanked on each side by a sensillum; dorsally without spinular ornamentation, with lateroventral long spinules posteriorly; ventrally with two sets of long spinules as shown. Caudal rami as long as anal somite, slightly divergent, semicylindrical; with seven setae as follows: lateral outer seta I short, arising about midway outer margin, slightly displaced ventrally; seta II slightly displaced dorsally, nearly three times as long as seta I; seta III slightly displaced ventrally, slightly longer than seta II; setae IV and V pinnate, broken off; seta VI arising at inner distal corner, slightly longer than seta III; dorsal seta VII arising close to inner margin, biarticulated.
Antennule (Figure 16A) eight-segmented, short and robust. All segments smooth, except for first segment with spinular rows as shown; with aesthetasc on third and last segments. With inner biarticulate seta on segments six and seven, none of them issuing from long expansion of segments. Relative length of segments as shown. Armature formula: 1-[1], 2-[6], 3-[7+(1+ae)], 4-[2], 5-[2], 6-[2], 7-[2], 8-[6+(1+ae)].
Antenna (Figure 16B) with small unornamented coxa. Basis as long as first endopodal segment, ornamented with proximal spinules as shown, with abexopodal seta as long as segment. Endopod two-segmented; first segment unornamented, with one long, pinnate, subdistal inner seta; second segment longer than first, with two sets of inner spinules as shown, and with shorter spinules subdistally, laterally with two strong geniculate spines and a well-developed geniculate seta, and seven distal elements. Exopod large, three-segmented; first segment with one seta; second segment shortest, with one seta; third segment longest, with one lateral and three distal setae as depicted.
Mandible (Figure 17A–C) with bi- and unicuspid teeth and one dorsal seta on gnathobase. Palp three-segmented composed of basis, exopod and endopod. Basis elongate, with one seta. Exopod one-segmented, with four setae as illustrated. Endopod one-segmented, longer than exopod, with five setae.
Maxillule (Figure 17D) with well-developed praecoxal arthrite with spinular row close to insertion of coxa, armed with two surface setae, and distally with eight strong spines and one small spiniform element. Coxa with epipodal seta; endite with four elements one of which spiniform and pinnate. Basis with some small spinules as shown, and five setae. Exopod one-segmented, small, with three setae. Endopod longer than exopod, with five setae as shown.
Maxilla (Figure 18A) with four endites on syncoxa as follows: two praecoxal endites fused basally of which proximal ornamented with subdistal spinules and with two elements, distal praecoxal endite with three setae; and two elongate coxal endites with three pinnate setae each. Allobasis drawn out into strong claw accompanied by one pinnate spine and one slender seta. Endopod one-segmented, with five elements as shown.
Maxilliped (Figure 18B) three-segmented. Syncoxa with outer log spinules, armed with one proximal short seta. Basis with some long spinules along hump-like inner extension, unarmed. Endopod small, one-segmented; with two lateral, slender setae of which subdistal shorter, and two long distal elements of which outer shorter.
P1 (Figure 19A) with bare intercoxal sclerite as shown. Praecoxa and coxa lost during dissection. Basis with medial spinules, between rami and close to base of inner element, with longer and slender spinules along inner margin; inner and outer spiniform elements with long spinules as shown. Exopod three-segmented, longer than endopod, but apical margin of EXP3 aligned with the apical margin of ENP2 due to a lower insertion site of EXP; EXP1 and EXP2 longest and about the same length, EXP2 shortest; exopodal segments with outer/subdistal spinules as shown; with proximal pore on EXP1; first segment with outer spine, without inner armature; second segment with outer spine and inner seta; third segment with three outer spines, two apical elements of which outer spiniform, and one inner proximal seta. Endopod two-segmented, distal segment longer than proximal; both segments with spinules as depicted; no pores detected; first segment with inner medial seta; second segment with two distal elements of which outer spiniform and shorter, and one inner seta arising below middle of segment.
P2 (Figure 19C) with bare intercoxal sclerite. Praecoxa lost during dissection. Coxa with anterior and posterior outer spinules as illustrated. Basis with medial sets of spinules, with spinules at base of outer spiniform element, between rami, and with longer spinules along inner margin. Exopod three-segmented, longer than endopod, and inserted at a slightly lower level than endopod; EXP1 longest and slightly longer than EXP3, EXP2 shortest; exopodal segments ornamented with spinules as shown; with subdistal pore on EXP2 and EXP3; EXP1 with outer spine and inner reduced seta; EXP2 armed as in preceding segment but inner seta well-developed; EXP3 with three outer spines, two distal elements of which outer spiniform, and two inner setae. Endopod three-segmented, reaching proximal third of EXP3; all segments with outer/subdistal spinules as depicted; with subdistal pore on ENP3; first and second segments subequal in length, with inner seta; third segment longest, with one distal inner seta, one distal element, and one distal outer spine.
P3–P4 (Figure 20A,B) with bare intercoxal sclerites. Praecoxa triangular, with spinular ornamentation as shown. Basis with few spinules at base of outer seta and along inner margin, with (P3) or without (P4) spinules between rami. Exopod three-segmented, longer and inserted at a lower level than endopod; EXP1 longest and slightly longer than EXP3, EXP2 shortest; exopodal segments ornamented with spinules as shown; with subdistal pore on EXP2 and EXP3; EXP1 with outer spine and inner seta (of P3 visibly shorter than that of P4); EXP2 with outer spine and with a well-developed inner seta; EXP3 with three outer spines, two distal elements of which outer spiniform, and two inner setae. Endopod two-segmented, of P3 reaching middle of EXP2, of P4 reaching proximal fifth of EXP2; segments with outer/subdistal spinules as depicted; no pores detected; P3 ENP1 half as long as P3 ENP2, P4 ENP1 and P4 ENP2 subequal in length; P3 ENP1 and P4 ENP1 with inner seta; P3 ENP2 with four elements (two inner setae, and two distal elements of which outer spiniform), P4 ENP2 oval, with one inner and one apical seta.
Both P5 (Figure 15B) separated, transversely elongate; baseoendopod and exopod fused. Baseoendopod with pore at the base of setophore of basal seta, with three elements on endopodal lobe of which outermost shortest. Exopod represented by two setae of which outermost shorter.
Armature formulae of P1–P4 as in Table 4.
Table 4.
Armature formulae of legs 1–4 of Peresime pryocta sp. nov.
Description of male. Male unknown.
Variability. Left P1 ENP2 of holotype without inner seta (Figure 19B), and P2 ENP2 and ENP3 fused, former division indicated by acute outer projection (remainder of outer distal projection of ENP2; arrowed in Figure 19D), and small scar close to inner seta (remainder of division above inner seta of ENP2; arrowed in Figure 19D). Left P2 ENP2 and ENP3 of paratype fused, with inner scar indicating former division between ENP2 and ENP3 (arrowed in Figure 19E), and right P2 ENP2 and ENP3 (not shown) fused completely.
Remarks. Peresime abyssalis is unique within the genus in (i) the reduced distal armature of A2 EXP (with one lateral and two distal setae in Pe. abyssalis but with one lateral and three distal setae on the other species), and (ii) in the reduction in the distal armature of P1 ENP2 (with one lateral and one distal seta in Pe. abyssalis but with one lateral and two distal setae in the other species). The males of the other three species (Pe. pryorae, Pe. reducta, and Pe. pryocta sp. nov.) remain unknown and the sexually dimorphic features cannot be compared.
The relationships between the species of Peresime are not clear. Peresime abyssalis and Pe. reducta share the six-segmented female antennule (it is eight-segmented in Pe. pryorae and Pe. pryocta sp. nov.). Peresime pryorae, Pe. reducta, and Pe. pryocta share the armature complements of A2 EXP (first and second segments with one seta, third segment with one lateral and three distal elements); Pe. abyssalis possesses one seta on the first and second segments, but the third segment possesses one lateral and two distal elements. Peresime abyssalis and Pe. pryorae possess six distal elements on A2 ENP2; Pe. pryocta sp. nov. possesses seven distal elements, the maximum setal number for Idyanthidimorpha [11]; Pe. reducta was described with five distal elements, but this requires confirmation. The mandibular exopod of Pe. abyssalis and Pe. pryorae bears three setae (one lateral and two apical setae), but the mandibular exopod of Pe. reducta and Pe. pryocta sp. nov. possess four setae (one lateral seta and three apical elements). Peresime abyssalis and Pe. pryocta sp. nov. share the presence of one epipodal seta on the coxa of the maxillule; this seta is absent in Pe. pryorae and Pe. reducta and the loss of this seta is regarded here as a synapomorphy for these two species. The apomorphic loss of the epipodal seta of the maxillulary coxa seems to have occurred convergently in the two species of Peresime above, in Z. reyssi, and in Acritozosime. The description of the maxilliped of Pe. abyssalis is incomplete. The armature formula of the maxillipedal syncoxa, basis an endopod of Pe. pryorae, Pe. reducta, and Pe. pryocta sp. nov. is 1;1;4, 0;1;2, and 1;0;4, respectively. The four species of the genus display the primitive heterogeneous spinulation pattern of the outer spines of P1 EXP, and the derived homogeneous spinulation pattern of the outer spines of P3–P4 EXP (see below). However, Pe. reducta was shown with a somewhat heterogeneous spinulation pattern of the outer spines of P2 EXP; the latter requires confirmation.
3.1.3. Genus Heterozosime Kim and Lee, 2021
Type species. Heterozosime tenuis Kim and Lee, 2021, by original designation.
Other species. Heterozosime klemie sp. nov.
Rediagnosis. As in Kim and Lee [1] (pp. 34–35), but the following amendments are introduced here:
- lateral armature o A2 ENP2 composed of two spines and one reduced short seta
- middle apical seta of P4 ENP3 well developed or reduced to a small slender seta
- armature formula of P1–P5 as follows:
| P1 | P2 | P3 | P4 | P5 | |
| EXP | I-0;III,II,2 or I-0;III,2,2 | I-1;I-1;II,I1,2 or I-1;I-1;III;I1,2 | I-1;I-1;II,I1,2 or I-1;I-1;III, I1,2 | I-1;I-1;II,I1,2 or I-1;I-1;III;I1,2 | 3+1 ss |
| ENP | 0-1;I,I,2 or 0-1;1,1,1 | 0-1;0-1;I,2,1♀ 0-1;0,Apo1,1♂ | 0-1;0,1;I,2,1 | 0-1;0-1;I,2,1 | 2 or 4♀ 2♂ |
| Apo, apophysis; ss, surface seta | |||||
- Heterozosime klemie sp. nov.
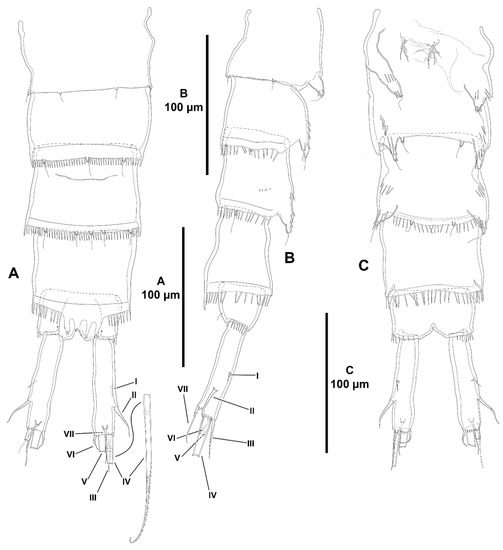
Figure 21.
Heterozosime klemie sp. nov., female holotype (ICML-EMUCOP-120812-01). (A), urosome, dorsal, P5-bearing somite omitted; (B), urosome lateral, P5-bearing somite omitted; (C), urosome, ventral, showing P6 and genital field, P5-bearing somite omitted. Roman numerals indicate the different caudal setae.
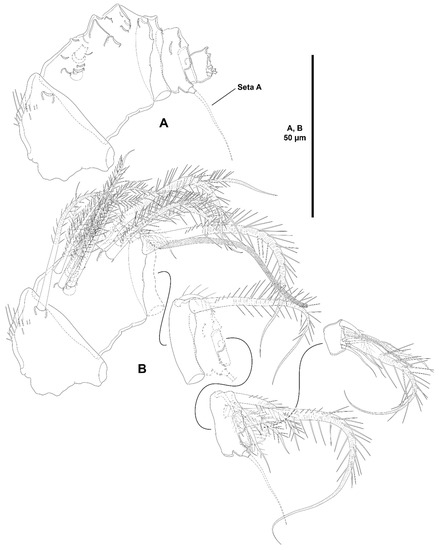
Figure 22.
Heterozosime klemie sp. nov., female holotype (ICML-EMUCOP-120812-01). (A), antennule, armature complements omitted for clarity, seta A indicated; (B), antennule showing armature complements.
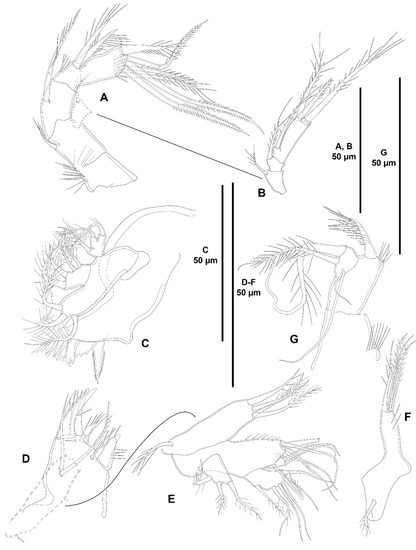
Figure 23.
Heterozosime klemie sp. nov., female holotype (ICML-EMUCOP-120812-01). (A), antenna, showing basis and endopod; (B), exopod of antenna; (C), mandible; (D), praecoxal arthrite of maxillule; (E), coxal endite and basal complex of maxillule; (F), coxal endite, another; (G), maxilliped.
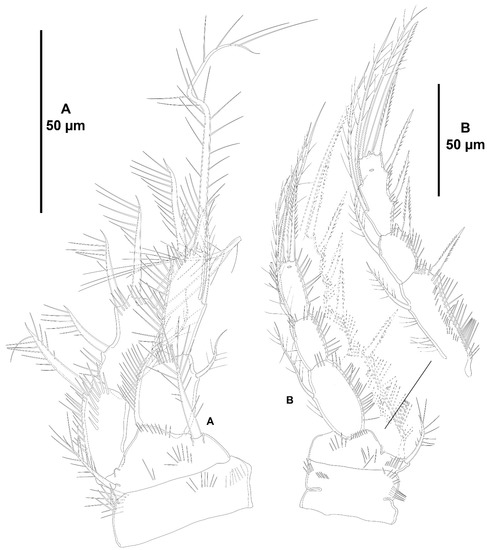
Figure 24.
Heterozosime klemie sp. nov., female holotype (ICML-EMUCOP-120812-01). (A), P1, anterior; (B), P2, anterior.
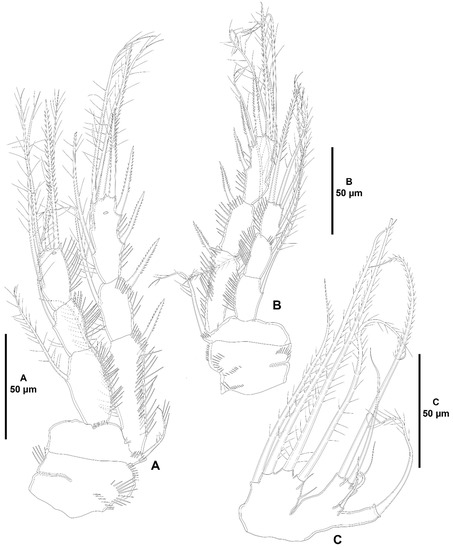
Figure 25.
Heterozosime klemie sp. nov., female holotype (ICML-EMUCOP-120812-01). (A), P3, anterior; (B), P4, anterior; (C), P5, anterior.
ZooBank Registration LSID
urn:lsid:zoobank.org:act:A238862B-E8E1-4F52-9401-51C67EB170FE
Type locality. Talud XV cruise; stn. 5C, eastern Pacific Ocean, off south-western coast of Baja California peninsula, Mexico, 23.230556°N, 110.900556°W; 987 m depth; organic matter, 2.08%, organic carbon, 1.21%, clay 5.73%, silt 33.72%, sand 60.56%; 12 August 2012.
Material examined. Female holotype (ICML-EMUCOP-120812-01) dissected and mounted onto eight slides. Samuel Gómez leg.
Etymology. The specific name is an anagram and arbitrary combination of Dr Jong Guk Kim’s and Dr Jimin Lee’s family names, authors of the genus Heterozosime [1]. Gender feminine.
Description of female. Body severely damaged, estimated total body length, 530 µm measured from anterior margin of body to posterior margin of caudal rami. Habitus tapering posteriad, with constriction between pro- and urosome.
Prosome (not shown) severely damaged, but similar to that of H. tenuis as described in Kim and Lee [1].
Urosome (Figure 21A–C) consisting of P5-bearing somite, genital double-somite, two free somites, and anal somite with caudal rami. Genital somite (second urosomite) and third urosomite separated dorsally, fused ventrally forming genital double-somite. Genital somite (anterior half of genital double-somite) without spinules dorsally and ventrally, posterodorsal margin minutely serrate, with posterior sensilla as shown, with paired ventrolateral sensillum-bearing pleural extensions ornamented with some spinules; P6 and genital field in the middle of somite; P6 with three setae of which outermost longest, medial shortest. Third urosomite (posterior half of genital double-somite) without surface spinules dorsally and ventrally; with spinules and sensilla along posterodorsal margin; ventrally with posterior margin minutely serrate and with ventral sensilla as shown; with paired ventrolateral sensillum-bearing pleural extensions ornamented with some spinules. Fourth urosomite with dorsal row of minute spinules proximally; laterally and ventrolaterally with sparse spinules as shown; with transverse row of spinules along its entire posterior margin; ventrally with sensilla as shown; with paired ventrolateral sensillum-bearing pleural extensions ornamented with spinules as illustrated. Fifth urosomite without surface spinules; with transverse row of spinules along its entire posterior margin but interrupted mediodorsally with pseudoperculum consisting of four crenate projections; with paired ventrolateral unornamented pleural extensions. Anal somite short, anal operculum covered by pseudoperculum of fifth somite not visible, with paired sensilla associated to operculum as illustrated; seemingly without surface spinular ornamentation dorsally and ventrally, but with ventrolateral row of spinules close to insertion site of caudal rami. The latter cylindrical, four times as long as wide, and three times as long as anal somite, slightly divergent; with seven setae as follows: lateral outer seta I short, arising midway outer margin, slightly displaced ventrally; lateral seta II arising at distal third of ramus; seta III issuing distally, slightly displaced ventrally, slightly longer than seta II; seta IV pinnate, posterior half rat-tail like; seta V broken off; seta VI short, arising at inner distal corner; dorsal seta VII arising close to posterior margin, biarticulated.
Antennule (Figure 22A,B) six-segmented, short and robust; second segment longest. All segments smooth, except for first segment with spinular rows as shown; with aesthetasc on second and last segments. With inner biarticulate seta issuing from long expansion of fourth segment (seta A in Figure 22A). Relative length of segments as shown. Armature formula: 1-[1], 2-[18+(1+ae)], 3-[2], 4-[7], 5-[1], 6-[6+(1+ae)].
Antenna (Figure 23A,B) with basis nearly as long as endopodal segments combined, with proximal spinules as shown, with short abexopodal seta. Endopod two-segmented; first segment unornamented, with one long, pinnate, medial inner seta; second segment longer than first, with spinules proximally on inner margin and subdistally, with two strong inner lateral spines and a slender seta, with six distal elements. Exopod large, three-segmented; first segment with one seta; second segment shortest, with one seta; third segment longest, with transverse row of spinules subdistally, with one lateral and three distal setae of different lengths.
Mandible (Figure 23C) with cuspid teeth and one dorsal, thick, pinnate seta on gnathobase. Palp three-segmented composed of basis, exopod and endopod. Basis elongate, with three setae. Exopod one-segmented, with three setae. Endopod one-segmented, with four setae.
Maxillule (Figure 23D–F) with well-developed praecoxal arthrite armed with two surface setae, and distally with eight strong spines and one setiform element. Coxa with epipodal seta; endite with four elements. Basis with some small spinules as shown; with two endites of which dorsal with five, ventral with two setae. Exopod one-segmented, with three setae. Endopod one-segmented, with six setae.
Maxilla lost during dissection.
Maxilliped (Figure 23G) three-segmented. Syncoxa with outer long spinules; unarmed. Basis with some long outer spinules subdistally, with one subdistal seta. Endopod small, one-segmented, with two lateral, slender setae of which subdistal shorter and proximal very long, and two long distal elements of which inner shorter.
P1 (Figure 24A) with medial anterior, and inner and outer posterior spinules on coxa. Basis with spinules medially and at base of endopod; with inner and outer spines. Exopod two-segmented, as long as endopod, but reaching insertion site of inner seta of ENP2 due to a lower insertion site on basis, both segments ornamented with spinules as shown; no pores detected; EXP1 slightly shorter than EXP2, with outer spine, without inner armature; EXP2 with three outer spines, two distal, and two inner setae. Endopod two-segmented, distal segment longer than proximal; both segments with spinules as depicted; no pores detected; ENP1 with inner subdistal seta; second segment with two distal elements of which outer shorter, and one seta arising midway inner margin of segment.
P2–P4 (Figure 24B and Figure 25A,B) with spinules on coxa as figured. Basis of P2 with spinules medially, at base of endopod and close to outer spiniform element; basis of P3 with spinules at base of endopod and at base of outer seta; basis of P4 as in P3 but without spinules at base of endopod, outer seta visibly longer than in P2 and P3. Exopod three-segmented, longer than endopod; EXP1 longest, EXP2 shortest; exopodal segments ornamented with spinules as shown; with subdistal pore on P2 EXP3 and P3 EXP3; EXP1 with outer spine and inner seta (inner seta of P2 comparatively shorter that in P3 and P4); EXP2 armed as in preceding segment; EXP3 with three outer spines, two distal elements of which outer spiniform, and two inner setae. Endopod three-segmented, of P2 reaching distal fourth of EXP3, of P3 reaching middle of EXP3, of P4 barely beyond distal margin of EXP2; all segments with spinules as depicted; with subdistal pore on P2 ENP3 and P3 ENP3; first segment longest (P2 and P3) or as long as second segment (P4); first and second segment with inner seta; third segment with three distal setae of which outer spiniform, and one inner element.
Both P5 (Figure 25C) separated; baseoendopod and exopod fused but separated by deep notch. Baseoendopod with outer basal seta arising from long setophore, with four elements on endopodal lobe. Exopod with three distal setae plus a surface seta between base of exopodal lobe and setophore of basal seta.
Armature formulae of P1–P4 as in Table 5.
Table 5.
Armature formulae of legs 1–4 of Heterozosime klemie sp. nov.
Description of male. Male unknown.
Remarks. The new species of Heterozosime presented here, H. klemie sp. nov., was attributed to that genus by the combination of (i) the six-segmented female antennule with aesthetasc on second segment; (ii) the presence of one seta on A2 EXP1; (iii) the presence of two spines and one small slender setae laterally on A2 ENP2; (iv) the mandibular palp with three, three and four setae on the basis, exopod, and endopod, respectively; (v) the presence of one epipodal seta on the coxa of the maxillule; (vi) the two-segmented maxillary endopod; (vii) the non-geniculate—stenopodial—maxilliped; (viii) the unarmed maxillipedal syncoxa; (ix) the two-segmented P1 EXP with two inner setae on P1 EXP2; (x) the two-segmented P1 ENP; (xi) the three-segmented P2–P4 EXP and ENP; (xii) the primitive and derived spinulation pattern on the outer spines of P1 EXP, and P2–P4 EXP, respectively; and (xiii) the female P5 EXP with three marginal and one outer surface seta. However, some differences between H. tenuis and H. klemie sp. nov. were observed in (i) the shape of distal elements of P1 EXP2 and P1 ENP2 (spiniform in H. tenuis but setiform in the new species); (ii) the armature complement of P1 ENP2 (with four elements in H. tenuis but three in H. klemie sp. nov.; (iii) the armature complement of P2–P4 EXP3 (with six elements in H. tenuis but seven in H. klemie sp. nov., i.e., H. klemie sp. nov. kept the plesiomorphic armature complement of three outer spines on the third exopodal segment of P–P4); (iv) shape of the medial distal seta on P4 ENP3 (reduced to a small seta in H. tenuis but well-developed in the new species); and (v) number of setae on the endopodal lobe of the female P5 (with two setae in H. tenuis but four elements in H. klemie sp. nov.). Unfortunately, the male of H. klemie sp. nov. remains unknown. The reduction in the medial seta of P4 ENP3, and the reduction in armature complement of the female P5 endopodal lobe seem to have occurred independently several times in Zosimeidae. The apomorphic reduced small seta of P4 ENP3 observed in H. klemie sp. nov., is also present in Z. montagnai, Z. mexicana sp. nov., and Z. pacifica (see above). The bisetose endopodal lobe of the female P5 is present, and probably synapomorphic, in few species of Zosime (Z. mexicana sp. nov., Z. montagnai, and Z. pacifica (typica-group, subordinate subgroup Aa), and in Z. valida, Z. destituta, and Z. changi (typica-group, subordinate subgroup Ba). The secondary loss of two setae of the female P5 endopodal lobe of H. tenuis is regarded here as apomorphic. The two-segmented condition of P1 EXP present in Acritozosime and Heterozosime is regarded here as a synapomorphy for both genera (see below).
3.1.4. Genus Acritozosime Kim and Lee, 2021
Type species. Acritozosime spinesco Kem and Lee, 2021, by original designation.
Other species. Acritozosime sigsbeeiensis sp. nov.
Rediagnosis. As in Kim and Lee [1] (p. 48), but the following amendments are introduced here:
- (i).
- lateral armature o A2 ENP2 composed of two spines and one well-developed seta
- (ii).
- syncoxa of maxilliped with one or two setae
- (iii).
- maxilliped geniculate or non-geniculate
- (iv).
- armature formula of P1–P5 as follows:
| P1 | P2 | P3 | P4 | P5 | |
| EXP | I-0;III,I1,1 | I-1;I-1;II,I1,1 or I-0;I-1;II;I1,1 | I-1;I-1;II,I1,2 or I-0;I-1;II, I1,2 | I-1;I-1;II,I1,2 | 3 |
| ENP | 0-0;0,I1,0 | 0-0;0,I1,0♀ 0-0;0,Apo1,0♂ | 0-0;0,2,1 | 0-1;0,2,1 | 3♀ 2♂ |
| Apo, apophysis | |||||
- Acritozosime sigsbeeiensis sp. nov.
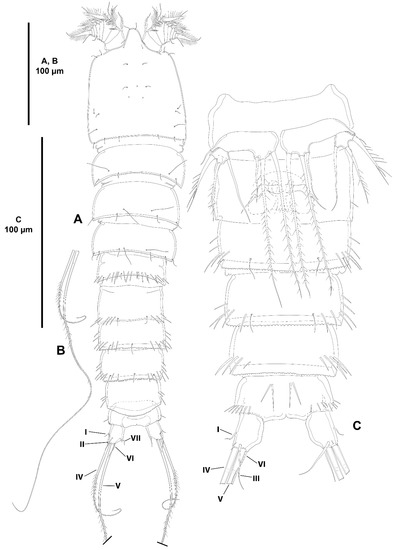
Figure 26.
Acritozosime sigsbeeiensis sp. nov., female holotype (ICML-EMUCOP-061816-02). (A), habitus, dorsal; (B), caudal setae IV and V; (C), urosome, ventral. Roman numerals indicate the different caudal setae.
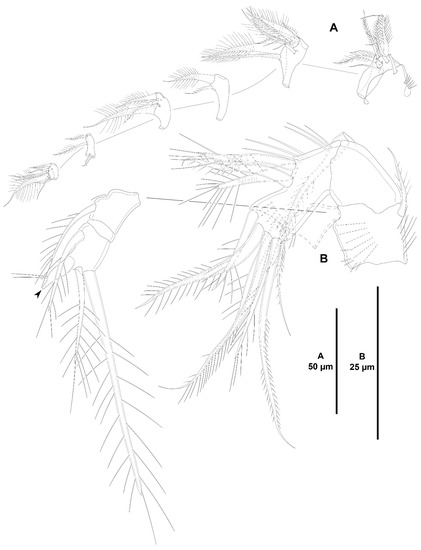
Figure 27.
Acritozosime sigsbeeiensis sp. nov., female holotype (ICML-EMUCOP-061816-02). (A), antennule; (B), antenna, reduced distal element of exopod indicated.
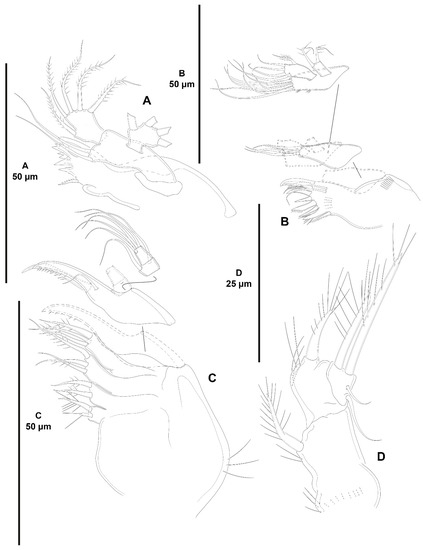
Figure 28.
Acritozosime sigsbeeiensis sp. nov., female holotype (ICML-EMUCOP-061816-02). (A), mandible; (B), maxillule; (C), maxilla; (D), maxilliped.
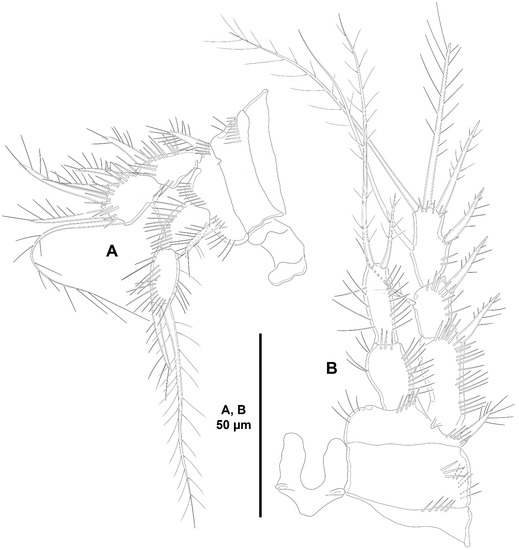
Figure 29.
Acritozosime sigsbeeiensis sp. nov., female holotype (ICML-EMUCOP-061816-02). (A), P1, anterior; (B), P2, anterior.
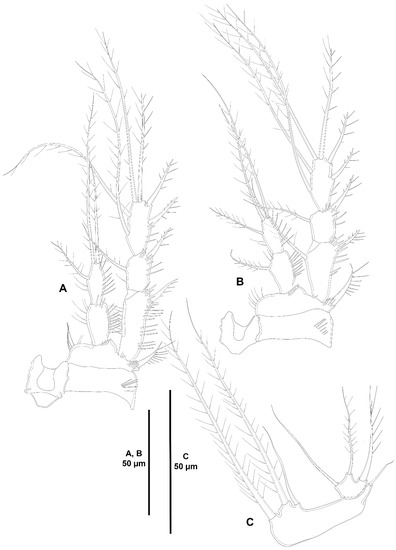
Figure 30.
Acritozosime sigsbeeiensis sp. nov., female holotype (ICML-EMUCOP-061816-02). (A), P3, anterior; (B), P4, anterior; (C), P5, anterior.
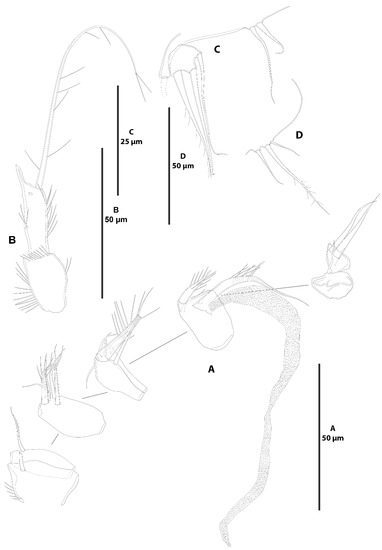
Figure 31.
Acritozosime sigsbeeiensis sp. nov., male allotype (ICML-EMUCOP-061816-03). (A), antennule; (B), P2 ENP, anterior; (C), P5; (D), P6.
ZooBank Registration LSID
urn:lsid:zoobank.org:act:E11D6186-C484-4034-A66B-B85F1D34DAD7
Type locality. Xiximi-5 cruise; stn. A8, Gulf of Mexico, eastern Sigsbee Deep, 25.1168° N, 89.0500° W; 3477 m depth; clay, 55.5%, silt, 23.0%, sand, 21.5%; 18 June 2016.
Material examined. Female holotype (ICML-EMUCOP-061816-02) dissected and mounted onto six slides; male allotype (ICML-EMUCOP-061816-03) dissected and mounted onto nine slides. Ivonne Martínez-Mendoza leg.
Etymology. The specific epithet comes from the family name of Commander Charles Dwight Sigsbee who discovered the abyssal plain during a mapping campaign of the Gulf of Mexico and refers to the place where the new species was found. The Latin suffix -iēnsis, is a variation of -ēnsis, of, from. It is in the nominative singular, gender feminine.
Description of female. Total body length, 425 µm measured from anterior margin of rostrum to posterior margin of caudal rami. Habitus (Figure 26A) subcylindrical, slightly tapering posteriad, with constriction between pro- and urosome.
Prosome (Figure 26A) consisting of cephalothorax and free P2–P4-bearing somites. Rostrum fused to cephalothorax, well developed, triangular, with pair of subdistal sensilla. Cephalothorax large, subquadrate; with lateral margins almost parallel in dorsal view; posterior margin smooth; with sensilla as shown. P2–P4-bearing somites without dorsal spinular ornamentation; posterior margin minutely serrate; with two dorsal long sensilla, and with shorter sensilla along posterior margin.
Urosome (Figure 26A–C) consisting of P5-bearing somite, genital double-somite, two free somites, and anal somite with caudal rami. P5-bearing somite with continuous transverse posterior row of long spinules; posterior margin minutely serrate; with posterior short sensilla as shown; ventrally without spinular ornamentation. Genital somite (second urosomite) and third urosomite separated dorsally, partially fused ventrally forming genital double-somite. Genital somite (anterior half of genital double-somite) with smooth posterodorsal margin; with dorsolateral posterior row of long spinules interrupted medially, and with few posterior sensilla, ventrally without spinular ornamentation, no sensilla detected; P6 and genital field in the middle of somite ventrally, covered with detritus, only one seta visible. Third urosomite (posterior half of genital double-somite) with serrate posterior margin coarser than in prosomites; ornamented as in previous somite but with some additional ventrolateral spinules; with sensilla as shown. Fourth urosomite largely as previous somite dorsally and ventrally. Fifth urosomite with ventrolateral spinules along posterior margin as depicted; posterior serrate margin less coarse than in preceding somites, extended medially forming a pseudoperculum; without sensilla. Anal somite short; with small operculum ornamented with minute spinules along its posterior margin, accompanying sensilla not observed due to accumulation of detritus; with small spinules on outer posterior corner, and with larger ventrolateral spinules; ventrally with two sets of two long spinules each. Caudal rami as long as anal somite, slightly divergent, semicylindrical proximally, tapering distally; with seven setae as follows: lateral outer seta I short, arising about midway outer margin; seta II displaced dorsally; seta III displaced ventrally, long; setae IV and V pinnate, rat-tail like, the latter longest; seta VI arising at inner distal corner, as long as seta II; dorsal seta VII arising close to inner margin, biarticulated.
Antennule (Figure 27A) seven-segmented; short and robust. All segments smooth, except for first segment with short spinular row; with aesthetasc on third and seventh segments. With two inner biarticulate setae on segment six, none of them issuing from long expansion of segment. Relative length of segments as shown. Armature formula: 1-[1], 2-[9], 3-[8+(1+ae)], 4-[2], 5-[3], 6-[4], 7-[5+(1+ae)].
Antenna (Figure 27B) with basis ornamented with spinules as shown, with abexopodal seta as long as segment. Endopod two-segmented; first segment slightly shorter than second, unornamented, with one long, pinnate, inner subdistal seta; second segment with two sets of long inner spinules, and with transverse row of shorter spinules subdistally, armed with two strong geniculate spiniform and one geniculate setiform element, and seven distal elements as shown. Exopod large, three-segmented; first segment as long as third, with two setae of which proximal very small; second segment shortest, unarmed; third segment with one lateral and three distal setae one of which short and thick (the latter arrowed in Figure 27B).
Mandible (Figure 28A) with multi- and unicuspid teeth and one dorsal pinnate seta on gnathobase. Palp three-segmented, composed of basis, exopod and endopod. Basis elongate, with two seemingly bare setae. Exopod one-segmented, with four setae. Endopod one-segmented, with one lateral, one subdistal, and three distal setae.
Maxillule (Figure 28B) with well-developed praecoxal arthrite ornamented with one spinular row close to insertion of coxa, and one short row on arthrite as depicted; armed with two surface setae, and distally with seven spines and one unipinnate seta. Coxal endite with three setae one of which pinnate. Basis with some small spinules as shown, and seven setae. Exopod one-segmented, small, with two apical setae. Endopod longer than exopod, with three distal setae, and one lateral seta on each side one of which shorter and pinnate.
Maxilla (Figure 28C) with large syncoxa ornamented with few long outer spinules; with four endites as follows: two praecoxal endites fused basally, each with two elements; and two elongate coxal endites with two pinnate and one bare seta each. Allobasis drawn out into strong unipinnate claw accompanied by one unipinnate spine and one slender seta. Endopod two-segmented; proximal segment with one anterior and one posterior seta; distal segment with one subdistal and three distal setae.
Maxilliped (Figure 28D) three-segmented. Syncoxa with proximal spinules as shown, armed with one pinnate seta. Basis with some long inner spinules and armed with one distal element. Endopod small, one-segmented, with two lateral, slender setae of which subdistal shorter, and two distal elements of different lengths.
P1 (Figure 29A) with bare intercoxal sclerite as shown. Praecoxa unornamented. Coxa rectangular, with some outer spinules as shown. Basis with spinules at the base of outer and inner pinnate spines and between rami, and with long slender spinules along inner margin. Exopod two-segmented, about as long as endopod but inserted at a lower level on basis; both segments with long outer/subdistal spinules as illustrated; no pores detected; first segment with outer spine, without inner armature; second segment with three outer spines of which proximal two subequal in length and subdistal visibly longer, two distal elements of which outer spiniform and inner setiform, and one short slender inner seta. Endopod two-segmented; both segments subequal in length and ornamented with long spinules as depicted; no pores detected; first segment unarmed; second segment with two distal elements of which outer spiniform, inner setiform and long.
P2–P4 (Figure 29B and Figure 30A,B) with bare intercoxal sclerites. Praecoxa of P2 with few transverse subdistal spinules, of P3 and P4 lost during dissection. Coxa with outer row of spinules anteriorly, of P2 with, of P3 and P4 seemingly without posterior outer spinules. Basis with spinules at the base of outer element and between rami, and with longer slender spinules along inner margin. Exopod three-segmented, EXP1 longest, EXP2 shortest; exopodal segments ornamented with outer/subdistal spinules as shown; no pores detected; P2–P4 EXP1 with outer spine, P2 and P3 EXP1 without, P4 EXP1 with inner seta; P2–P4 EXP2 with outer spine and inner seta; P2–P4 EXP3 with two outer spines, and two distal elements of which outer spiniform and inner setiform, and one (P2) or two (P3 and P4) inner setae. Endopod two-segmented; of P2 reaching proximal fourth of EXP3, of P3 and P4 barely reaching insertion site of inner seta of EXP2; each segment ornamented with spinules as shown; no pores detected; P2–P3 ENP1 without, P4 ENP1 with inner seta; P2 ENP2 with two distal elements of which outer spiniform and shorter, without inner armature; P3–P4 ENP2 with two setiform apical setae of which outer slightly shorter, with one inner seta.
Both P5 (Figure 26C and Figure 30C) separated; baseoendopod transversely elongate; baseoendopod and exopod distinct. Baseoendopod with outer basal seta arising from setophore; with three elements on endopodal lobe of which outermost shortest and bare. Exopod small; with three setae of which inner bare, outer shortest.
Armature formulae of P1–P4 as in Table 6.
Table 6.
Armature formulae of legs 1–4 of Acritozosime sigsbeeiensis sp. nov.
Description of male. Body badly damaged, largely as in female. Total body length, 250 µm measured from anterior margin of rostrum to posterior margin of caudal rami. Sexual dimorphism expressed in the antennules, segmentation of urosome (genital somite and third urosomite separated), P2 ENP, P5 and P6.
Antennule (Figure 31A) eight-segmented, subchirocer. All segments smooth except for first segment with row of spinules. Aesthetasc detected only on sixth segment. Armature formula: 1-[1], 2-[1], 3-[6], 4-[5], 5-[2], 6-[3+(1+ae)], 7-[1], 8-[4].
Antenna, mandible, maxillule, maxilla, and maxilliped (not shown) as in female.
P1 (not shown) as in female.
P2 as in female, except for two-segmented ENP (Figure 31B) with first segment unarmed, second segment with outer acute projection and armed with only one apical seta.
P3 and P4 (not shown) as in female.
P5 (Figure 31C) seemingly not fused to somite, transversely elongate; baseoendopod and exopod distinct. Endopodal lobe fully incorporated to baseoendopod, represented by two setae of which inner shorter. Exopod small, with one outer, one distal, and one inner seta as shown.
Both P6 seemingly functional, not fused to somite; with three setae of which inner shortest (Figure 31D).
Armature formulae of P1–P4 as in Table 6.
Remarks. The new species presented here was attributed to the genus Acritozosime on account of (i) the seven-segmented female antennule; (ii) the bisetose A2 EXP1; (iii) the presence of two spines and one long seta laterally on A2 ENP2; (iv) the presence of seven distal elements on A2 ENP2; (v) the mandibular basis, exopod, and endopod with two, four (one lateral, three distal elements), and five setae (one lateral, four distal elements); (vi) the presence of setal armature on the syncoxa of the maxilliped; (vii) the armature formulae of P1–P4 ENP and P1 EXP and P3–P4 EXP; (viii) the primitive homogeneous spinular ornamentation of the outer spines of P1–P4 EXP; and (ix) the female P5 EXP and endopodal lobe with three setae each. Acritozosime spinesco and A. sigsbeeiensis sp. nov. differ in (i) the maxilliped (with geniculation between syncoxa and basis, and probably between basis and endopod in A. spinesco but straight—stenopodial—without any sign of geniculation in A. sigsbeeiensis sp. nov.); (ii) the reduced armature complement of P2–P3 EXP1 (with inner seta in A. spinesco, unarmed in A. sigsbeeiensis sp. nov.); and (iii) the relative length of caudal seta V (nearly as long as the whole body in A. spinesco but visibly shorter in A. sigsbeeiensis sp. nov.). Kim and Lee [1] noticed that the loss of the inner element of the A2 ENP1, the loss of the epipodal seta of the maxillulary coxa, and the elongate caudal seta V are unambiguous features of A. spinesco. However, the presence of an inner seta on A2 ENP1 in A. sigsbeeiensis sp. nov. indicates that the loss of that seta in A. spinesco could be secondary and apomorphic for the latter. The apomorphic loss of the inner seta of A2 ENP1 occurred convergently in Z. thistlei. The apomorphic loss of the epipodal seta on the maxillulary syncoxa has been reported also for Z. reyssi, Pe. pryorae, and Pe. reducta and seems to have also occurred independently in Acritozosime. The elongate caudal seta V seems to be autapomorphic for A. spinesco [1]. Regarding the two-segmented P1 EXP, Kim and Lee [1] proposed independent origins of Acritozosime and Heterozosime (see below). However, the two-segmented condition of P1 EXP most probably occurred independently in Idyanthidae and Zosimeidae, and Acritozosime and Heterozosime probably share a common ancestor, where the two-segmented P1 EXP is a synapomorphy for both genera (see below).
3.2. Phylogenetics
Our phylogenetic analysis (Figure 32) with 88 characters (Table 1) and 36 zosimeid species, and six outgroup taxa (Table S1) showed that (i) the monotypic genus Pseudozosime is the adelphotaxon of Peresime; (ii) the genus Peresime is composed of two clades, abyssalis-reducta and pryocta-pryorae; (iii) Heterozosime is the adelphotaxon of Acritozosime; (iv) the subdivision of the genus Zosime into two groups (the incrassata- and the typica-group as proposed by Kim et al. [12] and modified by Pointner [13] is phylogenetically grounded, and the typica-group can be subdivided into two subgroups each with some natural subordinate subgroups; (v) the new species presented herein belong to the typica-group; (vi) Z. xiximi sp. nov. is closely related to Z. mediterranea and Z. atlantica, forming a well-defined clade (subordinate subgroup Ab); and (vii) Z. mexicana sp. nov. belongs to subordinate subgroup Aa and is closely related to Z. paratypica and Z. montagnai, forming a well-defined clade within the respective subordinate subgroup.
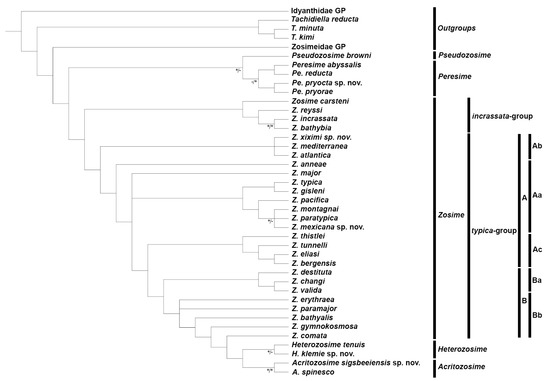
Figure 32.
Phylogenetic tree of the family Zosimeidae based on 88 morphological traits showing the different groupings of Zosime. Support values at nodes were estimated from the bootstrap (boot) of the maximum parsimony and BI posterior probabilities (pp), represented as boot/pp. * Nodes well-supported, boot > 85%; boot > 0.90. Values < 85% (boot) and < 0.90 (pp) are not shown or indicated by “-”.
4. Discussion
4.1. Phylogenetic Analysis
Most characters used in our analysis are self-explanatory. Following is the explanation of some characters used in our phylogenetic analysis.
- −
- Characters 2–5 (fusion of segments of the female antennule; see also Table S2). Seifried (2003) proposed the ground pattern of the female antennule of the Idyanthidimorpha and Idyanthidae with nine segments and armature formulae I-[1], II-[11], III-[10], IV-[4+ae], V-[3], VI-[4], VII-[2], VIII-[2], IX-[7+ae], and I-[1], II-[11], III-[10], IV-[4+ae], V-[3], VI-[3], VII-[2], VIII-[2], IX-[7+ae], respectively, and of Zosimeidae with eight segments and armature formula I-[1], II-[7], III-[9], IV-[3+ae], V-[1], VI-[4], VII-[4], VIII-[7+ae], being the tetrasetose segment VII of the ground pattern of Zosimeidae, the result of fusion of the idyanthid bisetose primitive segments VII and VIII.
- The female antennules in Zosimeidae are very compact and short, the boundaries between segments are sometimes difficult to see, some segments are ill-separated, and the setae are piled making the counting and homologation of the armature elements very difficult. However, there are two elements that are easy to identify, the proximal aesthetasc and an outer biarticulate seta arising from a strong outer extension of segment 4, 5, or 6 (hereafter referred to as seta A; see Figure 3A, Figure 9A and Figure 22A).The proximal aesthetasc is primitively located on the fourth segment in Idyanthidae and Zosimeidae [11], and is commonly fused basally to a single seta. This armature complex is located either on the second, third, or fourth actual segments in Zosimeidae (Table S2). Its position in the fourth segment indicates that there has not been any fusion of the proximal primitive segments I–IV like in Z. valida [26], Z. pacifica, and Z. destituta, and Pe. pryorae; its position on the third segment indicates fusion of primitive segments II and III like in Z. anneae, Z. xiximi sp. nov., Z. eliasi, Z. gymnokosmosa, Z. carsteni, Z. mexicana sp. nov., Z. montagnai, Z. thistlei, Z. atlantica, Z. changi, and Z. tunnelli, or fusion of primitive segments III and IV like in Pe. pryocta sp. nov. and A. sigsbeeiensis sp. nov.; its position on the second segment indicates that primitive segments II–IV became fused like in Z. comata, Heterozosime, and A. spinesco. However, the fact that (i) the aesthetasc lies on the third segment of the eight-segmented female antennule of Z. anneae, Z. xiximi. sp. nov., Z. eliasi, and Z. gymnokosmosa (these species underwent secondary fusion of primitive segments II and III forming the actual second segment), or in the second segment of the seven-segmented female antennule of A. spinesco (the aesthetasc-bearing second segment is the result of the fusion of primitive segments II–IV), and (ii) that the primitive segments II and III are separated in the seven-segmented female antennule of Z. valida, in the six-segmented female antennules of Z. pacifica, and Z. destituta, and in the eight-segmented female antennule of Pe. pryorae, indicates that the ground pattern of Zosimeidae includes a nine-segmented female antennule, instead of an eight-segmented appendage as in Seifried [11]. Moreover, some species display two setae on the antennulary segment that is most probably homologous to the primitive segment V (fourth segment in Z. xiximi sp. nov., Z. eliasi, Z. gymnokosmosa, Z. incrassata, Z. changi, Pe. pryocta sp. nov., and A. sigsbeeiensis sp. nov.; third segment in Z. comata, Heterozosime, and A. spinesco). Therefore, the armature formula of the ground pattern of Zosimeidae is assumed to be 1-[1], 2-[7], 3-[9], 4-[3+ae], 5-[2], 6-[4], 7-[2], 8-[2], 9-[7+ae]. Some other amendments in the armature complement of the antennulary segments are probable, but this is beyond the scope of the present discussion.Seta A is present in most members of the family, and it can be located either on the fourth, fifth, or sixth antennulary segment (Table S2). This seta can be traced back to the primitive segment VII of Idyanthidae. The outer seta on the actual seventh and eighth segments is still present in the nine-segmented antennule of Idyanthe australis Pallares, 1970. The fusion of the idyanthid primitive segments VII and VIII resulted in the actual seventh segment of the eight-segmented female antennule of Idyella kunzi Bodin, 1968, I. nilmaensis Kornev and Chertoprud, 2008, Tachidiella kimi Lee and Huys, 1999, and T. sarsi Kornev and Chertoprud, 2008, with two outer setae, amongst other inner elements. Zosime anneae and P. pryocta sp. nov. kept the—primitive—idyanthid bisetose sixth and seventh segments (homologous to the primitive segments VII and VIII) in their eight-segmented female antennules, and they became fused in A. sigsbeeiensis sp. nov. forming its tetrasetose sixth segment (see Figure 27A). The outer biarticulate seta of the sixth segment in Z. carsteni, Z. eliasi, and Z. gymnokosmosa, and in the fifth segment of Z. thistlei (homologous to the primitive segment VII) arises from poorly developed extension of the segment. This extension is well-developed in the sixth segment of Z. atlantica, Z. xiximi sp. nov., in the fifth segment of Z. changi, Z. tunnelli, Z. montagnai, Z. destituta, Z. pacifica, Z. mexicana, and in the fourth segment of Z. comata and H. klemie sp. nov. and is regarded here as apomorphic for these species.The position of the proximal aesthetasc and the position of seta A on the female antennule are used here as landmarks to better understand the segmentation patterns of the female antennule of Zosimeidae.The number of segments of the female antennule of Zosime varies from six to eight (see Table S2) and seems to be distributed randomly amongst the species of the typica-group and its subordinate subgroups; the number of segments of the female antennule varies from seven to eight in the incrassata-group, but the armature formula is known only for Z. carsteni. The female antennule is composed of six or eight segments in Peresime, but the armature formula is known only for Pe. pryorae and Pe. pryocta sp. nov. with eight-segmented antennules. The monotypic genus Pseudozosime has five-segmented female antennules, but the armature formula remains unknown. It is six-segmented in Heterozosime and seven-segmented in Acritozosime.The fusion pattern of the primitive antennulary segments of Zosimeidae is not clear. Briefly, fusion took place in primitive segments II and III, II–IV, IV and V, V and VI, V–VII, V–VIII, VI and VII, VII and VIII in Zosime; in V and VI in Pe. pryorae; in III and IV in Pe. pryocta sp. nov.; in II–IV, and VI and VII in H. tenuis and H. klemie sp. nov.; in II–IV in A. spinesco; and in III and IV, and VII and VIII in A. sigsbeeiensis sp. nov.The eight-segmented antennule of Z. anneae, Z. xiximi sp. nov., Z. eliasi, Z. gymnokosmosa, and Z. carsteni is the result of fusion of primitive segments II and III, resulting in the third and sixth segment with the proximal aesthetasc and seta A, respectively, except for Z. anneae in which the strong extension of the supporting segment from which seta A arises seems to have been suppressed.The seven-segmented female antennules in Zosime are the result of different processes. Zosime mexicana sp. nov., Z. montagnai, and Z. thistlei underwent fusion of primitive segments II and III, and IV and V, resulting in the third and fifth segment with the proximal aesthetasc and seta A, respectively. The seven-segmented female antennule of Z. atlantica, Z. changi, and Z. valida is the result of fusion of primitive segments II and III, and VII and VIII in Z. atlantica with the proximal aesthetasc and seta A on the third and sixth segment, respectively; II and III, and VI and VII in Z. changi with the proximal aesthetasc and seta A on the third and fifth segment, respectively; and V–VII in Z. valida with the proximal aesthetasc on the fourth segment (the position of seta A is inconclusive, but it is, in all probability, on the sixth segment).The six-segmented female antennules of Z. tunnelli, Z. comata, and Z. pacifica and Z. destituta is the result of fusion of primitive segments II and III, V and VI, and VII and VIII resulting in the third and fifth segments with the proximal aesthetasc and seta A, respectively, in Z. tunnelli; II–IV, and VI and VII in Z. comata with the proximal aesthetasc on the second segment and seta A on the fourth segment; and V–VIII in the latter two species with the proximal aesthetasc on the fourth segment and seta A on the fifth segment.The fusion pattern is also different in the eight-segmented female antennules of the two species of Peresime for which the armature formulae are known. It is the result of fusion of primitive segments V and VI in Pe. pryorae with the proximal aesthetasc and seta A on the fourth and sixth segment, respectively, but III and IV in Pe. pryocta sp. nov. with the proximal aesthetasc on the third segment (the development of the strong extension of the supporting segment from which seta A arises seems to have been suppressed).Both species of Heterozosime underwent fusion of primitive segments II–IV, and VI and VII, resulting in six-segmented antennules with the proximal aesthetasc on the second segment, and seta A on the fourth segment.The fusion pattern in both species of Acritozosime is different. The seven-segmented female antennule of A. spinesco seems to be the result of fusion of primitive segments II–IV, resulting in the second and fifth segment with the proximal aesthetasc and seta A, respectively; but in A. sigsbeeiensis sp. nov. the fusion took place in primitive segments III and IV, and VII and VIII, resulting in the third segment with the proximal aesthetasc (the outer extension of the supporting segment from which seta A arises seems to have been suppressed).The phylogenetic value of the fusion pattern between the species of each genus is not clear, but some species share a particular set of fusion events, e.g., (i) Z. anneae, Z. xiximi sp. nov., Z. eliasi, and Z. gymnokosmosa (the armature complement of the third segment of Z. carsteni—12 setae+ae—is significantly different); (ii) Z. mexicana sp. nov., Z. montagnai, and Z. thistlei; (iii) Z. pacifica and Z. destituta; (iv) H tenuis and H. klemie sp. nov. (Table S2).
- −
- Characters 6–10 (armature of A2 EXP). The A2 EXP1 is armed with one seta in all species of Zosime for which this ramus has been described (it is unknown in Z. gisleni, Z. major, Z. mediterranea, Z. erythraea, Z. bathyalis, and Z. reyssi), and in Peresime, Pseudozosime, and Heterozosime. Acritozosime is the only genus of the family with the plesiomorphic bisetose A2 EXP1 (see Kim and Lee [1] for an in-depth discussion of this character).
- The second exopodal segment bears one seta in all species of the typica-group of Zosime, in Peresime, Pseudozosime, and Heterozosime. This seta is absent in the incrassata-group except for Z. carsteni with one seta; it is absent also in Acritozosime. The loss of the seta of A2 EXP2 seems to have occurred independently several times within the family and is regarded here as synapomorphic for Z. incrassata, Z. bathybia and Z. reyssi, and apomorphic for Acritozosime (Table S1; Figure 32).The armature of EXP3 is variable. One inner lateral seta is present in all species of subgroup A of Zosime except for Z. atlantica and Z. xiximi sp. nov. (typica-group, subordinate subgroup Ab; the condition in Z. mediterranea remains unverifiable), and for the incrassata-group (except for Z. carsteni with one lateral inner seta). Zosime atlantica, Z. xiximi sp. nov., Z incrassata, Z. bathybia, and Z. reyssi underwent secondary—apomorphic—loss of this seta, and it is the most derived condition in the family. The inner lateral seta of A2 EXP3 is also present in all the other genera of the family.The distal armature complement of the A2 EXP3 varies from three to two. Three setae are present in some species of Zosime (Z. anneae, Z. paratypica, Z. mexicana sp. nov., and Z. pacifica (typica-group of Zosime, subordinate subgroup Aa), Z. atlantica and Z. xiximi sp. nov. (typica-group of Zosime, subordinate subgroup Ab), Z. thistlei (typica-group of Zosime, subordinate subgroup Ac), and Z. bathybia (incrassata-group)), in Pe. pryorae, Pe. reducta, and Pe. pryocta sp. nov., and in Heterozosime, and Acritozosime. Two setae are present in some species of Zosime (Z. typica and Z. montagnai (typica-group, subordinate subgroup Aa), and Z. eliasi, Z. bergensis, and Z. tunnelli (typica-group, subordinate subgroup Ac); the condition of Z. gisleni and Z. major (typica-group, subordinate subgroup Aa), and Z. mediterranea (typica-group, subordinate subgroup Ab) remain unverifiable)), in Z. carsteni, Z. incrassata, and Z. reyssi (incrassata-group of Zosime), and in Pe. abyssalis.
- −
- Character 11 (presence/absence of the inner seta of A2 ENP1). Most species of Zosimeidae possess one inner seta on A2 ENP1. This seta is present also in Idyanthidae. The inner seta of A2 ENP1 is missing in Z. paratypica, Z. thistlei, and A. spinesco. The loss of this element in two species of Zosime and in one species of Acritozosime seems to have occurred independently and is considered apomorphic for A. spinesco. It is also an apomorphy (potential synapomorphy?) for Z. paratypica and Z. thistlei in their respective subordinate subgroups.
- −
- Characters 12–13 (A2 ENP2, lateral armature). Most species of Zosimeidae display the armature of the second antennary endopod as described by Seifried [11] in her diagnosis and ground pattern of the family, i.e., with one proximal short (element I in Seifried [11]), and one subdistal spine (element III in Seifried [11]) set close to a reduced slender seta (element 4 in Seifried [11]). This setal arrangement occurs in Zosime except for some species with only two lateral spines (Seifried’s [11] elements I and III) on the second endopodal segment of the antenna, Z. anneae, Z. montagnai, Z. eliasi, Z. tunnelli, Z. valida, and Z. incrassata. It is unclear whether the absence of the small seta 4 is a true secondary loss or if this small seta has been overlooked in these species since it is often masked by element III. On the other hand, Peresime and Acritozosime deviates from the general ground pattern suggested by Seifried [11] for the family. The lateral armature of A2 ENP2 of Peresime and Acritozosime consists of three well-developed elements, i.e., spiniform elements I and III, and—primitive—well-developed setiform element 4. Element 4 is primitively well-developed and geniculate in Idyanthidae [11]. Elements I, III, and 4 are geniculate in Pe. pryocta sp. nov. and in A. sigsbeeiensis sp. nov., and their conditions in the other species of Peresime, and in A. spinesco remain uncertain. As noted above, the well-developed element 4 in Peresime and Acritozosime seems to be a plesiomorphy, and the reduction in seta 4 seems to have occurred independently in Zosime, Pseudozosime, and Heterozosime.
- −
- Character 14 (number of distal elements on A2 ENP2). The ground pattern of Idyanthidimorpha and Idyanthidae include seven distal elements on A2 ENP2 [11]. Most members of the family Zosimeidae underwent the synapomorphic loss of one element resulting on six distal setae/spines on A2 ENP2. Zosime atlantica, Z. incrassata, and Pe. reducta were described with five distal elements on A2 ENP2, but this is probably erroneous and requires confirmation. Acritozosime kept the plesiomorphic idyanthid armature complement of seven distal elements on A2 ENP2. Kim and Lee [1] believed that the presence of seven distal elements on A2 ENP2 is a plesiomorphy, and, therefore, ruled out the possibility of the extra seta being a novelty. This view is followed here.
- −
- Characters 15–19 (mandibular palp). The primitive mandibular basis of Idyanthidimorpha and Idyanthidae possesses four setae [11]. The family Zosimeidae underwent loss of one seta resulting in a trisetose basis. The primitive zosimeid trisetose basis is present in the typica-group of Zosime, and in Heterozosime. The mandibular basis of Acritozosime bears two setae and is regarded here as a potential autapomorphy for the genus. One seta is present in Peresime, and in the incrassata-group of Zosime, except for Z. carsteni. The latter kept the primitive trisetose condition. The reduced unisetose mandibular palp of Z. reyssi is unique in the family and is regarded here as autapomorphic for the species. The reduction from three to one seta seems to have occurred independently twice: the unisetose basis is a synapomorphy for Z. incrassata and Z. bathybia within Zosime and is an apomorphy for Peresime. The mandibular palp of Pseudozosime remains undescribed.
- The mandibular exopod is two-segmented in the ground pattern of Idyanthidimorpha and Idyanthidae, the proximal segment bearing four lateral—inner—setae, the distal segment with two apical setae [11]. The same architecture of that ramus is autapomorphic for Idyanthidae [11]. Both segments of the primitive idyanthidimorph mandibular exopod became fused and underwent loss of some setae, resulting in the—autapomorphic—one-segmented ramus in Zosimeidae, with three lateral—inner—setae and one distal element [11]. The one-segmented mandibular exopod with the primitive setal arrangement (three lateral and one distal element) is still present in Z. atlantica. The tetrasetose mandibular exopod is also present in P. reducta, Pe. pryocta sp. nov., A. tenuis, and A. sigsbeeiensis sp. nov. but the four exopodal setae of these species seem to have undergone secondary arrangement consisting of the displacement of the two distalmost lateral setae to the apical margin, giving the impression of one lateral—inner—and three distal setae. Almost all the species of the family underwent secondary loss of one seta, resulting in a trisetose ramus. The trisetose ramus is present in almost all species of Zosime, in Pe. abyssalis and Pe. pryorae, and Heterozosime. The bisetose mandibular exopod of Z. xiximi sp. nov. is autapomorphic for the species and is the most derived condition within the typica-group of Zosime. The knob-like, unisetose mandibular exopod in the incrassata-group was regarded as autapomorphic for that group by Pointner [13]. The one-segmented mandibular exopod with one seta is autapomorphic for Z. carsteni (incrassata-group of Zosime), the mandibular exopod absorbed into the basis and represented by one seta is a synapomorphy for Z. incrassata and Z. bathybia (incrassata-group of Zosime), and the reduced one-segmented mandibular palp of Z. reyssi with one seta only is autapomorphic for the species.The primitive idyanthidimorph and idyanthid mandibular endopod is one-segmented, with three lateral—inner—setae and three distal sets of three, two, and two setae fused basally [11]. Seifried [11] suggested that the tetrasetose mandibular endopod of Zosimeidae, with one lateral—inner—and three distal setae, is an autapomorphy for the family. However, the mandibular endopod of Peresime and Acritozosime bears one lateral and four distal elements, and the pentasetose mandibular endopod seems to be the autapomorphic condition for Zosimeidae. The—apomorphic—secondary reduction from four to three lateral setae is widespread in the family and seems to have occurred independently in Zosime (this condition is present in all species of the typica-group, and in Z. carsteni of the incrassata-group), and in Heterozosime. Two species of the incrassata-group of Zosime, Z. incrassata and Z. bathybia, underwent synapomorphic loss of two apical setae, resulting in a bisetose ramus, with one lateral and one distal seta. As noted above, the one-segmented unisetose mandibular palp is autapomorphic for Z. reyssi.
- −
- Character 20 (number of epipodal setae of the maxillulary coxa). The primitive idyanthidimorph maxillulary coxa is armed with two epipodal setae, and it is unisetose in Idyanthidae [11]. The primitive idyanthidimorph bisetose coxal epipodite is present in the ground pattern of Zosimeidae [11] and is still present in some species of Zosime (Z. anneae, Z. pacifica, Z. atlantica, and Z. valida, all of the typica-group of the genus). The secondary loss of one epipodal seta occurred independently in Zosime (this condition is present in the other species of the genus for which this appendage has been described; Z. reyssi was described without epipodal seta, but this requires confirmation), in two species of Peresime (Pe. abyssalis and Pe. pryocta sp. nov.), and in Heterozosime. The compete loss of the epipodal setae occurred independently in some species of Peresime (Pe. pryorae and Pe. reducta) and in Acritozosime. The bisetose condition of Z. anneae, Z. pacifica, Z. atlantica, and Z. valida is a plesiomorphy, and the unisetose condition of the other species of Zosime is regarded here as apomorphic within the genus. The unisetose condition of Pe. abyssalis and Pe. pryocta sp. nov. is primitive within that genus, and the unarmed coxal epipodite is a potential synapomorphy for Pe. pryorae and Pe. reducta. The complete loss of the epipodal armature in Acritozosime is regarded here as apomorphic for that genus.
- −
- Character 21 (number of segments of the maxillary endopod). Following Seifried [11], the ground pattern of the maxilla comprises a three-segmented endopod in Idyanthidimorpha and Idyanthidae, and a one-segmented endopod in Zosimeidae. However, some species of Zosime (Z. mexicana sp. nov., Z. xiximi sp. nov., Z. eliasi, Z. paramajor, Z. carsteni, and Z. bathybia; the condition of Z. incrassata is uncertain), H. tenuis, and both species of Acritozosime, display a two-segmented maxillary endopod. The secondary—apomorphic—fusion of both segments of the maxillary endopod seems to have occurred convergently in Zosime, Peresime, and Pseudozosime.
- −
- Characters 22–27 (maxilliped). Following Seifried [11], the ground pattern of the maxilliped in Idyanthidimorpha and Idyanthidae includes (i) the geniculation between syncoxa and basis and between basis and endopod; (ii) the syncoxa with two subdistal setae; (iii) the basis with one inner seta; and (iv) the two-segmented endopod with one seta and one thin and one large claw on proximal segment, and two outer small and two distal geniculate long setae on distal segment. The ground pattern of the maxilliped of Zosimeidae includes (i) the autapomorphic loss of geniculation between syncoxa and basis and between basis and endopod; (ii) the syncoxa with two subdistal setae; (iii) the basis with one inner seta; and (iv) the autapomorphic loss of the first endopodal segment and its armature, resulting in the one-segmented ramus with the primitive setal complement of two outer small elements and two geniculate long distal setae [11].
- The syncoxa is mostly unarmed in the family. Zosime eliasi is the only species of Zosime with one seta. One seta is also present in Pe. pryorae, Pe. pryocta sp. nov., and in A. sigsbeeiensis sp. nov. Acritozosime spinesco kept the full primitive armature complement of two setae. Kim and Lee [1] believed that the maxilliped of A. spinesco was the most plesiomorphic maxilliped in Zosimeidae and that it was similar to the idyanthid maxilliped in the geniculation between the syncoxa and basis and in the bisetose syncoxa and ruled out the possibility that the geniculation between the syncoxa and the basis and the two setae on the syncoxa were acquired secondarily. This view is followed here.The basis of the maxilliped bears one seta in most species of Zosime, except for Z. bathybia (incrassata-group) and Pe. pryocta sp. nov. with unarmed bases.The primitive tetrasetose maxillipedal endopod is present in most species of Zosime, in Pe. pryorae and Pe. pryocta sp. nov., in Heterozosime, and Acritozosime. Secondary reduction to three setae occurred independently in Pseudozosime, and in Zosime (Z. typica (typica-group, subordinate subgroup Aa), Z. eliasi and Z. bergensis (typica-group, subordinate subgroup Ac), Z. comata (typica-group, subordinate subgroup Bb), and in the two species of the incrassata-group of Zosime for which this appendage has been described, Z. carsteni, and Z. bathybia). The armature underwent reduction to two setae independently in Z. paratypica (typica-group of Zosime, subordinate subgroup Aa) and in Pe. reducta.
- −
- Character 28 (length:width ratio of caudal rami). Pointner [13] regarded the longer-than-wide caudal rami as (syn)apomorphic for Z. anneae and the typica-group. The longer-than-wide caudal rami is also present in Heterozosime and Acritozosime, and the plesiomorphic subquadrate caudal rami is present in the incrassata-group of Zosime, Peresime and Pseudozosime. The caudal rami seem to have undergone secondary elongation independently at least in the typica-group of Zosime, in Heterozosime, and in Acritozosime.
- −
- Characters 66–68 (ornamentation of outer spines of P1–P4 EXP). In her ground pattern of Zosimeidae, Seifried [11] noted the subdistal very long spinules on the outer and distal exopodal spines of P1. Pointner [13] noticed differences in the spinular ornamentation of the outer exopodal spines of P1 EXP of Zosime. Following Pointner [13], the typica-group of Zosime display the plesiomorphic outer exopodal spines of P1 EXP with distal and subdistal elongate spinules, and the incrassata-group possesses the derived outer spines of P1 EXP homogeneously pinnate (either with thin and pointed, or with thick and blunt spinules). The outer exopodal spines of P2–P4 show the derived condition in all species of Zosime, except for Z. gymnokosmosa in which the subdistal and distal spinules of the outer spines of P2–P4 EXP are somewhat longer than the proximal spinules (see Kim and Lee [27] (p. 229, Figure 13D; p. 231, Figure 15A–C). The outer spines of P1 EXP of Z. tunnelli display a mix of both conditions: the outer distal spine shows the plesiomorphic spinules, but the other spines of that ramus show the derived condition (see Yeom et al. [2] (p. 15, Figure 10A)). The plesiomorphic condition of P1 EXP is also present in Peresime, Pseudozosime, Heterozosime, and Acritozosime. The outer exopodal spines of P1 and P2 of Pe. reducta display the plesiomorphic condition, but the outer spines of P3 and P4 show the derived condition. Heterozosime shows the derived condition in P2–P4; Acritozosime displays the plesiomorphic condition.
- −
- Character 69 (number and length of the distal armature complements of the last segment of P4 ENP). Three distal setae on the last endopodal segment of P4 are present in all species of Zosime, except for Z. tunnelli, Z. gymnokosmosa, and Z. comata. Three distal setae are also present in H. tenuis and H. klemie sp. nov. The middle distal seta of P4 ENP3 is reduced to a very short seta in Z. mexicana sp. nov. (see Figure 13B, this study), Z. montagni (described as a tube-pore in Yeom et al. [2] (p. 8, Figure 5D)), and Z. pacifica (see Fiers [28] (p. 18–19, Figure 3d)). A similar reduction is present in H. tenuis and seems to have evolved independently. The reduction in the middle seta of P4 ENP3 is regarded here as synapomorphic for Z. mexicana sp. nov., Z. montagni, and Z. pacifica within Zosime, and apomorphic for H. tenuis against the primitive well-developed seta of H. klemie sp. nov. Zosime tunnelli and Z. gymnokosmosa share the—synapomorphic—loss of one distal seta of P4 ENP3; Z. comata underwent further loss of one more distal seta, for which one distal seta on P4 ENP3 is regarded here as autapomorphic within Zosime. The—apomorphic—loss of one distal seta on the last endopodal segment occurred also in the two-segmented endopod of Pseudozosime and Acritozosime. The most derived state amongst the genera of Zosimeidae is that of Peresime, in which the distal segment of the two-segmented endopod of P4 lost two setae, resulting in an unisetose distal margin of P4 ENP2; the latter is regarded here as autapomorphic for Peresime.
- −
- Characters 70–74 (P5). Seifried [11] showed that the ground pattern of P5 of Zosimeidae includes (i) separated baseoendopods in the females, but fused medially in the males; (ii) P5 EXP separated from the baseoendopod in both sexes; (iii) four setae on the female and male P5 EXP; and (iv) four setae on the endopodal lobe of females, but two setae in males.
- Both baseoendopods of the female P5 are separated throughout the family but became fused medially in Z. xiximi sp. nov. and Z. thistlei. The baseoendopods of both legs are fused medially in the males of most species of the family; both baseoendopods have been described as separate in the males of Z. pacifica, and Z. typica.Acritozosime sigsbeeiensis sp. nov. is the only species known to date in which both the female and male P5 EXP and BENP are separated; P5 EXP and BENP are also separated in the female of A. spinesco, but the male of the species has not been discovered yet. The female P5 EXP and BENP of two species of the incrassata-group of Zosime, Z. incrassata and Z. bathybia, are separated (the males of these species remain unknown). The exopod and baseoendopod of the female P5 of another species of the incrassata-group, Z. carsteni, are imperfectly separated and an anterior suture is still visible; P5 EXP and BENP are completely fused in the male. The exopod and baseoendopod of P5 are separated in the male of Z. pacifica but completely fused in the female. In this regard, Fiers [28] noted that the separation of both rami is most probably suppressed from early copepodid stages onwards in the females of most zosimeids. He [28] observed that the P5 of Z. pacifica appears at the copepodid III as a bud with three setae. The final armature complement of P5 in both the female and male (exopod with three marginal and one surface seta; endopod with two setae) appears at the copepodid IV and both rami undergo further subtle changes in length:width ratio during the molt to copepodid V. The exopod and baseoendopod remain fused in the adult female, but the exopod becomes separated from the baseoendopod in the male during the last molt from copepodid V to copepodid VI (adult). The female P5 EXP and BENP are fused in all other genera and species of the family; the same applies for the males of the species for which they have been described.The pair of fifth legs of the males of all genera and species for which the male is known are fused to the supporting somite. However, both P5 have been illustrated as being separate from the somite in the males of Z. typica and Z. montagnai; this requires confirmation.All the species of Zosime (except for Z. thistlei), and Heterozosime possess the full primitive armature complement of four setae on the female P5 EXP. Loss of one seta of the female P5 EXP—most probably the proximal outer element—occurred convergently in Z. thistlei (which is regarded as autapomorphic within Zosime), in two species of Peresime, Pe. pryorae and Pe. reducta (which is regarded here as a potential synapomorphy), and in the two species of Acritozosime. Further—potentially synapomorphic—reduction to two setae (most probably the proximal outer seta and one marginal element) on the female P5 EXP occurred in Pe. abyssalis and Pe. pryocta sp. nov. The condition of the female P5 EXP of P. browni Scott T., 1912 is uncertain. Scott T. [5] described the female P5 EXP with three marginal setae, but a small outer proximal bristle is present in his figure of the female P5 [5] (plate VIII, Figure 19). The outer proximal seta of the female P5 EXP of most species of Zosime with a tetrasetose ramus is displaced to the anterior surface of the ramus, resulting in three marginal setae and one proximal outer surface element. The four marginal setae on the female P5 EXP of Z. reyssi, Z. incrassata, Z bathybia, Z. carsteni, and Z. montagnai could be due to observational imprecisions, and the displacement of the proximal outer seta of the P5 EXP to a surface position on that ramus is a potential autapomorphy for Zosimeidae (see below).The tetrasetose male P5 EXP is present in most species of Zosime, and in the male of H. tenuis (the male of H. klemie remains undescribed). Secondary loss of one seta (most probably the proximal outer element) occurred in the male of two species of Zosime, Z. gisleni (the female P5 EXP possesses three marginal setae and one proximal outer surface seta) and Z. thistlei (the loss of one seta occurred also in the female), and in the male of A. sigsbeeiensis sp. nov. (the female P5 EXP is also trisetose). Further reduction to two setae on the male P5 EXP occurred in Pe. abyssalis (two setae are present also in the female of the species; the female of Pe. pryocta sp. nov. possess a bisetose P5 EXP, but the male of the species has not been described). Zosime gisleni seems to be unique in that the armature complement of the female and male P5 EXP is different, i.e., the female possesses three marginal setae and one proximal outer surface element, but the male possesses only three marginal setae. The proximal outer element of the male P5 EXP of Z. gisleni was most probably suppressed during the development from copepodid to the adult phase.The primitive tetrasetose baseoendopod of the female P5 is present in several species of Zosime (in two out of eight species of subordinate subgroup Aa (Z. anneae and Z. major), in all the species of subordinate subgroup Ab (Z. atlantica, Z. mediterranea, and Z. xiximi sp. nov.), in three out of four species of subordinate subgroup Ac (Z. eliasi, Z. tunnelli, Z. thistlei), in all the species of subordinate subgroup Bb (Z. paramajor, Z. erythraea, Z. comata, Z. bathyalis, and Z. gymnokosmosa), in two out of four species of the incrassata-group (Z. carsteni and Z. reyssi)), in P. browni, and in H. klemie sp. nov. The—secondary—loss of one seta resulting in a trisetose endopodal lobe occurred in Zosime (Z. gisleni and Z. typica (subordinate subgroup Aa), Z. bergensis (subordinate subgroup Ac), and Z. incrassata and Z. bathybia (incrassata-group)), in the four species of Peresime and in the two species of Acritozosime. Further—apomorphic—reduction to two setae occurred in Zosime (three species of subordinate subgroup Aa (Z. mexicana sp. nov., Z. montagni, and Z. pacifica), in all the species of subordinate subgroup Ba (Z. valida, Z. destituta, and Z. changi) for which is a potential synapomorphy), and in H. tenuis.The male P5 endopodal lobe bears the primitive armature complement of two setae in all genera and species of the family, except for Z. gymnokosmosa with one seta only. Kim and Lee [27] regarded the unisetose endopodal lobe of the male P5 of Z. gymnokosmosa as autapomorphic for the species.
- −
- Characters 86–88 (male P6). The ground pattern of the male P6 of Idyanthidae and Zosimeidae consists of three setae [11]. The innermost seta is very short in all the species within the family, but some differences were observed between the outer—basal—and the apical element. The outer—basal—and the apical—middle—element of Z. montagnai were described (they were not illustrated) as spiniform in Yeom et al. [2] (p. 9). Fiers [28] (p. 19) described the outer basal and the apical element of the male P6 of Z. pacifica as spinulose setae, but these elements look more like spiniform in his figure [28] (p. 20, Figure 4e). Pointner [13] (p. 2314) described the outer element of the male P6 of Z. eliasi as a plumose flexible seta, and the apical element as a bipinnate seta, but the latter was shown as a bipinnate spine in her figure [13] (p. 2313, Figure 20C). Due to these inconsistencies, these species were excluded from the present analysis. The primitive condition with setiform outer and apical elements on the male P6 is still present in Pe. abyssalis (the only species of that genus for which the male has been described), Z. thistlei, Z. carsteni, Z. valida, Z. gymnokosmosa, and Acritozosime sigsbeeiensis sp. nov. (the only species of that genus for which the male has been described). The most primitive condition amongst the above species is present in Pe. abyssalis, Z. thistlei, and Z. carsteni in which the outer and apical setiform elements are subequal in length, or the outer element slightly shorter than the apical seta. The apical seta underwent reduction in Z. valida and Z. gymnokosmosa. The outer seta of A. sigsbeeiensis underwent secondary reduction (it is half as long as the apical seta) and is regarded here as autapomorphic for the species. Heterozosime tenuis is unique within the family in the—potentially autapomorphic—transformation of the outer seta into a pinnate spine. The—apomorphic—transformation of the apical seta into a spine occurred convergently in some species of Zosime (Z. anneae, Z. xiximi sp. nov., and Z. changi), and in H. tenuis.
Whereas DNA-based phylogenetic studies are necessary to corroborate the relationships amongst the genera of Zosimeidae and their species, this study aims to reveal monophyletic clades using a morphological approach. Despite the inherent limitations due to homoplastic morphological characters, this study shed some light on the diversity of Zosimeidae in a phylogenetic context. The DNA sequences of zosimeids available in the National Center for Biotechnology Information (NCBI) database are still scarce. Only five individuals of Z. atlantica (COI), fifteen individuals of Z. cf. atlantica (18S and CytB), and one individual from a non-identified species (COI, 28S, 18S, H3) are available, making futile any attempt to study the phylogeny of the family from a molecular perspective.
4.2. The Family Zosimeidae
The taxonomic history of the family Zosimeidae has been dealt with in Huys and Clark [29]. Currently, the family Zosimeidae is composed of five genera, Acritozosime, Heterozosime, Peresime, Pseudozosime, and Zosime, the latter being the most species-rich with 27 taxa, the new species presented herein included. Seifried [11] (p. 100) gave a list of hypothetic autapomorphies for Zosimeidae. These are summarized below, with the ground pattern between square brackets when necessary for the upcoming discussion:
- (i).
- armature of A2 ENP2 composed of three lateral elements [two spines—spine I proximal to spine III (see Seifried [11] (p. 100, Figure 31)—and one tiny seta—seta 4 in Seifried [11] (p. 101, Figure 31)—close to spine III], and six distal setae
- (ii).
- loss of the lateral idyanthidimorph seta 2 of A2 ENP2 (see Seifried [11] (p. 89, Figure 26; p. 94, Figure 28))
- (iii).
- distal margin of the antennary endopod no beveled
- (iv).
- setae of the mandibular basis inserting on a bulge of inner border
- (v).
- mandibular endopod with one lateral and three apical setae [mandibular endopod one-segmented]
- (vi).
- mandibular exopod with three lateral and one distal seta [mandibular exopod one-segmented]
- (vii).
- (viii).
- exopod of the maxillule very short [with three setae]
- (ix).
- endites of the maxillary syncoxa inserted on distal half of syncoxa
- (x).
- endite of the (allo)basis of maxilla poorly developed
- (xi).
- maxillary endopod one-segmented with 5 setae
- (xii).
- syncoxa of maxilliped subquadrate, at most 1.5 times as long as wide [syncoxa with two setae]
- (xiii).
- maxilliped straight (stenopodial, without geniculation or flexure point between syncoxa and basis, and between basis and endopod)
- (xiv).
- endopod of the maxilliped one-segmented and directed outwardly; the idyanthidimorph first endopodal segment and its armature lost [endopod with two small outer and two geniculate distal setae]
- (xv).
- P1 ENP1 subquadrate; P1 ENP2 and ENP3 fused, with two inner setae and two terminal spines
- (xvi).
- outer basal seta of female and male P5 arising from long setophore
- (xvii).
- female P5 EXP and endopodal lobe separated by incision [endopodal lobe with two inner and two distal setae; exopod with four setae]
- (xviii).
- male P5 BENP very constricted [baseoendopods fused medially]
- (xix).
- male P5 endopodal lobe very small [with two small setae; exopod one-segmented with four setae (1,2,1)]
However, some zosimeid genera depart from the autapomorphic general scheme as described by Seifried [11] both in her list of autapomorphies and in the description of the ground pattern of the family.
Following Seifried [11], the lateral armature complement of A2 ENP2—one proximal short spine (element I in Seifried [11]), one subdistal longer spine (element III in Seifried [11]), and one tiny seta (element 4 in Seifried [11])—resulted from the autapomorphic loss of the small seta 2 (see Seifried [11] (Figure 28)) present in the ground pattern of Idyanthidae, the sister taxon of Zosimeidae. Some species of Zosime (Z. anneae, Z. montagnai, Z. eliasi, Z. tunnelli, Z. valida, and Z. incrassata) were described with only two lateral spines on the second segment of the antennary endopod. It is uncertain whether the small slender seta 4 commonly found close to the base of spine III underwent secondary loss or if it was overlooked, since it can be very small and is often masked by spine III. Moreover, the lateral armature of A2 ENP2 of some Zosimeidae, i.e., Peresime and Acritozosime, is composed of one proximal spine (presumably Seifried’s [11] spine I), one spine distal to spine I (presumably Seifried’s [11] spine III), and a well-developed spinulose seta comparable in length to the other two spines. Given the position of the latter element in Peresime and Acritozosime, it seems reasonable to suppose that it is homologous to the small slender seta 4 (see Seifried [11] (p. 100, Figure 31)) present in other zosimeids. However, it is not clear whether the long lateral seta 4 of A2 ENP2 present in Peresime and Acritozosime corresponds to the plesiomorphic long lateral seta 4 of Idyanthidae, or if the reduced small seta 4 present in most zosimeids underwent secondary—apomorphic—elongation in Peresime and Acritozosime resembling the plesiomorphic idyanthid long seta 4. In our opinion, the lateral long seta 4 of A2 ENP2 present in Peresime and Acritozosime is a plesiomorphy, and the secondary—apomorphic—reduction in the lateral element 4 of A2 ENP2 occurred independently in Zosime, Pseudozosime, and Heterozosime.
Seifried [11] noted that the primitive zosimeid mandibular exopod is composed of four setae (three lateral—inner, one distal seta). The tetrasetose primitive condition of the mandibular exopod is present in Z. atlantica, P. reducta, Pe. pryocta sp. nov., and A. tenuis, and A. sigsbeeiensis sp. nov. Zosime atlantica seems to have kept the primitive setal arrangement with three lateral—inner—and one distal seta. On the other hand, the four exopodal setae of P. reducta, Pe. pryocta sp. nov., A. tenuis, and A. sigsbeeiensis sp. nov. seem to have undergone secondary arrangement consisting of the displacement of the two distalmost lateral setae to the apical margin, giving the impression of one lateral—inner—and three distal setae. Almost all the species of the family underwent secondary loss of one seta, resulting in a trisetose ramus. The bisetose mandibular exopod of Z. xiximi sp. nov. is autapomorphic for the species and is the most derived condition within the typica-group. The one-segmented mandibular exopod with one seta is autapomorphic for Z. carsteni. The mandibular exopod absorbed into the basis and represented by one seta is a synapomorphy for Z. incrassata and Z. bathybia. The reduced one-segmented mandibular palp of Z. reyssi with one seta only is autapomorphic for the species.
Following Seifried [11], the primitive zosimeid mandibular endopod possesses four setae (one lateral—inner—and three distal setae). The pentasetose mandibular endopod of Peresime and Acritozosime (with one lateral—inner—and four distal setae) do not fit the list of autapomorphies and ground patterns for Zosimeidae and is regarded here as plesiomorphic. Another, less probable, alternative is that the extra seta on the mandibular endopod of Peresime and Acritozosime constitutes a novelty. The tetrasetose condition (with one lateral and three distal setae) of the mandibular endopod is comparatively more derived and is present in almost all the species of the family. The most derived condition is present in some species of the incrassata-group of Zosime. The mandibular endopod of Z. incrassata and Z. bathybia possesses two setae only (one lateral, one distal) and this condition is synapomorphic for these two species.
The two-segmented maxillary endopod in some species of Zosime, and in Heterozosime and Acritozosime, also do not fit the list of autapomorphies and is regarded here as symplesiomorphic. The alternative scenario where the one-segmented endopod of the maxilla underwent secondary—apomorphic—addition of one segment is less probable.
The geniculate maxillipeds of A. spinesco do not correspond to the list of autapomorphies for the family. The alternative scenario where the maxilliped of A. spinesco develops secondarily a geniculation between the syncoxa and the basis, and probably between the basis and the endopod, seems less probable. Therefore, the geniculate maxilliped of A. spinesco is regarded here as a plesiomorphy, and the straight—stenopodial—maxilliped, widely distributed in Zosimeidae, seems to be an apomorphy within the family.
Apart from the lateral armature of the second antennary exopod, the armature complement of the mandibular endopod, and the two-segmented maxillary endopod (see also Kim and Lee [1] for an in-depth analysis on Heterozosime and Acritozosime), the new species presented in this study match the autapomorphies of Zosimeidae, and were allocated to that family.
4.3. Key to the Genera of Zosimeidae
- 1a. P1 EXP three-segmented………………………………………………………………………………………………………………………………………………………………………………………………2
- 1b. P1 EXP two-segmented………………………………………………………………………………………………………………………………………………………………………………………………..4
- 2a. Female A1 six- to eight-segmented; P2 ENP three-segmented……………………………………………………………………………………………………………………………………………………3
- 2b. Female A1 five-segmented; P2 ENP two-segmented…………………………………………………………………………………………………………………………………………………Pseudozosime
- 3a. P3 ENP three-segmented………………………………………………………………………………………………………………………………………………………………………………………. Zosime
- 3b. P3–P4 ENP two-segmented…………………………………………………………………………………………………………………………………………………………………………………...Peresime
- 4a. Female A1 six-segmented; A2 EXP1 and EXP2 with one seta; A2 ENP2 with six distal elements; mandibular basis, EXP and ENP with 3,3,4 setae, respectively; maxillule with epipodal seta; syncoxa of maxilliped unarmed; P1 EXP2 with seven setae in all; P1 ENP1 with one seta, ENP2 with inner lateral seta and two or three distal elements; P2 EXP3 with two inner lateral setae; P2–P4 ENP three-segmented; female P5 EXP with three marginal elements and one outer surface seta; female P5 EXP and BENP fused…………………………………………………………………………………………………………………………………………………………………………………………………………….Heterozosime
- 4b. Female A1 seven-segmented; A2 EXP1 with two setae, EXP2 unarmed; A2 ENP2 with seven distal elements; mandibular basis, EXP and ENP with 2,4,5 setae, respectively; maxillule without epipodal seta; syncoxa of maxilliped armed; P1 EXP2 with six setae in all; P1 ENP1 unarmed, ENP2 without inner lateral seta and two distal elements; P2 EXP3 with one inner lateral seta; P2–P4 ENP two-segmented; female P5 EXP with three marginal elements (without outer surface seta); female P5 EXP and BENP separated………………………………………………………………………………………………………………………………………………………………………………………………………..Acritozosime
4.4. The Genus Zosime
The taxonomic history of the genus Zosime, the type genus of the family Zosimeidae, has been dealt with in Kim and Lee [27] (p. 210). The genus is distributed worldwide and shows an ample eurybathic distribution from shallow systems (<200 m depth) to the deep sea (>200 m depth) [2,6,7,8,9]. Recent efforts have yielded a significative increase in the number of species of Zosime from distant localities, i.e., Korea [12,27] and the Gulf of Mexico [2], and many more species from several places around the globe await to be described or their identities confirmed [6,7,8,9,11,30,31,32,33,34].
Despite some potential autapomorphies for Zosime detected by Koller and George [7], e.g., displacement of the outermost seta of the P5 EXP to the anterior surface of the ramus, and the elongate caudal rami, no real autapomorphies have been detected to date for the genus, and its monophyly is far from resolved. In this regard, Koller and George [7] noticed, for example, that Dinet [3] described the female P5 EXP of Z. reyssi (the males of this and many other species of Zosime remain unknown) with four marginal setae (the outermost proximal seta not displaced to the anterior surface of the exopod) both in his Figure 8 [3] (p. 565) and in the written description [3] (p. 566). The same seems to be the case for Z. incrassata and Z bathybia (the exopod and baseoendopod of the female P5 are separated in the latter two species), Z. montagnai, and Z. carsteni. The female P5 EXP of Z. montagnai was described with four marginal setae both in the written description [2] (p. 8) and in the respective figure [2] (p. 6, Figure 5E). Interestingly, the male of Z. montagnai was described with three marginal setae and one outer surface seta on the P5 EXP, both in the written description [2] (p. 9) and in the respective figure [2] (p. 6, Figure 5H). Similarly, the female P5 EXP of Z. tunnelli was shown with four marginal setae in Yeom et al.’s [2] Figure 10E (p. 15), but the ramus was described with three marginal setae and one outer surface element in the written description [2] (p. 17). In our opinion, the four marginal setae on the female P5 of Z. reyssi, Z. incrassata, Z bathybia, Z. carsteni, and Z. montagnai could be due to observational imprecisions and need to be verified. For example, the outermost seta of the female P5 EXP of Z. bathybia and Z. paratypica arises from a short protrusion or short setophore on the anterior surface of the ramus (see Bodin [24] (p. 44, plate XXIV) and Becker and Schriever [4] (p. 43, Figure 5)) but the distal margin of the setophore is aligned to the outer margin of the exopod giving the—false—impression that the seta arises from the outer margin of the ramus. Similarly, Pointner [13] described the female P5 of Z. carsteni with four setae on the exopod, the outermost proximal seta arising from a small lateral projection that seems to arise on the anterior surface of the ramus close to the outer margin; the male was described with three marginal and one outer surface seta. Whatever the case might be, three marginal setae and one outer surface seta in Zosime, on the female and male of Heterozosime and probably in the monotypic genus Pseudozosime (the condition of Ps. browni is inconclusive because the female P5 was described with three marginal setae only [5] (p. 541), but was shown with an additional outer proximal bristle [5] (plate VIII, Figure 19)), indicates that the displacement of the proximal outer seta of the P5 EXP to a surface position on that ramus is a potential autapomorphy for Zosimeidae (this condition is not present in its sister taxon Idyanthidae). The subsequent loss of the outer surface seta is regarded here as secondary and has occurred several times within the family.
The female and male P5 EXP of Z. thistlei is fused to the baseoendopod and possesses three marginal setae only [2]. The P5 EXP and BENP are fused in Peresime; Pe. pryorae and Pe. reducta share the female EXP with three marginal setae only; Pe. abyssalis and Pe. pryocta sp. nov. underwent additional loss of one distal seta of the female P5 EXP. Acritozosime shows the male and female P5 EXP and BENP separated, with three marginal setae only. The loss of the outer–surface–seta seems to have occurred independently at least three times in Zosimeidae, i.e., in Z. thistlei, Peresime, and Acritozosime, and is regarded here as apomorphic for each of them.
As an attempt to better understand the phylogenetic relationships between the species of the genus, Kim et al. [12] proposed three species groups within Zosime, the anneae-, incrassata- and typica-groups. They [12] proposed these species groups based upon several features in the females: segmentation of the antennule, armature formula of the antennary exopod, number of setae on the mandibular rami, armature of P3–P5, and length:width ratio of the caudal rami. The anneae-group included only Z. anneae; the incrassata-group included Z. bathybia, Z. incrassata, and Z. reyssi; the typica-group included Z. atlantica, Z. bathyalis, Z. bergensis, Z. destituta, Z. erythraea, Z. gisleni, Z. major, Z. mediterranea, Z. pacifica, Z. paramajor, Z. paratypica, Z. typica, and Z. valida. Additionally, they [12] foresaw the eventual subdivision of the typica-group based on differences in the armature complement of the mandibular rami, antennary exopod, and armature of swimming legs.
Pointner [13] suggested the mandibular endopod armed with one inner lateral, and three apical setae at most, as a true autapomorphy for Zosime to support its monophyly, but recent findings by Kim and Lee [1] and the results presented here show that a similar condition of the mandibular endopod is present in Heterozosime (see below). The apomorphic tetrasetose mandibular endopod might have evolved independently in Zosime and Heterozosime but underwent additional—synapomorphic—loss of two setae in Z. incrassata and Z. bathybia (incrassata-group of Zosime). The one-segmented unisetose mandibular palp of Z. reyssi (incrassata-group of Zosime) is regarded here as autapomorphic for the species. She [13] agreed with Kim et al.’s [12] typica- and incrassata-groups, to which she added Z. eliasi and Z. carsteni, respectively, and suggested the reduction in the mandibular exopod to a knob-like structure and its reduction in armature as an autapomorphy for the incrassata-group, against the plesiomorphic one-segmented, bilobate, and comparatively well-developed ramus with a lateral seta and a variable number of other setae of the typica-group. Pointner [13] also noticed differences between the typica- and incrassata-groups in (i) the spinular ornamentation of the outer exopodal spines of P1 EXP, i.e., with plesiomorphic, distal and subdistal elongate spinules on the outer exopodal spines of P1 in the typica-group versus the more derived, homogeneously pinnate (either with thin and pointed, or with thick and blunt spinules) in the incrassata-group; and (ii) length:width ratio of P2–P4 ENP1 and ENP2 (compact and square in the incrassata-group, and in Z. thistlei (the latter of the typica-group), but longer than wide in the typica-group (except for Z. thistlei). On the other hand, she [13] questioned the separation of the anneae- (Z. anneae) and the typica-group, since they share (i) the presence of a seta on the second exopodal segment of the antenna; (ii) the exopod and endopod of the mandible with three and four setae, respectively; (iii) the presence of two inner setae on P3–P4 EXP3; (iv) the P5 EXP with three apical and one surface seta (but see above some additional comments on the differences in shape and armature complement between the male and female of some species of Zosime) and P5 EXP and BENP fused; and (v) the longer-than-wide caudal rami. Pointner [13] regarded the latter two as (syn)apomorphies for Z. anneae and the typica-group versus the plesiomorphic subquadrate caudal rami in the incrassata-group. Additionally, she [13] regarded the separated rami (exopod and baseoendopod) of the female P5 in the incrassata-group as plesiomorphic versus the derived fused rami of the typica-group. As a result of her analysis, Pointner [13] (p. 2324) incorporated Z. anneae into the typica-group and amended Kim et al.’s [12] diagnosis of this species group. Later, Kim and Lee [27] (p. 235) expanded the diagnosis of the typica-group—length:width ratio of the female caudal rami 1.6–4.3—to include Z. changi. At the time of publication of Kim and Lee’s [27] study, the typica-group was defined by the combination of the following characters: (i) the six- to eight-segmented female antennule; (ii) the presence of one seta on the second segment of the antennary exopod; (iii) presence of three and four setae on the mandibular exopod and endopod, respectively; (iv) presence of two inner setae on P3–P4 EXP3; (v) armature complement of the female P5 exopod consisting of three marginal and one surface seta; and (vi) length:width ratio of caudal rami from 1.6 to 4.3; and the species groups of Zosime were composed as follows:
typica-group: Z. anneae, Z. atlantica, Z. bathyalis, Z. bergensis, Z. comata, Z. changi, Z. destituta, Z. eliasi, Z. erythraea, Z. gisleni, Z. gymnokosmosa, Z. major, Z. mediterranea, Z. montagnai, Z. pacifica, Z. paramajor, Z. paratypica, Z. thistlei, Z. tunnelli, Z. typica, and Z. valida.
incrassata-group: Z. bathybia, Z. carsteni, Z. incrassata, and Z. reyssi.
Zosime xiximi sp. nov. and Z. mexicana sp. nov. are here attributed to the typica-group, but the armature complement of the mandibular exopod of Z. xiximi sp. nov. underwent secondary reduction in the armature complement to two setae, and Z. atlantica kept the full, primitive armature complement of the mandibular exopod with four setae (three lateral, one distal), and the typica-group is herein redefined by the combination of several characters as follows: (i) female antennule six- to eight-segmented; (ii) second segment of antennary exopod with one seta; (iii) mandibular exopod and endopod with two–four, and four setae, respectively; (iv) P1 EXP with outer spines ornamented with distal and subdistal elongate spinules; (v) P3–P4 EXP3 with two inner setae; (vi) female P5 exopod with three marginal and one surface seta; (vii) caudal rami 1.6–4.3 times as long as wide.
We aimed at testing the naturalness of Kim et al.’s [12] and Pointner’s [13] groupings and at proposing further natural subgroups of the typica-group through a sound phylogenetic analysis of the species of the genera of the family Zosimeidae, using the ground pattern of Idyanthidae and Zosimeidae, three species of Tachidiella Sars G. O., 1909, and S. gladiator as outgroups. The results of our analyses are presented below and are illustrated in Figure 32.
Two main subgroups of the typica-group exist in terms of the armature complement of P1 EXP as the primary discriminant. Some other subordinate subgroups were identified within the main subgroups of the typica-group. These subgroups and their subordinate subgroups are defined below.
Typica-group, subgroup A. The larger subgroup (hereafter referred to as subgroup A; Figure 32) exhibits the relatively plesiomorphic armature complement of P1 EXP3 with six elements in all—one inner seta, two distal elements, and tree outer spines (the same is present in Z. carsteni of the incrassata-group of Zosime, and in Peresime; three outer spines are also present in the distal exopodal segment of Heterozosime and Acritozosime, but the exopod of these genera is two-segmented, and the outer armature complement of EXP3 (three spines) might be the result of the fusion of EXP2 with one outer spine and EXP3 with two outer spines, or fusion of EXP2 and EXP3 with one and three outer spines, respectively, and secondary loss of one of them). To subgroup A belong Z. xiximi sp. nov., Z. mediterranea, Z. atlantica, Z. anneae, Z major, Z. typica, Z. gisleni, Z. pacifica, Z. montagnai, Z. paratypica, Z. mexicana sp. nov., Z. thistlei, Z. tunnelli, Z. eliasi, and Z. bergensis.
Subordinate subgroup Aa. A subordinate subgroup of species of subgroup A (hereafter referred to as subordinate subgroup Aa; Figure 32) is composed of Z. anneae, Z major, Z. typica, Z. gisleni, Z. pacifica, Z. montagnai, Z. paratypica, and Z. mexicana sp. nov., and share the armature formulae of P1–P4 EXP/ENP (P1: 0,1,123/1,211; P2: 1,1,223/1,1,211; P3: 1,1,223/1,1,211; P4: 1,1,223/1,1,211).
Subordinate subgroup Ab. Another subordinate subgroup of subgroup A (hereafter referred to as subordinate subgroup Ab; Figure 32) is composed of Z. xiximi sp. nov., Z. mediterranea, and Z. atlantica. The species of this subordinate subgroup lost the lateroproximal inner seta of P1 ENP2, resulting in the armature formula 1,111. The armature formulae of P2–P4 of this subordinate subgroup is as in subordinate subgroup Aa, but the loss of the lateroproximal inner seta of P1 ENP2 in Z. atlantica, Z. mediterranea, and Z. xiximi sp. nov. is regarded here as derived for these species within subgroup A. The loss of the lateroproximal inner seta of P1 ENP2 occurs also in subordinate subgroup Ba. The armature formulae of P2–P4 is also like that in subordinate subgroup Ba, except for the loss of one outer spine of P2–P4 EXP3 in Z. changi, and the loss of the lateroproximal inner seta of P4 ENP3 in Z. destituta and Z. changi (the armature formulae of P1 ENP, and P2–P4 of subordinate subgroup Ab is identical to that of Z. valida).
Subordinate subgroup Ac. There is a third subordinate subgroup within subgroup A (hereafter referred to as subordinate subgroup Ac; Figure 32). It is composed of Z. thistlei, Z. tunnelli, Z. eliasi, and Z. bergensis. Subordinate subgroup Ac kept the lateroproximal inner seta of P1 ENP2 but lost one of the distal elements of this segment—most probably the distal inner plumose seta. The additional loss of the inner seta of P1 ENP1, and the lateroproximal inner seta of P2 ENP3 and one distal seta of P4 ENP3—most probably the middle seta—occurred in Z. thistlei and Z. tunnelli, respectively. The loss of the inner seta of P1 ENP1 in Z. thistlei is regarded here as apomorphic within Zosime and occurred independently in Acritozosime.
Typica-group, subgroup B. Subgroup B underwent loss of one outer spine of P1 EXP3, resulting in the armature formula 122—one inner and two distal elements, and two outer spines. Two outer spines are also present in Pseudozosime and in all the species of the incrassata-group of Zosime except for Z. carsteni. To subgroup B belong Z. destituta, Z changi, Z. valida, Z. erythraea, Z. paramajor, Z. bathyalis, Z. gymnokosmosa, and Z. comata (Figure 32).
Subordinate subgroup Ba. A subordinate subgroup of species of subgroup B (hereafter referred to as subordinate subgroup Ba; Figure 32) is composed of Z. destituta, Z changi, and Z. valida. These three species underwent loss of the lateroproximal inner seta of P1 ENP2 resulting in the armature formula of P1 ENP 1,111, like in subordinate subgroup Ab (Z. xiximi sp. nov., Z. mediterranea, and Z. atlantica). The loss of the lateroproximal inner seta of P1 ENP2 is regarded here as derived for Z. valida, Z. destituta, and Z. changi (subordinate subgroup Ba) within subgroup B. The armature formula of P2–P4 is like that in subordinate subgroup Aa except for the loss of one outer spine of P2–P4 EXP3 in Z. changi, and the inner lateral seta of P4 ENP3 in Z. destituta and Z. changi.
Subordinate subgroup Bb. Another subordinate subgroup of subgroup B (hereafter referred to as subordinate subgroup Bb; Figure 32) is composed of Z. erythraea, Z. paramajor, Z. bathyalis, Z. gymnokosmosa, and Z. comata. Zosime erythraea is unique within Zosime in the—autapomorphic—loss of the inner seta of P1 EXP2. The species of this subordinate subgroup are similar to those of subordinate subgroup Ac in the loss of one distal element of P1 ENP2 (see above) and seem to have occurred independently at least three times in Zosime (in subordinate subgroup Ac, in subordinate subgroup Bb, and in Z. reyssi of the incrassata-group). Zosime paramajor and Z. erythraea kept the armature complement of P2–P4 EXP/ENP (P2: 1,1,223/1,1,211; P3: 1,1,223/1,1,211; P4: 1,1,223/1,1,211). Zosime comata underwent loss of one outer spine in P2–P4 EXP3, and two setae of the P4 ENP3. The latter was regarded as autapomorphic for Z. comata in Kim and Lee [27], and a similar loss occurred convergently in Peresime. Zosime bathyalis and Z. gymnokosmosa also lost one outer spine of P2–P4 EXP3, but the former underwent additional loss of the lateroproximal inner seta of P2–P4 ENP3. Zosime gymnokosmosa kept the lateroproximal inner seta of P2–P4 ENP3 but underwent additional loss of one apical seta of the same segments.
4.5. Key to the Species of Zosime
- 1a. A2 EXP2 unarmed (except for Z carsteni with one seta), EXP3 without lateral seta (except for Z. carsteni with one lateral element); mandibular endopod with less than four setae (except for Z. carsteni with four setae); caudal rami short (length:width ratio < 1.6); outer spines of exopodal segments with homogeneous small spinules ………………………………….. 2 incrassata group
- 1b. A2 EXP2 always with one seta, EXP3 with one lateral seta and two or three distal elements (except for Z. atlantica and Z. xiximi sp. nov. without lateral seta); mandibular endopod with four setae; caudal rami elongate (length:width ratio > 1.6); at least one outer spine of exopodal segments with heterogeneous subdistal long spinules………………………………. 5 typica group
- 2a. Armature formula of A2 EXP 1,1,3 (EXP3 with one lateral and two distal elements); mandibular palp with trisetose basis, unisetose small, one-segmented exopod, and tetrasetose one-segmented endopod; female P5 EXP and BENP fused but with anterior suture between BENP and EXP, ENP with four setae………………………………………………………………….Z. carsteni
- 2b. Armature formula of A2 EXP 1,0,2–3 (EXP3 without lateral seta and with two or three distal elements); mandibular palp either with unisetose basis, exopod incorporated to basis and represented by one seta, and endopod with two setae (palp with less than four setae in all), or palp small, one-segmented with one seta; female P5 EXP and BENP fused or separated, ENP with three or four setae…………………………………………………………………………………………………………………………………………………………………………………………………….3
- 3a. Female A1 six-segmented; armature formula of A2 EXP 1,0,2; mandibular palp small, one-segmented, with one seta; P1 ENP2 without inner lateral seta and with two distal elements; P3 ENP3 with one inner lateral seta, and one inner distal, one distal, and one outer distal element; female P5 and BENP fused, ENP with four setae…………………………………………………………………………………………………………………………………………………………………………………………………………………..Z. reyssi
- 3b. Female A1 seven-segmented; armature formula of A2 EXP 1,0,2–3; mandibular palp with unisetose basis, exopod incorporated to basis and represented by one seta, endopod with two setae; P1 ENP2 with one inner lateral seta, and one inner distal, one distal, and one outer distal element; P3 ENP3 with two inner lateral setae, and one inner distal, one distal, and one outer distal element; female EXP and BENP separated, ENP with three setae ……………………………………………………………………………………………………………………………………………4
- 4a. A2 EXP3 with two distal setae……………………………………………………………………………………………………………………………………………………………………………Z. incrassata
- 4b. A2 EXP3 with three distal setae……………………………………………………………………………………………………………………………………………………………………………Z. bathybia
- 5a. Armature formula of P1 EXP 1,2,3 (with three outer spines)………………………………………………………………………………………………………………………………………6 subgroup A
- 5b. Armature formula of P1 EXP 1,2,2 (with two outer spines)……………………………………………………………………………………………………………………………………….20 subgroup B
- 6a. P1 ENP2 with four setae in all (one inner, one distal inner, one distal, and one outer distal seta)…………………………………………………………………………...7 subordinate subgroup Aa
- 6b. Armature complement of P1 ENP not like this…………………………………………………………………………………………………………………………………...14 subordinate subgroups Ab
- 7a. Middle apical seta of P4 ENP3 reduced……………………………………………………………………………………………………………………………………………………………………………..8
- 7b. Middle apical seta of P4 ENP3 well-developed…………………………………………………………………………………………………………………………………………………………………..10
- 8a. Female A1 six-segmented, ancestral segments II and III separated, proximal aesthetasc on segment 4, ancestral segments IV and V separated; maxillule with two epipodal setae………………………………………………………………………………………………………………………………………………………………………………………………………………..Z. pacifica
- 8b. Female A1 seven-segmented, ancestral segments II and III fused, proximal aesthetasc on segment 3, ancestral segments IV and V fused; maxillule with one epipodal seta…………………..9
- 9a. A2 EXP 3 with one lateral and three distal setae……………………………………………………………………………………………………………………………………………..Z. mexicana sp. nov.
- 9b. A2 EXP3 with one lateral and two distal setae………………………………………………………………………………………………………………………………………………………..Z. montagnai
- 10a. Female A1 eight-segmented; female P5 ENP with four setae………………………………………………………………………………………………………………………………………….Z. anneae
- 10b. Female A1 six- or seven-segmented; female P5 ENP with three or four setae………………………………………………………………………………………………………………………………11
- 11a. Female A1 seven-segmented; setal complement of female P5 ENP variable from three to four elements…………………………………………………………………………………..Z. paratypica
- 11b. Female A1 six-segmented………………………………………………………………………………………………………………………………………………………………………………………… 12
- 12a. Female P5 ENP with four setae……………………………………………………………………………………………………………………………………………………………………………..Z. major
- 12b. Female P5 ENP with three setae…………………………………………………………………………………………………………………………………………………………………………………..13
- 13a. Male P5 EXP with four setae in all (three marginal, 1 surface element)………………………………………………………………………………………………………………………………Z. typica
- 13b. Male P5 EXP with three setae in all (without surface element)………………………………………………………………………………………………………………………………………..Z. gisleni
- 14a. P1 ENP2 without inner lateral seta, with tree distal elements (inner distal, distal, and outer distal)……………………………………………………………………..15 subordinate subgroup Ab
- 14b. P1 ENP2 with inner lateral seta, and two distal elements………………………………………………………………………………………………………………………17 subordinate subgroup Ac
- 15a. Female A1 six-segmented; baseoendopods of both pairs of female P5 separated……………………………………………………………………………………………………………Z. mediterranea
- 15b. Female A1 seven- or eight-segmented; baseoendopods of both pairs of female P5 fused medially or separated………………………………………………………………………………………16
- 16a. Female A1 seven-segmented; A2 ENP2 with five elements distally; mandibular exopod with four setae; maxillule with two epipodal setae; baseoendopods of both pairs of female P5 separated ………………………………………………………………………………………………………………………………………………………………………………………………………...Z. atlantica
- 16b. Female A1 eight-segmented; A2 ENP2 with six elements distally; mandibular exopod with two setae; maxillule with one epipodal seta; baseoendopods of both pairs of female P5 fused medially ………………………………………………………………………………………………………………………………………………………………………………………………….Z. xiximi sp. nov.
- 17a. Female antennule seven-segmented, ancestral segments IV and V fused; A2 EXP3 with one lateral and three distal setae; A2 ENP1 unarmed; P1 ENP1 unarmed; female and male P5 EXP with three marginal setae (without surface outer element); baseoendopods of both pairs of female P5 fused medially……………………………………………………………………………Z. thistlei
- 17b. Segmentation of female antennule different, other characters not combined……………………………………………………………………………………………………………………………….18
- 18a. Female A1 eight-segmented; syncoxa of maxilliped with one seta……………………………………………………………………………………………………………………………………..Z. eliasi
- 18b. Female A1 six-segmented; syncoxa of maxilliped unarmed…………………………………………………………………………………………………………………………………………………..19
- 19a. Endopod of maxilliped with three elements; P2 ENP3 with four elements in all (one inner lateral seta, and one inner distal, one distal, and one outer distal element); outer exopodal spines of P1 with heterogeneous subdistal long spinules; female P5 ENP with three setae…………………………………………………………………………………………………………..Z. bergensis
- 19b. Endopod of maxilliped with four elements; P2 ENP3 with three elements in all (without inner lateral armature); heterogeneous subdistal long spinules present only on the distalmost outer spine of P1 EXP; female P5 ENP with four setae……………………………………………………………………………………………………………………………………………………...Z. tunnelli
- 20a. P1 ENP2 without inner lateral seta, with three distal elements (inner distal, distal, and outer distal element)…………………………………………………………………………………………21
- 20b. P1 ENP2 with inner lateral seta and two distal elements………………………………………………………………………………………………………………………………………………………23
- 21a. P2–P4 EXP3 with two outer spines………………………………………………………………………………………………………………………………………………………………………...Z. changi
- 21b. P2–P4 EXP3 with three outer spines………………………………………………………………………………………………………………………………………………………………………………22
- 22a. Female A1 seven-segmented; maxillule with two epipodal setae; P4 ENP3 with four seta in all (one inner lateral seta, and one inner distal, one distal, and one outer distal element)……………………………………………………………………………………………………………………………………………………………………………………………………………..Z. valida
- 22b. Female A1 six-segmented; maxillule with one epipodal seta; P4 ENP3 with three seta in all (without inner lateral armature)……………………………………………………………Z. destituta
- 23a. P2–P4 EXP3 with three outer spines……………………………………………………………………………………………………………………………………………………………………………....24
- 23b. P2–P4 EXP3 with two outer spines………………………………………………………………………………………………………………………………………………………………………………..25
- 24a. P1 EXP2 with one inner seta…………………………………………………………………………………………………………………………………………………………………………….Z. paramajor
- 24b. P1 EXP2 unarmed…………………………………………………………………………………………………………………………………………………………………………………………Z. erythraea
- 25a. Female A1 six-segmented; P2–P3 ENP3 with or without inner lateral seta but with three distal elements (inner distal, distal, and outer distal element); P4 ENP3 with two (one inner lateral and one distal seta) or three setae in all (without inner lateral seta, and with three distal elements)………………………………………………………………………………………………….26
- 25b. Female A1 eight-segmented; P2–P4 ENP3 with one inner lateral seta and two distal elements………………………………………………………………………………………….Z. gymnokosmosa
- 26a. P2–P3 ENP3 with four elements in all (one inner lateral seta, and one inner distal, one distal, and one outer distal element); P4 ENP3 with two elements in all (one inner lateral and one distal seta)………………………………………………………………………………………………………………………………………………………………………………………………………….Z. comata
- 26b. P2–P4 ENP3 with three elements in all (without inner lateral seta)………………………………………………………………………………………………………………………………...Z. bathyalis
4.6. The Genera Peresime and Pseudozosime
The monotypic genus Pseudozosime was created by Scott T. [5] for Ps. browni from South Orkney Islands. The genus was originally attributed to the Cerviniidae Sars G. O., 1903 (=Aegisthidae Giesbrecht, 1892) and is unique in the combination of a two-segmented P2 ENP and in the imperfectly separated rami of the female P5 (with an anterior suture dividing the exopod and baseoendopod). The two-segmented P2 ENP occurred independently in Acritozosime (Table S1) and is regarded as apomorphic for these two genera. The presence of an anterior suture dividing the female P5 exopod and baseoendopod is present also in Z. carsteni (Table S1). On the other hand, our results showed that Pseudozosime is the sister taxon of Peresime. These two genera share the two-segmented P3–P4 ENP (Table S1). Two-segmented endopods of P3 and P4 occurred convergently in Acritozosime, but the latter differs from Pseudozosime and Peresime in many important characters (see Table S1).
The genus Peresime was created by Dinet [3] for P. abyssalis from Walvis Ridge (south-western Africa). The genus was originally attributed to the family Tisbidae Stebbing, 1910, and Pe. abyssalis was unique in the three-segmented P2 ENP and the two-segmented P3–P4 ENP. Later on, Seifried [11] erected and diagnosed the family Zosimeidae and removed Peresime, Pseudozosime and Zosime from Idyanthidae and reallocated them into her newly created family. The genus Peresime is currently composed of four species, Pe. abyssalis, Pe. pryorae, Pe. reducta, and Pe. pryocta sp. nov.
The following characters are present in the ground pattern of Peresime: (i) lateral armature of the A2 ENP2 composed of three spines, (ii) four setae on the mandibular exopod (the reduction from four to three setae in Pe. abyssalis and Pe. pryorae is regarded here as secondary within the genus), (iii) five setae (one lateral, four apical) on the mandibular endopod, (iv) one seta on the syncoxa of the maxilliped (secondary reduction leading to the unarmed syncoxa occurred in Pe. reducta; the maxilliped of Pe. abyssalis has not been described yet), (v) caudal rami subquadrate, (vi) armature formulae of P2–P4 EXP (1,1,223), (vii) presence of the lateroproximal inner seta on P1 ENP2, (viii) three-segmented P2 ENP, and two-segmented P3–P4 ENP, (ix) presence of two lateroproximal inner setae on P3 ENP2 (the same was observed in Z. incrassata and Z. bathybia of the incrassata-group of Zosime), (x) heterogeneous ornamentation of the outer spines of P1 EXP (but also in P2 EXP in Pe. reducta), (xi) P5 baseoendopods not fused medially in the female, (xii) endopodal lobe of the male P5 with two setae, (xiii) outer basal seta of the male P6 setiform (the males of Pe. pryorae, Pe. abyssalis, and Pe. pryocta remain undescribed).
The following derived states are present in the genus: (i) one-segmented endopod of the maxilla (present in several species of Zosime; see above), (ii) reduced distal armature complement on the P1 ENP2 from three to two setae (an additional seta was secondarily lost in Pe. abyssalis, for which is regarded here as autapomorphic) as in subordinate subgroups Ac and Bb of the typica-group of Zosime, and in Z. reyssi (incrassata-group of Zosime), in H. klemie sp. nov., and Acritozosime, (iii) loss of the lateroproximal inner seta of P2 ENP3 as in Z. tunnelli and Z. bathyalis, and Z. carsteni (typica- and incrassata-group of Zosime, respectively), and Acritozosime, (iv) two-segmented P3 and P4 endopods as in Pseudozosime and Acritozosime, (v) reduction from three to two distal setae on P3 ENP2 as in Acritozosime, (vi) reduction in the armature complement to one distal seta on P4 ENP2 (regarded here as apomorphic for the genus; a similar reduction on the three-segmented endopod occurred convergently in Z. comata (typica-group of Zosime)), (vii) homogeneous spinular ornamentation on the outer spines of P3–P4 EXP, (viii) P2–P4 ENP longer than wide, (xi) reduction from four to three setae on the female P5 endopodal lobe, (x) caudal rami elongated.
4.7. Key to the Species of Peresime
- 1a. Female A1 six-segmented; A2 ENP2 with five or six distal elements……………………………………………………………………………………………………………………………………………2
- 1b. Female A1 eight-segmented; A2 ENP2 with six or seven distal elements………………………………………………………………………………………………………………………………………3
- 2a. A2 EXP3 with four setae; A2 ENP2 with five distal elements; mandibular exopod with four setae; maxillule without epipodal seta; P1 ENP2 with three setae in all; female P5 EXP with three setae in all…………………………………………………………………………………………………………………………………………………………………………………………………..Pe reducta
- 2b. A2 EXP3 with three setae; A2 ENP2 with six distal elements; mandibular exopod with three setae; maxillule with one epipodal seta; P1 ENP2 with two setae in all; female P5 EXP with two setae in all………………………………………………………………………………………………………………………………………………………………………………………………….Pe. abyssalis
- 3a. A2 ENP2 with six distal setae; mandibular exopod with three setae; maxillule without epipodal seta; basis of maxilliped with one seta; female P5 EXP with three setae………………………………………………………………………………………………………………………………………………………………………………………………………………Pe. pryorae
- 3b. A2 ENP2 with seven distal setae; mandibular exopod with four setae; maxillule with one epipodal seta; basis of maxilliped unarmed; female P5 EXP with two setae……………………………………………………………………………………………………………………………………………………………………………………………………..Pe. pryocta sp. nov.
4.8. The Genus Heterozosime
The genus Heterozosime was created recently by Kim and Lee [1] for H. tenuis found at 78.7 m depth in the southern Sea of Korea, and is characterized by: (i) the female antennule six-segmented with aesthetasc on second segment; (ii) the two-segmented P1 EXP with two inner setae on P1 EXP2; (iii) P1 ENP two-segmented; (iv) P2–P4 EXP and ENP three-segmented; (v) the reduction from three to two outer spines on P2–P4 EXP3; (vi) the primitive heterogeneous spinulation pattern on the outer spines of P1 EXP, and derived homogeneous pattern on the outer spines of P2–P4 EXP; (vii) the presence of one seta on A2 EXP1; (viii) the presence of two spines and one small slender setae laterally on the A2 ENP2; (ix) the mandibular palp with three, three (one lateral, two apical), and four (one lateral, three apical) setae on the mandibular basis, exopod, and endopod, respectively; (x) the presence of one epipodal seta on the coxal endite of the maxillule; (xi) the maxillary endopod two-segmented; (xii) the maxillipedal syncoxa unarmed; and (xiii) the male and female P5 EXP with three marginal and one outer surface setae, and endopodal lobe with two setae in both sexes. Following Kim and Lee [1], Heterozosime displays the autapomorphies for Zosimeidae as defined by Seifried [11] and is very similar to Zosime, Z. changi, Z. comata, Z. bathyalis, and Z. gymnokosmosa (typica-group, subgroup B as defined above) in the presence of two outer spines on P2–P4 EXP3. Kim and Lee [1] noticed some similarities in the dimorphic two-segmented male P2 ENP of H. tenuis and Z. changi (the male P2 ENP is three-segmented in Z. comata and Z. gymnokosmosa). As noticed by Kim and Lee [1], the two-segmented P1 EXP is present also in some Idyanthidae (Styracothorax Huys, 1993, Aspinothorax Moura and Martínez Arbizu, 2003, and Meteorina George, 2004), and in Acritozosime. They [1] also noticed that the armature formula of P1 EXP of A. spinesco is as in the copepodid IV of Z. pacifica, i.e., 0,223, and suggested that the two-segmented condition of P1 of A. spinesco and H. tenuis is the result of suppression of the division between the middle and distal segments during ontogeny. The second and third exopodal segments of P1 became fused in H. tenuis without loss of any armature element, but P1 EXP2 and EXP3 became fused in A. spinesco with additional loss of one of the inner setae, either the inner seta of EXP2 or EXP3, during ontogeny. Kim and Lee [1] argued that this is strong evidence to support independent origins of Acritozosime and Heterozosime; thus, the two-segmented P1 EXP is a clear autapomorphy for each of these genera. They reinforced their hypothesis based on the occurrence of several unambiguous autapomorphies and some plesiomorphic states in A. spinesco, which also indicate its basal position within Zosimeidae. However, the fact that the two-segmented P1 EXP is present in Styracothorax, Aspinothorax, and Meteorina (Idyanthidae), but also in Heterozosime and Acritozosime (Zosimeidae) indicates that the two-segmented P1 EXP possibly evolved independently twice in Idyanthidimorpha. In our opinion, there is not enough evidence to suggest the separate and independent evolution of the two-segmented P1 EXP in Heterozosime and Acritozosime (with one ancestor for each genus, instead of a common ancestor for these two genera), and could be a synapomorphy for Heterozosime and Acritozosime, rather than an autapomorphy for each of them as suggested by Kim and Lee [1]. Our results (Figure 32) support this view. The new species proposed here, H. klemie sp. nov. is different from the type species H. tenuis in several aspects and a rediagnosis of the genus is justified (see above).
4.9. Key to the Species of Heterozosime
- 1a. P1 ENP2 with four setae in all (one inner lateral and three distal elements); P2–P4 EXP3 with six elements in all (with two outer spines); medial apical seta on P3 ENP3 reduced; female P5 ENP with two setae………………………………………………………………………………………………………………………………………………………………………………………………..H. tenius
- 1b. P1 ENP2 with three setae in all (one inner lateral and two distal elements); P2–P4 EXP3 with seven elements in all (with three outer spines); medial apical seta on P3 ENP3 well-developed; female P5 ENP with four setae…………………………………………………………………………………………………………………………………………………………..H. klemie sp. nov.
4.10. The Genus Acritozosime
The genus Acritozosime was created recently by Kim and Lee [1] for A. spinesco, found at a depth of 5317 m in the East Mariana Basin (north-western Pacific). Kim and Lee [1] (p. 54) detected some similarities between their newly created genus and Idyanthidae in (i) A2 EXP1 with two setae; (ii) A2 ENP2 with seven distal setae; and (iii) maxilliped with geniculation between syncoxa and basis, and two setae on syncoxa. At the same time, they [1] (p. 56) observed that Acritozosime is similar to Zosimeidae in terms of the (i) shape and kind of armature complements of the female antennule; (ii) lateral armature complement of A2 ENP2 with tree elements; (iii) mandibular endopod with one lateral seta only; (iv) one-segmented mandibular exopod with four setae; (v) maxillulary exopod smaller than the endopod; (vi) position of the maxillary endites; (vii) shape of the endopod of the maxilliped and of its armature complements; (viii) length:width ratio of P1 ENP1; and (ix) shape of the female P5. As a result of their analyses, Kim and Lee [1] (p. 56) attributed Acritozosime to the family Zosimeidae, suggested some relationship between their newly created genus and Pseudozosime, and observed that the (i) loss of the inner element of the A2 ENP1, (ii) loss of the epipodal seta of the maxillulary coxa, and (iii) caudal seta V elongated are unambiguous features of A. spinesco.
As noted above, Kim and Lee [1] favored independent origins of Acritozosime and Heterozosime. However, we are inclined to believe that the two-segmented condition of P1 EXP occurred independently in Idyanthidae and Zosimeidae, and that Acritozosime and Heterozosime share a common ancestor, with the two-segmented P1 EXP being a synapomorphy for both genera.
The new species proposed here, A. sigsbeeiensis sp. nov., is different from the type species A. spinesco in several aspects and a rediagnosis of the genus is justified (see above).
4.11. Key to the Species of Acritozosime
- 1a. A2 ENP1 without inner seta; maxilliped geniculate, syncoxa with two setae; P2–P3 EXP1 with one inner seta………………………………………………………………………………A. spinesco
- 1b. A2 ENP1 with inner seta; maxilliped non-geniculate, syncoxa with one seta; P2–P3 EXP1 unarmed……………………………………………………………………………A. sigsbeeiensis sp. nov.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d15030363/s1, Figure S1: Preliminary phylogenetic tree of the family Zosimeidae showing the position of Styracothorax gladiator (Styracothoracidae) and the different groupings of Zosime.; Table S1: Matrix used for the phylogenetic analysis; Table S2: Segmentation pattern of the female antennule in Zosimeidae.
Author Contributions
Conceptualization, S.G.; investigation, S.G.; writing—original draft preparation, S.G.; preparation of figures, S.G.; preparation of list of characters and characters matrix, S.G. and B.Y.-R.; writing—review and editing, S.G. and B.Y.-R.; phylogenetic analysis, B.Y.-R.; funding acquisition, S.G.; project administration, S.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Secretaría de Energía (SENER)-Hydrocarbon Trust of the National Council on Science and Technology (Consejo Nacional de Ciencia y Tecnología [CONACyT]) and Petróleos Mexicanos (PEMEX) through project 201441 of the Gulf of Mexico Research Consortium (CIGoM), and by the Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica (PAPIIT) of the Dirección General de Asuntos del Personal Académico of the Universidad Nacional Autónoma de México (UNAM-DGAPA-PAPIIT) through project IN202116. Xiximi-5 cruise took place on board R/V “Justo Sierra of the UNAM. Talud IV and XV cruises on board R/V “El Puma” of the UNAM were financed by CONACyT, projects 31805-N and 179467, respectively, and ship time was provided by the Coordinación de la Investigación Científica (CIC)-UNAM. Article processing charge was financed by the Instituto de Ciencias del Mar y Limnología of the Universidad Nacional Autónoma de México.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The voucher specimens of the species presented here were deposited in the Copepoda collection of the Instituto de Ciencias del Mar y Limnología, Unidad Académica Mazatlán (ICML-EMUCOP).
Acknowledgments
We are grateful to Axayácatl Rocha-Olivares (Centro de Investigación Científica y de Educación Superior de Ensenada [CICESE]), Ivonne Martínez-Mendoza, Celia Bisbal, Samantha Guerrero, Ivette Lara, and Viridiana Tapia for taking and sorting of meiofauna samples during the Xiximi-5 cruise, and for putting the benthic harpacticoids at the disposal of the first author. Our thanks go to Karen Itzel Rivera Sánchez for preliminary identification of the material from the Gulf of Mexico. We are grateful to scientists, students, and crew members of R/V “Justo Sierra” and “El Puma” (UNAM) for their help and support during oceanographic cruises.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Kim, J.G.; Lee, J. Proposal of two new genera of the family Zosimeidae Seifried (Copepoda: Harpacticoida): Heterozosime gen. nov. and Acritozosime gen. nov. Eur. J. Taxon. 2021, 760, 32–60. [Google Scholar] [CrossRef]
- Yeom, J.; Rohal Lupher, M.; Lee, W. Four new species of Zosimeidae (Copepoda: Harpacticoida) from the southwestern Gulf of Mexico. Diversity 2022, 14, 198. [Google Scholar] [CrossRef]
- Dinet, A. Espèces nouvelles de copépodes harpacticoïdes (Crustacea) des sédiments profonds de la dorsale de Walvis. Arch. Zool. Expérimentale Générale 1974, 115, 549–577. [Google Scholar]
- Becker, K.-H.; Schriever, G. Eidonomie und Taxonomie abyssaler Harpacticoidea (Crustacea, Copepoda) Teil III. 13 Neue Tiefsee-Copepoda Harpacticoidea der Familien Canuellidae, Crviniidae, Tisbidae, Thalestridae, Diosaccidae und Ameiridae. “Meteor” Forschungsergebnisse 1979, Reihe D-Bi, 38–62. [Google Scholar]
- Scott, T. The Entomostraca of the Scottish National Antarctic Expedition, 1902–1904. Trans. Linn. Soc. Edinburgh. 1912, XLVIII, 521–599. [Google Scholar] [CrossRef]
- Gómez, S.; Rivera-Sánchez, K.I. The Deep-Water Benthic Harpacticoida (Copepoda) of the Americas. In Deep-Sea Pycnogonids and Crustaceans of the Americas; Hendrickx, M.E., Ed.; Springer: Cham, Switzerland, 2020; pp. 47–125. ISBN 978-3-030-58409-2. [Google Scholar]
- Koller, S.; George, K.H. Description of a new species of Zosime Boeck, 1872 (Copepoda: Harpacticoida: Zosimeidae) from the Great Meteor Seamount, representing one of the few eurybathic Harpacticoida among the distinct plateau and deep-sea assemblages. Meiofauna Mar. 2011, 19, 109–126. [Google Scholar]
- Kitahashi, T.; Kawamura, K.; Kojima, S.; Shimanaga, M. Assemblages gradually change from bathyal to hadal depth: A case study on harpacticoid copepods around the Kuril Trench (north-west Pacific Ocean). Deep. Res. Part I Oceanogr. Res. Pap. 2013, 74, 39–47. [Google Scholar] [CrossRef]
- Kitahashi, T.; Kawamura, K.; Kojima, S.; Shimanaga, M. Bathymetric Patterns of α and β diversity of harpacticoid copepods at the genus level around the Ryukyu Trench, and turnover diversity between trenches around Japan. Prog. Oceanogr. 2014, 123, 54–63. [Google Scholar] [CrossRef]
- Gómez, S.; Morales-Serna, F.N. Updated checklist of published and unpublished records of harpacticoid copepods (Crustacea: Copepoda: Harpacticoida) from Mexico. Proc. Biol. Soc. Washingt. 2014, 127, 99–121. [Google Scholar] [CrossRef]
- Seifried, S. Phylogeny of Harpacticoida (Copepoda): Revision of “Maxillipedasphalea” and Exanechentera; Cuvillier Verlag: Göttingen, Germany, 2003. [Google Scholar]
- Kim, J.G.; Jung, T.W.; Yoon, S.M. A new species of Zosime (Copepoda: Harpacticoida: Zosimeidae) from shallow subtidal habitats in Korea, with notes on the genus. Zootaxa 2016, 4136, 323–334. [Google Scholar] [CrossRef]
- Pointner, K. Description of two new species of Zosime (Copepoda: Harpacticoida: Zosimeidae), including remarks on its phylogeny and distribution on the Great Meteor Seamount Plateau (north-east Atlantic Ocean). J. Nat. Hist. 2017, 51, 2283–2330. [Google Scholar] [CrossRef]
- Yoder, M.; De Ley, I.T.; King, I.W.; Mundo-Ocampo, M.; Mann, J.; Blaxter, M.; Poiras, L.; De Ley, P. DESS: A versatile solution for preserving morphology and extractable DNA of nematodes. Nematology 2006, 8, 367–376. [Google Scholar] [CrossRef]
- Somerfield, P.J.; Warwick, R.M. Meiofauna in Marine Pollution Monitoring Programmes. A Laboratory Manual; Ministry of Agriculture, Fisheries and Food Directorate of Fisheries Research: Great Britain, UK, 1996.
- de Jonge, V.N.; Bouwman, L.A. A simple density separation technique for quantitative isolation of meiobenthos using the colloidal silica Ludox-TM. Mar. Biol. 1977, 42, 143–148. [Google Scholar] [CrossRef]
- Huys, R.; Boxshall, G.A. Copepod Evolution; The Ray Society: London, UK, 1991. [Google Scholar]
- Sewell, R.B.S. The littoral and semi-parasitic Cyclopoida, the Monstrilloida and Notodelphyoida. Sci. Reports John Murray Exped. Publ. Br. Museum Natural Hist. 1949, 9, 17–199. [Google Scholar]
- Lang, K. Marine Harpacticiden von der Campbell-Insel und einigen anderen südlichen Inseln. Acta Univ. Lund. New Ser. Avd. 2 Kungl. Fysiogr. Sällskapets Handl. 1934, 30, 1–56. [Google Scholar]
- Felsenstein, J. PHYLIP (Phylogeny Inference Package), Version 3.6; Department of Genome Sciences at University of Washington: Washington, DC, USA, 2005. Available online: http://evolution.genetics.washington.edu/phylip.html (accessed on 22 January 2023).
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive tree of life (ITOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, W1, W293–W296. [Google Scholar] [CrossRef]
- ICZN (International Commission on Zoological Nomenclature). International Code of Zoological Nomenclature, 4th ed.; The International Trust for Zoological Nomenclature: London, UK, 1999. [Google Scholar]
- Bodin, P. Copépodes Harpacticoïdes des etages bathyal et abyssal du Golfe de Gascogne. Mémoires du Muséum Natl. d´Histoire Nat. Sér. 1968, 55, 1–107. [Google Scholar]
- Monard, A. Les harpacticoïdes marins de la région d’Alger et de Castiglione. Bull. la Stn. d’Aquiculture Pêche Castiglione 1937, 1935, 9–93. [Google Scholar]
- Kornev, P.N.; Chertoprud, E.C. Copepod Crustaceans of the Order Harpacticoida of the White Sea: Morphology, Systematics, Ecology; Biology Faculty, M.S.U., Ed.; Tovarishchestvo Nauchnikh Izdanii KMK: Moscow, Russia, 2008. [Google Scholar]
- Kim, J.G.; Lee, J. New records of benthic harpacticoids from Korea: I. Three new species of the genus Zosime Boeck (Copepoda: Harpacticoida: Zosimeidae). Mar. Biol. Res. 2019, 15, 210–239. [Google Scholar] [CrossRef]
- Fiers, F. Three new harpacticoid copepods from the Santa Maria Basin off the Californian Pacific coast (Copepoda, Harpacticoida). Beaufortia 1991, 42, 13–47. [Google Scholar]
- Huys, R.; Clark, P.F. Case 3467 Zosimidae Seifried, 2003 (Crustacea, Copepoda, Harpacticoida): Proposed emendation of spelling to Zosimeidae to remove homonymy with Zosiminae Alcock, 1898 (Crustacea, Decapoda, Xanthidae). Bull. Zool. Nomencl. 2009, 66, 24–29. [Google Scholar] [CrossRef]
- Mu, F.H.; Somerfield, P.J.; Warwick, R.M.; Zhang, Z.N. Large-scale spatial patterns in the community structure of benthic harpacticoid copepods in the Bohai Sea, China. Raffles Bull. Zool. 2002, 50, 17–26. [Google Scholar]
- Ma, L.; Li, X. Benthic harpacticoid copepods of Jiaozhou Bay, Qingdao. Chinese J. Oceanol. Limnol. 2017, 35, 1127–1133. [Google Scholar] [CrossRef]
- Chertoprud, E.S.; Gómez, S.; Gheerardyn, H. Harpacticoida (Copepoda) fauna and the taxocene diversity of the South China Sea. Oceanology 2009, 49, 488–498. [Google Scholar] [CrossRef]
- Shimanaga, M.H.K.; Shirayama, Y. Temporal patterns in diversity and species composition of deep-sea benthic copepods in bathyal Sagami Bay, Central Japan. Mar. Biol. 2004, 144, 1097–1110. [Google Scholar]
- George, K.H.; Pointner, K.; Packmor, J. The benthic Copepoda (Crustacea) of Anaximenes Seamount (eastern Mediterranean Sea)—Community structure and species distribution. Prog. Oceanogr. 2018, 165, 299–316. [Google Scholar] [CrossRef]
































Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).