

Morphometric Analysis of a Trapdoor Spider (Araneae, Idiopidae) across Different Brazilian Biomes Reveals the Geographic Variation of Spiders from the Caatinga Biome

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Definition and Measurement of Morphometric Variables
2.2. Linear Morphometric Analysis
2.3. Geometric Morphometric Analysis
2.3.1. Geometric Morphometric Analysis Based on Outlines
2.3.2. Geometric Morphometric Analysis Based in Landmarks
3. Results
3.1. Linear Morphometry
3.2. Geometric Morphometry
3.2.1. Copulatory Bulb
3.2.2. Sternum
3.2.3. Eye Arrangement
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mayr, E. Populations, Species, and Evolution: An Abridgment of Animal Species and Evolution; Harvard University Press: Cambridge, MA, USA, 1970; Volume 19. [Google Scholar]
- Gould, S.J.; Johnston, R.F. Geographic variation. Annu. Rev. Ecol. Syst. 1972, 3, 457–498. [Google Scholar] [CrossRef]
- Endler, J.A. Geographic Variation, Speciation and Clines; MPB-10; Princeton University Press: Princeton, NJ, USA, 1977; Volume 10. [Google Scholar]
- Chown, S.L.; Gaston, K.J. Body size variation in insects: A macroecological perspective. Biol. Rev. 2010, 85, 139–169. [Google Scholar] [CrossRef]
- Werneck, F.P. The diversification of eastern South American open vegetation biomes: Historical biogeography and perspectives. Quat. Sci. Rev. 2011, 30, 1630–1648. [Google Scholar] [CrossRef]
- Rull, V.; Carnaval, A.C. (Eds.) Neotropical Diversification: Patterns and Processes; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Reyment, R.A. Morphometrics: An historical essay. In Morphometrics for Nonmorphometricians; Elewa, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 9–24. [Google Scholar] [CrossRef]
- Zelditch, M.L.; Swiderski, D.L.; Sheets, H.D. Geometric Morphometrics for Biologists: A Primer; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Klingenberg, C.P. Evolution and development of shape: Integrating quantitative approaches. Nat. Rev. Genet. 2010, 11, 623–635. [Google Scholar] [CrossRef]
- Zúniga-Reinoso, Á.; Benítez, H.A. The overrated use of the morphological cryptic species concept: An example with Nyctelia darkbeetles (Coleoptera: Tenebrionidae) using geometric morphometrics. Zool. Anz.—J. Comp. Zool. 2015, 255, 47–53. [Google Scholar] [CrossRef]
- Struck, T.H.; Feder, J.L.; Bendiksby, M.; Birkeland, S.; Cerca, J.; Gusarov, V.I.; Kistenich, S.; Larsson, K.H.; Liow, L.H.; Nowak, M.D.; et al. Finding evolutionary processes hidden in cryptic species. Trends Ecol. Evol. 2018, 33, 153–163. [Google Scholar] [CrossRef] [Green Version]
- Recoder, R.S.; Ribeiro, M.C.; Rodrigues, M.T. Spatial variation in morphometry in Vanzosaura rubricauda (Squamata, Gymnophthalmidae) from open habitats of South America and its environmental correlates. S. Am. J. Herpetol. 2013, 8, 186–197. [Google Scholar] [CrossRef]
- Álvarez-Varas, R.; Véliz, D.; Vélez-Rubio, G.M.; Fallabrino, A.; Zárate, P.; Heidemeyer, M.; Godoy, D.A.; Benítez, H.A. Identifying genetic lineages through shape: An example in a cosmopolitan marine turtle species using geometric morphometrics. PLoS ONE 2019, 14, e0223587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilaseca, C.; Méndez, M.A.; Pinto, C.F.; Lemic, D.; Benítez, H.A. Unraveling the morphological variation of Triatoma infestans in the peridomestic habitats of Chuquisaca Bolivia: A Geometric Morphometric Approach. Insects 2021, 12, 185. [Google Scholar] [CrossRef]
- Nattero, J.; Piccinali, R.V.; Macedo Lopes, C.; Hernández, M.L.; Abrahan, L.; Lobbia, P.A.; Rodríguez, C.S.; Carbajal de la Fuente, A.L. Morphometric variability among the species of the Sordida subcomplex (Hemiptera: Reduviidae: Triatominae): Evidence for differentiation across the distribution range of Triatoma sordida. Parasites Vectors 2017, 10, 4124. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.H.; Johnson, N.A.; Inoue, K.; Doyle, R.D.; Randklev, C.R. Integrative taxonomy reveals a new species of freshwater mussel, Potamilus streckersoni sp. nov. (Bivalvia: Unionidae): Implications for conservation and management. Syst. Biodivers. 2019, 17, 331–348. [Google Scholar] [CrossRef] [Green Version]
- Aglagane, A.; Tofilski, A.; Er-Rguibi, O.; Laghzaoui, E.M.; Kimdil, L.; El Mouden, E.H.; Fuchs, S.; Oleksa, A.; Aamiri, A.; Aourir, M. Geographical variation of honey bee (Apis mellifera L. 1758) populations in South-Eastern Morocco: A geometric morphometric analysis. Insects 2022, 13, 288. [Google Scholar] [CrossRef]
- Ferreira, V.S.; Aguiar CM, L.; Costa, M.A.; Silva, J.G. Morphometric analysis of populations of Centris aenea lepeletier (Hymenoptera: Apidae) from Northeastern Brazil. Neotrop. Entomol. 2011, 40, 97–102. [Google Scholar] [CrossRef] [Green Version]
- da Silva AC, S.; Nunes, L.A.; de Lima Batista, W.; Lhano, M.G. Morphometric variation among males of Orphulella punctata (De Geer, 1773)(Acrididae: Gomphocerinae) from different biomes in Brazil. J. Orthoptera Res. 2018, 27, 163–171. [Google Scholar] [CrossRef]
- Regueira, J.C.; Damasceno, E.M.; Iannuzzi, L. Shape variation of Cydianerus latruncularius (Coleoptera, Curculionidae) across biomes and sexes. Zool. Anz. 2020, 289, 96–107. [Google Scholar] [CrossRef]
- Cordeiro-Estrela, P.; Baylac, M.; Denys, C.; Marinho-Filho, J. Interspecific patterns of skull variation between sympatric Brazilian vesper mice: Geometric morphometrics assessment. J. Mammal. 2006, 87, 1270–1279. [Google Scholar] [CrossRef]
- Coyle, F.A. Systematics of the trapdoor spider genus Aliatypus (Araneae: Antrodiaetidae). Psyche 1974, 81, 431–500. [Google Scholar] [CrossRef] [Green Version]
- Coyle, F.A. A revision of the funnelweb mygalomorph spider subfamily Ischnothelinae (Araneae, Dipluridae). In Bulletin of the American Museum of Natural History; No. 226; American Museum of Natural History: New York, NY, USA, 1995. [Google Scholar]
- Crews, S.C.; Hedin, M. Studies of morphological and molecular phylogenetic divergence in spiders (Araneae: Homalonychus) from the American southwest, including divergence along the Baja California Peninsula. Mol. Phylogenet. Evol. 2006, 38, 470–487. [Google Scholar] [CrossRef] [PubMed]
- Entling, W.; Schmidt-Entling, M.H.; Bacher, S.; Brandl, R.; Nentwig, W. Body size–climate relationships of European spiders. J. Biogeogr. 2010, 37, 477–485. [Google Scholar] [CrossRef]
- Puzin, C.; Leroy, B.; Pétillon, J. Intra-and inter-specific variation in size and habitus of two sibling spider species (Araneae: Lycosidae): Taxonomic and biogeographic insights from sampling across Europe. Biol. J. Linn. Soc. 2014, 113, 85–96. [Google Scholar] [CrossRef]
- Krehenwinkel, H.; Graze, M.; Rödder, D.; Tanaka, K.; Baba, Y.G.; Muster, C.; Uhl, G. A phylogeographical survey of a highly dispersive spider reveals eastern Asia as a major glacial refugium for Palaearctic fauna. J. Biogeogr. 2016, 43, 1583–1594. [Google Scholar] [CrossRef]
- Wong, M.K.; Woodman, J.D.; Rowell, D.M. Short-range phenotypic divergence among genetically distinct parapatric populations of an Australian funnel-web spider. Ecol. Evol. 2017, 7, 5094–5102. [Google Scholar] [CrossRef] [PubMed]
- Korba, J.; Opatova, V.; Calatayud-Mascarell, A.; Enguídanos, A.; Bellvert, A.; Adrián, S.; Sánchez-Vialas, A.; Arnedo, M.A. Systematics and phylogeography of western Mediterranean tarantulas (Araneae: Theraphosidae). Zool. J. Linn. Soc. 2022, 196, 845–884. [Google Scholar] [CrossRef]
- Bond, J.E.; Stockman, A.K. An integrative method for delimiting cohesion species: Finding the population-species interface in a group of Californian trapdoor spiders with extreme genetic divergence and geographic structuring. Syst. Biol. 2008, 57, 628–646. [Google Scholar] [CrossRef] [Green Version]
- Costa-Schmidt, L.E.; de Araújo, A.M. Genitalic variation and taxonomic discrimination in the semi-aquatic spider genus Paratrechalea (Araneae: Trechaleidae). J. Arachnol. 2010, 38, 242–249. [Google Scholar] [CrossRef]
- Fernández-Montraveta, C.; Marugan-Lobon, J. Geometric morphometrics reveals sex-differential shape allometry in a spider. PeerJ 2017, 5, e3617. [Google Scholar] [CrossRef] [Green Version]
- Valdez-Mondragón, A.; Navarro-Rodríguez, C.I.; Solís-Catalán, K.P.; Cortez-Roldán, M.R.; Juárez-Sánchez, A.R. Under an integrative taxonomic approach: The description of a new species of the genus Loxosceles (Araneae, Sicariidae) from Mexico City. ZooKeys 2019, 892, 93. [Google Scholar] [CrossRef]
- Wilson, J.D.; Zapata, L.V.; Barone, M.L.; Cotoras, D.D.; Poy, D.; Ramírez, M.J. Geometric morphometrics reveal sister species in sympatry and a cline in genital morphology in a ghost spider genus. Zool. Scr. 2021, 50, 485–499. [Google Scholar] [CrossRef]
- Lapinski, W.; Walther, P.; Tschapka, M. Morphology reflects microhabitat preferences in an assemblage of neotropical wandering spiders. Zoomorphology 2015, 134, 219–236. [Google Scholar] [CrossRef]
- Wolff, J.O.; Wierucka, K.; Paterno, G.B.; Coddington, J.A.; Hormiga, G.; Kelly, M.B.; Herberstein, M.E.; Ramírez, M.J. Stabilized morphological evolution of spiders despite mosaic changes in foraging ecology. Syst. Biol. 2022, 71, 1487–1503. [Google Scholar] [CrossRef]
- Kallal, R.J.; Moore, A.J.; Hormiga, G. The shape of weaver: Investigating shape disparity in orb-weaving spiders (Araneae, Araneidae) using geometric morphometrics. Evol. Biol. 2019, 46, 317–331. [Google Scholar] [CrossRef]
- Bond, J.E.; Beamer, D.A. A morphometric analysis of mygalomorph spider carapace shape and its efficacy as a phylogenetic character (Araneae). Invertebr. Syst. 2006, 20, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Brandt, S.; Sole, C.; Lyle, R.; Pirk, C. Geometric Morphometric Analysis of Ocular Patterns as a Species Identifier in the South African Endemic Trapdoor Spider Genus Stasimopus Simon, 1892 (Araneae, Mygalomorphae, Stasimopidae). Evol. Biol. 2023, 1–15. [Google Scholar] [CrossRef]
- Ríos-Tamayo, D. Variación intra-específica en machos de Actinopus sp. (Mygalomorphae: Actinopodidae) del norte argentino. Acta Zoológica Lilloana 2016, 60, 78–88. [Google Scholar]
- Bond, J.E.; Hedin, M.C.; Ramirez, M.G.; Opell, B.D. Deep molecular divergence in the absence of morphological and ecological change in the Californian coastal dune endemic trapdoor spider Aptostichus simus. Mol. Ecol. 2001, 10, 899–910. [Google Scholar] [CrossRef]
- Hendrixson, B.E.; Bond, J.E. Testing species boundaries in the Antrodiaetus unicolor complex (Araneae: Mygalomorphae: Antrodiaetidae):“paraphyly” and cryptic diversity. Mol. Phylogenet. Evol. 2005, 36, 405–416. [Google Scholar] [CrossRef]
- Cooper, S.J.; Harvey, M.S.; Saint, K.M.; Main, B.Y. Deep phylogeographic structuring of populations of the trapdoor spider Moggridgea tingle (Migidae) from southwestern Australia: Evidence for long-term refugia within refugia. Mol. Ecol. 2011, 20, 3219–3236. [Google Scholar] [CrossRef] [PubMed]
- Mason, L.D.; Wardell-Johnson, G.; Main, B.Y. The longest-lived spider: Mygalomorphs dig deep, and persevere. Pac. Conserv. Biol. 2018, 24, 203–206. [Google Scholar] [CrossRef] [Green Version]
- Opatova, V.; Hamilton, C.A.; Hedin, M.; De Oca, L.M.; Král, J.; Bond, J.E. Phylogenetic systematics and evolution of the spider infraorder Mygalomorphae using genomic scale data. Syst. Biol. 2020, 69, 671–707. [Google Scholar] [CrossRef]
- Fonseca-Ferreira, R.; Guadanucci JP, L.; Yamamoto, F.U.; Brescovit, A.D. Taxonomic revision of the Neotropical spiders of the genus Idiops Perty, 1833 (Araneae, Idiopidae), with description of four new species. Eur. J. Taxon. 2021, 780, 1–71. [Google Scholar] [CrossRef]
- Atkinson, D. Temperature and organism size: A biological law for ectotherms. Adv. Ecol. Res. 1994, 25, 1–58. [Google Scholar] [CrossRef]
- Horne, C.R.; Hirst, A.G.; Atkinson, D. Temperature-size responses match latitudinal-size clines in arthropods, revealing critical differences between aquatic and terrestrial species. Ecol. Lett. 2015, 18, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Lira, A.F.; Foerster, S.I.; Albuquerque, C.M.; Moura, G.J. Contrasting patterns at interspecific and intraspecific levels in scorpion body size across a climatic gradient from rainforest to dryland vegetation. Zoology 2021, 146, 125908. [Google Scholar] [CrossRef]
- Cushman, J.H.; Lawton, J.H.; Manly BF, J. Latitudinal patterns in European ant assemblages: Variation in species richness and body size. Oecologia 1993, 95, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Remmert, H. Body size of terrestrial arthropods and biomass of their populations in relation to the abiotic parameters of their milieu. Oecologia 1981, 50, 12–13. [Google Scholar] [CrossRef]
- Gouveia, S.F.; Correia, I. Geographical clines of body size in terrestrial amphibians: Water conservation hypothesis revisited. J. Biogeogr. 2016, 43, 2075–2084. [Google Scholar] [CrossRef]
- Kumar, A. A tragic loss: Fire at Instituto Butantan, Brazil. Toxicon Off. J. Int. Soc. Toxinology 2010, 56, 1528–1529. [Google Scholar] [CrossRef]
- IBGE. Biomas e Sistema Costeiro-Marinho do Brasil: Compatível com a Escala 1: 250 000; Série Relatórios Metodológicos; v. 45; IBGE: Rio de Janeiro, Brazil, 2019. [Google Scholar]
- Strauss, R.E. Discriminating groups of organisms. In Morphometrics for Nonmorphometricians; Springer: Berlin/Heidelberg, Germany, 2010; pp. 73–91. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Kuhl, F.P.; Giardina, C.R. Elliptic Fourier features of a closed contour. Comput. Graph. Image Process. 1982, 18, 236–258. [Google Scholar] [CrossRef]
- Bonhomme, V.; Picq, S.; Gaucherel, C.; Claude, J. Momocs: Outline analysis using R. J. Stat. Softw. 2014, 56, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Rohlf, F.J. tpsDig. 2001. Available online: https://life2.bio.sunysb.edu/ee/rohlf/software.html (accessed on 13 July 2022).
- Klingenberg, C.P. MorphoJ: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Main, B.Y. Adaptations to arid habitats by mygalomorph spiders. In Evolution of the Flora and Fauna of Arid Australia; Barker, W.R., Greenslade, P.J., Eds.; Australian Systematic Botany Society: Frewville, Australia, 1982. [Google Scholar]
- Canals, M.; Veloso, C.; Solís, R. Adaptation of the spiders to the environment: The case of some Chilean species. Front. Physiol. 2015, 6, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werneck, F.P.; Costa, G.C.; Colli, G.R.; Prado, D.E.; Sites, J.W., Jr. Revisiting the historical distribution of Seasonally Dry Tropical Forests: New insights based on palaeodistribution modelling and palynological evidencegeb. Glob. Ecol. Biogeogr. 2011, 20, 272–288. [Google Scholar] [CrossRef]
- Bucher, E.H. Chaco and Caatinga—South American arid savannas, woodlands and thickets. In Ecology of Tropical Savannas; Springer: Berlin/Heidelberg, Germany, 1982; pp. 48–79. [Google Scholar] [CrossRef]
- Prado, D.E. As caatingas da América do Sul. Ecol. Conserv. Caatinga 2003, 2, 3–74. [Google Scholar]
- Silva, J.M.C.D.; Barbosa, L.C.F.; Leal, I.R.; Tabarelli, M. The Caatinga: Understanding the challenges. In Caatinga; Springer: Cham, Switzerland, 2017; pp. 3–19. [Google Scholar] [CrossRef]
- de Oliveira, F.R.C.; Passos, D.C.; Borges-Nojosa, D.M. Ecology of the lizard Gonatodes humeralis (Sphaerodactylidae) in a coastal area of the Brazilian semiarid: What differs from the Amazonian populations? J. Arid. Environ. 2021, 190, 104506. [Google Scholar] [CrossRef]
- McCluney, K.E.; Date, R.C. The effects of hydration on growth of the house cricket, Acheta domesticus. J. Insect Sci. 2008, 8, 32. [Google Scholar] [CrossRef] [Green Version]
- Grizante, M.B.; Brandt, R.; Kohlsdorf, T. Evolution of body elongation in gymnophthalmid lizards: Relationships with climate. PLoS ONE 2012, 7, e49772. [Google Scholar] [CrossRef] [Green Version]
- Vanzolini, P.E. Problemas faunísticos do Cerrado. Simpósio Sobre o Cerrado 1963, 307, 1. [Google Scholar]
- Magalhaes, I.L.; Oliveira, U.; Santos, F.R.; Vidigal, T.H.; Brescovit, A.D.; Santos, A.J. Strong spatial structure, Pliocene diversification and cryptic diversity in the Neotropical dry forest spider Sicarius cariri. Mol. Ecol. 2014, 23, 5323–5336. [Google Scholar] [CrossRef]
- Werneck, F.P.; Gamble, T.; Colli, G.R.; Rodrigues, M.T.; Sites, J.W., Jr. Deep diversification and long-term persistence in the South American ‘dry diagonal’: Integrating continent-wide phylogeography and distribution modeling of geckos. Evol. Int. J. Org. Evol. 2012, 66, 3014–3034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca, E.M.; Gehara, M.; Werneck, F.P.; Lanna, F.M.; Colli, G.R.; Sites, J.W., Jr.; Rodrigues, M.T.; Garda, A.A. Diversification with gene flow and niche divergence in a lizard species along the South American “diagonal of open formations”. J. Biogeogr. 2018, 45, 1688–1700. [Google Scholar] [CrossRef]
- Thome MT, C.; Carstens, B.C.; Rodrigues, M.T.; Alexandrino, J.; Haddad, C.F. Genomic data from the Brazilian sibilator frog reveal contrasting pleistocene dynamics and regionalism in two South American dry biomes. J. Biogeogr. 2021, 48, 1112–1123. [Google Scholar] [CrossRef]
- Moura CC, D.M.; Araujo HF, D.; Aleixo, A.; Wink, M.; Fernandes, A.M. The role of landscape change and paleoclimatic events in shaping the evolutionary history of the Polioptila gnatcatchers (Passeriformes, Polioptilidae) with emphasis on species associated with open habitats. J. Avian Biol. 2018, 49, jav-012409. [Google Scholar] [CrossRef]
- Rocha, A.V.; Cabanne, G.S.; Aleixo, A.; Silveira, L.F.; Tubaro, P.; Caparroz, R. Pleistocene climatic oscillations associated with landscape heterogeneity of the South American dry diagonal explains the phylogeographic structure of the narrow-billed woodcreeper (Lepidocolaptes angustirostris, Dendrocolaptidae). J. Avian Biol. 2020, 51, 1–13. [Google Scholar] [CrossRef]
- Newton, L.G.; Starrett, J.; Jochim, E.E.; Bond, J.E. Phylogeography and cohesion species delimitation of California endemic trapdoor spiders within the Aptostichus icenoglei sibling species complex (Araneae: Mygalomorphae: Euctenizidae). Ecol. Evol. 2023, 13, e10025. [Google Scholar] [CrossRef]
- Newton, L.G.; Starrett, J.; Hendrixson, B.E.; Derkarabetian, S.; Bond, J.E. Integrative species delimitation reveals cryptic diversity in the southern Appalachian Antrodiaetus unicolor (Araneae: Antrodiaetidae) species complex. Mol. Ecol. 2020, 29, 2269–2287. [Google Scholar] [CrossRef]
- Satler, J.D.; Carstens, B.C.; Hedin, M. Multilocus species delimitation in a complex of morphologically conserved trapdoor spiders (Mygalomorphae, Antrodiaetidae, Aliatypus). Syst. Biol. 2013, 62, 805–823. [Google Scholar] [CrossRef] [Green Version]
- Harvey, M.S.; Main, B.Y.; Rix, M.G.; Cooper, S.J. Refugia within refugia: In situ speciation and conservation of threatened Bertmainius (Araneae: Migidae), a new genus of relictual trapdoor spiders endemic to the mesic zone of south-western Australia. Invertebr. Syst. 2015, 29, 511–553. [Google Scholar] [CrossRef]
- Ferretti, N.E.; Soresi, D.S.; Gonzalez, A.; Arnedo, M. An integrative approach unveils speciation within the threatened spider Calathotarsus simoni (Araneae: Mygalomorphae: Migidae). Syst. Biodivers. 2019, 17, 439–457. [Google Scholar] [CrossRef]
- Opatova, V.; Bond, J.E.; Arnedo, M.A. Uncovering the role of the Western Mediterranean tectonics in shaping the diversity and distribution of the trap-door spider genus Ummidia (Araneae, Ctenizidae). J. Biogeogr. 2016, 43, 1955–1966. [Google Scholar] [CrossRef]
- Opatova, V.; Arnedo, M.A. Spiders on a hot volcanic roof: Colonisation pathways and phylogeography of the Canary Islands endemic trap-door spider Titanidiops canariensis (Araneae, Idiopidae). PLoS ONE 2014, 9, e115078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eberhard, W.G.; Huber, B.A.; Briceño, R.D.; Salas, I.; Rodriguez, V. One size fits all? Relationships between the size and degree of variation in genitalia and other body parts in twenty species of insects and spiders. Evolution 1998, 52, 415–431. [Google Scholar] [CrossRef] [PubMed]
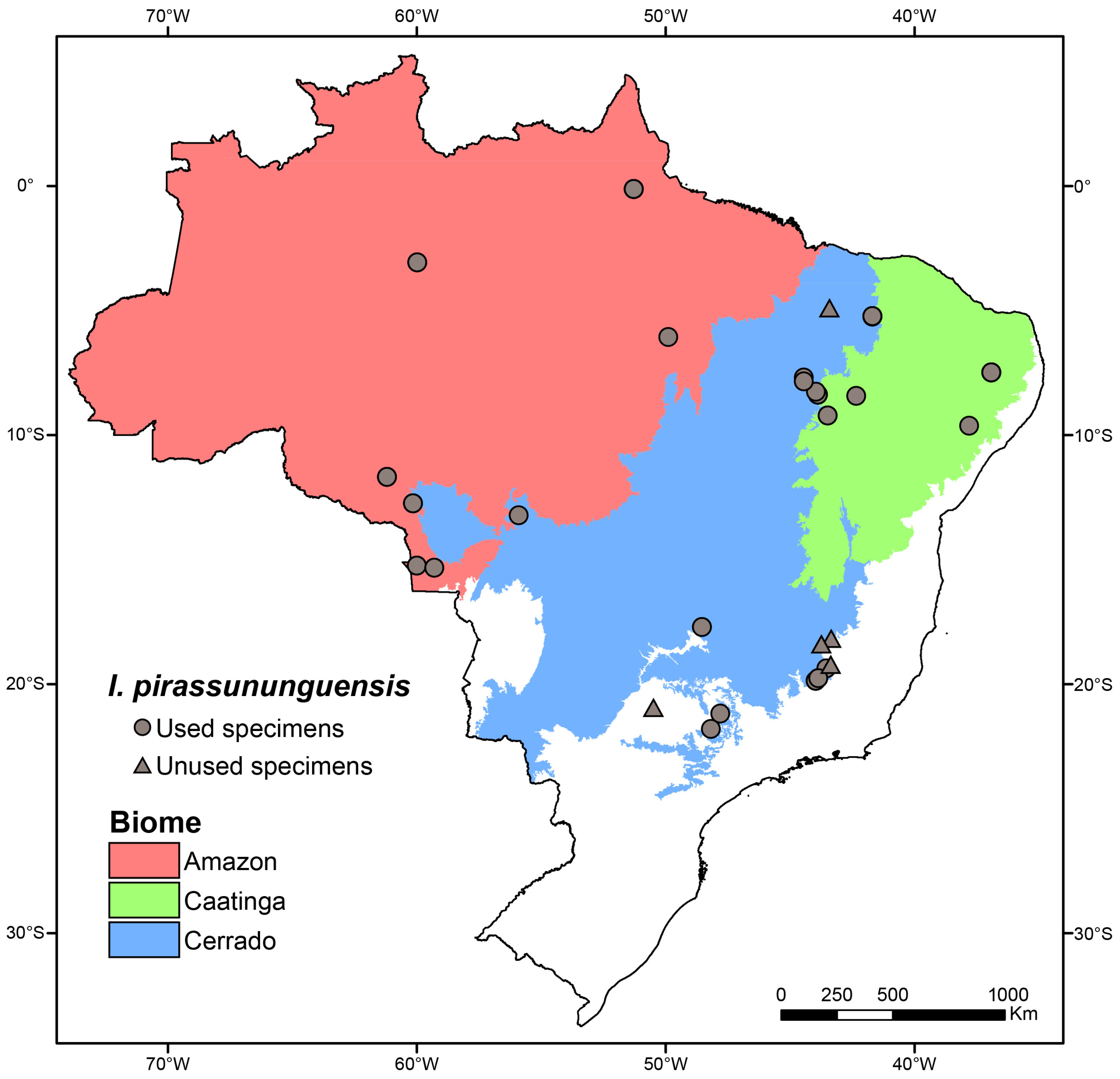




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurements | Amazon (n: 20) | Caatinga (n: 18) | Cerrado (n: 26) | Total (n: 64) |
|---|---|---|---|---|
| Carapace length | 6.96 ± 0.14 (5.53–8.04) | 7 ± 0.11 (5.94–8.12) | 7.08 ± 0.12 (5.64–8.2) | 7 ± 0.07 (5.53–8.2) |
| Carapace width | 6.51 ± 0.16 (4.97–7.56) | 6.5 ± 0.11 (5.39–7.31) | 6.65 ± 0.16 (5.07–8) | 6.56 ± 0.08 (4.97–8) |
| Sternum length | 4 ± 0.09 (3.10–4.60) | 4.09 ± 0.07 (3.35–4.73) | 4.02 ± 0.07 (3.45–4.8) | 4.07 ± 0.04 (3.10–4.8) |
| Sternum width | 3.68 ± 0.09 (2.66–4.34) | 3.53 ± 0.07 (2.82–4.09) | 3.62 ± 0.07 (2.94–4.4) | 3.61 ± 0.05 (2.66–4.4) |
| Leg 1 length | 23.4 ± 0.58 (18.36–27.1) | 23 ± 0.27 (21.17–25.63) | 23.52 ± 0.5 (17.89–27.08) | 23.34 ± 0.28 (17.89–27.08) |
| Leg 2 length | 20 ± 0.5 (15.51–23.13) | 17.5 ± 0.6 (13.92–21.54) | 20 ± 0.43 (14.48–23.1) | 19.31 ± 0.3 (13.92–23.13) |
| Leg 3 length | 17.6 ± 0.46 (12.68–20.6) | 15.37 ± 0.58 (12.23–20.46) | 17.47 ± 0.36 (13.32–20.45) | 16.93 ± 0.28 (12.23–20.6) |
| Leg 4 length | 23.95 ± 0.62 (18.26–27.96) | 20.56 ± 0.72 (16.62–26.84) | 23.67 ± 0.5 (18.15–28.15) | 22.88 ± 0.38 (16.62–28.15) |
| Pedipalp length | 12.96 ± 0.27 (10.25–14.9) | 11.08 ± 0.42 (9.01–14.85) | 12.7 ± 0.25 (10.48–15.16) | 12.33 ± 0.2 (9.01–15.16) |
| a. Linear measurements (PERMANOVA) | b. Bulb copulatory (contingency table) | ||||||
| Biome | Amazon | Caatinga | Cerrado | Biome | Amazon | Caatinga | Cerrado |
| Amazon | - | <0.01 | 0.914 | Amazon | 65% | 10% | 25% |
| Caatinga | 14.74 * | - | <0.01 | Caatinga | 0% | 78% | 22% |
| Cerrado | 1.16 | 14.16 * | - | Cerrado | 11.50% | 11.50% | 77% |
| c. Sternum (Mahalanobis distance) | d. Sternum (Procrustes distance) | ||||||
| Biome | Amazon | Caatinga | Cerrado | Biome | Amazon | Caatinga | Cerrado |
| Amazon | - | <0.01 | 0.0135 | Amazon | - | <0.01 | 0.583 |
| Caatinga | 2.49 * | - | <0.01 | Caatinga | 0.03 * | - | <0.01 |
| Cerrado | 1.56 | 2.2 * | - | Cerrado | 0 | 0.03 * | - |
| e. Eye arrangement (Mahalanobis distance) | f. Eye arrangement (Procrustes distance) | ||||||
| Biome | Amazon | Caatinga | Cerrado | Biome | Amazon | Caatinga | Cerrado |
| Amazon | - | <0.01 | 0.0912 | Amazon | - | <0.01 | 0.324 |
| Caatinga | 3.86 * | - | <0.01 | Caatinga | 0.03 * | - | <0.01 |
| Cerrado | 1.3 | 3.17 * | - | Cerrado | 0.02 | 0.03 * | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fonseca-Ferreira, R.; Morales, M.J.A.; Carvalho, L.S.; Guadanucci, J.P.L. Morphometric Analysis of a Trapdoor Spider (Araneae, Idiopidae) across Different Brazilian Biomes Reveals the Geographic Variation of Spiders from the Caatinga Biome. Diversity 2023, 15, 861. https://doi.org/10.3390/d15070861
Fonseca-Ferreira R, Morales MJA, Carvalho LS, Guadanucci JPL. Morphometric Analysis of a Trapdoor Spider (Araneae, Idiopidae) across Different Brazilian Biomes Reveals the Geographic Variation of Spiders from the Caatinga Biome. Diversity. 2023; 15(7):861. https://doi.org/10.3390/d15070861
Chicago/Turabian StyleFonseca-Ferreira, Rafael, Millke Jasmine A. Morales, Leonardo S. Carvalho, and José Paulo L. Guadanucci. 2023. "Morphometric Analysis of a Trapdoor Spider (Araneae, Idiopidae) across Different Brazilian Biomes Reveals the Geographic Variation of Spiders from the Caatinga Biome" Diversity 15, no. 7: 861. https://doi.org/10.3390/d15070861
APA StyleFonseca-Ferreira, R., Morales, M. J. A., Carvalho, L. S., & Guadanucci, J. P. L. (2023). Morphometric Analysis of a Trapdoor Spider (Araneae, Idiopidae) across Different Brazilian Biomes Reveals the Geographic Variation of Spiders from the Caatinga Biome. Diversity, 15(7), 861. https://doi.org/10.3390/d15070861