

Paleoclimatic Reconstruction Based on the Late Pleistocene San Josecito Cave Stratum 720 Fauna Using Fossil Mammals, Reptiles, and Birds


Abstract
:1. Introduction
2. Materials and Methods
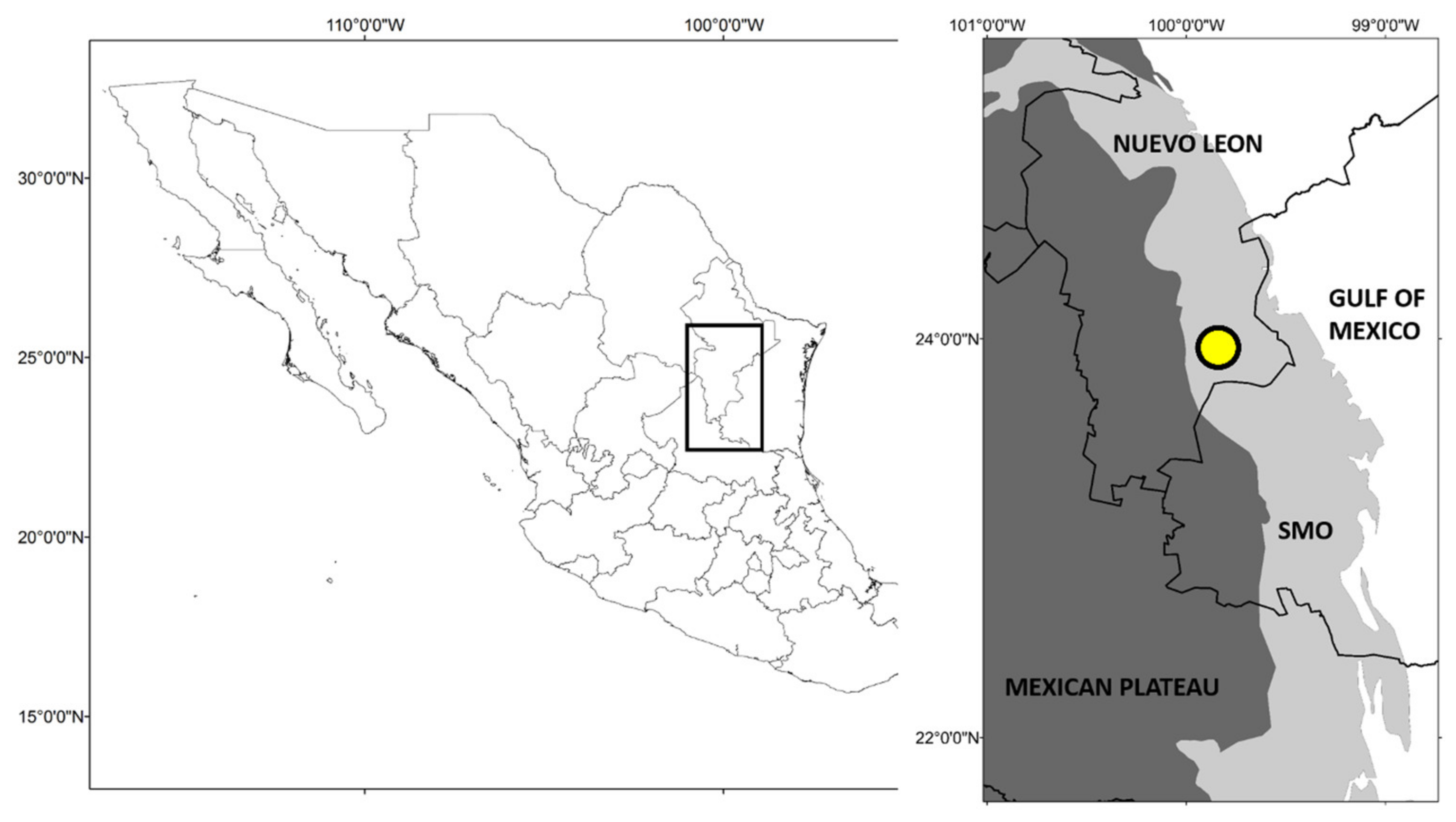
2.1. Study Area
2.2. Material
2.3. Ocurrence Data
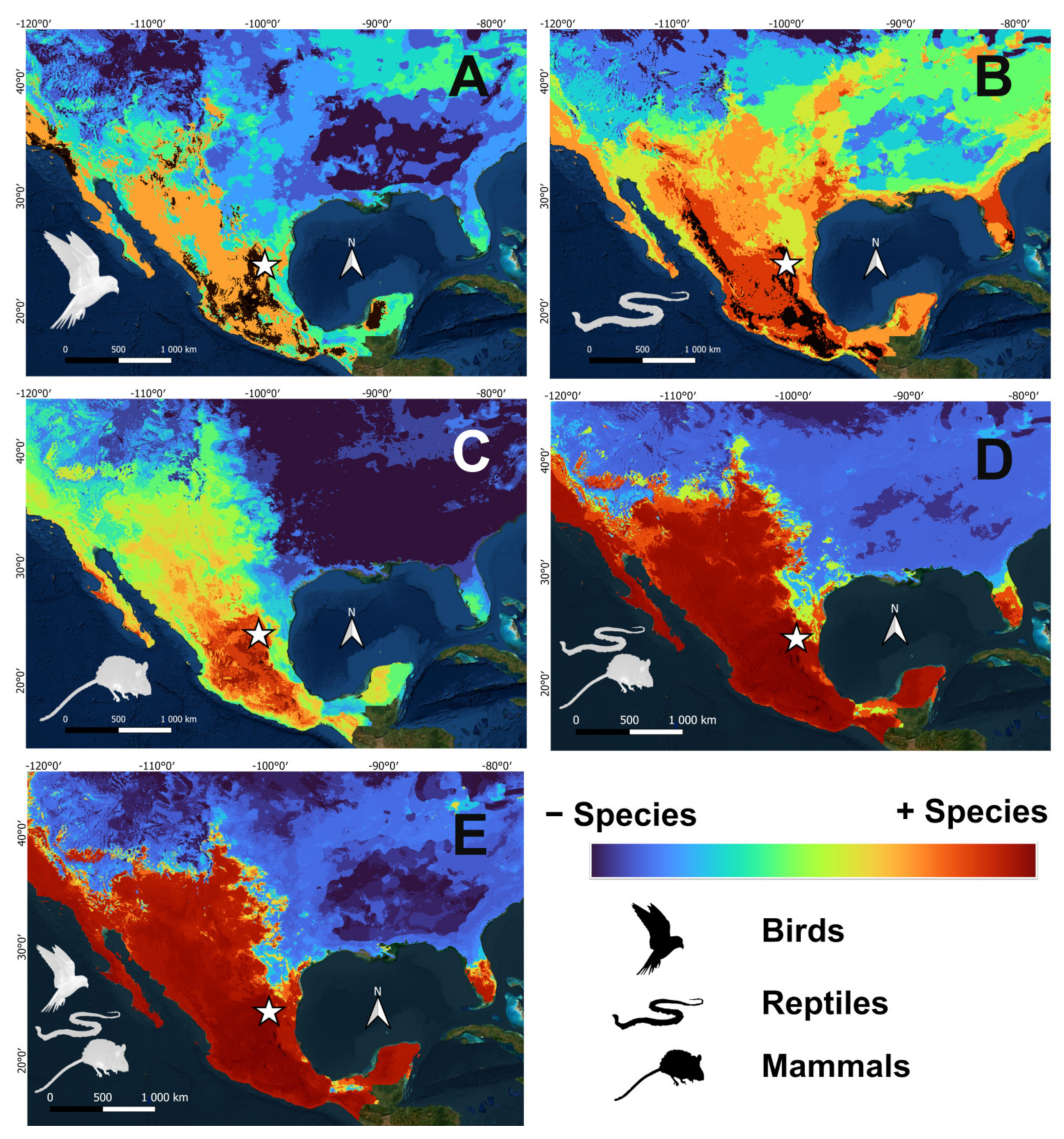
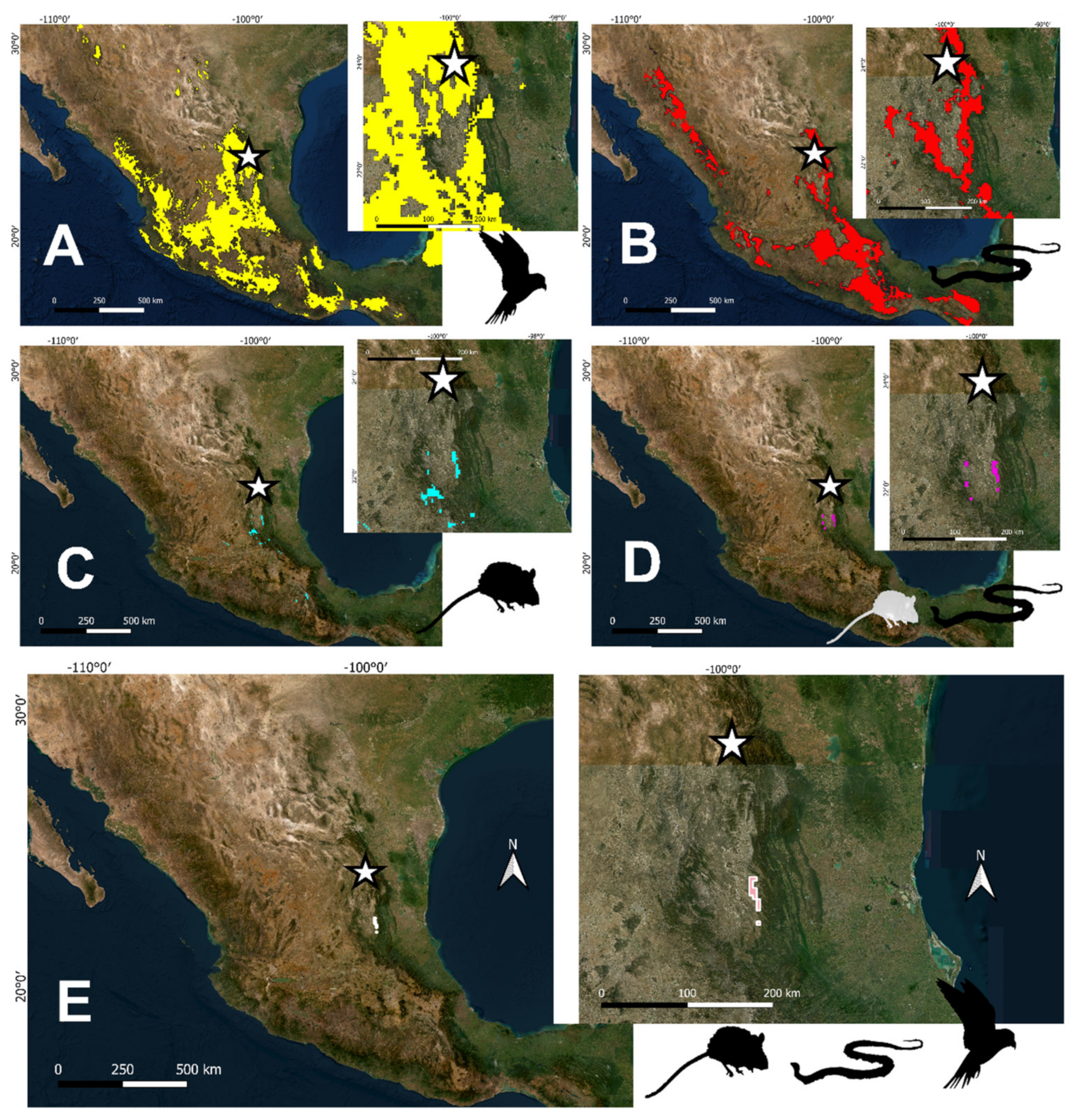
2.4. Potential Distribution Maps
2.5. Paleoclimatic Reconstruction
3. Results
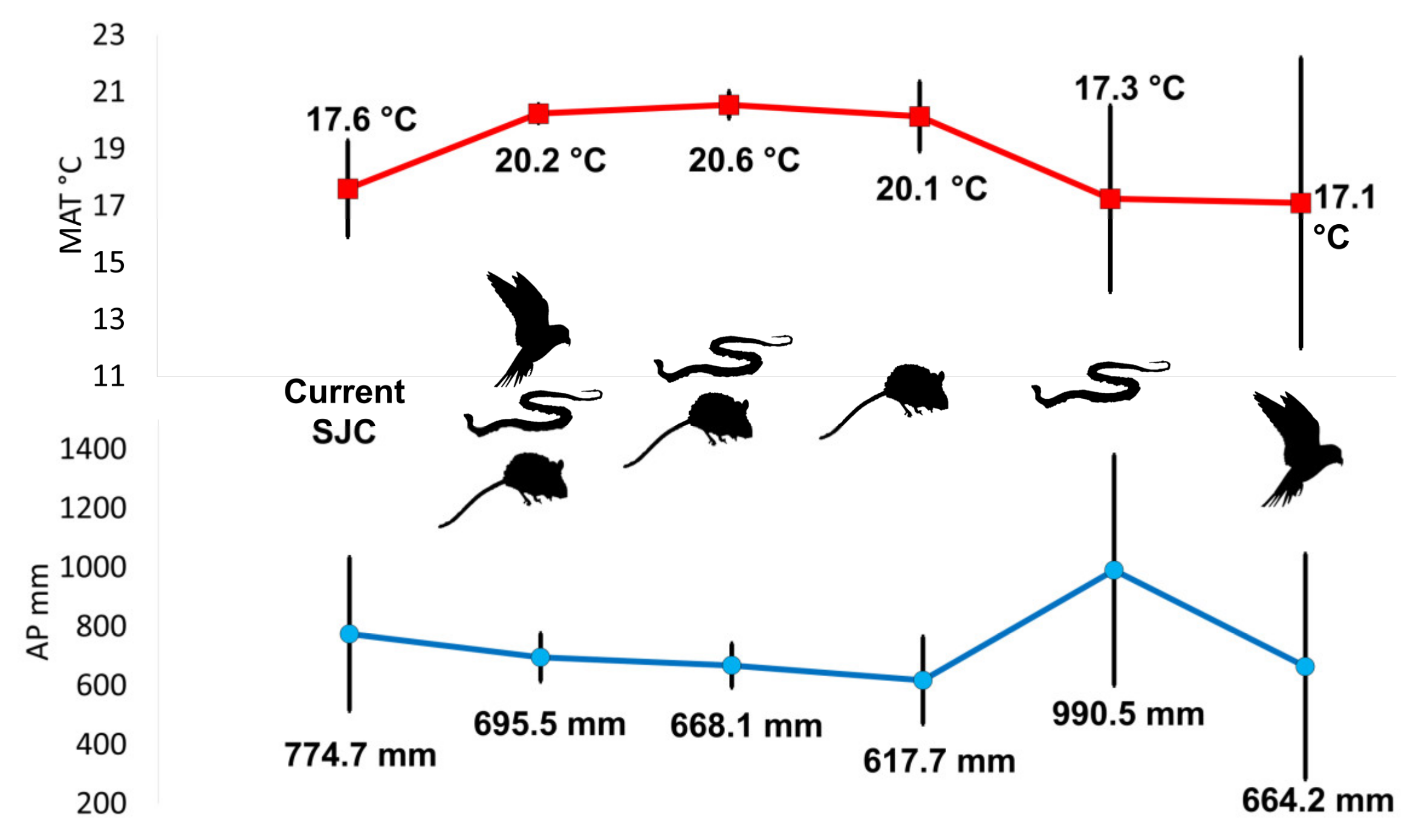
3.1. Paleoclimatic Inferences of the Late Pleistocene from San Josecito Cave
3.2. Paleoclimatic Inferences between Vertebrate Groups
4. Discussion
4.1. Paleoclimatic Inferences between Vertebrate Groups
4.2. Paleoclimatic Inferences between Vertebrate Groups
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schweitzer, M.H. Paleontology in the 21st Century. Curr. Biol. 2023, 12, 487. [Google Scholar] [CrossRef] [PubMed]
- Dillon, E.M.; Dunne, E.M.; Womack, T.M.; Kouvari, M.; Larina, E.; Claytor, J.R.; Ivkic, A.; Juhn, M.; Carmona, P.S.M.; Robson, S.K.; et al. Challenges and directions in analytical paleobiology. Paleobiology 2023, 3, 1–17. [Google Scholar] [CrossRef]
- Kiessling, W.; Smith, J.A.; Raja, N.B. Improving the relevance of paleontology to climate change policy. Proc. Natl. Acad. Sci. USA 2023, 120, e2201926119. [Google Scholar] [CrossRef]
- Bradley, R.S. Paleoclimatology: Reconstructing Climates of the Quaternary, 3rd ed.; Academic Press: Oxford, UK, 2015; p. 675. [Google Scholar]
- Blois, J.L.; Zarnetske, P.L.; Fitzpatrick, M.C.; Finnegan, S. Climate change and the past, present, and future of biotic interactions. Science 2013, 341, 499–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eronen, J.T.; Polly, P.D.; Fred, M.; Damuth, J.; Frank, D.C.; Mosbrugger, V.; Scheidegger, C.; Stenseth, N.C.; Fortelius, M. Ecometrics: The traits that bind the past and present together. Integr. Zool. 2010, 5, 88–101. [Google Scholar] [CrossRef]
- Polly, P.D.; Eronen, J.T.; Fred, M.; Dietl, G.P.; Mosbrugger, V.; Scheidegger, C.; Frank, D.C.; Damuth, J.; Stenseth, N.C.; Fortelius, M. History matters: Ecometrics and integrative climate change biology. Proc. R. Soc. B Biol. Sci. 2011, 278, 1131–1140. [Google Scholar] [CrossRef] [PubMed]
- Vermillion, W.A.; Polly, P.D.; Head, J.J.; Eronen, J.T.; Lawing, A.M. Ecometrics: A trait-based approach to paleoclimate and paleoenvironmental reconstruction. In Methods in Paleoecology: Reconstructing Cenozoic Terrestrial Environments and Ecological Communities, 1st ed.; Croft, D.A., Su, D.F., Simpson, S.W., Eds.; Springer Nature: Basel, Switzerland, 2018; pp. 373–394. [Google Scholar] [CrossRef]
- Balk, M.A.; Deck, J.; Emery, K.F.; Walls, R.L.; Reuter, D.; LaFrance, R.; Arroyo-Cabrales, J.; Barrett, P.; Blois, J.; Boileau, A.; et al. A solution to the challenges of interdisciplinary aggregation and use of specimen-level trait data. iScience 2022, 25, 105101. [Google Scholar] [CrossRef]
- Blain, H.A.; Cruz-Silva, J.A.; Arenas, J.M.J.; Margari, V.; Roucoux, K. Towards a Middle Pleistocene terrestrial climate reconstruction based on herpetofaunal assemblages from the Iberian Peninsula: State of the art and perspectives. Quat. Sci. Rev. 2018, 191, 167–188. [Google Scholar] [CrossRef] [Green Version]
- Jonzén, N.; Lindén, A.; Ergon, T.; Knudsen, E.; Vik, J.O.; Rubolini, D.; Piacentini, D.; Brinch, C.; Spina, F.; Karlsson, L.; et al. Rapid advance of spring arrival dates in long-distance migratory birds. Science 2006, 312, 1959–1961. [Google Scholar] [CrossRef]
- Charmantier, A.; McCleery, R.H.; Cole, L.R.; Perrins, C.; Kruuk, L.E.B.; Sheldon, B.C. Adaptative phenotypic plasticity in response to climate change in a wild bird population. Science 2008, 320, 800–803. [Google Scholar] [CrossRef] [Green Version]
- Youngflesh, C.; Socolar, J.; Amaral, B.R.; Arab, A.; Guralnick, R.P.; Hurlbert, A.H.; LaFrance, R.; Mayor, S.J.; Miller, D.A.W.; Tingley, M.W. Migratory strategy drives species-level variation in bird sensitivity to vegetation green-up. Nat. Ecol. Evol. 2021, 5, 987–994. [Google Scholar] [CrossRef]
- McLean, N.; Kruuk, L.E.; Van Der Jeugd, H.P.; Leech, D.; van Turnhout, C.A.; van de Pol, M. Warming temperatures drive at least half of the magnitude of long-term trait changes in European birds. Proc. Natl. Acad. Sci. USA 2022, 119, e2105416119. [Google Scholar] [CrossRef]
- Arroyo-Cabrales, J.; Polaco, O.J. Caves and the Pleistocene vertebrate paleontology of Mexico. In Ice Age Cave Faunas of North America, 1st ed.; Schubert, B.W., Mead, J.I., Graham, R.W., Eds.; Indiana University Press: Bloomington, IN, USA, 2003; pp. 273–291. [Google Scholar]
- Nunez, E.E.; Macfadden, B.J.; Mead, J.I.; Baez, A. Ancient forests and grasslands in the desert: Diet and habitat of Late Pleistocene mammals from Northcentral Sonora, Mexico. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 297, 391–400. [Google Scholar] [CrossRef]
- Pérez-Crespo, V.A.; Arroyo-Cabrales, J.; Alva-Valdívia, L.M.; Morales-Puente, P.; Cienfuegos-Alvarado, E.; Otero, F.J. Estado actual de la aplicación de los marcadores biogeoquímicos en paleoecología de mamíferos del Pleistoceno tardío de México. Archaeobios 2012, 6, 53–64. [Google Scholar]
- Cuenca-Bescós, G.; Lawrence, G.; Straus González Morales, M.R.; García Pimienta, J.C. The reconstruction of past environments through small mammals: From the Mousterian to the Bronze Age in El Mirón Cave (Cantabria Spain). J. Archaeol. Sci. 2009, 36, 947–955. [Google Scholar] [CrossRef]
- Baca, M.; Popovic, D.; Lemanik, A.; Bañuls-Cardona, S.; Conard, N.J.; Cuenca-Bescós, G.; Desclaux, E.; Fewlass, H.; Garcia, J.T.; Hadravova, T.; et al. Ancient DNA reveals interstadials as a driver of common vole population dynamics during the last glacial period. J. Biogeogr. 2023, 50, 183–196. [Google Scholar] [CrossRef]
- Stock, C. The Cave of San Josecito, Mexico. Eng. Sci. 1943, 6, 10–14. [Google Scholar]
- Arroyo-Cabrales, J.; Johnson, E.; Ralph, R.W. An initial reappraisal of San Josecito Cave and its Late Pleistocene fauna. Curr. Res. Pleistocene 1989, 6, 63–64. [Google Scholar]
- Arroyo-Cabrales, J.; Johnson, E.; Cruz, J.A. San Josecito Cave and Its Paleoecological Contributions for Quaternary Studies in Mexico. Quaternary 2021, 4, 34. [Google Scholar] [CrossRef]
- GBIF. The Global Biodiversity Information Facility Ocurrence Download. Available online: https://doi.org/10.15468/dl.mqz2g8; https://doi.org/10.15468/dl.4cedb9; https://doi.org/10.15468/dl.tnfnn2; https://doi.org/10.15468/dl.qdxbwq (accessed on 19 January 2021); https://doi.org/10.15468/dl.66mfbe; https://doi.org/10.15468/dl.f7y54g; https://doi.org/10.15468/dl.zvyqn8; https://doi.org/10.15468/dl.ajd4bv; https://doi.org/10.15468/dl.4fr4gy; https://doi.org/10.15468/dl.mmp74y; https://doi.org/10.15468/dl.7wnckf; https://doi.org/10.15468/dl.gp4s6s; https://doi.org/10.15468/dl.kq95r8; https://doi.org/10.15468/dl.z7tz3n; https://doi.org/10.15468/dl.c56a7m; https://doi.org/10.15468/dl.ppbwyq; https://doi.org/10.15468/dl.6dx2nc; https://doi.org/10.15468/dl.fx2trk; https://doi.org/10.15468/dl.6bjgx2; https://doi.org/10.15468/dl.w9a6ww; https://doi.org/10.15468/dl.qc5b73; https://doi.org/10.15468/dl.5x4y4z; https://doi.org/10.15468/dl.e9jvhh; https://doi.org/10.15468/dl.xe7q6t; https://doi.org/10.15468/dl.87te5y; https://doi.org/10.15468/dl.ga4m2m; https://doi.org/10.15468/dl.uxbfe3; https://doi.org/10.15468/dl.75sj32; https://doi.org/10.15468/dl.k4kyh2; https://doi.org/10.15468/dl.yp6b6h; https://doi.org/10.15468/dl.dg4g72; https://doi.org/10.15468/dl.6m2x5e; https://doi.org/10.15468/dl.9j87yt; https://doi.org/10.15468/dl.uy285t; https://doi.org/10.15468/dl.ypz3cc (accessed on 27 July 2021).
- Kass, J.M.; Pinilla-Buitrago, G.E.; Paz, A.; Johnson, B.A.; Grisales-Betancur, V.; Meenan, S.I.; Attali, D.; Broennimann, O.; Galante, P.J.; Maitner, B.S.; et al. Wallace 2: A shiny app for modeling species niches and distributions redesigned to facilitate expansion via module contributions. Ecography 2022, 3, e06547. [Google Scholar] [CrossRef]
- Osorio-Olvera, L.; Lira-Noriega, A.; Soberón, J.; Townsend, P.A.; Falconi, M.; Contreras-Díaz, R.G.; Martínez-Meyer, E.; Barve, V.; Barve, N. ntbox: An R package with graphical user interface for modeling and evaluating multidimensional ecological niches. Methods Ecol. Evol. 2020, 11, 1199–1206. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species. Version 2021-2. Available online: https://www.iucnredlist.org (accessed on 3 September 2021).
- BirdLife International. Available online: http://www.birdlife.org (accessed on 3 September 2021).
- Campbell, J.A.; Lamar, W.W.; Brodie, E.D. The Venomous Reptiles of the Western Hemisphere; Comstock Publishing Associates: Ithaca, NY, USA, 2004; Volume 1, p. 528. [Google Scholar]
- Martínez-Méndez, N.; Méndez-De la Cruz, F.R. Molecular phylogeny of the Sceloporus torquatus species-group (Squamata: Phrynosomatidae). Zootaxa 2007, 1609, 53–68. [Google Scholar] [CrossRef] [Green Version]
- Mulcahy, D.G. Phylogeography and species boundaries of the western North American Nightsnake (Hypsiglena torquata): Revisiting the subspecies concept. Mol. Phylogenet. Evol. 2008, 46, 1095–1115. [Google Scholar] [CrossRef]
- Leache, A.D. Species tree discordance traces to phylogeographic clade boundaries in North American fence lizards (Sceloporus). Syst. Biol. 2009, 58, 547–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ernst, C.H. Storeria. CAAR 2012, 900, 900.1–900.14. [Google Scholar]
- Grummer, J.A. Testing Species Limits and Inferring the Species Tree for the Sceloporus scalaris Species Group (Squamata: Phrynosomatidae). Master’s Thesis, San Diego State University, San Diego, CA, USA, June 2013. [Google Scholar]
- Leache, A.D.; Harris, R.B.; Maliska, M.E.; Linkem, C.W. Comparative species divergence across eight triplets of spiny lizards (Sceloporus) using genomic sequence data. GBE 2013, 5, 2410–2419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulcahy, D.G.; Martínez-Gómez, J.E.; Aguirre-León, G.; Cervantes-Pasqualli, J.A.; Zug, G.R. Rediscovery of an endemic vertebrate from the remote Islas Revillagigedo in the Eastern Pacific Ocean: The Clarión Nightsnake lost and found. PLoS ONE 2014, 9, e97682. [Google Scholar] [CrossRef]
- Campillo-García, G.; Flores-Villela, O.; Butler, B.O.; Vinasco, J.A.V.; Corona, F.R. Hidden diversity within a polytypic species: The enigmatic Sceloporus torquatus Wiegmann, 1828 (Reptilia, Squamata, Phrynosomatidae). Vertebr. Zool. 2021, 71, 781–798. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W.; Zimmermann, N.E. Habitat Suitability and Distribution Models: With Applications in R, 1st ed.; Cambridge University Press: Cambridge, UK, 2017; p. 462. [Google Scholar] [CrossRef]
- Naimi, B.; Araújo, M.B. sdm: A reproducible and extensible R platform for species distribution modelling. Ecography 2016, 39, 368–375. [Google Scholar] [CrossRef] [Green Version]
- Di Cola, V.; Broennimann, O.; Petitpierre, B.; Breiner, F.T.; d’Amen, M.; Randin, C.; Engler, R.; Pottier, J.; Pio, D.; Dubuis, A.; et al. ecospat: An R package to support spatial analyses and modeling of species niches and distributions. Ecography 2017, 40, 774–787. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Cruz, J.A.; Arroyo-Cabrales, J.; Reynoso, V.H. Reconstructing the paleoenvironment of Loltún Cave, Yucatán, Mexico, with Pleistocene amphibians and reptiles and their paleobiogeographic implications. Rev. Mex. Cienc. Geol. 2016, 33, 342–354. [Google Scholar] [CrossRef]
- Cruz, J.A.; Alarcón, D.I.; Figueroa-Castro, D.M.; Castañeda-Posadas, C. Fossil pigmy rattlesnake inside the mandible of an American mastodon and use of fossil reptiles for the paleoclimatic reconstruction of a Pleistocene locality in Puebla, Mexico. Quat. Int. 2021, 574, 116–126. [Google Scholar] [CrossRef]
- Cruz, J.A.; Prado, J.L.; Arroyo-Cabrales, J. The mutual ecogeographical range and paleoclimatic reconstruction during the Late Pleistocene-Holocene in the Pampas (Argentina) using meso and microvertebrate fossils. Holocene 2021, 31, 983–992. [Google Scholar] [CrossRef]
- Hernández-Hernández, M.J.; Cruz, J.A.; Castañeda-Posadas, C. Paleoclimatic and vegetation reconstruction of the miocene southern Mexico using fossil flowers. J. South Am. Earth Sci. 2020, 104, 102827. [Google Scholar] [CrossRef]
- Medina-Castañeda, C.I.; Bravo-Cuevas, V.M.; Cruz, J.A. Turtles from the Late Pleistocene of Hidalgo and Puebla and their paleobiogeographic and paleoclimatic significance. Quat. Int. 2022, 634, 111–123. [Google Scholar] [CrossRef]
- QGIS Development Team. QGIS Geographic Information System. Open Source Geospatial Foundation. 2021. Available online: https://qgis.osgeo.org/ (accessed on 26 May 2021).
- Fick, S.E.; Hijmans, R.J. Worldclim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Hijmans, R. Raster: Geographic Data Analysis and Modeling. R Package Version 3.6-20. 2023. Available online: https://CRAN.R-project.org/package=raster (accessed on 3 September 2021).
- Keddy, P.A.; Laughlin, D.C. A Framework for Community Ecology: Species Pools, Filters and Traits, 1st ed.; Cambridge University Press: Cambridge, UK, 2021; p. 360. [Google Scholar] [CrossRef]
- Escalante, T.; Rodriguez-Tapia, G.; Morrone, J.J. Toward a biogeographic regionalization of the Nearctic region: Area nomenclature and digital map. Zootaxa 2020, 5027, 351–375. [Google Scholar] [CrossRef]
- Escalante, T.; Morrone, J.J. Evolutionary biogeography and the regionalization of the Neotropics: A perspective from mammals. Mastozool. Neotrop. 2020, 27, 5–14. [Google Scholar] [CrossRef]
- López-García, J.M.; Blain, H.A.; Cuenca-Bescós, G.; Ruiz-Zapata, M.B.; Dorado-Valiño, M.; Gil-García, M.J.; Valdeolmillos, A.; Ortega, A.I.; Carretero, J.M.; Arsuaga, J.L.; et al. Palaeoenvironmental and palaeoclimatic reconstruction of the latest Pleistocene of El Portalón site, Sierra de Atapuerca, northwestern Spain. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 292, 453–464. [Google Scholar] [CrossRef]
- López-García, J.M.; Blain, H.A.; Allué, E.; Bañuls, S.; Bargalló, A.; Martín, P.; Morales, J.I.; Pedro, M.; Rodríguez, A.; Solé, A.; et al. First fossil evidence of an “interglacial refugium” in the Pyrenean region. Naturwissenschaften 2010, 97, 753–761. [Google Scholar] [CrossRef] [PubMed]
- López-García, J.M.; Cuenca-Bescós, G.; Blain, H.A.; Álvarez-Lao, D.; Uzquiano, P.; Adán, G.; Arbizu, M.; Arsuaga, J.L. Palaeoenvironment and palaeoclimate of the Mousterian–Aurignacian transition in northern Iberia: The small-vertebrate assemblage from Cueva del Conde (Santo Adriano, Asturias). J. Hum. Evol. 2011, 61, 108–116. [Google Scholar] [CrossRef] [PubMed]
- López-García, J.M.; Blain, H.A.; Cuenca-Bescós, G.; Alonso, C.; Alonso, S.; Vaquero, M. Small vertebrates (Amphibia, Squamata, Mammalia) from the late Pleistocene-Holocene of the Valdavara-1 cave (Galicia, northwestern Spain). Geobios 2011, 44, 253–269. [Google Scholar] [CrossRef]
- López-García, J.M.; Blain, H.A.; Bennàsar, M.; Euba, I.; Bañuls, S.; Bischoff, J.; López-Ortega, E.; Saladié, P.; Uzquiano, P.; Vallverdú, J. A multiproxy reconstruction of the palaeoenvironment and palaeoclimate of the Late Pleistocene in northeastern Iberia: Cova dels Xaragalls, Vimbodí-Poblet, Paratge Natural de Poblet, Catalonia. Boreas 2012, 41, 235–249. [Google Scholar] [CrossRef]
- López-García, J.M.; Blain, H.A.; Sanz, M.; Daura, J. A coastal reservoir of terrestrial resources for neanderthal populations in north-eastern Iberia: Palaeoenvironmental data inferred from the small-vertebrate assemblage of Cova del Gegant, Sitges, Barcelona. J. Quat. Sci. 2012, 27, 105–113. [Google Scholar] [CrossRef]
- López-García, J.M.; Blain, H.A.; Bennàsar, M.; Sanz, M.; Daura, J. Heinrich event 4 characterized by terrestrial proxies in southwestern Europe. Clim. Past 2013, 9, 1053–1064. [Google Scholar] [CrossRef] [Green Version]
- López-García, J.M.; Blain, H.A.; Julià, R.; Maroto, J. Environment and climate during MIS 7 and their implications for the late Middle Pleistocene hominins: The contribution of Mollet cave, Serinyà, Girona, northeastern Iberian Peninsula. Quat. Int. 2014, 337, 4–10. [Google Scholar] [CrossRef] [Green Version]
- López-García, J.M.; Blain, H.A.; Bennàsar, M.; Fernández-García, M. Environmental and climatic context of Neanderthal occupation in southwestern Europe during MIS3 inferred from the small-vertebrate assemblages. Quat. Int. 2014, 326, 319–328. [Google Scholar] [CrossRef]
- López-García, J.M.; Fernández-García, M.; Blain, H.A.; Sanz, M.; Daura, J. MIS 5 environmental and climatic reconstruction in northeastern Iberia using the small-vertebrate assemblage from the terrestrial sequence of Cova del Rinoceront (Castelldefels, Barcelona). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 451, 13–22. [Google Scholar] [CrossRef]
- López-García, J.M.; Blain, H.A.; Sánchez-Bandera, C.; Cohen, J.; Lebreton, L.; Montuire, S.; Stewart, J.R.; Desclaux, E. Multi-method approach using small vertebrate assemblages to reconstruct the Marine Isotope Stage 6 climate and environment of the Lazaret cave sequence (Maritime Alps, Nice, France). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2021, 577, 110529. [Google Scholar] [CrossRef]
- Bañuls-Cardona, S.; López-García, J.M.; Blain, H.A.; Salomó, A.C. Climate and landscape during the Last Glacial Maximum in southwestern Iberia: The small-vertebrate association from the Sala de las Chimeneas, Maltravieso, Extremadura. C. R. Palevol. 2012, 11, 31–40. [Google Scholar] [CrossRef]
- Rey-Rodríguez, I.; López-García, J.M.; Bennasar, M.; Bañuls-Cardona, S.; Blain, H.A.; Blanco-Lapaz, Á.; Rodríguez-Álvarez, X.P.; de Lombera-Hermida, A.; Díaz-Rodríguez, M.; Ameijenda-Iglesias, A.; et al. Last Neanderthals and first Anatomically Modern Humans in the NW Iberian Peninsula: Climatic and environmental conditions inferred from the Cova Eirós small-vertebrate assemblage during MIS 3. Quat. Sci. Rev. 2016, 151, 185–197. [Google Scholar] [CrossRef]
- Steadman, D.W.; Arroyo-Cabrales, J.; Johnson, E.; Guzman, A.F. New information on the late Pleistocene birds from San Josecito cave, Nuevo León, Mexico. Condor 1994, 96, 577–589. [Google Scholar] [CrossRef] [Green Version]
- Mead, J.I.; Arroyo-Cabrales, J.; Johnson, E. Pleistocene lizards (Reptilia: Squamata) from San Josecito Cave, Nuevo León, México. Copeia 1999, 1999, 163–173. [Google Scholar] [CrossRef]
- Van Devender, T.R. Late Quaternary vegetation and climate of the Chihuahuan Desert, United States and Mexico. In Packrat Middens: The Last 40,000 Years of Biotic Change, 1st ed.; Betancourt, J.L., Van Devender, T.R., Martin, P.S., Eds.; Arizona University Press: Tucson, AZ, USA, 1990; pp. 104–133. [Google Scholar]
- Caballero, M.; Lozano-García, S.; Ortega-Guerrero, B.; Correa-Metrio, A. Quantitative estimates of orbital and millennial scale climatic variability in central Mexico during the last ∼40,000 years. Quat. Sci. Rev. 2019, 205, 62–75. [Google Scholar] [CrossRef]
- Lozano-García, S.; Torres-Rodríguez, E.; Figueroa-Rangel, B.; Caballero, M.; Sosa-Nájera, S.; Ortega-Guerrero, B.; Acosta-Noriega, C. Vegetation history of a Mexican Neotropical basin from the late MIS 6 to early MIS 3: The pollen record of Lake Chalco. Quat. Sci. Rev. 2022, 297, 107830. [Google Scholar] [CrossRef]
- Wright, K.T. Reconstructing Rainfall Variability in Northeast Mexico in the Late-Pleistocene and Holocene Using Multiple Speleothem Geochemical Proxies. Ph.D. Thesis, University of California, Irvine, CA, USA, 2021. [Google Scholar]
- Pearce-Higgins, J.W.; Eglington, S.M.; Martay, B.; Chamberlain, D.E. Drivers of climate change impacts on bird communities. J. Anim. Ecol. 2015, 84, 943–954. [Google Scholar] [CrossRef]
- Moretti, J.A.; Johnson, E. Small rails from the late Quaternary of the Southern High Plains and their paleoenvironmental context. Ibis 2023, in press. [Google Scholar] [CrossRef]
- Oswald, J.A.; Steadman, D.W. The changing diversity and distribution of dry forest passerine birds in northwestern Peru since the last ice age. Auk 2015, 132, 836–862. [Google Scholar] [CrossRef]
- Steadman, D.W.; Albury, N.A.; Kakuk, B.; Mead, J.I.; Soto-Centeno, J.A.; Singleton, H.M.; Franklin, J. Vertebrate community on an ice-age Caribbean Island. Proc. Natl. Acad. Sci. USA 2015, 112, E5963–E5971. [Google Scholar] [CrossRef] [PubMed]
- Steadman, D.W.; Franklin, J. Origin, paleoecology, and extirpation of bluebirds and crossbills in the Bahamas across the last glacial–interglacial transition. Proc. Natl. Acad. Sci. USA 2017, 114, 9924–9929. [Google Scholar] [CrossRef] [PubMed]
- Tomek, T.; Bochenski, Z.M.; Socha, P.; Stefaniak, K. Continous 300,000-year fossil record changes in the ornithofauna of Bisnik Cave, Poland. Palaeontol. Electron. 2012, 15, 2A–20A. [Google Scholar] [CrossRef]
- Carrera, L.; Scarponi, D.; Martini, F.; Sarti, L.; Pavia, M. Mid-Late Pleistocene Neanderthal landscapes in southern Italy: Palaeocological contributions of the avian assemblage from Grotta del Cavallo, Apulia, Southern Italy. Palaeogeog. Palaeoclimatol. Palaeoecol. 2021, 567, 110256. [Google Scholar] [CrossRef]
- Núñez-Lahuerta, C.; Galán, J.; Cuenca-Bescós, G.; García-Medrano, P.; Cáceres, I. A bird asseblage across the MIS9/8 boundary: The Middle Pleistocene of Galería (Atapuerca). Quat. Sci. Rev. 2022, 293, 107708. [Google Scholar] [CrossRef]
- Pavia, M.; Val, A.; Carrera, L.; Steininger, C.M. Fossil birds from Cooper’s D aid in reconstructing the Early Pleistocene palaeoenvironment in the Cradle of Humankind (Gauteng, South Africa). J. Hum. Evol. 2022, 167, 103185. [Google Scholar] [CrossRef] [PubMed]
- Brattstrom, B. The climate of the past. Eng. Sci. 1956, 19, 22–23. [Google Scholar]
- Vitt, L.J.; Caldwell, J.P. Herpetology: An Introductory Biology of Amphibians and Reptiles, 4th ed.; Academic Press: San Jose, CA, USA, 2014; p. 776. [Google Scholar] [CrossRef]
- Böhm, M.; Cook, D.; Ma, H.; Davidson, A.D.; García, A.; Tapley, B.; Pearce-Kelly, P.; Carr, J. Hot and bothered: Using trait-based approaches to assess climate change vulnerability in reptiles. Biol. Conserv. 2016, 204, 32–41. [Google Scholar] [CrossRef]
- Diele-Viegas, L.M.; Figueroa, R.T.; Vilela, B.; Rocha, C.F.D. Are reptiles toast? A worldwide evaluation of Lepidosauria vulnerability to climate change. Clim. Change 2020, 159, 581–599. [Google Scholar] [CrossRef]
- Porch, N. Climate space, bioclimatic envelopes and coexistence methods for the reconstruction of past climates: A method using Australian beetles and significance for Quaternary reconstruction. Quat. Sci. Rev. 2010, 29, 633–647. [Google Scholar] [CrossRef]
- Araújo, M.B.; Nogués-Bravo, D.; Diniz-Filho, J.A.F.; Haywood, A.M.; Valdes, P.J.; Rahbek, C. Quaternary climate changes explain diversity among reptiles and amphibians. Ecography 2008, 31, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Hadly, E.A.; Spaeth, P.A.; Li, C. Niche conservatism above the species level. Proc. Natl. Acad. Sci. USA 2009, 106, 19707–19714. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.R.; Polly, P.D. A reevaluation of the Harrodsburg Crevice fauna (Late Pleistocene of Indiana, USA) and the climatic implications of its mammals. J. Vertebr. Paleontol. 2013, 33, 410–420. [Google Scholar] [CrossRef]
- Polly, P.D.; Eronen, J.T. Mammal associations in the Pleistocene of Britain: Implications of ecological niche modelling and a method for reconstructing palaeoclimate. In The Ancient Human Occupation of Britain, 1st ed.; Ashton, N., Lewis, S., Stringer, C., Eds.; Elsevier: London, UK, 2011; pp. 279–304. [Google Scholar] [CrossRef]
- Villa, A.; Blain, H.A.; Delfino, M. The Early Pleistocene herpetofauna of Rivoli Veronese (Northern Italy) as evidence for humid and forested glacial phases in the Gelasian of Southern Alps. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 490, 393–403. [Google Scholar] [CrossRef]
- Villa, A.; Blain, H.A.; van den Hoek Ostende, L.W.; Delfino, M. Fossil amphibians and reptiles from Tegelen (Province of Limburg) and the early Pleistocene palaeoclimate of The Netherlands. Quat. Sci. Rev. 2018, 187, 203–219. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Orden | Familia | Especie |
|---|---|---|---|
| Birds | Accipitriformes | Accipitridae | Aquila chrysaetos |
| Galliformes | Phasianidae | Cyrtonyx montezumae | |
| Gruiformes | Rallidae | Rallus limicola | |
| Charadriiformes | Charadriidae | Pluvialis cf. P. squatarola | |
| Strigiformes | Strigidae | Megascops asio/M. kennicottii | |
| Asio otus | |||
| Ciccaba virgata | |||
| Passeriformes | Corvidae | Gymnorhinus cyanocephalus | |
| Corvus corax | |||
| Icteridae | Sturnella sp. | ||
| Reptiles | Squamata | Anguidae | Barisia ciliaris |
| Barisia imbricata | |||
| Phrynosomatidae | Phrynosoma modestum | ||
| Phrynosoma orbiculare | |||
| Sceloporus spp. | |||
| Colubridae | Tantilla sp. | ||
| Heterodon cf. H. simus | |||
| Hypsiglena | |||
| Storeria sp. | |||
| Viperidae | Crotalus spp. | ||
| Mammals | Soricomorpha | Soricidae | Sorex altoensis |
| Chiroptera | Phyllostomidae | Desmodus stocki | |
| Leptonycteris nivalis | |||
| Vespertilionidae | Myotis californicus | ||
| Myotis thysanodes | |||
| Rodentia | Cricetidae | Microtus mexicanus | |
| Neotoma albigula | |||
| Neotoma albigula | |||
| Peromyscus difficilis | |||
| Peromyscus labecula | |||
| Peromyscus levipes | |||
| Reithrodontomy megalotis | |||
| Lagomorpha | Leporidae | Sylvilagus floridanus |
| MAT °C (Average ± SD) | AP mm (Average ± SD) | |
|---|---|---|
| Current San Josecito Cave | 17.6 ± 1.69 | 774.68 ± 259.85 |
| Difference | 0 | 0 |
| All vertebrates | 20.24 ± 0.32 | 695.5 ± 80.95 |
| Difference | +2.64 | −79.175 |
| Mammals and reptiles | 20.55 ± 0.48 | 668.1 ± 74.18 |
| Difference | +2.95 | −106.58 |
| Mammals | 20.14 ± 1.22 | 617.7 ± 147.66 |
| Difference | +2.54 | −156.98 |
| Reptiles | 17.25 ± 3.27 | 990.5 ± 389.77 |
| Difference | −0.35 | +215.82 |
| Birds | 17.1 ± 5.1 | 664.2 ± 380.87 |
| Difference | −0.5 | −110.48 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruz, J.A.; Velasco, J.A.; Arroyo-Cabrales, J.; Johnson, E. Paleoclimatic Reconstruction Based on the Late Pleistocene San Josecito Cave Stratum 720 Fauna Using Fossil Mammals, Reptiles, and Birds. Diversity 2023, 15, 881. https://doi.org/10.3390/d15070881
Cruz JA, Velasco JA, Arroyo-Cabrales J, Johnson E. Paleoclimatic Reconstruction Based on the Late Pleistocene San Josecito Cave Stratum 720 Fauna Using Fossil Mammals, Reptiles, and Birds. Diversity. 2023; 15(7):881. https://doi.org/10.3390/d15070881
Chicago/Turabian StyleCruz, J. Alberto, Julián A. Velasco, Joaquín Arroyo-Cabrales, and Eileen Johnson. 2023. "Paleoclimatic Reconstruction Based on the Late Pleistocene San Josecito Cave Stratum 720 Fauna Using Fossil Mammals, Reptiles, and Birds" Diversity 15, no. 7: 881. https://doi.org/10.3390/d15070881
APA StyleCruz, J. A., Velasco, J. A., Arroyo-Cabrales, J., & Johnson, E. (2023). Paleoclimatic Reconstruction Based on the Late Pleistocene San Josecito Cave Stratum 720 Fauna Using Fossil Mammals, Reptiles, and Birds. Diversity, 15(7), 881. https://doi.org/10.3390/d15070881