

Vanilla planifolia Andrews (Orchidaceae): Labellum Variation and Potential Distribution in Hidalgo, Mexico

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Geographic Location
2.2. Species Distribution
2.3. Species Distribution Modeling
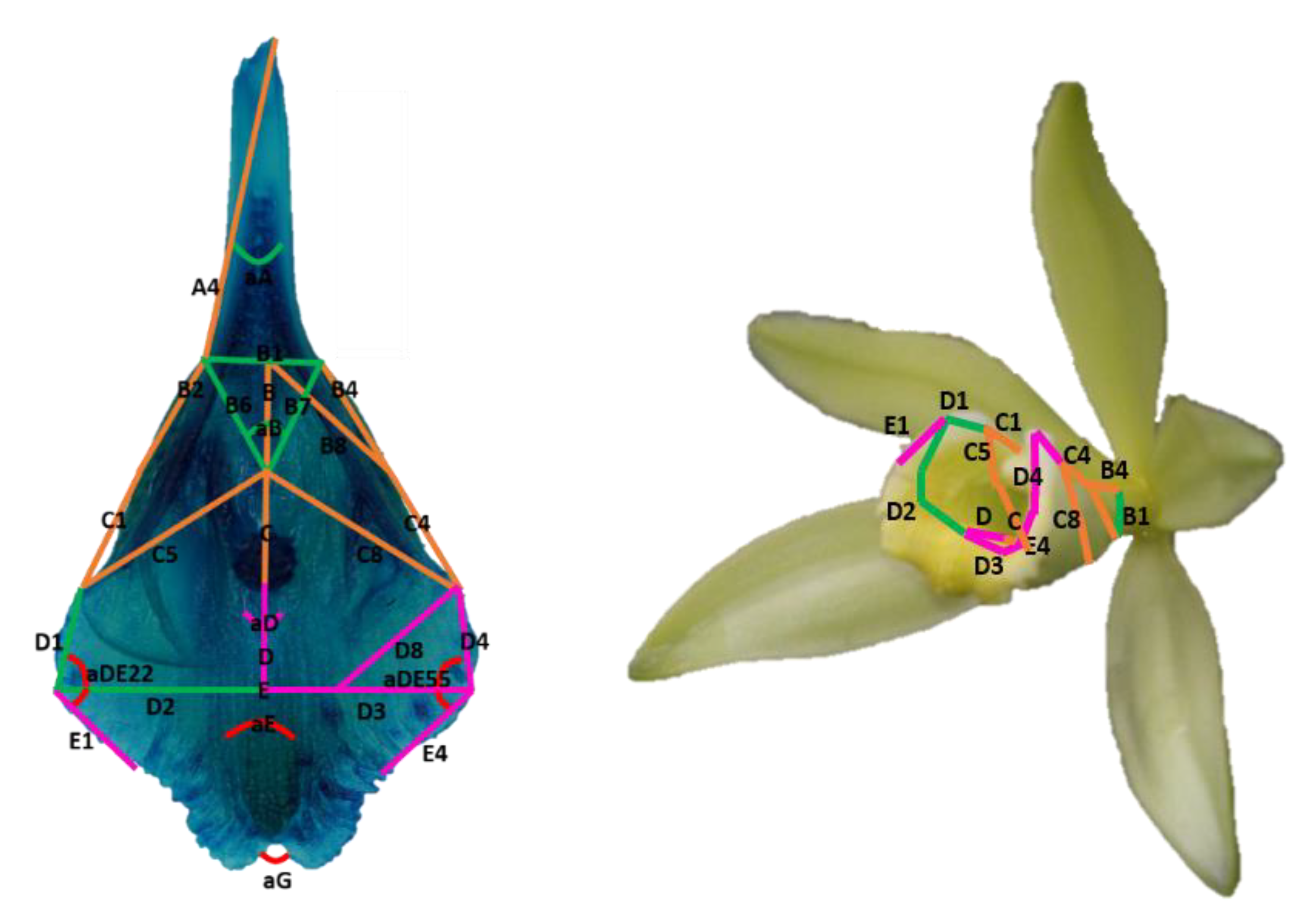
2.4. Morphological Characterization of the Flower
2.5. Statistical and Numerical Analysis
3. Results
3.1. Potential Distribution of Vanilla planifolia Andrews
Location of the Populations of Vanilla planifolia Andrews
3.2. Potential Distribution
3.3. Effect of Environmental Variables
3.4. Labellum Characterization
3.5. Diversity Distribution
3.6. Diversity Clustering
4. Discussion
4.1. Potential Distribution of Vanilla planifolia Andrews
4.1.1. Potential Distribution Model
4.1.2. Environmental Variables That Define the Potential Distribution
4.2. Labellum Characterization
4.2.1. Labellum Morphotypes
4.2.2. Geographic Distribution of Morphotypes
4.3. Final Considerations on the Labellum Variation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | M I | M II | M III | M IV | M V | Variable | M I | M II | M III | M IV | M V |
|---|---|---|---|---|---|---|---|---|---|---|---|
| A1 | 2.68 | 2.46 | 2.31 | 2.66 | 2.48 | D | 7.97 | 8.49 | 8.2 | 8.11 | 7.63 |
| A2 | 17.12 | 17.17 | 16.55 | 17.03 | 16.04 | E1 | 7.25 | 6.55 | 8.32 | 7.68 | 7.18 |
| A3 | 17.16 | 17.23 | 16.72 | 17.09 | 16.18 | E2 | 7.36 | 6.63 | 6.07 | 6.3 | 6.47 |
| A4 | 17.41 | 17.41 | 16.62 | 17.29 | 16.23 | E3 | 6.62 | 6.94 | 4.55 | 5.71 | 5.58 |
| A5 | 17.64 | 17.6 | 17.22 | 17.57 | 16.7 | E4 | 6.33 | 6.81 | 5.75 | 6.78 | 5.94 |
| A | 17.11 | 17.16 | 16.59 | 17.04 | 16.07 | E5 | 5.26 | 5.31 | 4.85 | 4.88 | 4.78 |
| b1 | 2.46 | 2.3 | 2.22 | 2.38 | 2.38 | E6 | 6.64 | 6.34 | 6.08 | 6.05 | 6.02 |
| b2 | 9.64 | 9.77 | 8.94 | 9.54 | 8.94 | E7 | 6.12 | 6.54 | 4.96 | 5.66 | 5.42 |
| b3 | 7.11 | 7.01 | 6.09 | 6.65 | 6.67 | E8 | 5.18 | 5.34 | 4.45 | 4.78 | 4.56 |
| b4 | 9.82 | 9.79 | 9.28 | 9.58 | 9.2 | E | 4.55 | 4.74 | 4.25 | 4.34 | 4.13 |
| b5 | 10.99 | 11.06 | 10.01 | 10.81 | 10.22 | F1 | 3.57 | 3.2 | 3.42 | 3.05 | 3.23 |
| b6 | 9 | 9.97 | 8.65 | 9.05 | 8.43 | F2 | 5.38 | 4.8 | 4.24 | 4.5 | 4.63 |
| b7 | 8.89 | 10.04 | 8.44 | 9.01 | 8.28 | F3 | 5.1 | 4.91 | 3.88 | 4.34 | 4.43 |
| b8 | 11.24 | 11.08 | 10.49 | 10.86 | 10.57 | F4 | 2.98 | 3.46 | 2.38 | 2.74 | 2.64 |
| B | 8.53 | 8.57 | 8.25 | 8.54 | 8 | F5 | 6.01 | 5.49 | 4.94 | 5.12 | 5.23 |
| C1 | 9.88 | 10 | 9.05 | 9.58 | 9.05 | F6 | 7.92 | 7.16 | 6.71 | 6.82 | 6.97 |
| C2 | 12.41 | 12.07 | 10.34 | 11.42 | 11.26 | F7 | 7.08 | 7.42 | 5.08 | 6.2 | 6.03 |
| C3 | 11.94 | 12.07 | 9.64 | 10.98 | 10.76 | F8 | 5.77 | 5.59 | 4.63 | 4.99 | 5.04 |
| C4 | 9.97 | 9.94 | 9.21 | 9.6 | 9.15 | F | 2.69 | 2.65 | 2.53 | 2.47 | 2.41 |
| C5 | 14.77 | 14.81 | 12.8 | 13.97 | 13.51 | G1 | 3.88 | 3.61 | 2.69 | 3.42 | 3.19 |
| C6 | 11.43 | 11.1 | 10.58 | 10.88 | 10.65 | G2 | 4.71 | 4.2 | 4.49 | 4.07 | 4.3 |
| C7 | 10.92 | 11.11 | 9.93 | 10.65 | 10.2 | G3 | 3.79 | 3.75 | 2.96 | 3.36 | 3.18 |
| C8 | 14.93 | 14.75 | 13.11 | 14.04 | 13.68 | G4 | 3.64 | 3.16 | 3.23 | 3.13 | 3.11 |
| C | 8.55 | 8.59 | 8.26 | 8.47 | 8.01 | G5 | 3.34 | 3.14 | 2.67 | 2.98 | 2.88 |
| D1 | 7.69 | 8.83 | 7.17 | 7.86 | 7.22 | G | 2.54 | 2.38 | 2.22 | 2.34 | 2.09 |
| D2 | 12.77 | 11.23 | 12.55 | 12.44 | 12.07 | aA | 24.9 | 23.16 | 22.42 | 24.5 | 25.21 |
| D3 | 11.12 | 11.53 | 9.12 | 11.04 | 10.13 | aB | 32.09 | 29.98 | 29.98 | 31.16 | 33.11 |
| D4 | 8.47 | 8.41 | 9.26 | 8.46 | 8.14 | aD | 58.3 | 54.31 | 46.36 | 52.44 | 54.58 |
| D5 | 11.02 | 11.57 | 10.11 | 10.82 | 10.29 | aE | 88.17 | 84.65 | 77.28 | 83.78 | 86.17 |
| D6 | 9.15 | 9.56 | 8.68 | 8.96 | 8.52 | aDE22 | 127.81 | 141.76 | 112.12 | 120.28 | 122.5 |
| D7 | 9.16 | 9.51 | 9.06 | 9.01 | 8.6 | aDE55 | 140.41 | 139.1 | 130.68 | 129.01 | 134.54 |
| D8 | 11.2 | 11.36 | 10.79 | 10.94 | 10.54 | aG | 85.75 | 81.11 | 105.52 | 79.48 | 90.47 |
References
- Schlüter, P.M.; Soto-Arenas, M.A.; Harris, S.A. Genetic variation in Vanilla planifolia (Orchidaceae). Econ. Bot. 2007, 61, 328. [Google Scholar] [CrossRef]
- Hu, Y.; Resende, M.F.R.; Bombarely, A.; Brym, M.; Bassil, E.; Chambers, A.H. Genomics-based diversity analysis of Vanilla species using a Vanilla planifolia draft genome and Genotyping-By-Sequencing. Sci. Rep. 2019, 9, 3416. [Google Scholar] [CrossRef]
- Soto Arenas, M.A.; Dressler, R.L.; Cameron, K.; Cribb, P.; Hágsater, E.; Salazar, G.; Solano, R. A revision of the Mexican and Central American species of Vanilla plumier ex miller with a characterization of their its region of the nuclear ribosomal DNA. Lankesteriana 2009, 9, 285–354. [Google Scholar] [CrossRef]
- Herrera-Cabrera, B.E.; Salgado Garciglia, R.; Manuel, V.; Higuera, O.; Jair Barrales-Cureño, H.; Delgado Alvarado, A.; Montiel-Montoya, J.; Diaz-Bautista, M.; Albino, R.A.; Reyes, C. Producción y caracterización de vainilla (Vanilla planifolia) en función de la concentración de vainillina. Rev. Iberoam. Cienc. 2022, 9, 46–62. [Google Scholar]
- Soto-Arenas, M.A.S.; Cribb, P. A new infrageneric classification and synopsis of the genus Vanilla Plum. ex mill. (Orchidaceae: Vanillinae). Lankesteriana 2009, 9, 355–398. [Google Scholar] [CrossRef]
- Herrera-Cabrera, B.E.; Salazar-Rojas, V.M.; Delgado-Alvarado, A.; Contreras, J.; Contreras, C.; Cervantes-Vargas, J. Use and conservation of Vanilla planifolia J. in the Totonacapan Region, México. Eur. J. Environ. Sci. 2012, 2, 43–50. [Google Scholar] [CrossRef]
- Bello-Bello, J.J.; García-García, G.G.; Iglesias-Andreu, L. Conservación de vainilla (Vanilla planifolia Jacks.) bajo condiciones de lento crecimiento in vitro. Rev. Fitotec. Mex. 2015, 38, 165–171. [Google Scholar] [CrossRef]
- Householder, E.; Janovec, J.; Mozambite, A.B.; Maceda, J.H.; Wells, J.; Valega, R.; Maruenda, H.; Christenson, E. Diversity, natural history, and conservation of Vanilla (Orchidaceae) in amazonian wetlands of Madre de Dios, Peru. J. Bot. Res. Inst. Tex. 2010, 4, 227–243. [Google Scholar]
- Ellestad, P.; Pérez-Farrera, M.A.; Buerki, S. Genomic insights into cultivated mexican Vanilla planifolia reveal high levels of heterozygosity stemming from hybridization. Plants 2022, 11, 2090. [Google Scholar] [CrossRef]
- Faleiro, F.V.; Machado, R.B.; Loyola, R.D. Defining spatial conservation priorities in the face of land-use and climate change. Biol. Conserv. 2013, 158, 248–257. [Google Scholar] [CrossRef]
- Peterson, A.T.; Papes, M.; Eaton, M. Transferability and model evaluation in ecological niche modeling: A comparison of GARP and MaxEnt. Ecography 2007, 30, 550–560. [Google Scholar] [CrossRef]
- Kozak, K.H.; Graham, C.H.; Wiens, J.J. Integrating GIS-based environmental data into evolutionary biology. Trends Ecol. Evol. 2008, 23, 141–148. [Google Scholar] [CrossRef]
- Chen, G.; Kéry, M.; Zhang, J.; Ma, K. Factors affecting detection probability in plant distribution studies. J. Ecol. 2009, 97, 1383–1389. [Google Scholar] [CrossRef]
- Bertolini, V.; Damon, A.; Valle Mora, J.; Natanael Rojas Velázquez, A. Distribution and ecological patterns of orchids in Monte Pel-legrino Reserve, Palermo (Sicily, Italy). Biodivers. J. 2012, 3, 375–384. [Google Scholar]
- Kalkvik, H.M.; Stout, I.J.; Doonan, T.J.; Parkinson, C.L. Investigating niche and lineage diversification in widely distributed taxa: Phylogeography and ecological niche modeling of the Peromyscus maniculatus species group. Ecography 2012, 35, 54–64. [Google Scholar] [CrossRef]
- Kolanowska, M.; Szlachetko, D.L. Niche conservatism of Eulophia alta, a trans-Atlantic orchid species. Acta Soc. Bot. Pol. 2014, 83, 51–57. [Google Scholar] [CrossRef]
- Kumar, S.; Stohlgren, T.J. MaxEnt modeling for predicting suitable habitat for threatened and endangered tree Canacomyrica monticola in New Caledonia. J. Ecol. Nat. Environ. 2009, 1, 94–98. [Google Scholar]
- Murray-Smith, C.; Brummitt, N.A.; Oliveira-Filho, A.T.; Bachman, S.; Moat, J.; Lughadha, E.M.N.; Lucas, E.J. Plant diversity hotspots in the Atlantic coastal forests of Brazil. Conserv. Biol. 2009, 23, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Lehtomäki, J.; Moilanen, A. Methods and workflow for spatial conservation prioritization using Zonation. Environ. Model. Softw. 2013, 47, 128–137. [Google Scholar] [CrossRef]
- Yang, X.Q.; Kushwaha, S.P.S.; Saran, S.; Xu, J.; Roy, P.S. MaxEnt modeling for predicting the potential distribution of medicinal plant, Justicia adhatoda L. in Lesser Himalayan foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Fandohan, B.; Assogbadjo, A.E.; Glèlè Kakaï, R.L.; Sinsin, B. Effectiveness of a protected areas network in the conservation of Tamarindus indica (Leguminosea–Caesalpinioideae) in Benin. Afr. J. Ecol. 2011, 49, 40–50. [Google Scholar] [CrossRef]
- Phillips, S.B.; Aneja, V.P.; Kang, D.; Arya, S.P. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with MaxEnt: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Wan, J.; Wang, C.; Han, S.; Yu, J. Planning the priority protected areas of endangered orchid species in northeastern China. Biodivers. Conserv. 2014, 23, 1395–1409. [Google Scholar] [CrossRef]
- Flores Jiménez, Á.; Reyes López, D.; García, D.J.; Romero Arenas, O.; Antonio, J.; Tapia, R.; Lara, M.H.; Silva, A.P. Diversidad de Vanilla spp. (Orchidaceae) y sus perfiles bioclimáticos en México. Rev. Biol. Trop. 2017, 65, 975–987. [Google Scholar] [CrossRef]
- Santillán-Fernández, A.; Cabrera, M.T.; Martínez Sánchez, A.; Ángel, L.M.; Vásquez Bautista, N.; Mejía, S.L. Potencial productivo de Vanilla planifolia Jacks en el Totonacapan, México, mediante técnicas geográficas. Rev. Mex. Cienc. Agrícolas 2019, 10, 789–802. [Google Scholar] [CrossRef]
- Hernández-Ruíz, J.; Herrera-Cabrera, B.E.; Delgado-Alvarado, A.; Salazar-Rojas, V.M.; Bustamante-Gonzalez, Á.; Campos-Contreras, J.E.; Ramírez-Juarez, J. Distribución potencial y características geográficas de poblaciones silvestres de Vanilla planifolia(Orchidaceae) en Oaxaca, México. Rev. Biol. Trop. 2016, 64, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Reyes Hernández, H.; Trinidad García, K.L.; Herrera Cabrera, B.E. Caracterización del ambiente de los vainillales y área potencial para su cultivo en la Huasteca Potosína. Biotecnia 2018, 20, 49–57. [Google Scholar] [CrossRef]
- Trinidad García, K.L.; Reyes Hernández, H.; Martínez Salazar, R.I.; Galarza Rincón, E.; Trinidad García, K.L.; Reyes Hernández, H.; Martínez Salazar, R.I.; Galarza Rincón, E. Distribución de Vanilla planifolia Jacks. ex Andrews y acciones para su conservación en la Huasteca Potosina. Rev. Mex. Cienc. For. 2019, 10, 108–134. [Google Scholar] [CrossRef]
- Armenta-Montero, S.; Menchaca-García, R.; Pérez-Silva, A.; Velázquez-Rosas, N. Changes in the Potential Distribution of Vanilla planifolia Andrews under Different Climate Change Projections in Mexico. Sustainability 2022, 14, 2881. [Google Scholar] [CrossRef]
- Lubinsky, P.; Bory, S.; Hernández Hernández, J.; Kim, S.C.; Gómez-Pompa, A. Origins and dispersal of cultivated vanilla (Vanilla planifolia Jacks. [Orchidaceae]). Econ. Bot. 2008, 62, 127–138. [Google Scholar] [CrossRef]
- Cortéz-Marin, A.L.; Aceves-Navarro, L.A.; Arteaga-Ramírez, R.; Vázquez-Peña, M.A. Zonificación agroecológica para aguacate en la zona central de Venezuela. Terra Latinoam. 2005, 23, 159–166. [Google Scholar]
- Martínez, M.Á.; Evangelista, V.; Basurto, F.; Mendoza, M.; Cruz-Rivas, A. Flora útil de los cafetales en la Sierra Norte de Puebla, México. Rev. Mex. Biodivers. 2007, 78, 15–40. [Google Scholar] [CrossRef]
- Pedroso, H.L.; Rocha-Filho, L.C.; Lomônaco, C. Variación fenotípica de plantas del Cerrado (Sabana brasileña) frente a la heterogeneidad ambiental. Ecosistemas 2010, 19, 24–36. [Google Scholar]
- Vargas-Amado, G.; Castro-Castro, A.; Harker, M.; Villaseñor, J.L.; Ortiz, E.; Rodríguez, A. Distribución geográfica y riqueza del género Cosmos (Asteraceae: Coreopsideae). Rev. Mex. Biodivers. 2013, 84, 536–555. [Google Scholar] [CrossRef]
- De Oliveira, R.T.; da Silva Oliveira, J.P.; Macedo, A.F. Vanilla beyond Vanilla planifolia and Vanilla × tahitensis: Taxonomy and Historical Notes, Reproductive Biology, and Metabolites. Plants 2022, 11, 3311. [Google Scholar] [CrossRef]
- Villanueva-Viramontes, S.; Hernández-Apolinar, M.; Carnevali Fernández-Concha, G.; Dorantes-Euán, A.; Dzib, G.R.; Martínez-Castillo, J. Vanilla planifolia silvestre y sus parientes en la Península de Yucatán, México: Análisis sistemáticos con ISSR e ITS. Bot. Sci. 2017, 95, 169–187. [Google Scholar] [CrossRef]
- Flanagan, N.S.; Navia-Samboni, A.; González-Pérez, E.N.; Mendieta-Matallana, H. Distribution and conservation of vanilla crop wild relatives: The value of local community engagement for biodiversity research. Neotrop. Biol. Conserv. 2022, 17, 205–227. [Google Scholar] [CrossRef]
- Hernandez-Ruiz, J.; Delgado-Alvarado, A.; Salazar-Rojas, V.M.; Herrera-Cabrera, B.E. Morphological variation of the labellum of Vanilla planifolia Andrews (Orchidaceae) in Oaxaca, Mexico. Rev. La Fac. Cienc. Agrar. UNCuyo 2020, 52, 160–175. [Google Scholar]
- Hernández-Ruiz, J.; Herrera-Cabrera, B.E.; Delgado-Alvarado, A. Variación morfológica del labelo de Vanilla pompona (Orchidaceae) en Oaxaca, México. Rev. Mex. Biodivers. 2019, 90, 16. [Google Scholar] [CrossRef]
- Lima-Morales, M.; Herrera-Cabrera, B.E.; Delgado-Alvarado, A. Intraspecific variation of Vanilla planifolia (Orchidaceae) in the Huasteca region, San Luis Potosí, Mexico: Morphometry of floral labellum. Plant Syst. Evol. 2021, 307, 40. [Google Scholar] [CrossRef]
- Flanagan, N.S.; Mosquera-Espinosa, A.T. An integrated strategy for the conservation and sustainable use of native Vanilla species in Colombia. Lankesteriana 2016, 16, 201–218. [Google Scholar] [CrossRef]
- Herrera, J. The Variability of Organs Differentially Involved in Pollination, and Correlations of Traits in Genisteae (Leguminosae: Papilionoideae). Ann. Bot. 2001, 88, 1027–1037. [Google Scholar] [CrossRef]
- Brock, M.T.; Weinig, C. Plasticity and environment-specific covariances: An investigation of floral-vegetative and within flower correlations. Evolution 2007, 61, 2913–2924. [Google Scholar] [CrossRef]
- Herrera, J. Flower Size Variation in Rosmarinus officinalis: Individuals, Populations and Habitats. Ann. Bot. 2005, 95, 431–437. [Google Scholar] [CrossRef]
- Chiron, G.R.; Guignard, G.; Barale, G. Contribution of Morphometry to the Taxonomy of Baptistonia Barb. Rodr. (Orchidaceae). Candollea 2010, 65, 45–62. [Google Scholar] [CrossRef]
- Margońska, H.B.; Kozieradzka-Kiszkurno, M.; Brzezicka, E.; Haliński, Ł.P.; Davies, K.L.; Lipińska, M.M. Crepidium sect. Crepidium (Orchidaceae, Malaxidinae)—Chemical and Morphological Study of Flower Structures in the Context of Pollination Processes. Plants 2021, 10, 2373. [Google Scholar] [CrossRef]
- Shipunov, A.B.; Bateman, R.M. Geometric morphometrics as a tool for understanding Dactylorhiza (Orchidaceae) diversity in European Russia. Biol. J. Linn. Soc. 2005, 85, 1–12. [Google Scholar] [CrossRef]
- Sobel, J.M.; Streisfeld, M.A. Flower color as a model system for studies of plant evo-devo. Front. Plant Sci. 2013, 4, 321. [Google Scholar] [CrossRef]
- Salazar-Rojas, V.M.; Herrera-Cabrera, B.E.; Soto-Arenas, M.Á.; Castillo-González, F. Morphological variation in Laelia anceps subsp. dawsonii f. chilapensis Soto-Arenas Orchidaceae in traditional home gardens of Chilapa, Guerrero, Mexico. Genet. Resour. Crop Evol. 2010, 57, 543–552. [Google Scholar] [CrossRef]
- Priyanka, V.; Kumar, R.; Dhaliwal, I.; Kaushik, P. Germplasm Conservation: Instrumental in Agricultural Biodiversity-A Review. Sustainability 2021, 13, 6743. [Google Scholar] [CrossRef]
- Villavicencio Nieto, M.Á.; Pérez Escandón, E.B. Vegetación e inventario de la flora útil de la Huasteca y la zona Otomí-Tepehua de Hidalgo. Cienc. Univ. 2010, 1, 23–33. [Google Scholar]
- Leoncio, J.; García, M. Lucha campesina en la Huasteca hidalguense. Un estudio regional. Estud. Agrar. 2013, 19, 17–90. [Google Scholar]
- Ceja-Romero, J.; Mendoza-Ruiz, A.; López-Ferrari, A.R.; Espejo-Serna, A.; Pérez-García, B.; García-Cruz, J. Las epífitas vasculares del Estado de Hidalgo, México: Diversidad y distribución. Acta Botánica Mex. 2010, 93, 1–39. [Google Scholar] [CrossRef]
- Cruz-Cárdenas, G.; López-Mata, L.; Villaseñor, J.L.; Ortiz, E. Potential species distribution modeling and the use of principal component analysis as predictor variables. Rev. Mex. Biodivers. 2014, 85, 189–199. [Google Scholar] [CrossRef]
- Drake, J.M.; Beier, J.C. Ecological niche and potential distribution of Anopheles arabiensis in Africa in 2050. Malar. J. 2014, 13, 213. [Google Scholar] [CrossRef] [PubMed]
- CONABIO—Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. Available online: http://www.conabio.gob.mx/informacion/gis/ (accessed on 18 May 2022).
- Kumar, S.; Graham, J.; West, A.M.; Evangelista, P.H. Using district-level occurrences in MaxEnt for predicting the invasion potential of an exotic insect pest in India. Comput. Electron. Agric. 2014, 103, 55–62. [Google Scholar] [CrossRef]
- Fitzgibbon, A.; Pisut, D.; Fleisher, D. Evaluation of Maximum Entropy (MaxEnt) Machine Learning Model to Assess Relationships between Climate and Corn Suitability. Land 2022, 11, 1382. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in MaxEnt: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef]
- Gunawan, G.; Sulistijorini, S.; Chikmawati, T.; Sobir, S. Predicting suitable areas for Baccaurea angulata in Kalimantan, Indonesia using MaxEnt modelling. Biodiversitas J. Biol. Divers. 2021, 22, 2646–2653. [Google Scholar] [CrossRef]
- Padalia, H.; Srivastava, V.; Kushwaha, S.P.S. Modeling potential invasion range of alien invasive species, Hyptis suaveolens (L.) Poit. in India: Comparison of MaxEnt and GARP. Ecol. Inform. 2014, 22, 36–43. [Google Scholar] [CrossRef]
- Radović, S.; Urošević, A.; Hočevar, K.; Vuleta, A.; Manitašević Jovanović, S.; Tucić, B. Geometric morphometrics of functionally distinct floral organs in Iris pumila: Analyzing patterns of symmetric and asymmetric shape variations. Arch. Biol. Sci. 2017, 69, 223–231. [Google Scholar] [CrossRef]
- Caiza Guamba, J.C.; Corredor, D.; Galárraga, C.; Herdoiza, J.P.; Santillán, M.; Segovia-Salcedo, M.C. Geometry morphometrics of plant structures as a phenotypic tool to differentiate Polylepis incana Kunth. and Polylepis racemosa Ruiz & Pav. reforested jointly in Ecuador. Neotrop. Biodivers. 2021, 7, 121–134. [Google Scholar] [CrossRef]
- Ibacache, M.V.T.; Soto, G.M.; Galdames, I.S. Morfometría geométrica y el estudio de las formas biológicas: De la morfología descriptiva a la morfología cuantitativa. Int. J. Morphol. 2010, 28, 977–990. [Google Scholar] [CrossRef]
- McPherson, J.M.; Jetz, M. Effects of species’ ecology on the accuracy of distribution models. Ecography 2007, 30, 135–151. [Google Scholar] [CrossRef]
- Evangelista, P.H.; Kumar, S.; Stohlgren, T.J.; Jarnevich, C.S.; Crall, A.W.; Norman, J.B.; Barnett, D.T. Modelling invasion for a habitat generalist and a specialist plant species. Divers. Distrib. 2008, 14, 808–817. [Google Scholar] [CrossRef]
- Jaryan, V.; Datta, A.; Uniyal, S.K.; Kumar, A.; Gupta, R.C.; Singh, R.D. Modelling potential distribution of Sapium sebiferum—An invasive tree species in western Himalaya. Curr. Sci. 2013, 105, 1282–1288. [Google Scholar]
- Shcheglovitova, M.; Anderson, R.P. Estimating optimal complexity for ecological niche models: A jackknife approach for species with small sample sizes. Ecol. Modell. 2013, 269, 9–17. [Google Scholar] [CrossRef]
- Barrera-Rodríguez, A.; Herrera-Cabrera, B.E.; Jaramillo-Villanueva, J.L.; Escobedo-Garrido, S.; Bustamante-González, A. Caracterización de los sistemas de producción de vainilla (Vanilla planifolia A.) bajo naranjo y en malla sombra en el Totonacapan. Trop. Subtrop. Agroecosyst. 2009, 10, 199–212. [Google Scholar]
- García-González, A.; Damon, A.; Esparza, O.; Ligia, G.; Valle-Mora, J. Population structure of Oncidium poikilostalix (Orchidaceae), in coffee plantations. Lankesteriana Int. J. Orchid. 2011, 11, 23–32. [Google Scholar]
- Moorhead, L.C.; Philpott, S.M.; Bichier, P. Epiphyte biodiversity in the coffee agricultural matrix: Canopy stratification and distance from forest fragments. Conserv. Biol. 2010, 24, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, C. Diversity and abundance of orchid bees (Hymenoptera: Apidae, Euglossini) in a tropical rainforest succession. Neotrop. Entomol. 2009, 38, 66–73. [Google Scholar] [CrossRef]
- Lozano, F.D.; Schwartz, M.W. Patterns of rarity and taxonomic group size in plants. Biol. Conserv. 2005, 126, 146–154. [Google Scholar] [CrossRef]
- Tsiftsis, S.; Tsiripidis, I.; Trigas, P. Identifying important areas for orchid conservation in Crete. Eur. J. Environ. Sci. 2012, 1, 28–37. [Google Scholar] [CrossRef]
- Condit, R. Spatial patterns in the distribution of tropical tree species. Science 2000, 288, 1414–1418. [Google Scholar] [CrossRef]
- Casazza, G.; Zappa, E.; Mariotti, M.G.; Médail, F.; Minuto, L. Ecological and historical factors affecting distribution pattern and richness of endemic plant species: The case of the Maritime and Ligurian Alps hotspot. Divers. Distrib. 2008, 14, 47–58. [Google Scholar] [CrossRef]
- Kijowska-Oberc, J.; Staszak, A.M.; Kamiński, J.; Ratajczak, E. Adaptation of Forest Trees to Rapidly Changing Climate. Forests 2020, 11, 123. [Google Scholar] [CrossRef]
- Oguz, M.C.; Aycan, M.; Oguz, E.; Poyraz, I.; Yildiz, M. Drought Stress Tolerance in Plants: Interplay of Molecular, Biochemical and Physiological Responses in Important Development Stages. Physiologia 2022, 2, 180–197. [Google Scholar] [CrossRef]
- SEMARNAT NORMA Oficial Mexicana NOM-059-SEMARNAT-2010, Protección Ambiental-Especies Nativas de México de Flora y fauna Silvestres-Categorías de Riesgo y Especificaciones para su Inclusión, Exclusión o Cambio-Lista de Especies en Riesgo. Available online: https://www.dof.gob.mx/normasOficiales/4254/semarnat/semarnat.htm (accessed on 23 January 2023).
- Soto-Arenas, M.A.; Solano-Gómez, A.R. Ficha técnica de Vanilla planifolia. In Información Actualizada Sobre Las Especies de Orquídeas del PROY-NOM-059-ECOL2000. Bases de Datos SNIB-CONABIO. Proyecto No. W029, 1st ed.; Soto-Arenas, M.A., Ed.; Instituto Chinoin A.C., Herbario de la Asociación Mexicana de Orquideología A.C.: México City, Mexico, 2007; pp. 1–9. [Google Scholar]
- Paiaro, V.; Oliva, G.E.; Cocucci, A.A.; Sérsic, A.N. Geographic patterns and environmental drivers of flower and leaf variation in an endemic legume of Southern Patagonia. Plant Ecol. Divers. 2012, 5, 13–25. [Google Scholar] [CrossRef]
- Pigliucci, M. Evolution of phenotypic plasticity: Where are we going now? Trends Ecol. Evol. 2005, 20, 481–486. [Google Scholar] [CrossRef]
- Hodgins, K.A.; Barrett, S.C.H. Geographic variation in floral morphology and style-morph ratios in a sexually polymorphic daffodil. Am. J. Bot. 2008, 95, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Pélabon, C.; Osler, N.C.; Diekmann, M.; Graae, B.J. Decoupled phenotypic variation between floral and vegetative traits: Distinguishing between developmental and environmental correlations. Ann. Bot. 2013, 111, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Givnish, T.J. Ecological constraints on the evolution of plasticity in plants. Evol. Ecol. 2002, 16, 213–242. [Google Scholar] [CrossRef]
- Blinova, I.V. Intra- and interspecific morphological variation of some European terrestrial orchids along a latitudinal gradient. Russ. J. Ecol. 2012, 43, 111–116. [Google Scholar] [CrossRef]
- Ramírez, N.; Nassar, J.M.; Valera, L.; Garay, V.; Briceño, H.; Quijada, M.; Moret, Y.A.; Montilla, J. Variación morfométrica floral en Pachira quinata (Jacq.) W.Alverson (Bombacaceae). Acta Botánica Venez. 2010, 33, 83–102. [Google Scholar]
- Mccormick, M.K.; Jacquemyn, H. Research review What constrains the distribution of orchid populations? New Phytol. 2014, 202, 392–400. [Google Scholar] [CrossRef]
- Damon, A.; Hernández-Ramírez, F.; Riggi, L.; Verspoor, R.; Bertolini, V.; Lennartz-Walker, M.; Wiles, A.; Burns, A. Pollination of euglossinophylic epiphytic orchids in agroecosystems and forest fragments in southeast Mexico. Eur. J. Environ. Sci. 2012, 2, 5–14. [Google Scholar] [CrossRef]
- Benitez-Vieyra, S.; Medina, A.M.; Cocucci, A.A. Variable selection patterns on the labellum shape of Geoblasta pennicillata, a sexually deceptive orchid. J. Evol. Biol. 2009, 22, 2354–2362. [Google Scholar] [CrossRef]
- Gaskett, A.C. Floral shape mimicry and variation in sexually deceptive orchids with a shared pollinator. Biol. J. Linn. Soc. 2012, 106, 469–481. [Google Scholar] [CrossRef]
- Bory, S.; Grisoni, M.; Duval, M.F.; Besse, P. Biodiversity and preservation of vanilla: Present state of knowledge. Genet. Resour. Crop Evol. 2008, 55, 551–571. [Google Scholar] [CrossRef]
- Bateman, R.M.; Rudall, P.J. Evolutionary and Morphometric Implications of Morphological Variation Among Flowers within an Inflorescence: A Case-Study Using European Orchids. Ann. Bot. 2006, 98, 975–993. [Google Scholar] [CrossRef]
- Savriama, Y.; Gómez, J.M.; Perfectti, F.; Klingenberg, C.P. Geometric morphometrics of corolla shape: Dissecting components of symmetric and asymmetric variation in Erysimum mediohispanicum (Brassicaceae). New Phytol. 2012, 196, 945–954. [Google Scholar] [CrossRef] [PubMed]
- Solís-Montero, L.; Vallejo-Marín, M. Does the morphological fit between flowers and pollinators affect pollen deposition? An experimental test in a buzz-pollinated species with anther dimorphism. Ecol. Evol. 2017, 7, 2706–2715. [Google Scholar] [CrossRef]
- Ordano, M.; Fornoni, J.; Boege, K.; Domínguez, C.A. The adaptive value of phenotypic floral integration. New Phytol. 2008, 179, 1183–1192. [Google Scholar] [CrossRef]
- Gong, Y.B.; Huang, S.Q. Floral symmetry: Pollinator-mediated stabilizing selection on flower size in bilateral species. Proc. R. Soc. B Biol. Sci. 2009, 276, 4013–4020. [Google Scholar] [CrossRef] [PubMed]
- Borba, E.L.; Shepherd, G.J.; Van Den Berg, C.; Semir, J. Floral and Vegetative Morphometrics of Five Pleurothallis (Orchidaceae) Species: Correlation with Taxonomy, Phylogeny, Genetic Variability and Pollination Systems. Ann. Bot. 2002, 90, 230. [Google Scholar] [CrossRef]
- Lozano-Rodríguez, M.A.; Luna-Rodríguez, M.; Pench-Canché, J.M.; Menchaca-García, R.A.; Cerdán-Cabrera, C.R. Visit frequency of Euglossine bees (Hymenoptera: Apidae) to mature fruits of Vanilla planifolia (Orchidaceae). Acta Bot. Mex. 2022, 129, e2001. [Google Scholar] [CrossRef]
- Pansarin, E. Vanilla flowers: Much more than food-deception. Bot. J. Linn. 2021, 20, 1–17. [Google Scholar] [CrossRef]
- Andriamihaja, C.F.; Botomanga, A.; Misandeau, C.; Ramarosandratana, A.V.; Grisoni, M.; Da Silva, D.; Pailler, T.; Jeannoda, V.H.; Besse, P. Integrative taxonomy and phylogeny of leafless Vanilla orchids from the South-West Indian Ocean region reveal two new Malagasy species. J. Syst. Evol. 2022, 61, 80–90. [Google Scholar] [CrossRef]
- Ellestad, P.; Perez-Farrera, M.A.; Forest, F.; Buerki, S. Uncovering haplotype diversity in cultivated Mexican vanilla species. Am. J. Bot. 2022, 109, 1120–1138. [Google Scholar] [CrossRef] [PubMed]
- Grisoni, M.; Nany, F. The beautiful hills: Half a century of vanilla (Vanilla planifolia Jacks. ex Andrews) breeding in Madagascar. Genet. Resour. Crop Evol. 2021, 68, 1691–1708. [Google Scholar] [CrossRef]











| Code | Environmental Variables | Units |
|---|---|---|
| Bio1 | Annual mean temperature | °C |
| Bio2 | Mean diurnal range | °C |
| Bio3 | Isothermality | Dimensionless |
| Bio4 | Temperature seasonality | CV |
| Bio5 | Max temperature of the warmest month | °C |
| Bio6 | Min temperature of the coldest month | °C |
| Bio7 | Temperature annual range | °C |
| Bio8 | Mean temperature of the wettest quarter | °C |
| Bio9 | Mean temperature of the driest quarter | °C |
| Bio10 | Mean temperature of the warmest quarter | °C |
| Bio11 | Mean temperature of the coldest quarter | °C |
| Bio12 | Annual precipitation | mm |
| Bio13 | Precipitation of the wettest month | mm |
| Bio14 | Precipitation of the driest month | mm |
| Bio15 | Precipitation seasonality | CV |
| Bio16 | Precipitation of the wettest quarter | mm |
| Bio17 | Precipitation of the driest quarter | mm |
| Bio18 | Precipitation of the warmest quarter | mm |
| Bio19 | Precipitation of the coldest quarter | mm |
| Cover | Vegetation cover | 16 types |
| Alt | Altitude | m |
| Municipality | Locality | Accessions | Number of Flowers (Repetition) |
|---|---|---|---|
| Atlapexco | Itzocal | S1 | 20 |
| S2 | 13 | ||
| Huizotlaco | S3 | 1 | |
| S4 | 7 | ||
| San Isidro | S5 | 27 | |
| S6 | 26 | ||
| Huejutla | Contepec | S7 | 19 |
| S8 | 18 | ||
| Tezahual | S9 | 20 | |
| Xocotitla | S10 | 12 | |
| Poxtla | S11 | 5 | |
| S12 | 9 | ||
| S13 | 3 | ||
| Pahuatlán | S14 | 30 | |
| Ichcatepec | S15 | 20 | |
| Jaltocán | Tlanepantla | S16 | 20 |
| Mirador | S17 | 16 | |
| Huejutla | Coacuilco | S18 | 17 |
| S19 | 10 | ||
| S20 | 14 | ||
| S21 | 11 | ||
| S22 | 10 |
| Municipality | Locality | Accession | Altitude | Weather * | Vegetation * |
|---|---|---|---|---|---|
| Atlapexco | Itzocal | S1 | 370 | Am(f) Warm and wet | Agricultural use |
| S2 | 382 | ||||
| Huizotlaco | S3 | 285 | |||
| S4 | 273 | ||||
| San Isidro | S5 | 394 | |||
| S6 | 350 | ||||
| Huejutla | Contepec | S7 | 406 | (A)C(m)(f) Semiwarm-temperate humid | Tropical or subtropical evergreen broadleaf forest |
| S8 | 352 | ||||
| Tezahual | S9 | 414 | |||
| Xocotitla | S10 | 391 | |||
| Poxtla | S11 | 312 | Agricultural use | ||
| S12 | 367 | ||||
| S13 | 331 | ||||
| Pahuatlán | S14 | 381 | |||
| Ichcatepec | S15 | 545 | |||
| Jaltocán | Tlanepantla | S16 | 482 | ||
| Mirador | S17 | 316 | Am(f) Warm and wet | ||
| Huejutla | Coacuilco | S18 | 420 | A(f) Warm humid coldest month less than 18 °C | Tropical or subtropical evergreen broadleaf forest |
| S19 | 400 | ||||
| S20 | 473 | (A)C(fm) Semi-warm humid of group C | |||
| S21 | 398 | ||||
| S22 | 423 |
| Variable | Contribution (%) |
|---|---|
| Precipitation of driest month (Bio14) | 43 |
| Vegetal cover (Cover) | 14.9 |
| Precipitation of the driest quarter (Bio17) | 7.2 |
| Temperature seasonality (Bio4) | 7 |
| Precipitation seasonality (Bio15) | 6.5 |
| Mean temperature of the wettest quarter (Bio8) | 5.8 |
| Mean temperature of the driest quarter (Bio9) | 5.3 |
| Annual mean temperature (Bio1) | 4.9 |
| Mean diurnal range (Bio2) | 2.7 |
| Altitude (Alt) | 1.3 |
| Precipitation of the wettest quarter (Bio16) | 0.7 |
| Temperature annual range (Bio7) | 0.6 |
| Variable | Mean | Coefficient of Variation | Mean Square | Variable | Mean | Coefficient of Variation | Mean Square | ||
|---|---|---|---|---|---|---|---|---|---|
| Accessions | Error | Accessions | Error | ||||||
| A1 | 2.56 | 10.15 | 0.52 *** | 0.06 | D | 7.93 | 6.05 | 2.52 *** | 0.23 |
| A2 | 16.76 | 3.46 | 4.08 *** | 0.33 | E1 | 7.1 | 11.7 | 3.18 *** | 0.69 |
| A3 | 16.85 | 3.65 | 3.52 *** | 0.37 | E2 | 6.81 | 11.04 | 4.47 *** | 0.56 |
| A4 | 17 | 3.6 | 4.70 *** | 0.37 | E3 | 6.23 | 9.27 | 5.70 *** | 0.33 |
| A5 | 17.33 | 4.04 | 3.39 *** | 0.49 | E4 | 6.3 | 12.62 | 3.08 *** | 0.63 |
| A | 16.77 | 3.51 | 3.84 *** | 0.34 | E5 | 5.04 | 8.37 | 1.51 *** | 0.17 |
| B1 | 2.38 | 5.07 | 0.16 *** | 0.01 | E6 | 6.29 | 8.62 | 2.07 *** | 0.29 |
| B2 | 9.4 | 4.06 | 2.21 *** | 0.14 | E7 | 5.9 | 7.6 | 2.95 *** | 0.2 |
| B3 | 6.87 | 6.08 | 2.14 *** | 0.17 | E8 | 4.94 | 7.4 | 2.01 *** | 0.13 |
| B4 | 9.57 | 4.12 | 1.37 *** | 0.15 | E | 4.4 | 6.48 | 1.17 *** | 0.08 |
| B5 | 10.71 | 4.21 | 2.96 *** | 0.2 | F1 | 3.32 | 14.37 | 1.21 *** | 0.22 |
| B6 | 8.93 | 5.28 | 5.14 *** | 0.22 | F2 | 4.94 | 7.64 | 2.53 *** | 0.14 |
| B7 | 8.83 | 5.7 | 6.12 *** | 0.25 | F3 | 4.77 | 7.01 | 2.25 *** | 0.11 |
| B8 | 10.94 | 4.2 | 1.81 *** | 0.21 | F4 | 2.91 | 13.36 | 1.49 *** | 0.15 |
| B | 8.36 | 3.49 | 1.02 *** | 0.08 | F5 | 5.57 | 6.85 | 2.76 *** | 0.14 |
| C1 | 9.58 | 5.1 | 2.65 *** | 0.23 | F6 | 7.34 | 10.84 | 4.89 *** | 0.63 |
| C2 | 11.87 | 5.82 | 6.83 *** | 0.47 | F7 | 6.7 | 9.38 | 6.02 *** | 0.39 |
| C3 | 11.45 | 5.39 | 7.56 *** | 0.38 | F8 | 5.42 | 6.3 | 2.56 *** | 0.11 |
| C4 | 9.69 | 5.32 | 2.11 *** | 0.26 | F | 2.56 | 7.25 | 0.49 *** | 0.03 |
| C5 | 14.24 | 5.12 | 7.31 *** | 0.53 | G1 | 3.58 | 13.4 | 1.94 *** | 0.23 |
| C6 | 11.06 | 5.76 | 2.60 *** | 0.4 | G2 | 4.45 | 14.89 | 2.06 *** | 0.43 |
| C7 | 10.63 | 4.53 | 2.71 *** | 0.23 | G3 | 3.51 | 13.6 | 2.03 *** | 0.22 |
| C8 | 14.43 | 4.99 | 6.26 *** | 0.51 | G4 | 3.35 | 11.29 | 1.15 *** | 0.14 |
| C | 8.37 | 3.6 | 1.01 *** | 0.09 | G5 | 3.13 | 11.68 | 0.86 *** | 0.13 |
| D1 | 7.74 | 10.8 | 5.30 *** | 0.7 | G | 2.35 | 10.09 | 0.75 *** | 0.05 |
| D2 | 12.2 | 10.07 | 8.31 *** | 1.51 | aA | 24.59 | 5.3 | 17.64 *** | 1.69 |
| D3 | 10.83 | 10.98 | 8.29 *** | 1.41 | aB | 31.86 | 5.32 | 35.28 *** | 2.88 |
| D4 | 8.31 | 10.61 | 2.93 *** | 0.77 | aD | 55.82 | 6.78 | 162.33 *** | 14.36 |
| D5 | 10.84 | 6.81 | 4.96 *** | 0.54 | aE | 86.62 | 3.96 | 122.95 *** | 11.8 |
| D6 | 8.97 | 4.73 | 2.86 *** | 0.18 | aDE22 | 127.87 | 8.16 | 972.33 *** | 109.06 |
| D7 | 8.99 | 4.87 | 2.70 *** | 0.19 | aDE55 | 137.05 | 8.56 | 967.25 *** | 137.88 |
| D8 | 10.94 | 6.56 | 4.17 *** | 0.51 | aG | 86.31 | 13.18 | 679.38 *** | 129.45 |
| Variable | PC1 * | PC2 * | PC3 * | Variable | PC1 * | PC2 * | PC3 * |
|---|---|---|---|---|---|---|---|
| A1 | 0.094 | 0.103 | −0.062 | D | 0.081 | −0.207 | 0.236 |
| A2 | 0.149 | −0.071 | 0.04 | E1 | −0.034 | 0.104 | 0.261 |
| A3 | 0.145 | −0.069 | 0.048 | E2 | 0.125 | 0.197 | −0.01 |
| A4 | 0.151 | −0.062 | 0.033 | E3 | 0.146 | −0.011 | −0.176 |
| A5 | 0.141 | −0.04 | 0.044 | E4 | 0.079 | −0.1 | −0.216 |
| A | 0.148 | −0.069 | 0.039 | E5 | 0.137 | 0.02 | −0.053 |
| B1 | 0.066 | 0.217 | 0.14 | E6 | 0.132 | 0.122 | 0.084 |
| B2 | 0.15 | −0.077 | 0.058 | E7 | 0.145 | −0.073 | −0.157 |
| B3 | 0.118 | 0.078 | 0.092 | E8 | 0.147 | −0.043 | −0.09 |
| B4 | 0.159 | −0.008 | 0.04 | E | 0.139 | −0.08 | −0.023 |
| B5 | 0.147 | −0.039 | 0.081 | F1 | 0.084 | 0.182 | 0.096 |
| B6 | 0.105 | −0.214 | −0.015 | F2 | 0.138 | 0.175 | 0.01 |
| B7 | 0.104 | −0.231 | −0.006 | F3 | 0.146 | 0.091 | −0.108 |
| B8 | 0.157 | 0.054 | 0.062 | F4 | 0.121 | −0.127 | −0.196 |
| B | 0.147 | −0.075 | 0.03 | F5 | 0.142 | 0.152 | 0.01 |
| C1 | 0.157 | −0.07 | 0.018 | F6 | 0.125 | 0.191 | 0.018 |
| C2 | 0.147 | 0.08 | 0.061 | F7 | 0.146 | −0.014 | −0.179 |
| C3 | 0.147 | −0.001 | −0.002 | F8 | 0.148 | 0.08 | −0.089 |
| C4 | 0.154 | −0.02 | −0.007 | F | 0.132 | 0.02 | 0.001 |
| C5 | 0.156 | −0.005 | 0.033 | G1 | 0.14 | 0.114 | −0.092 |
| C6 | 0.132 | 0.092 | 0.122 | G2 | 0.092 | 0.124 | 0.104 |
| C7 | 0.148 | −0.066 | 0.052 | G3 | 0.137 | 0.049 | −0.121 |
| C8 | 0.155 | 0.041 | 0.024 | G4 | 0.121 | 0.183 | 0.053 |
| C | 0.15 | −0.068 | 0.035 | G5 | 0.14 | 0.105 | −0.06 |
| D1 | 0.096 | −0.243 | −0.048 | G | 0.133 | 0.085 | −0.034 |
| D2 | 0.044 | 0.245 | 0.156 | aA | −0.01 | 0.249 | −0.02 |
| D3 | 0.112 | −0.017 | −0.246 | aB | −0.046 | 0.226 | 0.096 |
| D4 | 0.028 | −0.052 | 0.392 | aD | 0.089 | 0.202 | −0.224 |
| D5 | 0.13 | −0.132 | 0.098 | aE | 0.067 | 0.196 | −0.099 |
| D6 | 0.138 | −0.141 | 0.119 | aDE22 | 0.104 | −0.15 | −0.071 |
| D7 | 0.119 | −0.141 | 0.211 | aDE55 | 0.063 | −0.015 | 0.186 |
| D8 | 0.108 | −0.079 | 0.269 | aG | −0.063 | 0.017 | 0.205 |
| PC1 | PC2 | PC3 | |||||
| Eigenvalue | 36.56 | 8.48 | 5.6 | ||||
| Variance (%) | 57.13 | 13.26 | 8.76 | ||||
| Accumulative variance (%) | 57.13 | 70.4 | 79.15 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maceda, A.; Delgado-Alvarado, A.; Salazar-Rojas, V.M.; Herrera-Cabrera, B.E. Vanilla planifolia Andrews (Orchidaceae): Labellum Variation and Potential Distribution in Hidalgo, Mexico. Diversity 2023, 15, 678. https://doi.org/10.3390/d15050678
Maceda A, Delgado-Alvarado A, Salazar-Rojas VM, Herrera-Cabrera BE. Vanilla planifolia Andrews (Orchidaceae): Labellum Variation and Potential Distribution in Hidalgo, Mexico. Diversity. 2023; 15(5):678. https://doi.org/10.3390/d15050678
Chicago/Turabian StyleMaceda, Agustín, Adriana Delgado-Alvarado, Víctor M. Salazar-Rojas, and B. Edgar Herrera-Cabrera. 2023. "Vanilla planifolia Andrews (Orchidaceae): Labellum Variation and Potential Distribution in Hidalgo, Mexico" Diversity 15, no. 5: 678. https://doi.org/10.3390/d15050678
APA StyleMaceda, A., Delgado-Alvarado, A., Salazar-Rojas, V. M., & Herrera-Cabrera, B. E. (2023). Vanilla planifolia Andrews (Orchidaceae): Labellum Variation and Potential Distribution in Hidalgo, Mexico. Diversity, 15(5), 678. https://doi.org/10.3390/d15050678