Abstract
Studies of the paleopathology of the vertebrae provide an interesting, oblique approach to their paleobiology and even paleoethology. They tell us about possible ethological causes such as accidental blows with objects, social interactions within a group, and defense against predators, etc. There are numerous works on the anatomical and phylogenetic aspects of sauropod dinosaurs, and in recent years paleopathological studies have also increased. Here, we describe the injuries recorded in the caudal vertebrae of two indeterminate titanosaurids, and undertake a compilation and analysis of the pathological fossil record worldwide, focusing on the tails of sauropods. Two pathologies have been identified as present in the indeterminate titanosaurids under study: a possible case of spondyloarthropathy in MAU-Pv-LI-601, and a case of suppurative spinal osteomyelitis in MAU-Pv-LJ-472/1. Both titanosaurids are from Gondwana. In the world registry of pathologies associated with the tails of sauropod dinosaurs, it is observed that titanosaurs are the sauropods with the highest percentage of diagnosed pathologies (69% including the two new records from the province of Neuquén) and that all of these have been described in Gondwanan specimens.
1. Introduction
Sauropods are one of the most abundant groups of dinosaurs in the Jurassic-Cretaceous fossil record, representing the largest terrestrial quadrupedal megaherbivores that have ever existed on the planet [1]. Their remains have been found on all continents, including Antarctica [2]. They originated in the Late Triassic, were predominant in the Middle Jurassic, and became one of the main faunal components towards the end of the Cretaceous [3,4]. Of all known sauropod groups, titanosaurs were the only group to survive until the Late Cretaceous, reaching their greatest diversity on the Gondwanan continents and especially in South America [5,6].
Over the last decade, numerous works have been published on sauropod dinosaurs, mainly on titanosaurs. These have deepened our knowledge of anatomical and phylogenetic aspects [7,8,9,10,11], in addition to paleobiological aspects [12,13,14,15,16,17]. Recent years have seen an increase in studies on paleopathology in general [18,19,20,21,22,23,24,25,26,27] and on sauropod paleopathology in particular [28,29,30,31]. These studies provide valuable information on the historical record of the injuries suffered by these dinosaurs and how they affected their paleobiology and paleoecology [32,33].
In Gondwana, the paleopathological record of sauropod tails is rich and mainly focuses on two countries: Brazil and Argentina. Here we study two titanosaurid caudal vertebrae from two paleontological sites in Argentina, Loma de los Jotes and La Invernada. Loma de los Jotes is located about 13 km southwest of the town of Rincón de los Sauces, in the province of Neuquén (Figure 1). A middle caudal vertebra (MAU-Pv-LJ-472/1) was found here, together with other sauropod remains in an area of approximately 30 m2. The site is located in a sector of low relief that would correspond to the alluvial descent system developed on the northwestern front of the Sierra del Auca Mahuída, which drains towards the valley of the Colorado River. Notable is a low relief and the fact that it is partially covered by modern (eolian) deposits. In terms of its lithostratigraphic identity, it corresponds to the Sierra Barrosa Formation (Coniacian), Upper Cretaceous (Río Neuquen Subgroup, Neuquen Group), integrated by sedimentary deposits formed by quartzitic to quartz-lithic sandstones, fine to very fine, whitish in color and very friable. These psammites have a slight content of pelitic matrix, which gives them a light greenish hue. They present a good textural maturity, with rounded clasts and a good selection, which gives them a very uniform granulometric appearance. The second caudal vertebra is an anterior vertebra (MAU-Pv-LI-601) and comes from the La Invernada site, located about 50 km southwest of the town of Rincón de los Sauces, in the province of Neuquén (Figure 1). This material was found isolated in an area of great fossiliferous abundance, comprising sedimentary deposits made up of reddish-orange quartz sandstones with little matrix, which are interspersed with pelitic horizons accompanied by gypsum deposits [34]. These outcrops are assigned to the Bajo de la Carpa Formation (Santonian), and Late Cretaceous (Río Colorado Subgroup, Neuquen Group). The depositional environment of the Bajo de la Carpa Formation corresponds to a river and floodplains, with the participation of eolian deposits [35].
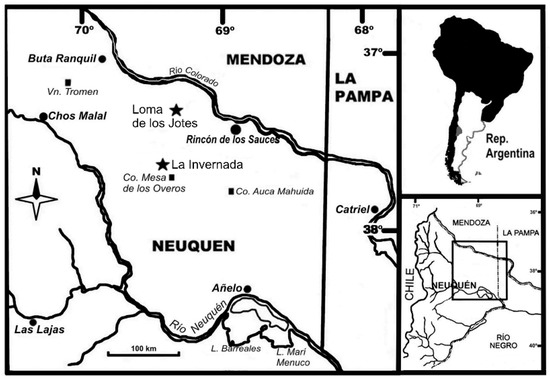
Figure 1.
Map showing the location of La Invernada and Loma de los Jotes paleontological sites (Rincón de los Sauces city, Neuquén province, Argentina).
Here we present a complete revision of the pathological record of sauropod tails and a new contribution to the paleopathological record of titanosaur caudal vertebrae, from the Upper Cretaceous of Argentine Patagonia.
2. Material and Methods
2.1. Institutional Abbreviations
MAU, Museo Municipal Argentino Urquiza (Rincón de los Sauces, Neuquén, Argentina).
2.2. Material
The specimens studied are two caudal vertebrae (MAU-Pv-LI-601 and MAU-Pv-LJ-472/1; Figure 2) belonging to two indeterminate sauropods from different Upper Cretaceous localities (the sites of La Invernada and Loma de los Jotes) to the southwest of the town of Rincón de los Sauces (Neuquén Province, Patagonia, Argentina).
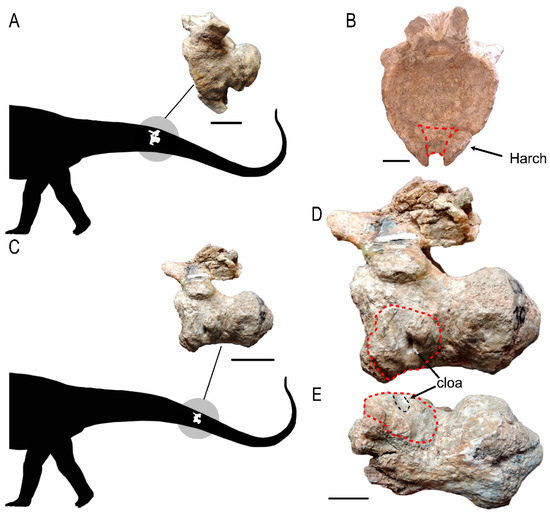
Figure 2.
Caudal vertebrae showing the delimitation of the pathological area: (A) localization of MAU-Pv-LI-601 within a graphical scheme of the tail of an indeterminate sauropod; and (B) an anterior view of the vertebral centrum; (C) MAU-Pv-LJ-472/1 located within the graphical scheme of the tail; (D) MAU-Pv-LJ-472/1 in lateral view and (E) in ventral view. The extent of pathological tissue on the vertebral surface is marked by a red dashed line; the cloaca is marked by a black arrow in lateral view (D) and by a black dashed line in ventral view (E). The haemal arch is marked by a black arrow in the anterior view (B). Abbreviations: Cloa, cloaca; Harch, haemal arch. Scale bars equal 5 cm in (A,C) and 3 cm in (B,D,E).
The anterior caudal vertebra MAU-Pv-LI-601 (Figure 2B,E) comes from the Bajo de la Carpa Formation (Santonian) in the Río Colorado Subgroup (Neuquén Group) of the Neuquén Basin. The middle caudal vertebra MAU-Pv-LJ-472/1 (Figure 2A,C,D) comes from the Sierra Barrosa Formation (Coniacian) in the Río Neuquén Subgroup (Neuquén Group) of the Neuquén Basin.
2.3. CT Scan
Both bones were CT-scanned using axial computed tomography (Aquilion Lightning CT system), at the Clínica y Maternidad Eva Perón, Rincón de los Sauces (Neuquén Province, Argentina). Settings for a differential diagnosis of bone pathologies were used (120 kV and 219 mA with 0.8 slice thickness). The CT scan images were saved in DICOM format images and analyzed with 3D Slicer 5.0.3 software. The image data are stored on DVD and deposited with the specimen at the Museo Municipal Argentino Urquiza, Rincón de los Sauces (Neuquén, Argentina).
3. Description
3.1. External Morphology
The caudal vertebrae are assigned to titanosaur sauropods on account of the combination of an anterior neural arch and well-developed procoelous articulations, a condition frequently observed in the clade during the Late Cretaceous [36].
MAU-Pv-LI-601 is an anterior caudal vertebra (Figure 2B,E), as shown by the development of the posterolateral transverse process and the presence of articulation for the haemal arch [37]. This vertebra shows a small periosteal reaction of the bone tissue (formation of pathological tissue) and is located on the ventral side. The haemal arch, of which only about 5 cm of the proximal end is preserved, is ankylosed to the ventral surface of the centrum by this pathological tissue, as can be observed in the posterior view (Figure 2E). This tissue filled the most proximal section of the haemal canal. The subperiosteal surface of the vertebra has a rough texture and is covered with shallow, irregular pits, probably due to infection.
MAU-Pv-LJ-472/1 is a vertebra from the middle section of the tail, as shown by the reduction of the transverse process and the absence or reduction of the articulation facets for the haemal arch [38] (Figure 2A,C,D). Its vertebral centrum shows an extensive periosteal reaction of the bone tissue associated with an open channel, subcircular in shape, in the anterior border of the periosteal tissue (Figure 2C). The vertical channel has smooth borders and a length of 30.59 mm. Its most ventral section penetrates about 5.28 mm into the vertebral centrum (Figure 2C). This abscess was possibly a drainage hole (cloaca) for pus. The subperiosteal surface of the vertebra shows signs of an infection, namely a rough texture covered with shallow, irregular pits (Figure 2C,D).
3.2. Internal Morphology (CT Data)
For each vertebra, three cross-sectional CT images covering the pathological area were studied to analyze the internal morphology. In MAU-Pv-LI-601 the CT scan images are not very clear, but it can be seen how the periosteal tissue, with a less dense appearance internally, surrounds the healthy tissue of the haemal arch, which can be delineated precisely (Figure 3B). In MAU-Pv-LJ-472/1 the periosteal reaction shows a decrease in bone density too (Figure 3A). On the lateral surface, there is a cavity that communicates with the external surfaces of the periosteal tissue. This opening corresponds to the erosion observed in the proximal section (Figure 2C). These CT images do not reveal any trace of a possible fracture or trauma.
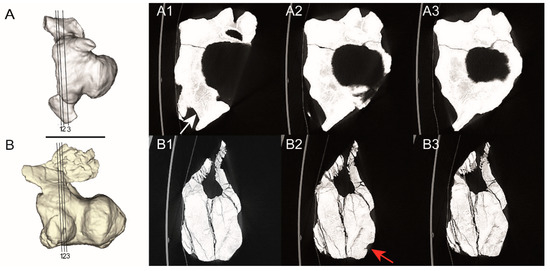
Figure 3.
3D models of the caudal vertebrae and the sequence of CT scan images in the serial axial section for MAU-Pv-LI-601 (A) and MAU-Pv-LJ-472/1 (B). (A1–A3) corresponding to the proximal end of the vertebral centrum MAU-Pv-L I-601 and (B1–B3) corresponding to the proximal end of the vertebral centrum of MAU-Pv-L J-472/1. In (A1) the pathological tissue is marked by a white arrow, and in (B2) the cloaca is marked by a red arrow. Scale bars equal 10 cm.
4. Differential Diagnosis
Analysis of the two caudal vertebrae reveals the following abnormalities on the basis of computed tomography and external macroscopic observations. In MAU-Pv-LI-601 there is: (1) ankylosis of the haemal arch, and (2) lower density of the pathological bone tissue. In MAU-Pv-LJ-472/1 there is: (1) lower density of the pathological bone tissue, (2) an amorphous mass of rough-textured bone with an irregular, shallow depression, and (3) an elliptical erosion (cloaca). Based on these radiological and morphological characteristics, we rule out fractures due to the lack of evidence in the CT images, since no disruption in the continuity of the bone is observed [39,40].
Developmental disorders produce an abnormal proliferation of cells, and to rule them out, it is necessary to undertake an analysis of the pattern of bone destruction and the nature and extent of the medullary, cortical, or periosteal reaction or disruption, as well as of the calcification of the matrix of the tumor [40]. In this way, one can rule out hemolytic anemia and hemangiomas, since these affect the cortical tissue by thinning it, and malignant tumors (osteosarcoma, chondrosarcoma, and hemangiosarcoma [41]) produced by errors in the regulation of cell division [41] because they use thinning and present a spiculated periosteal reactions [40].
These characteristics make us tentatively propose a case of infection in MAU-Pv-LJ-472/1 and a possible case of spondyloarthropathy in MAU-Pv-LI-601.
4.1. Infections
According to Jacobson [42], infectious diseases are produced by the entrance and proliferation of pathogens in an organism’s body (e.g., fungal agents, bacteria, parasites). This may give rise to several reactions, such as inflammation, pain, and sometimes pus. Infections that occur in bone are called osteitis (i.e., inflammation of the bone) or osteomyelitis (i.e., inflammation of bone marrow) and these are not very common in the dinosaur fossil record [27,43]. One possible cause of osteomyelitis is a trauma that affects the soft tissue or bone, resulting in an open wound through which pathogens enter and from which they may spread through the bloodstream [21,44,45].
According to [40,43] in a mammalian immune system model osteomyelitis is classified taking into account whether or not there is a production of pus, being categorized as pyogenic or suppurative, when there is pus, and as non-pyogenic or non-suppurative) when there is no pus production. In addition, depending on the type of response of the body, it can be categorized as acute (a new infection), subacute (caused by an open wound), or chronic (a recurring infection; [40]), where acute and subacute responses cause a periosteal reaction, cortical irregularity, and demineralization. By contrast, a chronic case is characterized by the presence of thick, sclerotic, irregular bone and a swollen periosteal surface [46]. With respect to a reptilian (including birds) immune system model response, there would be the formation of small fibrin cysts (fibrisces) at the origin of infection, which would tend to calcify in advanced stages ([27] and references).
Osteomyelitis is recognized by a periosteal reaction, cortical irregularity, and demineralization. The suppurative type is further differentiated from the non-suppurative by the presence of elliptical abscesses with a draining function [40]. Among the variants of osteomyelitis, spinal osteomyelitis is recognized as an infection that is located in the vertebrae and ribs and is characterized by the presence of irregular growth of new bone with a pitted texture [22].
4.2. Spondyloarthropathy
Spondyloarthropathy is a chronic aseptic inflammatory process that affects multiple articulations in the vertebrae. Sometimes this paleopathology can be confused with diffuse idiopathic skeletal hyperostosis (DISH). The latter is a non-inflammatory enthesitis of the vertebral column that can produce ankyloses and is characterized externally by a “pasted-on” or “dripped candle-wax” appearance, and by ligamentous calcification and ossification that is accentuated in the anterolateral aspect of the spine and the bone insertion areas of tendons, ligaments (especially of the longitudinal ligament) and joint capsules, although it can also be seen in other parts of the body, such as the phalanges [47,48]. In the case of this paleopathology, the fusion of the vertebrae is asymmetrical, and the articulated surfaces appear unaffected [40]. Moreover, it is recognized by its presence in tomographic images as a “fluid” ossification at the anterolateral margins of the vertebrae [49].
Spondyloarthropathy encompasses several rheumatic diseases including spondylitis ankylosans, psoriatic arthritis, juvenile spondyloarthropathy, and reactive arthritis [50]. According to Cisneros et al. [49], the diagnostic criteria are as follows: zygapophyseal or sacroiliac joint erosion or ankylosis; asymmetrical arthritis patterns; reactive new bone formation; syndesmophytes (which are either marginal or are calcifications/ossifications within the annulus fibrosus); or peripheral joint ankylosis. One might also add reduced trabecular density, ossification of tendons and ligaments, and calcification of the intervertebral disc [19,40]. Caution is due since ligamentous ossification can also be produced by diffuse idiopathic skeletal hyperostosis (DISH; [40]), but in this case, the reaction of diffuse idiopathic skeletal hyperostosis is located in the lateral sides of the vertebral centrum and not in the area between branches in the haemal arch. Moreover, another criterion for ruling out diffuse idiopathic skeletal hyperostosis is that in this case ossification of the annulus fibrosus must occur in combination with typical syndesmophytes [49].
4.3. MAU-Pv-LI-601
4.4. MAU-Pv-LJ-472/1
5. Discussion
5.1. The Paleopathological Record of Sauropod Tails
As regards the diversity of the worldwide fossil record of paleopathologies in sauropod tails, nine different diseases, traumas, and disorders (infection, congenital malformation, spondyloarthropathy, diffuse idiopathic skeletal hyperostosis, tumor, cystic lesion, enthesophyte, fracture, and amputation) have been diagnosed so far (Table 1; Supplementary File S1). Regarding the distribution of this paleopathological record between the two supercontinents (Laurasia and Gondwana), we have found that there are more pathological fossil remains diagnosed in Gondwana, with the work carried out in Brazil and Argentina standing out (Figure 4; Table 1; Supplementary File S1 [28,29,30,50,51,52]).

Table 1.
Summary of the record of pathologies associated with the tails of sauropod dinosaurs distinguished by supercontinents (Laurasia and Gondwana) and by types of pathology. *, fracture associate to arch haemal.
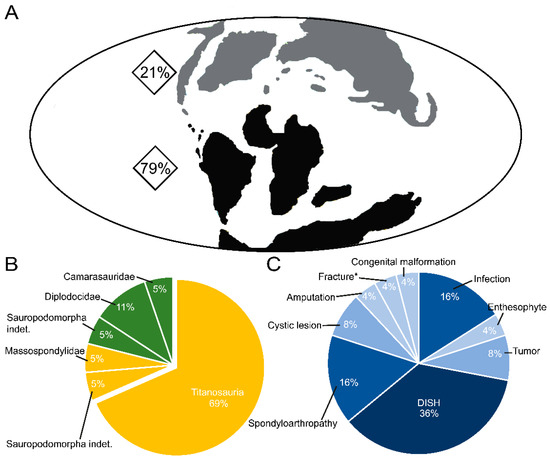
Figure 4.
Distribution of pathologies associated with sauropod dinosaur tails: (A) percentage by supercontinents (Laurasia and Gondwana), (B) pie chart of the pathologies analyzed by clade, and (C) by disease. *, presence of fracture associated with a haemal arch.
The tail is one of the body parts most at risk of injury throughout an animal’s life. After reviewing and analyzing the data obtained from the bibliography on paleopathologies in tails, and further taking into account the difficulty in many cases of distinguishing between diffuse idiopathic skeletal hyperostosis (DISH) and spondyloarthropathy, it can be observed that together they account for just over 50% of the pathological occurrences in the fossil record (Supplementary File S1). On the other hand, if these paleopathologies are distinguished, it emerges that the most commonly recorded paleopathology is diffuse idiopathic skeletal hyperostosis (DISH; 36%, nine records), which affects the Laurasian clades Diplodocidae and Camarasauridae and Gondwanan titanosaurs (this paper; Table 1; Figure 4; Supplementary File S1). This disease is usually recorded between approximately caudal 18 and 23. The spondyloarthropathy record corresponds to 16% of the total, and this paleopathology has been diagnosed more frequently in titanosaurs from the Gondwana supercontinent (three out of four records) and indeterminate sauropodomorphs from the Laurasia supercontinent (this paper; Table 1; Figure 4; Supplementary File S1). Infections (the second most commonly recorded palaeopathology) have been recorded in equal measure to spondyloarthropathy (16% of the total), but only in titanosaurs from the Gondwana supercontinent (four records; Table 1; Figure 4; Supplementary File S1).
Finally, cystic lesions and tumors (8% of the total) and congenital malformations, amputations, fractures, and enthesophytes (4%) are the least common pathologies; these affect titanosaurs and sauropodomorphs (Figure 4; Table 1; Supplementary File S1). The records of these diseases are found in both supercontinents.
Titanosaurs are the sauropod clade with the highest number of diagnosed tail pathologies worldwide, with 69% of the total (Figure 4; Table 1; Supplementary File S1), and all these records are from Gondwana. Among the titanosaurs, the family Saltasauridae stands out as showing the greatest diversity of diseases, with three affecting the same individual [30]. But it is important to bear in mind that these results could be an artifact due to the fragmentary record of pathologies and/or titanosaurs, and the absence of extensive pathological descriptions of all sauropod bones, precise diagnoses, etc. (Table 1).
5.2. Paleobiological Implications of the Presence of Paleopathologies on Sauropod Tails
The size and shape of the tails of some clades of sauropod dinosaurs have always attracted attention, especially regarding the function they might fulfill beyond balance (e.g., the diplodocid flagellicaudatan sauropods and the use of their tails as whips, [53]). Studying the paleopathologies associated with tails could help to elucidate their possible functions, since the presence of some paleopathologies can affect the biomechanics of the tail, and this, in turn, is reflected in the paleopathology of the animal.
Taking into account that the fossil record of paleopathologies in sauropod tails is very scarce and fragmentary, trying to decipher their implications for biomechanics and paleoethology, i.e., for the functionality of their associated muscles, is very complex. To date, few explanations have been given. These few include the presence of male-female sexual dimorphism, the defensive use of the tail against predators (interspecific interactions), and the involvement of the tail during mating or aggressive interactions with conspecifics (intraspecific interactions) [40,53]. Accordingly, the paleobiological implications of the bone fusion that may result either from diffuse idiopathic skeletal hyperostosis (DISH) or from spondyloarthropathy have been associated with sexual dimorphism in male sauropods related to the posture mechanism, either due to rivalry during mating or compatibility [40,54,55]. It should be noted that diffuse idiopathic skeletal hyperostosis (DISH) and spondyloarthropathy diseases have similarities, and due to the fragmentary nature of the material, it is sometimes difficult to distinguish between them in the fossil record, as mentioned above.
In the present case study, it is possible that the musculature has been affected. If we compare it with closely related groups [Extant Phylogenetic Bracketing (EPB) approach] such as crocodiles and lepidosaurs [56], it can be inferred that the affected musculature would have been the hypaxial muscles of the tail, such as M. caudofemoralis longus and M. ilio-ischiocaudalis [56,57]. When the pathology is located in the insertion zone of the lateral aspect of the vertebral centrum next to the haemal arch (as in MAU-Pv-LI-601), the main muscle involved would be M. caudofemoralis longus. However, likely, spondyloarthropathy did not cause pain, implying that any reduction in the animal’s motion had a protective function [40]. On the other hand, M. caudofemoralis longus in MAU-Pv-LI-601 would not have been very developed in this area as is characteristic of titanosaurids, so it is reasonable to assume that the influence on its movement would not have been very great. However, in MAU-Pv-LJ-472/1, it is likely that the only muscle affected by the infection was the M. ilio-ischiocaudalis, since given the tentative location of the vertebra in the middle of the tail, it is most likely that the transition between the M. caudofemoralis longus and the M. ilio-ischiocaudalis had already occurred. In crocodiles, this muscle is involved in regulating the speed of locomotion, so it could have played a crucial role in escaping from a potential predator [58,59]. Regarding the possible pain in the case of infections, these injuries may have caused pain initially, probably restricting the mobility of the posterior end of the tail. Due to the fragmentary nature of the material, we do not know whether this lesion was the direct cause of the death of the titanosaur to which the vertebra MAU-Pv-LJ-472/1 belonged, but it is very likely that this pathology would have affected it in its daily life.
6. Conclusions
The results obtained in our analysis of the pathologies of two caudal vertebrae belonging to indeterminate titanosaurs from the Upper Cretaceous of North Patagonia indicate one case of spondyloarthropathy (MAU-Pv-LI-601) and one case of suppurative spinal osteomyelitis (MAU-Pv-LJ-472/1) The infection in MAU-Pv-LJ-472/1 is the second case of suppurative osteomyelitis in the supercontinent Gondwana. The spondyloarthropathy in MAU-Pv-LI-601 increases a problematic record which is characterized by the difficulty of distinguishing between diffuse idiopathic skeletal hyperostosis (DISH) and spondyloarthropathy pathologies when we are faced with fragmentary fossil records. Our study thus sheds new light on the paleopathologies to which Gondwanan titanosaurs were subjected and increases their record. As regards the specimens to which the caudal vertebrae under study belong, our analysis shows that in the case of MAU-Pv-LJ-472/1, the infection could have caused pain, discomfort, and probably restricted the mobility of the posterior end of the tail. In MAU-Pv-LI-601, by contrast, spondyloarthropathy did not cause pain or any reduction in mobility.
In addition, this work compiles for the first time the worldwide records of pathologies (diseases and traumas) present in the tails of sauropod dinosaurs throughout the Mesozoic, serving as a database for future paleobiological and paleoethological work.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d15030464/s1, File S1: Worldwide fossil record of paleopathologies in sauropod tails Supplementary Data.
Author Contributions
Conceptualization, P.C.-C.; methodology, P.C.-C. and J.G.-D.; software, J.G.-D.; CT scanning, L.S.F.; writing—original draft preparation, P.C.-C., L.S.F., J.G.-D. and J.I.C.; writing—review and editing, P.C.-C., L.S.F., J.G.-D. and J.I.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by [Spanish Ministerio de Ciencia e Innovación and the European Regional Development Fund (AEI/FEDER EU)] grant number [PID2021-122612OB-I00].
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The CT scan raw data of the vertebrae are available on MorphoSource (https://www.morphosource.org/projects/000502935?locale=en). All the CT scan raw data are available there upon request. Request for CT raw data should be addressed to Penélope Cruzado-Caballero.
Acknowledgments
We thank Martín Moyano, Director of the Clínica y Maternidad Eva Perón, Rincón de los Sauces (Neuquén Province, Argentina), the technician Norberto Rugoso for making possible the CT scanning of the specimen, and the Dra. Ariana Paulina-Carabajal assisted during the scanning work of MAU-Pv-LI-601 and MAU-Pv-LJ-472/1. We also thank two anonymous reviewers for improvements to this manuscript. The authors sincerely thank Rupert Glasgow, who revised the translation of the text into English.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sander, P.M.; Christian, A.; Clauss, M.; Fechner, R.; Gee, C.T.; Griebeler, E.M.; Gunga, H.C.; Hummel, J.; Mallison, H.; Perry, S.F.; et al. Biology of the sauropod dinosaurs: The evolution of gigantism. Biol. Rev. Camb. Philos. Soc. 2011, 86, 117–155. [Google Scholar] [CrossRef] [PubMed]
- Cerda, I.; Paulina Carabajal, A.; Salgado, L.; Coria, R.A.; Reguero, M.A.; Tambussi, C.P.; Moly, J.J. The first record of a sauropod dinosaur from Antarctica. Sci. Nat. 2012, 99, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Upchurch, P.; Barrett, P.M.; Dodson, P. Sauropoda. In The Dinosauria, 2nd ed.; Weishampel, D.B., Dodson, P., Osmólska, H., Eds.; University of California Press: Berkeley, CA, USA, 2004; pp. 295–322. [Google Scholar]
- Barrett, P.M.; Benson, R.B.J.; Upchurch, P. Dinosaur of Dorset: Part II, the Sauropod dinosaur (Saurischia, Sauropoda) with additional comments on the theropods. Proc. Dorset Nat. Hist. Archaeol. Soc. 2010, 131, 113–126. [Google Scholar]
- Bonaparte, J.F. The early radiation and phylogenetic relationships of the Jurassic sauropod dinosaurs, based on vertebral anatomy. In The Beginning of the Age of Dinosaurs; Padian, K., Ed.; Cambridge University Press: Cambridge, UK, 1986; pp. 247–258. [Google Scholar]
- Powell, J.E. Revision of South American Titanosaurid dinosaururs: Palaeobiological, palaeobiogeographical and phylogenetic aspects. In Records of the Queen Victoria Museum; Queen Victoria Museum and Art Gallery: Launceston, Australia, 2003; 173p. [Google Scholar]
- Mannion, P.D.; Upchurch, P.; Jin, X.; Zheng, W. New information on the Cretaceous sauropod dinosaurs of Zhejiang Province, China: Impact on Laurasian titanosauriform phylogeny and biogeography. R. Soc. Open Sci. 2019, 6, 191057. [Google Scholar] [CrossRef]
- Filippi, L.S.; Salgado, L.; Garrido, A.C. A new giant basal titanosaur sauropod in the Upper Cretaceous (Coniacian) of the Neuquén Basin, Argentina. Cretac. Res. 2019, 100, 61–81. [Google Scholar] [CrossRef]
- Hechenleitner, E.M.; Leuzinger, L.; Martinelli, A.G.; Rocher, S.; Fiorelli, L.E.; Taborda, J.R.A.; Salgado, L. Two Late Cretaceous sauropods reveal titanosaurian dispersal across South America. Commun. Biol. 2020, 3, 622. [Google Scholar] [CrossRef]
- Gallina, P.A.; Canale, J.L.; Carballido, J.L. The earliest known titanosaur sauropod dinosaur. Ameghiniana 2021, 58, 35–51. [Google Scholar] [CrossRef]
- Pérez-Moreno, A.; Carballido, J.l.; Otero, A.; Salgado, L.; Calvo, J.O. The axial skeleton of Rinconsaurus caudamirus (Sauropoda: Titanosauria) form the Late Cretaceous of Patagonia, Argentina. Ameghiniana 2022, 59, 1–46. [Google Scholar]
- Paulina-Carabajal, A. Neuroanatomy of titanosaurid dinosaurs from the Upper Cretaceous of Patagonia, with comments on endocranial variability within Sauropoda. Anat. Rec. 2012, 295, 2141–2156. [Google Scholar] [CrossRef]
- Paulina-Carabajal, A.; Filippi, L.S.; Knoll, F. Neuroanatomy of the titanosaurian sauropod Narambuenatitan palomoi from the Upper Cretaceous of Patagonia, Argentina. Publicación De La Electrónica De La Asoc. Paleontológica Argent. 2020, 20, 1–9. [Google Scholar]
- García, R.A.; Salgado, L.; Fernández, M.S.; Cerda, I.A.; Paulina Carabajal, A.; Otero, A.; Coria, R.A.; Fiorelli, L.E. Paleobiology of Titanosaurs: Reproduction, development, histology, pneumaticity, locomotion and neuroanatomy from the South American fossil record. Ameghiniana 2015, 52, 29–68. [Google Scholar] [CrossRef]
- Cerda, I.; Chinsamy, A.; Pol, D.; Apaldetti, C.; Otero, A.; Powell, J.E.; Martínez, R.N. Novel insight into the origin of the gorwth dynamic of saurpod dinosaurs. PLoS ONE 2017, 12, e0179707. [Google Scholar] [CrossRef]
- González, R.; Cerda, I.A.; Filippi, L.S.; Salgado, L. Early growth dynamics of titanosaur sauropods inferred from bone histology. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 537, 109404. [Google Scholar] [CrossRef]
- da Silva Vidal, L.; Luiz Gomes da Costa Pereira, P.V.; Tavares, S.; Brusatte, S.L.; Paglarelli Bergqvist, L.; dos Anjos Candeiro, C.R. Investigating the enigmatic Aeolosaurini clade: The caudal biomechanics of Aeolosaurus maximus (Aeolosaurini/Sauropoda) using the neutral pose method and the first case of protonic tail condition in Sauropoda. Hist. Biol. 2020, 33, 1836–1856. [Google Scholar] [CrossRef]
- Rothschild, B.M. Scientifically rigorous reptile and amphibian osseous pathology: Lessons for forensic herpetology from comparative and paleoepathology. Appl. Herpetol. 2009, 6, 47–79. [Google Scholar] [CrossRef]
- Arbour, V.M.; Currie, P.J. Tail and pelvis pathologies of ankylosaurian dinosaurs. Hist. Biol. 2011, 23, 375–390. [Google Scholar] [CrossRef]
- Rothschild, B.M.; Schultze, H.P.; Pellegrini, R. (Eds.) Herpetological Osteopathology: Annotated Bibliography of Amphibians and Reptiles; Springer Science: New York, NY, USA, 2012. [Google Scholar]
- Peterson, J.E.; Vittore, C.P. Cranial Pathologies in a Specimen of Pachycephalosaurus. PLoS ONE 2012, 7, e36227. [Google Scholar] [CrossRef]
- Tanke, D.H.; Rothschild, B.M. Paleopathology in Late Cretaceous Hadrosauridae from Alberta, Canada with comments on a putative Tyrannosaurus bite injury on an Edmontosaurus tail. In Hadrosaurs; Eberth, D.A., Evans, D.C., Eds.; Indiana University Press: Bloomington, IN, USA, 2014; pp. 540–572. [Google Scholar]
- Kappelman, J.; Ketcham, R.A.; Pearce, S.; Todd, L.; Akins, W.; Colbert, M.W.; Feseha, M.; Maisano, J.A.; Witzel, A. Perimortem fractures in Lucy suggest mortality from fall out of tall tree. Nature 2016, 537, 503. [Google Scholar] [CrossRef]
- Dumbrava, M.D.; Rothschild, B.M.; Weishampel, D.B.; Csiki-Sava, Z.; Andrei, R.A.; Acheson, K.A.; Codrea, V.A. A dinosaurian facial deformity and the first occurrence of ameloblastoma in the fossil record. Sci. Rep. 2016, 6, 29271. [Google Scholar] [CrossRef]
- Hearn, L.; Williams, A.C. Pain in dinosaurs: What is the evidence? Philos. Trans. R. Soc. B 2019, 374, 20190370. [Google Scholar] [CrossRef]
- Cruzado-Caballero, P.; Díaz-Martínez, I.; Rothschild, B.; Bedell, M.; Pereda-Suberbiola, X. A limping dinosaur in the Late: Pathologies in the pes of the neornithischian Othnielosaurus consors from the Morrison Formation (Upper Jurassic, USA). Hist. Biol. 2020, 33, 1753–1759. [Google Scholar] [CrossRef]
- Cruzado-Caballero, P.; Lecuona, A.; Cerda, I.A.; Díaz-Martínez, I. Osseous paleopathologies of Bonapartesaurus rionegrensis (Ornithopoda, Hadrosauridae) from Allen Formation (Upper Cretaceous) of Patagonia Argentina. Cretac. Res. 2021, 124, 104800. [Google Scholar] [CrossRef]
- Martinelli, A.G.; Teixeira, V.P.A.; Marinho, T.S.; Fonseca, P.H.M.; Cavellani, C.L.; Araujo, A.J.G.; Ribeiro, L.C.B.; Ferraz, M.L.F. Fused mid-caudal vertebrae in the titanosaur Uberabatitan ribeiroi from the Late Cretaceous of Brazil and other bone lesions. Lethaia 2015, 48, 456–462. [Google Scholar] [CrossRef]
- García, R.A.; Cerda, I.A.; Heller, M.; Rothschild, B.M.; Zurriaguz, V. The first evidence of osteomyelitis in a sauropod dinosaur. Lethaia 2016, 50, 227–236. [Google Scholar] [CrossRef]
- González, R.; Gallina, P.A.; Cerda, I.A. Multiple paleopathologies in the dinosaur Bonitasaura salgadoi (Sauropoda: Titanosauria) from the Upper Cretaceous of Patagonia, Argentina. Cretac. Res. 2017, 79, 159–170. [Google Scholar] [CrossRef]
- Clayton, R.J. Description of unusual pathological disorders on pubes and associated left femur from a Diplodocus specimen. Paludicola 2018, 11, 179–187. [Google Scholar]
- Tschopp, E.; Wings, O.; Frauenfelder, T.; Rothschild, B.M. Pathological phalanges in a camarasaurid sauropod dinosaur and implications on behaviour. Acta Palaeontol. Pol. 2016, 61, 125–134. [Google Scholar] [CrossRef]
- Brum, A.S.; Bandeira, K.L.; Holgado, B.; Souza, L.G.; Pêgas, R.V.; Sayão, J.M.; Campos, D.A.; Kellner, A.W. Palaeohistology and palaeopatology of an Aeolosaurini (Sauropoda: Titanosauria) from Morro do Cambambe (Upper Cretaceous, Brazil). [Paleohistología y paleopatología de un Aelosaurini (Sauropoda: Titanosauria) de Morro do Cambambe (Cretácico Superior, Brasil)]. Span. J. Palaeontol. 2021, 36, 1–17. [Google Scholar]
- Garrido, A. Estratigrafía del Grupo Neuquén, Cretácico Superior de la Cuenca Neuquina (Argentina): Nueva propuesta de ordenamiento litoestratigráfico. Rev. Del Mus. Argent. De Cienc. Nat. 2010, 12, 121–177. [Google Scholar] [CrossRef]
- Leanza, H.A.; Hugo, C.A. Cretaceous red beds from southern Neuquén Basin (Argentina): Age, distribution and stratigraphic discontinuities. Publicación Electrónica De La Asoc. Paleontológica Argent. 2001, 7, 117–122. [Google Scholar]
- Filippi, L.S.; García, R.A.; Garrido, A. A new titanosaur sauropod dinosaur from Upper Cretaceous of North Patagonia, Argentina. Acta Palaeontol. Pol. 2011, 56, 505–520. [Google Scholar] [CrossRef]
- Filippi, L.S.; Garrido, A. Pitekunsaurus macayai gen. et sp. nov., nuevo titanosaurio (Saurischia, Sauropoda) del Cretácico Superior de la Cuenca Neuquina, Argentina. Ameghiniana 2008, 45, 575–590. [Google Scholar]
- Gallina, P.A.; Apesteguía, S. Postcranial anatomy of Bonitasaura salgadoi (Sauropoda, Titanosauria) from the Late Cretaceous of Patagonia. J. Vertebr. Paléontol. 2015, 35, e924957. [Google Scholar] [CrossRef]
- Mahajan Mahajan, T.; Ganguly, S.; Para, P.A. Fracture Management in Animals: A Review. J. Chem. Biol. Phys. Sci. 2015, 5, 4053–4057. [Google Scholar]
- Rothschild, B.M.; Martin, L.D. Skeletal Impact of Disease: Bulletin 33. In New Mexico Museum of Natural History and Science; Authority of the State of New Mexico: Alburquerque, NM, USA, 2006; Volume 33. [Google Scholar]
- de Sousa Barbosa, F.H.; Porpino, K.D.O.; Rothschild, B.M.; Silva, R.C.D.; Capone, D. First cancer in an extinct Quaternary non-human mammal. Hist. Biol. 2020, 33, 2878–2882. [Google Scholar] [CrossRef]
- Jacobson, E.R. (Ed.) Infectious Diseases and Pathology of Reptiles. In Viruses and Viral Diseases of Reptiles; CRC Press: Boca Raton, FL, USA, 2007; pp. 395–460. [Google Scholar]
- Anné, J.; Garwood, R.J.; Lowe, T.; Withers, P.J.; Manning, P.L. Interpreting pathologies in extant and extinct archosaurs using micro-CT. PeerJ 2015, 3, e1130. [Google Scholar] [CrossRef]
- Hanna, R.R. Multiple injury and infection in a sub-adult theropod dinosaur Allosaurus fragilis with comparisons to Allosaurus pathology in the Clevland-Lloyd Quarry collection. J. Vertebr. Paleontol. 2002, 22, 76–90. [Google Scholar] [CrossRef]
- Ramírez-Velasco, A.A.; Morales-Salinas, E.; Hernández-Rivera, R.; Tanke, D.H. Spinal and rib osteopathy in Huehuecanauhtlus tiquichensis (Ornithopoda: Hadrosauroidea) from the Late Cretaceous in Mexico. Hist. Biol. 2017, 29, 208–222. [Google Scholar] [CrossRef]
- Resnick, D.; Niwayama, G. Diagnosis of Bone and Joint Disorders; WB Saunders Co.: Philadelphia, PA, USA, 1981; Volume 3, pp. 2058–2079. [Google Scholar]
- de Souza Barbosa, F.H.; da Costa Ribeiro, I.; da Costa, P.V.L.G.; Bergqvist, L.P. Vertebral lesions in a titanosaurian dinosaur from the Lower-Upper Cretaceous of Brazil. Geobios 2018, 51, 385–389. [Google Scholar] [CrossRef]
- Mader, R.; Sarzi-Puttini, P.; Atzeni, F.; Olivieri, I.; Pappone, N.; Verlaan, J.J.; Buskila, D. Extraspinal manifestations of diffuse idiopathic skeletal hyperostosis. Rheumatology 2009, 48, 1478–1481. [Google Scholar] [CrossRef]
- Cisneros, J.C.; Gomes Cabral, U.; de Beer, F.; Damiani, R.; Costa Fortier, D. Spondarthritis in the Triassic. PLoS ONE 2010, 5, e13425. [Google Scholar] [CrossRef] [PubMed]
- Witzmann, F.; Schwarz-Wings, D.; Hampe, O.; Fritsch, G.; Asbach, P. Evidence of spondyloarthropathy in the spine of a phytosaur (Reptilia: Archosauriformes) from the Late Triassic of Halberstadt, Germany. PLoS ONE 2014, 9, e85511. [Google Scholar] [CrossRef] [PubMed]
- Gallina, P.A. Histología ósea del titanosaurio Bonitasaura salgadoi (Dinosauria: Sauropoda) del Cretácico superior de Patagonia. Ameghiniana 2012, 49, 289–302. [Google Scholar] [CrossRef]
- Rothschild, B.M.; Berman, D.S. Fusion of caudal vertebrae in Late Jurassic sauropods. J. Vertebr. Paleontol. 1991, 11, 29–36. [Google Scholar] [CrossRef]
- Conti, S.; Tschopp, E.; Mateus, O.; Zanoni, A.; Masarati, P.; Sala, G. Multibody analysis and soft tissue strength refute supersonic dinosaur tail. Sci. Rep. 2022, 12, 19245. [Google Scholar] [CrossRef] [PubMed]
- Butler, R.J.; Yates, A.M.; Rauhut, O.W.; Foth, C. A pathological tail in a basal sauropodomorph dinosaur from South Africa: Evidence of traumatic amputation? J. Vertebr. Paleontol. 2013, 33, 224–228. [Google Scholar] [CrossRef]
- Xing, L.; Rothschild, B.M.; Ran, H.; Miyashita, T.; Persons, W.S.; Sekiya, T.; Zhang, J.; Tao Wang, T.; Dong, Z. Vertebral fusion in two Early Jurassic sauropodomorph dinosaurs from the Lufeng Formation of Yunnan, China. Acta Palaeontol. Pol. 2015, 60, 643–649. [Google Scholar] [CrossRef][Green Version]
- Weston, D.A. Nonspecific infection in paleopathology: Interpreting periosteal reactions. In A Companion to Paleopathology; Grauer, A.L., Ed.; Wiley-Blackwell: Oxford, UK, 2012; pp. 492–512. [Google Scholar]
- Díez Díaz, V.; Demuth, O.E.; Schwarz, D.; Mallison, H. The tail of the Late Jurassic sauropod Giraffatitan brancai: Digital reconstruction of its epaxial and hypaxial musculature, and implications for tail biomechanics. Front. Earth Sci. 2020, 8, 160. [Google Scholar] [CrossRef]
- Ibiricu, L.M.; Lamanna, M.C.; Lacovara, K.J. The influence of caudofemoral musculature on the titanosaurian (Saurischia: Sauropoda) tail skeleton: Morphological and phylogenetic implications. Hist. Biol. 2014, 26, 454–471. [Google Scholar] [CrossRef]
- Otero, A.; Gallina, P.A.; Canale, J.I.; Haluza, A. Sauropod haemal arches: Morphotypes, new classification and phylogenetic aspects. Hist. Biol. 2012, 24, 243–256. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).