


Genetic Diversity and Population Structure of the Slender Racer (Orientocoluber spinalis) in South Korea

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
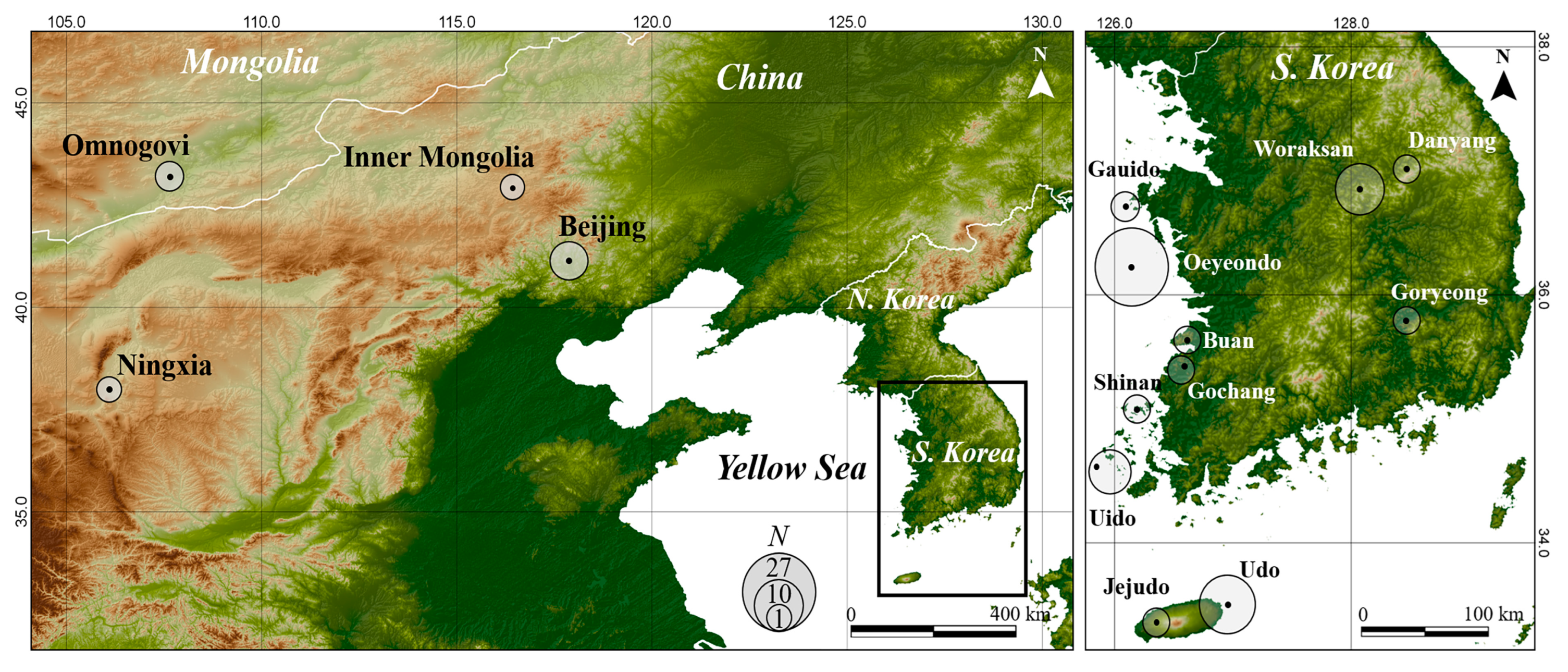
2.1. Sample Collection
2.2. PCR Amplification and Sequencing
2.3. Population Genetic Analysis
3. Results
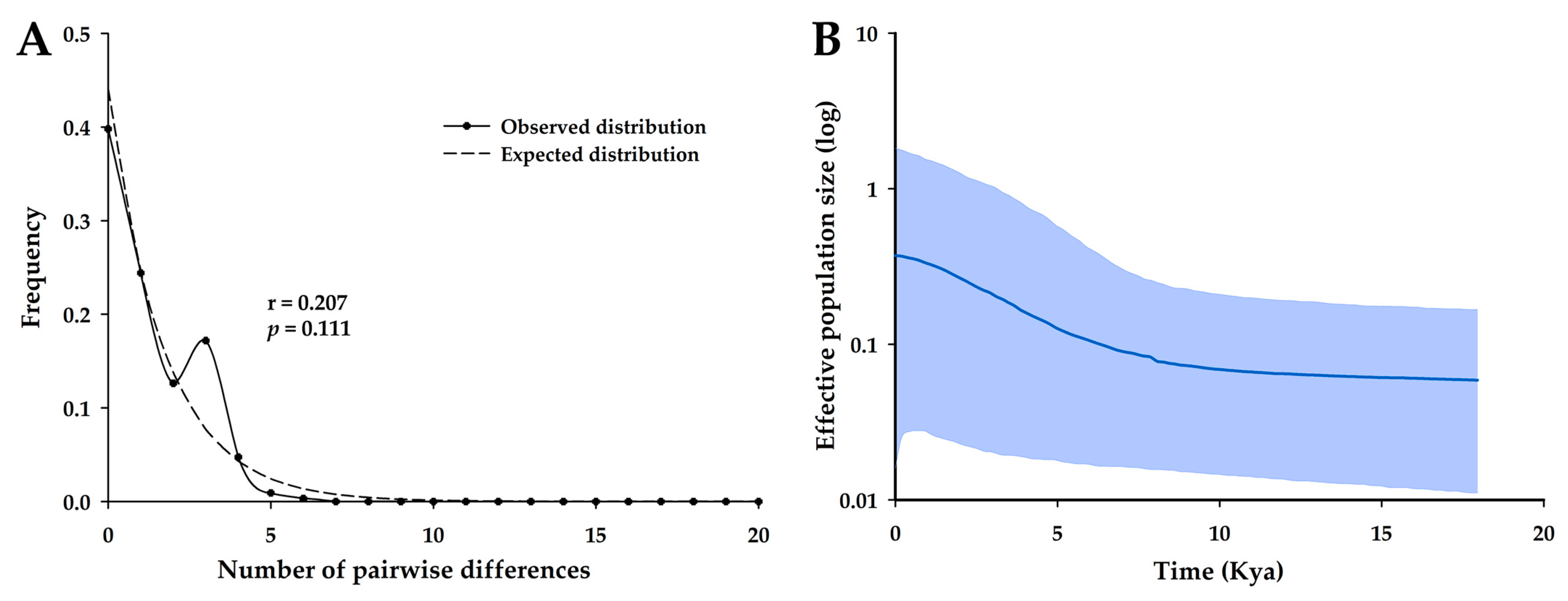
3.1. Genetic Diversity
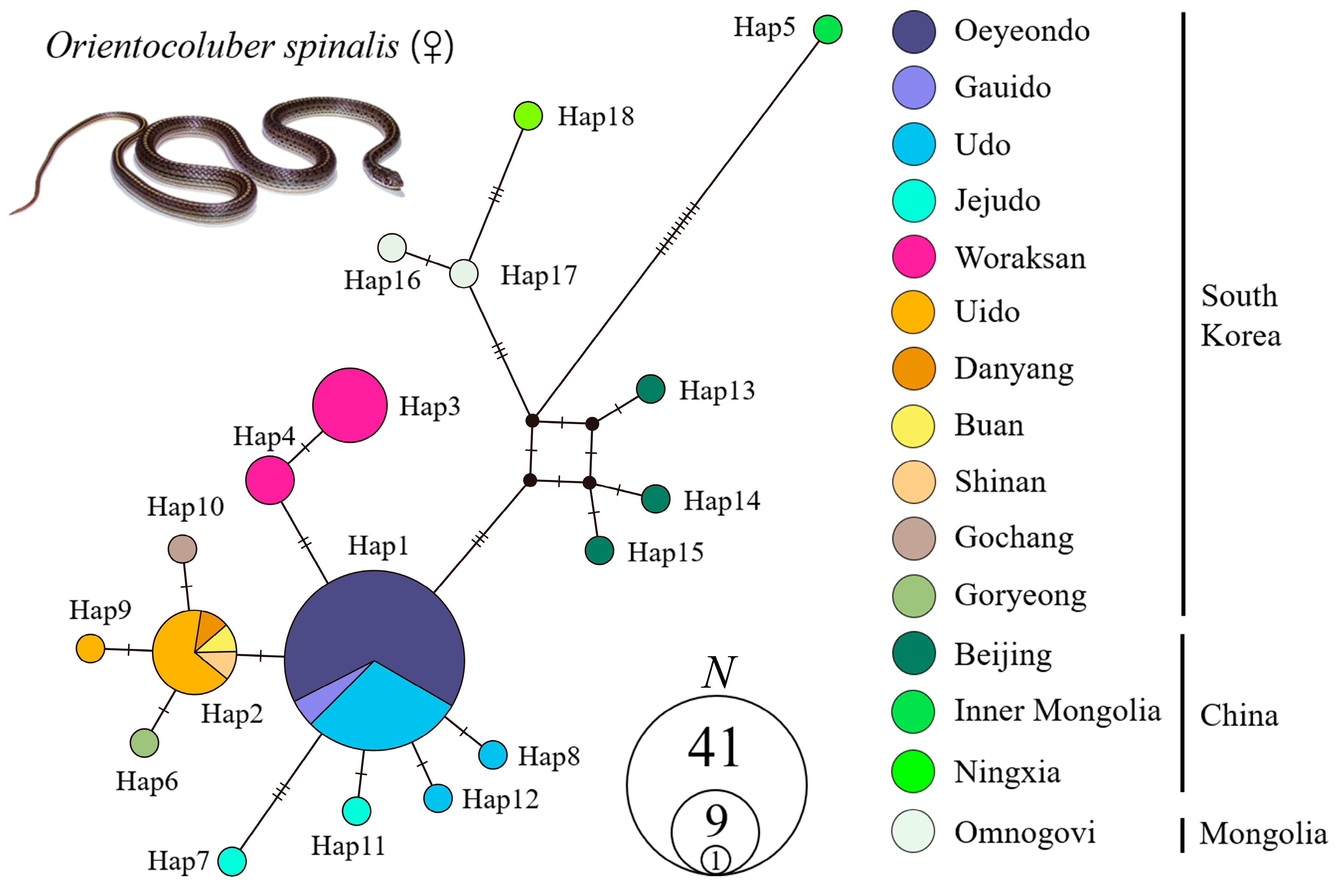
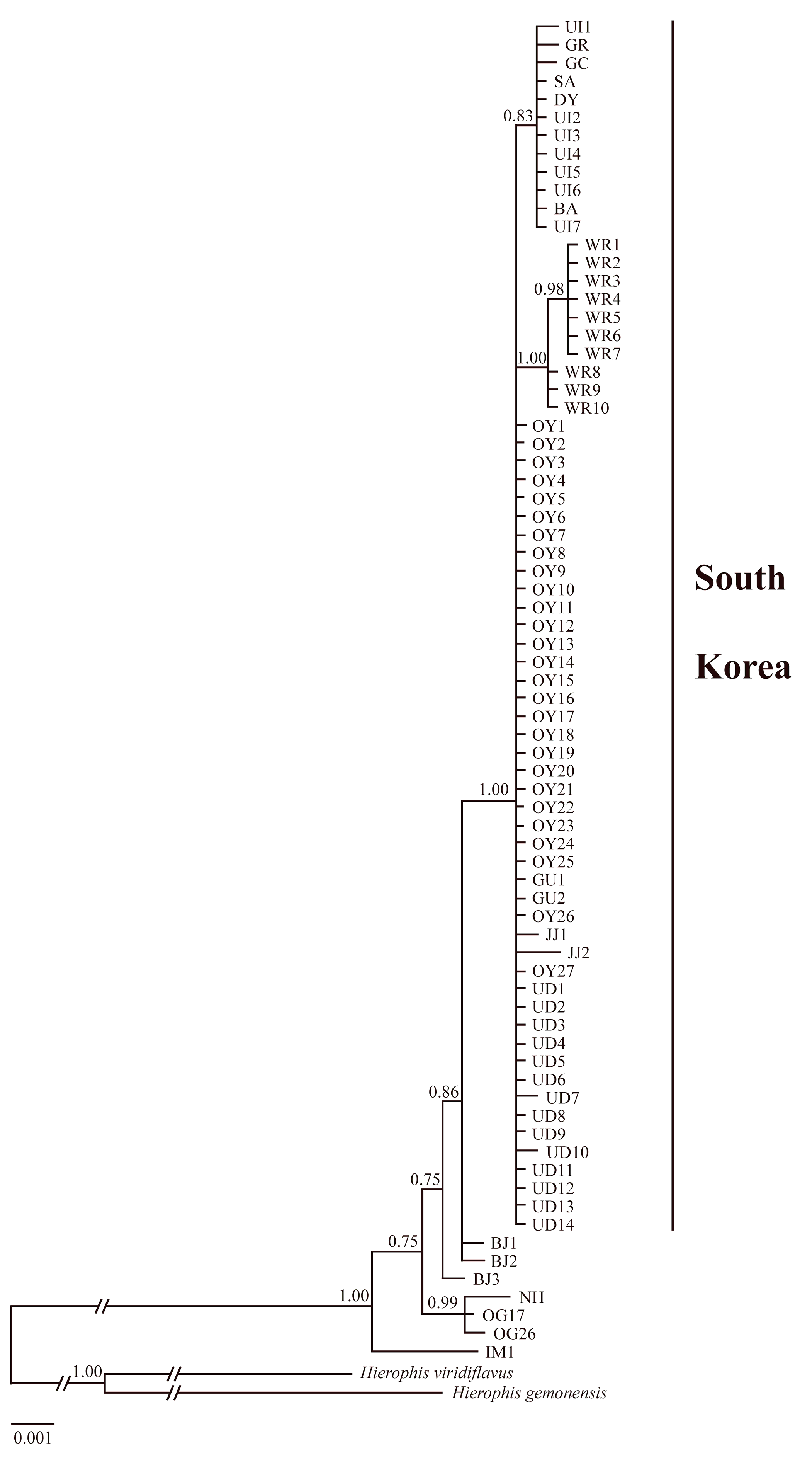
3.2. Genetic Structure
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Drury, W.H. Rare species. Biol. Conserv. 1974, 6, 162–169. [Google Scholar] [CrossRef]
- Cunningham, R.B.; Lindenmayer, D.B. Modeling count data of rare species: Some statistical issues. Ecology 2005, 5, 1135–1142. [Google Scholar] [CrossRef]
- Sodhi, N.S.; Brook, B.W.; Bradshaw, C.A.J. Causes and Consequences of Species Extinctions. In The Princeton Guide to Ecology, 1st ed.; Simon, A.L., Stephen, R., Carpenter, H., Charles, J.G., Ann, P.K., Michel, L., Jonathan, B.L., Brian, W., David, S.W., Eds.; Princeton University Press: Princeton, NJ, USA, 2009; pp. 520–541. ISBN 978-0691156040. [Google Scholar]
- Canales-Delgadillo, J.C.; Scott-Morales, L.; Korb, J. The influence of habitat fragmentation on genetic diversity of a rare bird species that commonly faces environmental fluctuations. J. Avian Biol. 2012, 43, 168–176. [Google Scholar] [CrossRef]
- Vermeij, G.J.; Grosberg, R.K. Rarity and persistence. Ecol. Lett. 2018, 21, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Frankham, R.; Ballou, J.D.; Ralls, K.; Eldridge, M.D.E.; Dudash, M.R.; Fenster, C.B.; Lacy, R.C.; Sunnucks, P. Genetic Management of Fragmented Animal and Plant Populations, 1st ed.; Oxford University Press: New York, NY, USA, 2017; ISBN 978-0198783404. [Google Scholar]
- DeWoody, J.A.; Harder, A.M.; Mathur, S.; Willoughby, J.R. The long-standing significance of genetic diversity in conservation. Mol. Ecol. 2021, 30, 4147–4154. [Google Scholar] [CrossRef]
- Gómez-Sánchez, D.; Olalde, I.; Sastre, N.; Enseñat, C.; Carrasco, R.; Marques-Bonet, T.; Lalueza-Fox, C.; Leonard, J.A.; Vilà, C.; Ramírez, O. On the path to extinction: Inbreeding and admixture in a declining grey wolf population. Mol. Ecol. 2017, 27, 3599–3612. [Google Scholar] [CrossRef] [PubMed]
- Khosravi, R.; Malekian, M.; Hemami, M.R.; Silva, T.L.; Brito, J.C. Low genetic diversity in the vulnerable Goitred gazelle, Gazella subgutturosa (Cetartiodactyla: Bovidae), in Iran: Potential genetic consequence of recent population declines. Zool. Middle East 2019, 65, 104–115. [Google Scholar] [CrossRef]
- Wood, D.A.; Rose, J.P.; Halstead, B.J.; Stoelting, R.E.; Swaim, K.E.; Vandergast, A.G. Combining genetic and demographic monitoring better informs conservation of an endangered urban snake. PLoS ONE 2020, 15, e0231744. [Google Scholar] [CrossRef]
- Keyghobadi, N.; Crawford, L.A.; Desjardins, S. High genetic drift in endangered northern peripheral populations of the Behr’s hairstreak butterfly (Satyrium behrii). Insect. Syst. Divers. 2021, 14, 403–411. [Google Scholar] [CrossRef]
- Soltis, P.S.; Gitzendanner, M.A. Molecular systematics and the conservation of rare species. Conserv. Biol. 1999, 13, 471–483. [Google Scholar] [CrossRef]
- Steiner, C.C.; Putnam, A.S.; Hoeck, P.E.A.; Ryder, O.A. Conservation genomics of threatened animal species. Annu. Rev. Anim. Biosci. 2013, 1, 261–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hohenlohe, P.A.; Funk, W.C.; Rajora, O.P. Population genomics for wildlife conservation and management. Mol. Ecol. 2021, 30, 62–82. [Google Scholar] [CrossRef] [PubMed]
- Ruane, S.; Austin, C.C. Phylogenomics using formalin-fixed and 100+ year-old intractable natural history specimens. Mol. Ecol. Resour. 2017, 17, 1003–1008. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.C.; Messenger, K.R.; Koo, K.S.; Lee, S.C.; Hou, M.; Borzée, A. How threatened is Scincella huanrenensis? An update on threats and trends. Conservation 2021, 1, 58–72. [Google Scholar] [CrossRef]
- Pilliod, D.S.; Hallock, L.A.; Miller, M.P.; Mullins, T.D.; Haig, S.M. Conservation genetics of the imperiled Striped whipsnake in Washington, USA. Herpetol. Conserv. Biol. 2020, 15, 597–610. [Google Scholar]
- Shaffer, H.B.; Gidiş, M.; McCartney-Melstad, E.; Neal, K.M.; Oyamaguchi, H.M.; Tellez, M.; Toffelmier, E.M. Conservation genetics and genomics of amphibians and reptiles. Annu. Rev. Anim. Biosci. 2015, 3, 113–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.Y. Current status and distribution of reptiles in the Republic of Korea. Korean J. Environ. Biol. 2007, 25, 124–138. [Google Scholar]
- Macias, D.; Shin, Y.C.; Borzée, A. An update on the conservation status and ecology of Korean terrestrial squamates. J. Nat. Conserv. 2021, 60, 125971. [Google Scholar] [CrossRef]
- Kharin, V.E.; Akulenko, M.V. Rare and little-known snakes in North-Eastern Eurasia. 1. On a new record of Slender racer–Hierophis spinalis (Colubridae) in the Russian Far East. Curr. Stud. Herpetol. 2008, 8, 160–169. [Google Scholar]
- Park, I.K.; Borzée, A.; Park, J.J.; Min, S.H.; Zhang, Y.P.; Li, S.R.; Park, D.S. Past, present, and future predictions on the suitable habitat of the Slender racer (Orientocoluber spinalis) using species distribution models. Ecol. Evol. 2022, 12, e9169. [Google Scholar] [CrossRef]
- Kim, L.T.; Han, G.H. Animal of Chosun: Amphibian and Reptiles; Science and Technology Publisher: Pyongyang, Republic of Korea, 2009; p. 138. ISBN 978-9946-1-0065-4. [Google Scholar]
- Park, I.K.; Park, J.J.; Min, S.H.; Grajal-Puche, A.; Park, D.S. Predation of the Japanese keelback (Hebius vibakari Boie, 1826) by the Slender racer (Orientocoluber spinalis Peters, 1866). J. Ecol. Environ. 2021, 45, 19. [Google Scholar] [CrossRef]
- Nagy, Z.T.; Lawson, R.; Joger, U.; Wink, M. Molecular systematics of racers, whipsnakes, and relatives (Reptilia: Colubridae) using mitochondrial and nuclear markers. J. Zool. Syst. Evol. Res. 2004, 42, 223–233. [Google Scholar] [CrossRef]
- Schätti, B.; Monsch, P. Systematics and phylogenetic relationships of whip snakes (Hierophis Fitzinger) and Zamenis andreana Werner, 1917 (Reptilia: Squamata: Colubrinae). Rev. Suisse Zool. 2004, 111, 239–256. [Google Scholar] [CrossRef]
- Kharin, V.E. Rare and little-known snakes of the north-eastern Eurasia. 3. On the taxonomic status of the Slender racer Hierophis spinalis (Serpentes: Colubridae). Curr. Stud. Herpetol. 2011, 11, 173–179. [Google Scholar]
- Maslova, I.V.; Akulenko, M.V.; Portnyagina, E.Y.; Pokhiyuk, N.E.; Rogashevskaya, D.A. Rare and endangered amphibians and reptiles of Primorsky Krai (Russian Far East). Biota. Environ. Nat. Areas. 2021, 41, 102–121. [Google Scholar] [CrossRef] [PubMed]
- Terbish, K.H.; Munkhbayar, K.H.; Clark, E.L.; Munkhbat, J.; Monks, E.M. Mongolian red list of reptiles and amphibians. In Regional Red List Series; Munkhbaatar, M., Baillie, J.E.M., Borkin, L., Batsaikhan, N., Samiya, R., Semenov, D.V., Eds.; Zoological Society of London: London, UK, 2011; Volume 5, p. 50. [Google Scholar]
- Kubykin, R.A.; Zima, Y.A. Animals. In Red Book of the Republic of Kazakhstan; Konzhyk: Almaty, Kazakhstan, 2010; Volume 1, pp. 74–75. [Google Scholar]
- National Institute of Biological Resources. Amphibians and Reptiles. In Red Data Book of Republic of Korea; National Institute of Biological Resources: Incheon, Republic of Korea, 2019; Volume 2. [Google Scholar]
- Park, J.J.; Park, I.K.; Ra, N.Y.; Min, S.H.; Park, D.S. Complete mitochondrial genome of the Slender racer (Orientocoluber spinalis Peters, 1866; Squamata, Colubridae). Mitochondrial DNA B Resour. 2020, 5, 2693–2694. [Google Scholar] [CrossRef]
- Burbrink, F.T.; Lawson, R.; Slowinski, J.B. Mitochondrial DNA phylogeography of the polytypic North American rat snake (Elaphe obsoleta): A critique of the subspecies concept. Evolution 2000, 54, 2107–2118. [Google Scholar] [CrossRef] [PubMed]
- Fontanella, F.M.; Feldman, C.R.; Siddall, M.E.; Burbrink, F.T. Phylogeography of Diadophis punctatus: Extensive lineage diversity and repeated patterns of historical demography in a trans-continental snake. Mol. Phylogenet. Evol. 2008, 46, 1049–1070. [Google Scholar] [CrossRef] [PubMed]
- Mezzasalma, M.; Dall’Asta, A.; Loy, A.; Cheylan, M.; Lymberakis, P.; Zuffi, M.A.; Tomović, L.; Odierna, G.; Guarino, F.M. A sisters’ story: Comparative phylogeography and taxonomy of Hierophis viridiflavus and H. gemonensis (Serpentes, Colubridae). Zool. Scr. 2015, 44, 495–508. [Google Scholar] [CrossRef]
- Meier, N.; Dubey, S.; Glaizot, O.; Schmitz, A.; Zambelli, N.; Ursenbacher, S. Where are you from? Origin determination of the introduced Green whipsnake, Hierophis viridiflavus (Squamata: Colubridae), in Switzerland. Herpetol. Notes 2022, 15, 335–344. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadagkar, S.R.; Rosenberg, M.S.; Kumar, S. Inferring species phylogenies from multiple genes: Concatenated sequence tree versus consensus gene tree. J. Exp. Zool. B Mol. Dev. Evol. 2005, 304, 64–74. [Google Scholar] [CrossRef]
- Rozas, J.; Sánchez-DelBarrio, J.C.; Messeguer, X.; Rozas, R. DnaSP, DNA polymorphism analyses by the coalescent and other methods. Bioinformatics 2003, 19, 2496–2497. [Google Scholar] [CrossRef] [Green Version]
- Petit, R.J.; Mousadik, A.E.; Pons, O. Identifying populations for conservation on the basis of genetic markers. Conserv. Biol. 1998, 12, 844–855. [Google Scholar] [CrossRef]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef]
- Fu, Y.X. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef]
- Fu, Y.X.; Li, W.H. Statistical tests of neutrality of mutations. Genetics 1993, 133, 693–709. [Google Scholar] [CrossRef]
- Harpending, H.C.; Sherry, S.T.; Rogers, A.R.; Stoneking, M. The genetic structure of ancient human populations. Curr. Anthropol. 1993, 34, 483–496. [Google Scholar] [CrossRef]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; de Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, R.P.; Yang, Z. Rate variation and estimation of divergence times using strict and relaxed clocks. BMC Evol. Biol. 2011, 11, 271. [Google Scholar] [CrossRef] [Green Version]
- Ho, S.Y.; Duchêne, S. Molecular-clock methods for estimating evolutionary rates and timescales. Mol. Ecol. 2014, 23, 5947–5965. [Google Scholar] [CrossRef]
- Daza, J.M.; Smith, E.N.; Páez, V.P.; Prkinson, C.L. Complex evolution in the neotropics: The origin and diversification of the widespread genus Leptodeira (Serpentes: Colubridae). Mol. Phylogenet. Evol. 2009, 53, 653–667. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Ho, S.Y.W.; Phillips, M.J.; Rambaut, A. Relaxed phylogenetics and dating with confidence. PLoS Biol. 2006, 4, e88. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. FigTree 1.4.4. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 3 September 2022).
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Posada, D.; Crandall, K.A. MODELTEST: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [Green Version]
- Posada, D.; Buckley, T.R. Model selection and model averaging in phylogenetics: Advantages of Akaike information criterion and Bayesian approaches over likelihood ratio tests. Syst. Biol. 2004, 53, 793–808. [Google Scholar] [CrossRef] [PubMed]
- Avella, I.; Castiglia, R.; Senczuk, G. Who are you? The genetic identity of some insular populations of Hierophis viridiflavus s. l. from the Tyrrhenian Sea. Acta Herpetol. 2017, 12, 209–214. [Google Scholar] [CrossRef]
- Machado, L.; Harris, D.J.; Salvi, D. Biogeographic and demographic history of the Mediterranean snakes Malpolon monspessulanus and Hemorrhois hippocrepis across the strait of Gibraltar. BMC Ecol. Evol. 2021, 21, 210. [Google Scholar] [CrossRef]
- Case, T.J.; Bolger, D.T.; Richman, A.D. Reptilian extinctions: The last ten thousand years. In Conservation Biology; Fiedler, P.L., Jain, S.K., Eds.; Springer: Boston, MA, USA, 1992; pp. 91–125. ISBN 978-0412019517. [Google Scholar]
- Frankham, R. Do island populations have less genetic variation than mainland populations? Heredity 1997, 78, 311–327. [Google Scholar] [CrossRef] [Green Version]
- Hoorn, C.; Mosbrugger, V.; Mulch, A.; Antonelli, A. Biodiversity from mountain building. Nat. Geosci. 2013, 6, 154. [Google Scholar] [CrossRef] [Green Version]
- Jablonski, D.; Nagy, Z.T.; Avcı, A.; Olgun, K.; Kukushkin, O.V.; Safaei-Mahroo, B.; Jandzik, D. Cryptic diversity in the smooth snake (Coronella austriaca). Amphibia-Reptilia 2019, 40, 179–192. [Google Scholar] [CrossRef]
- Koo, H.J.; Cho, H.G. Changes in detrital sediment supply to the central Yellow Sea since the last deglaciation. Ocean Sci. 2020, 16, 1247–1259. [Google Scholar] [CrossRef]
- Lee, B.R.; Yoo, D.G.; Lee, G.S. High-resolution sequence stratigraphy and evolution of the Jeju strait shelf, Korea, since the last glacial maximum. Mar. Pet. Geol. 2022, 135, 105389. [Google Scholar] [CrossRef]
- Rogers, A.R.; Harpending, H. Population growth makes waves in the distribution of pairwise genetic differences. Mol. Biol. Evol. 1992, 9, 552–569. [Google Scholar] [CrossRef]
- Shannon, F.A. The reptiles and amphibian of Korea. Herpetologica 1956, 12, 22–49. [Google Scholar]
- Ray, N.; Adams, J. A GIS-based vegetation map of the world at the last glacial maximum (25,000-15,000 BP). Internet Archaeol. 2001, 11. [Google Scholar] [CrossRef] [Green Version]
- d’Alpoim Guedes, J.; Austerman, J.; Mitrovica, J.X. Lost foraging opportunities for east Asian hunter-gatherers due to rising sea level since the last glacial maximum. Geoarchaeology 2016, 31, 255–266. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality ID | Country | Locality | No. of Samples | Haplotype | GenBank Accession Number | |
|---|---|---|---|---|---|---|
| Cytb | ND4 | |||||
| KR1 | South Korea | Oeyeondo, Chungnam | 27 | Hap1 | OQ290820 | OQ290083 |
| KR2 | Udo, Jeju | 14 | Hap1 Hap8 Hap12 | OQ290820 OQ290824 | OQ290837, OQ290838 | |
| KR3 | Woraksan, Chungbuk | 10 | Hap3 Hap4 | OQ290816 NC049067 | OQ290825, OQ290083, NC049067 | |
| KR4 | Uido, Jeonnam | 7 | Hap2 Hap9 | OQ290823 | OQ290836, OQ290825, OQ290083 | |
| KR5 | Jejudo, Jeju | 2 | Hap7 Hap11 | OQ290820 OQ290822 | OQ290083, OQ290831, OQ290835 | |
| KR6 | Gauido, Chungnam | 2 | Hap1 | OQ290820 | OQ290083 | |
| KR7 | Buan, Jeonbuk | 1 | Hap2 | OQ290823 | OQ290083 | |
| KR8 | Danyang, Chungbuk | 1 | Hap2 | OQ290823 | OQ290083 | |
| KR9 | Gochang, Jeonbuk | 1 | Hap10 | OQ290817 | OQ290083 | |
| KR10 | Goryeong, Gyeongbuk | 1 | Hap6 | OQ290815 | OQ290083 | |
| KR11 | Shinan, Jeonnam | 1 | Hap2 | OQ290823 | OQ290083 | |
| CN1 | China | Beijing | 3 | Hap13 | OQ290818 | OQ290826, OQ290827, OQ290828 |
| CN2 | Inner Mongolia Autonomous Region | 1 | Hap14 | OQ290819 | OQ290829, OQ290834 | |
| CN3 | Ningxia Hui Autonomous Region | 1 | Hap18 | AY486924 | AY487056 | |
| MN1 | Mongolia | Omnogovi | 2 | Hap16 Hap17 | OQ290821 | OQ290832, OQ290833 |
| Total | 74 | |||||
| Country | South Korea | China | Mongolia | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Haplotype | OY | UD | WR | UI | JJ | GU | BA | DY | GC | GR | SA | BJ | IM | NH | OG |
| Hap1 | 27 | 12 | 2 | ||||||||||||
| Hap2 | 6 | 1 | 1 | 1 | |||||||||||
| Hap3 | 7 | ||||||||||||||
| Hap4 | 3 | ||||||||||||||
| Hap5 | 1 | ||||||||||||||
| Hap6 | 1 | ||||||||||||||
| Hap7 | 1 | ||||||||||||||
| Hap8 | 1 | ||||||||||||||
| Hap9 | 1 | ||||||||||||||
| Hap10 | 1 | ||||||||||||||
| Hap11 | 1 | ||||||||||||||
| Hap12 | 1 | ||||||||||||||
| Hap13 | 1 | ||||||||||||||
| Hap14 | 1 | ||||||||||||||
| Hap15 | 1 | ||||||||||||||
| Hap16 | 1 | ||||||||||||||
| Hap17 | 1 | ||||||||||||||
| Hap18 | 1 | ||||||||||||||
| Total | 1 | 3 | 2 | 2 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 3 | 1 | 1 | 2 |
| Population | N | NH | Ps | h ± SD | π ± SD | HR | D | FS | FD* | FF* |
|---|---|---|---|---|---|---|---|---|---|---|
| Oeyeondo | 27 | 1 | 0 | 0.000 ± 0.000 | 0.0000 ± 0.0000 | 0.000 | NA | 0.000 | 0.000 | 0.000 |
| Udo | 14 | 3 | 2 | 0.275 ± 0.148 | 0.0002 ± 0.0001 | 1.000 | −1.481 | −1.475 | −1.827 | −1.827 |
| Woraksan | 10 | 2 | 1 | 0.467 ± 0.132 | 0.0003 ± 0.0001 | 0.992 | 0.820 | 0.818 | 0.804 | 0.898 |
| Uido | 7 | 2 | 1 | 0.286 ± 0.196 | 0.0002 ± 0.0001 | 1.000 | −1.006 | −0.095 | −1.049 | −1.101 |
| Combined Korean population | 67 | 11 | 12 | 0.602 ± 0.064 | 0.0007 ± 0.0001 | −1.398 | −4.557 † | −3.153 † | −2.874 † |
| Country | South Korea | ||||
|---|---|---|---|---|---|
| Population | Oyen (n = 27) | Udo (n = 14) | Woraksan (n = 10) | Uido (n = 7) | |
| South Korea | Oeyeondo | ||||
| Udo | 0.01 | ||||
| Woraksan | 0.15 | 0.16 | |||
| Uido | 0.06 | 0.07 | 0.20 | ||
| China and Mongolia | Combined foreign population | 0.42 | 0.43 | 0.57 | 0.48 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, H.; Park, I.-K.; Kim, J.; Park, J.; Lee, S.-C.; Park, D. Genetic Diversity and Population Structure of the Slender Racer (Orientocoluber spinalis) in South Korea. Diversity 2023, 15, 543. https://doi.org/10.3390/d15040543
Jeong H, Park I-K, Kim J, Park J, Lee S-C, Park D. Genetic Diversity and Population Structure of the Slender Racer (Orientocoluber spinalis) in South Korea. Diversity. 2023; 15(4):543. https://doi.org/10.3390/d15040543
Chicago/Turabian StyleJeong, Hojun, Il-Kook Park, Jongsun Kim, Jaejin Park, Sang-Cheol Lee, and Daesik Park. 2023. "Genetic Diversity and Population Structure of the Slender Racer (Orientocoluber spinalis) in South Korea" Diversity 15, no. 4: 543. https://doi.org/10.3390/d15040543
APA StyleJeong, H., Park, I.-K., Kim, J., Park, J., Lee, S.-C., & Park, D. (2023). Genetic Diversity and Population Structure of the Slender Racer (Orientocoluber spinalis) in South Korea. Diversity, 15(4), 543. https://doi.org/10.3390/d15040543