Abstract
Background: This work studied the occurrence, density, and diversity of yeast species associated with the leaves and fruits of Sabicea brasiliensis and Anacardium humile. Methods: Fragments of leaves and fruits were subjected to maceration and placed under agitation in peptone water. One hundred µL aliquots of each of the decimal dilutions were plated on YM chloramphenicol agar medium. The different morphotypes were identified through sequence analysis of the 26S rDNA. Results: A total of 83 yeast isolates were recovered from the organs sampled, 40 from fruits and 19 from the leaves of S. brasiliensis, while 14 were recovered from fruits and 10 from the leaves of A. humile. The isolates were identified as belonging to two phyla, six classes, eight orders, and eight families. Most isolates, at 71 (84.3%), belong to Ascomycota, with Saccharomycetes dominant in fruits and Dothideomycetes in leaves. The remaining 12 (15.7%) isolates belong to the phylum Basidiomycota. The most frequent genera were Aureobasidium, Candida, Rhodotorula, and Wickerhamiella. The Wickerhamiella isolates made possible the subsequent description of a new species within this genus. Conclusions: Our data reveal the yeast species that inhabit different Cerrado plant organs and indicate that these hosts can be considered an important habitat for yeast diversity, including new and/or previously unreported species in this biome.
1. Introduction
The phyllosphere, a region that encompasses all the aerial parts of plants [1], is colonized by a diverse pool of species from numerous microbial groups, which include archaea, bacteria, fungi, and viruses [2,3,4,5,6]. In regard to the fungal species present in the phyllosphere, yeasts are the main group of this kingdom, while filamentous fungi usually occur as dormant spores [2].
The yeast communities associated with the phylloplane of plant species from various natural and agricultural ecosystems have been intensively studied throughout the world [7,8,9,10] and their ecological roles, occurrence, and species diversity have been widely discussed [10,11]. For example, it is known that the taxonomic structure, relative abundance, and occurrence of yeast species present in the phylloplane depend on a number of factors, such as host taxonomic identity [12], nutrient availability [13], and seasonality [14,15]. In Brazilian biomes, there are still few studies involving the characterization of the yeast community associated with the phylloplane, mainly of plant species endemic to the Brazilian neotropical savanna (Cerrado).
The Brazilian neotropical savanna is the second largest biome in the country, comprising an area of more than 2.0 million km2 [16] and is considered a global hotspot due to its biological richness, high degree of endemicity, and high anthropic pressure to which it has been suffering [17]. In this biome, communities of yeast and filamentous fungi have been reported as epiphytes or endophytes associated with the leaves and fruits of some native plant species [18,19,20]. However, even with the effort and work of the authors mentioned above, data on the occurrence and distribution of yeasts in native plants of the Cerrado are still very limited, especially considering the great plant diversity of this biome that can host these microorganisms.
Anacardium humile and Sabicea brasiliensis are species with high occurrence in Cerrado regions of Brazil and belong to the families Anacardiaceae and Rubiaceae, respectively (https://floradobrasil.jbrj.gov.br/reflora/ accessed on 28 November 2022). Both have nutritional, medicinal and economic importance. Primarily used as food, they are also employed in popular medicine to treat inflammation, infections and with antioxidant properties [21,22,23,24,25]. Due to the fact that these Cerrado species show antagonistic (antimicrobial activity), biological and socioeconomic interests, it becomes necessary to also generate biological information about their mycobiota, in an attempt at an adequate biotechnological exploration through the future use of the microorganisms that these plants host in their tissues. Despite the widespread popular use of plants from these genera, only Anacardium humile has had part of its mycobiota studied by our team using samples from another region of the Brazilian Cerrado [19].
Based on this, the occurrence and diversity of yeasts in leaves and fruits of two species of plants native to the Brazilian neotropical savanna that occur in the Midwest Region, Federal District, Brazil, were evaluated through cultivation techniques and molecular identification. The objectives were (1) to isolate, identify and evaluate the population density of cultivable yeasts and (2) to evaluate the diversity and distribution of yeast species in the leaves and fruits of the host plant, noting whether there are yeast species in both the yeast communities of the evaluated host plants and in different organs of the same host (leaves and fruits).
2. Materials and Methods
2.1. Sample Processing
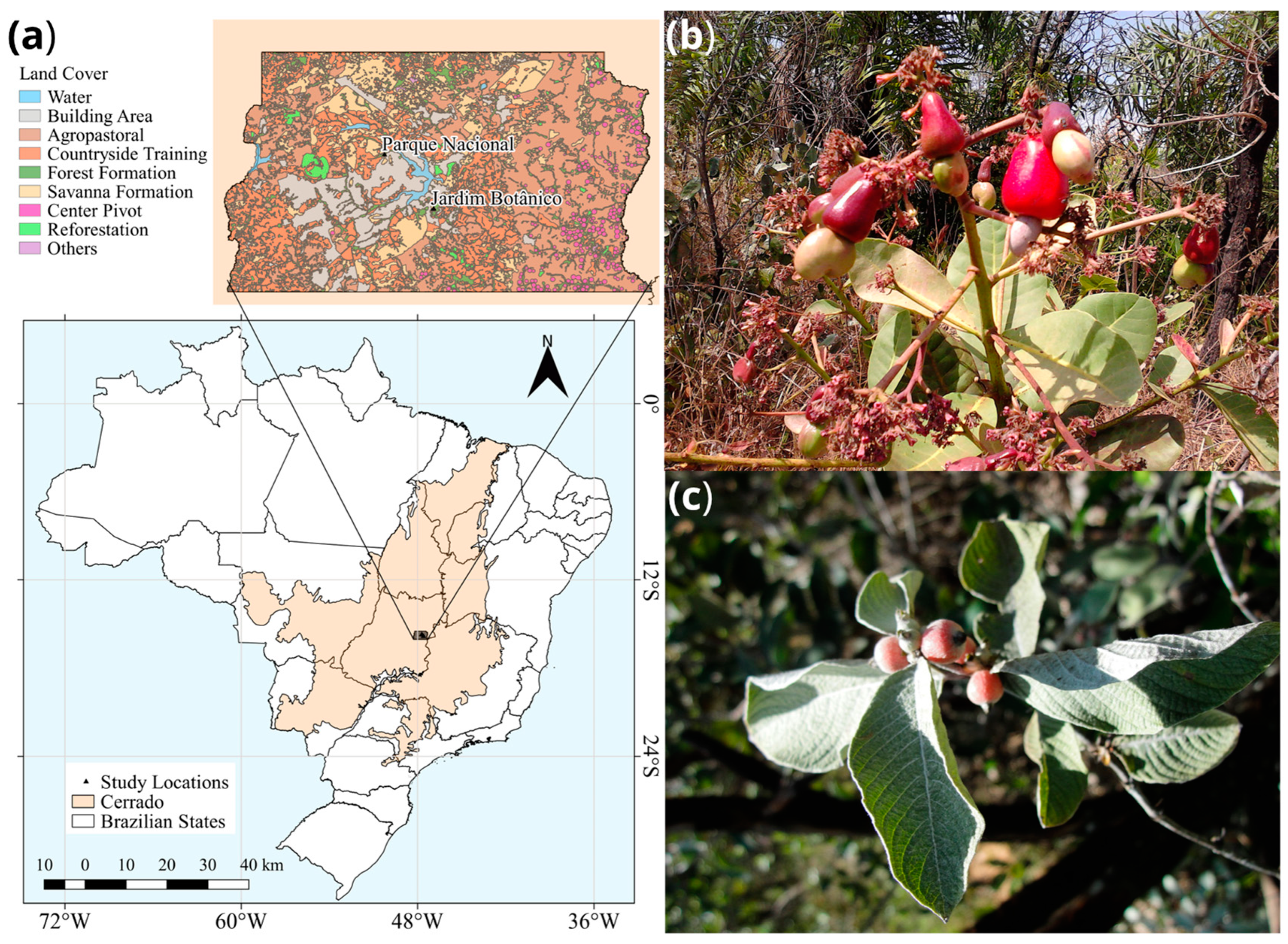
Two plant species native to the Cerrado were collected in the Ecological Reserves of the Federal District, Brazil: Sabicea brasiliensis (Rubiaceae), geographic coordinates 15°54′0″ S 47°51′30″ W and Anacardium humile (Anacardiaceae), geographic coordinates S 15°47′ W 47°56′ (Figure 1). Mature healthy leaves and fruits were collected aseptically and processed within 48 h.
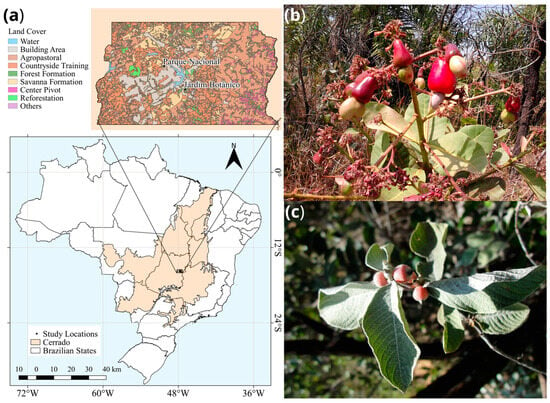
Figure 1.
Origin of samples and characteristics of the study site in the Midwest Region, Federal District, Brazil, and host species analyzed. Panel (a) shows the area of the Cerrado biome where leaves and fruits were sampled. In (b) the leaves and fruits of Anacardium humile are shown, while in (c) the leaves and fruits of Sabicea brasiliensis are shown. Panel (a) was created in QGIS software version 3.26.2.
2.2. Isolation of Yeasts
Samples of two species of plants were collected, comprising three different trees for each plant species and three different samples for each tree of the same plant species. The samples were cut into small pieces, ground and placed in Erlenmeyer flasks containing 0.8% peptone aqueous solution (w/v) (Bio-Rad, Hercules, CA, USA) in a 9:1 ratio of peptone solution volume to sample volume. The samples were agitated on a rotary shaker at 150 rpm for 25 min. Later, each peptone solution was diluted by factors of 10−1 up to 10−3 in culture tubes containing 9 mL of peptone solution. Aliquots of 100 μL from these serial dilutions were pipetted onto and spread on Petri dishes; three Petri dishes were used per dilution. The yeast was isolated on Petri dishes with YM agar (0.3% yeast extract, 0.3% malt extract, 0.5% peptone, 1% glucose, 2% agar) containing 100 mg mL−1 chloramphenicol. The Petri dishes were incubated in a growth chamber at 28 °C for a period of 5 to 7 days, and, after incubation, colonies were counted and expressed as colony-forming units (CFU). Representative colonies were selected according to their cultural characteristics (colony color, size, surface, texture, border type and growth rate) and picked for purification 5 and 7 days after processing the samples; these time points were chosen to allow for the growth of the yeast colonies and to avoid the growth of filamentous fungi on the plates. Each isolate was purified by subsequent streaking on the same medium with chloramphenicol (100 mg mL−1) and microscope observation. Pure yeast cultures were catalogued and preserved at −80 °C in cryovials containing 700 μL of YM liquid culture medium and 300 μL of 50% glycerol (v/v).
2.3. Identification of Isolates
DNA was extracted from yeast culture samples as described by Makimura et al. [26], with some modifications. The isolates were grown in 9 mL of YM liquid culture medium (pH 5.6) for 48 h at 28 °C without shaking, after which they were centrifuged for 5 min at 12,000 rpm. Cells were lysed by agitation at 2500 rpm with 0.1 g of glass beads in 200 μL of extraction buffer for 1.0 min. The culture was centrifuged for 10 min at 12,000 rpm after adding 200 μL of phenol:chloroform:isoamyl alcohol at 25:24:1 (v/v). DNA in the supernatant was precipitated with isopropyl alcohol for 1 h at −20 °C. After that, the DNA samples were washed and centrifuged twice with each of 70% and 100% ethanol. The precipitated DNA was then suspended in 30 μL of Milli-Q H2O and stored at −20 °C until further use.
The yeast isolates were grouped according to cultural morphological aspects, production of amyloid compounds, assimilation of carbon and nitrogen sources, growth at different temperatures, and tolerance to sodium chloride (NaCl) [27] and also grouped utilizing the pattern of bands generated by MSP-PCR with the M13 primer (data not shown) [28]. A total of 83 yeast isolates, representative isolates for each group, were selected to be identified molecularly. Fragments of the D1/D2 domain of the rDNA large subunit (LSU) were amplified as described by Kurtzman and Robnett [29] using the primers NL1 (5′-GCATATCAATAAGCG GAG GAAAAG-3′) and NL4 (5′-GGTCCGTGTTTCAAGACGG-3′) [15]. The resulting PCR products were purified using a USB® ExoSAP-IT® Kit (Thermo Fisher Scientific, Waltham, Massachusetts, USA) according to the manufacturer’s instructions, and the purified PCR products were then sequenced at the Catholic University of Brazil using the Sanger method. The sequences that were obtained were compared with sequences in the GenBank database (http://www.ncbi.nlm.nih.gov/) using Basic Local Alignment Search Tool (BLAST) (https://blast.ncbi.nlm.nih.gov/Blast.cgi accessed on 18 October 2022). The sequences were deposited in Genbank. We considered sequence homology of more than 97% to classify isolates with species and homology less than 97% to classify isolates by genus.
2.4. Relative Abundance and Diversity Metric
Relative abundance analyzes and yeast community diversity indices were performed using the R software version 4.1.3, and the graphs were generated using the “ggplot” package, except the visualization of the taxonomic network of species occurrence and sharing, which was performed in CYTOSCAPE version 3.9.1 [30].
The relative abundance and abundance curve were calculated comparing the ratio between the absolute frequency of a given species over the sum of the absolute frequency of all species. To observe the sharing of species between host plants and the type of organ analyzed, the data were organized in columns, the first column referring to the species found and the second the identity of the host plant and type of organ analyzed. The file was uploaded to CYTOSCAPE, where the first column was named “Target node” and column two as “Source node”, and the species sharing network was visualized.
The alpha and beta diversity metrics were calculated using the “vegan”, “betapart” and “BiodiversityR” packages. To assess alpha diversity, the Pielou evenness [31] and the Hill series (q) were used [32,33]. Beta diversity (similarities and dissimilarities) between communities was demonstrated using the Jaccard distance, and Sorensen beta diversity [34].
3. Results
3.1. Isolation and Population Density of Yeasts in Sabicea brasiliensis and Anacardium humile
A total of 83 yeast isolates were recovered from the two types of organs sampled from the two plant hosts studied, 40 from fruits and 19 from leaves of the host S. brasiliensis, and 14 from fruits and 10 from leaves of the host A. humile (Table 1).

Table 1.
Isolated yeasts, organ of isolation and distribution among hosts.
The count of CFU x g−1 of fruits/yeast leaves showed variation between the species of host plants and type of organ analyzed, ranging from 3.1 × 101 to 2.2 × 102 CFU x g−1 for the fruits and 1.7 × 101 to 6.0 × 101 CFU.g−1 for the leaves (Table 2). When the organ analyzed was the fruit, S. brasiliensis presented a higher average of colony-forming units per gram of fruit (2.6 × 102 CFU.g−1), about ten times more than A. humile (3.1 × 101 CFU.g−1). As for the leaf tissue, A. humile presented a higher average of CFU, 6.0 × 101 CFU.g−1, when compared with S. brasiliensis, which presented an average of 1.7 × 101 CFU.g−1 of the leaf.
Table 2.
Density of yeasts per gram of tissue (UFC.g organ−1) in leaves and fruits of Cerrado native plants: S. brasiliensis and A. humile.
3.2. Identification and Composition of Yeasts in the Two Hosts
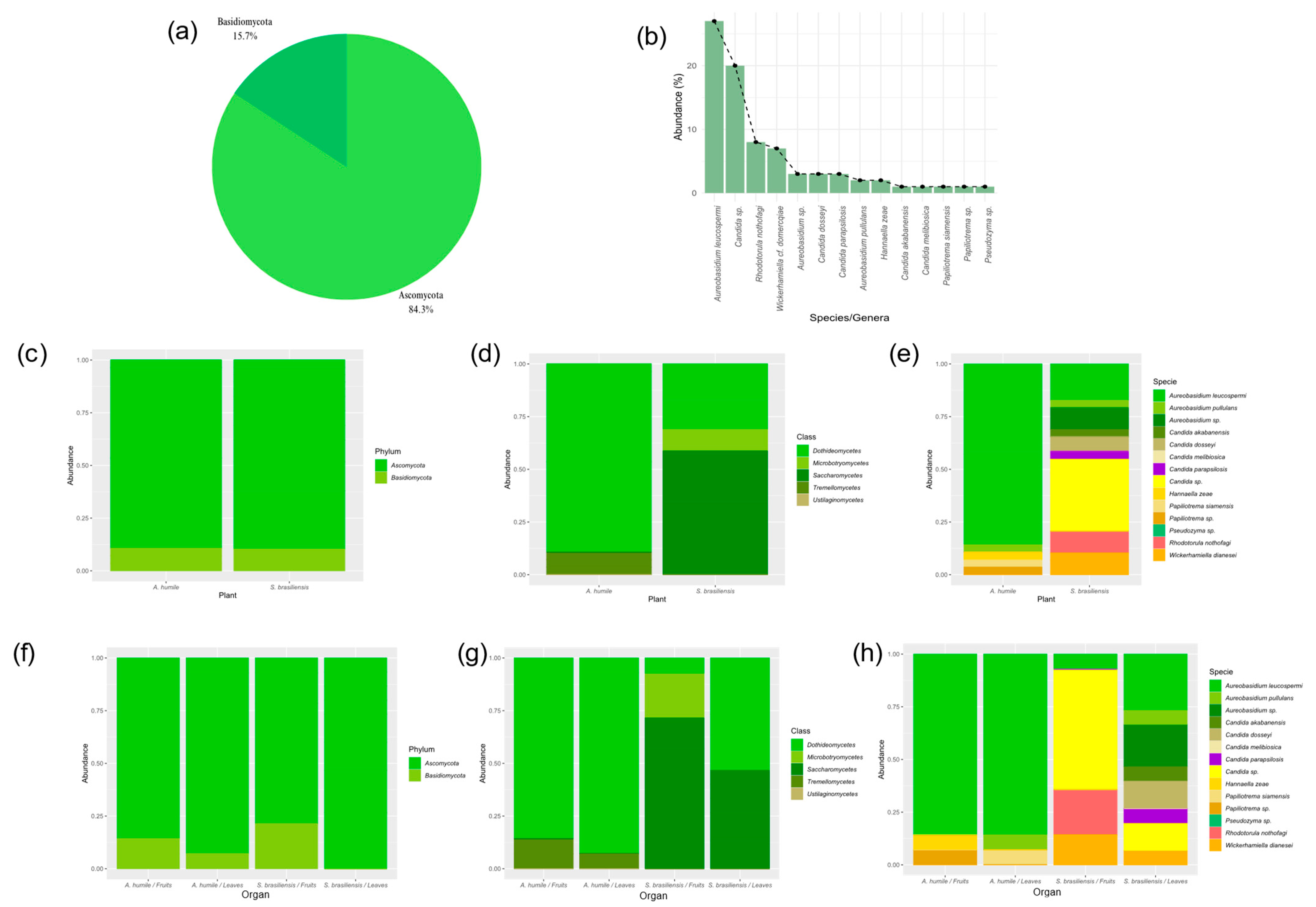
Based on the sequencing of the LSU D1/D2 region, the isolates were classified into two phyla, five classes and seven genera. Of the 83 isolates, 71 belong to the phylum Ascomycota, while 12 belong to the phylum Basidiomycota (Figure 2a). The most abundant genera were Aureobasidium, Candida, Rhodotorula, and Wickerhamiella, with Candida showing the greatest diversity of species (Figure 2b). There were differences in the abundance of genera and in the diversity of isolated species among the hosts (Figure 2c). In both hosts and in the organs analyzed there was a prevalence of the phylum Ascomycota (Figure 2c), with most isolates belonging to the classes Saccharomycetes and Dothideomycetes (Figure 2d). Six genera and nine species were isolated from S. brasiliensis and three species belonging to three genera were isolated from A. humile (Figure 2e). Isolates belonging to the phylum Basidiomycota were recovered from both hosts, but with lower frequency and abundance (Figure 2f). In the fruits of S. brasiliensis, there was a predominance of species belonging to the genera Candida and Wickerhamiella, while in the leaves there was a predominance of Aureobasidium and Wickerhamiella (Figure 2g,h). As for the leaves and fruits of A. humile, there was a predominance of the genus Aureobasidium.
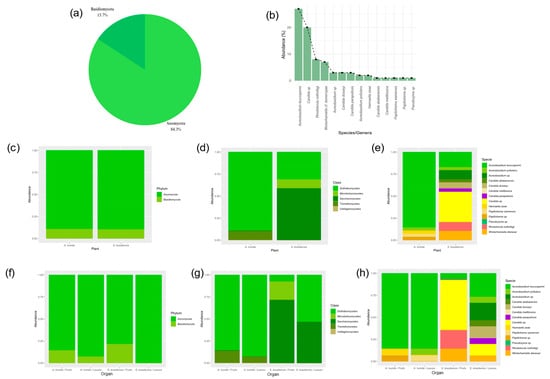
Figure 2.
Taxonomic composition and relative abundance of yeast community in leaves and fruits of Anacardium humile and Sabicea brasiliensis. (a) Percentage of yeasts belonging to the phyla Ascomycota and Basidiomycota of the total yeast community. (b) Relative abundance curve of the total yeast community. (c) Relative abundance of the phyla Ascomycota and Basidiomycota associated with the two host species. (d) Composition and relative abundance of the yeast community at the taxonomic level of the class. (e) Relative abundance of the yeast community associated with the two host species. (f) Relative abundance of the phyla Ascomycota and Basidiomycota associated with leaves and fruits of both host species. (g) Relative abundance of the yeast community, at the taxonomic level of the class, associated with the leaves and fruits of both host species. (h) Relative abundance of the yeast community associated with leaves and fruits of both host species. Figure created in software R version 4.1.1.
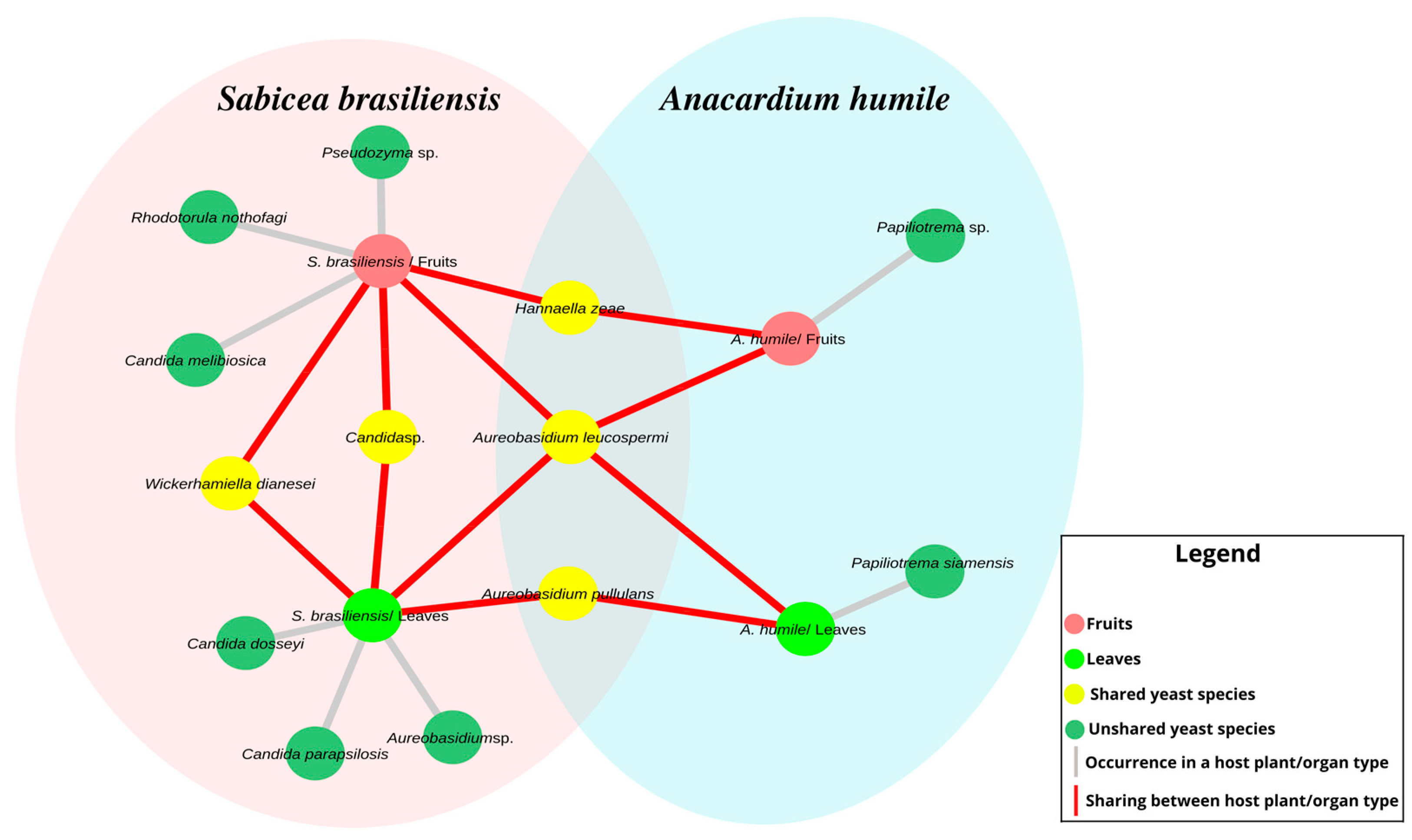
In addition to the differences in the relative abundance of yeasts between the hosts, differences were observed between the occurrence and sharing of species between leaves and fruits for both plant species (Figure 3). The species of Candida dosseyi, C. parapsilopsis and Aureobasidium sp. colonize only the leaves of the host S. brasiliensis, while the Pseudozyma genus C. melibiosica and R. nothofagi species are exclusive to the fruits. The fruits and leaves of A. humile share only isolates of Aureobasidium and Papiliotrema. The only species shared between the two hosts were A. pullulans, A. leucospermi and H. zeae, with A. leucospermi found on the leaves and fruits of both hosts.
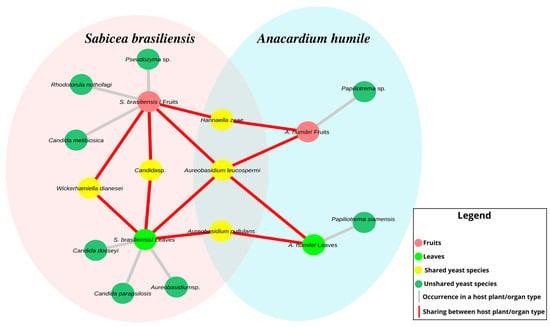
Figure 3.
Architecture of the distribution network of yeast between the two host plant species and the organ types analyzed. The pink ellipse represents the host S. brasiliensis, while the green ellipse represents the host A. humile. Pink nodes represent fruits and lime green nodes represent leaves. The green nodes represent the yeasts species not shared among the host plants, while those in yellow show the shared yeasts species. Gray edges indicate the occurrence of the species within a host type and organ type, while those in red show species sharing. The relative abundance of each species/genus was not considered. Figure created in CYTOSCAPE version 3.9.1.
3.3. Alpha and Beta Diversity
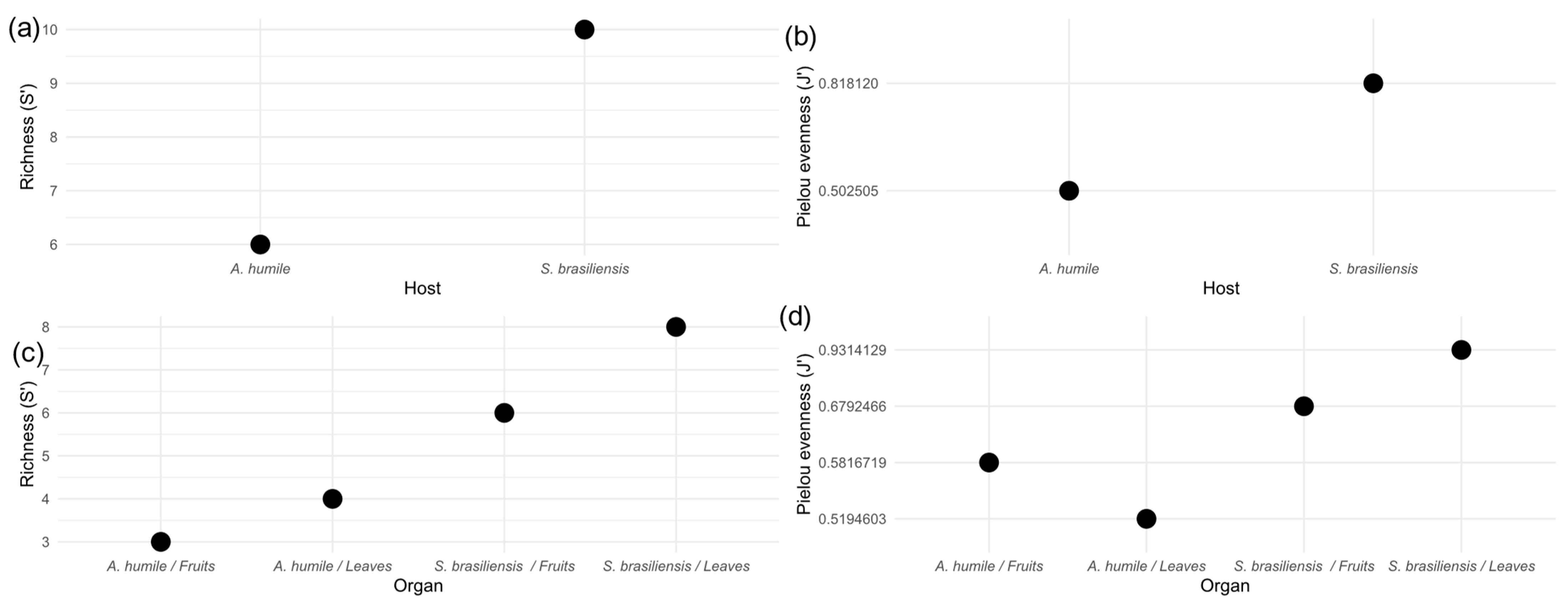
The cultured yeast community associated with the two host plants analyzed here showed differences between the alpha diversity metrics. The Species Richness index (S’) shows that the community present in S. brasiliensis is richer in yeast species than the community associated with A. humile (Figure 4a). When the community was evaluated between the different organs of the two plant host species, the highest values of species richness were observed in the fruits and leaves of S. brasiliensis (Figure 4c). When the yeast communities were evaluated for equity, the Pielou evenness index showed that the community in S. brasiliensis has the highest equability value (Figure 4b), with the highest value observed in the community associated with leaves of this species (Figure 4d). The lowest evenness value was observed in A. humile leaves. This finding suggests that the species of cultivable yeasts present in the leaves and fruits of S. brasiliensis have closer abundance values, while in A. humile the community in these organs is composed of a dominant group.
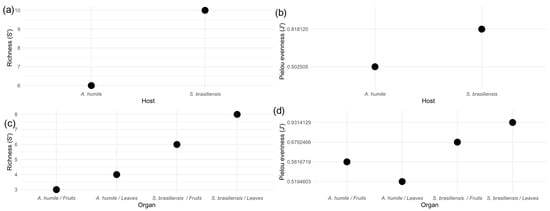
Figure 4.
Alpha diversity indices of the community of yeasts associated with aerial parts (leaves and fruits) of Anacardium humile and Sabicea brasiliensis. (a,c) Species richness (S’). (b,d) Pielou evenness. Figure created in software R version 4.1.1.
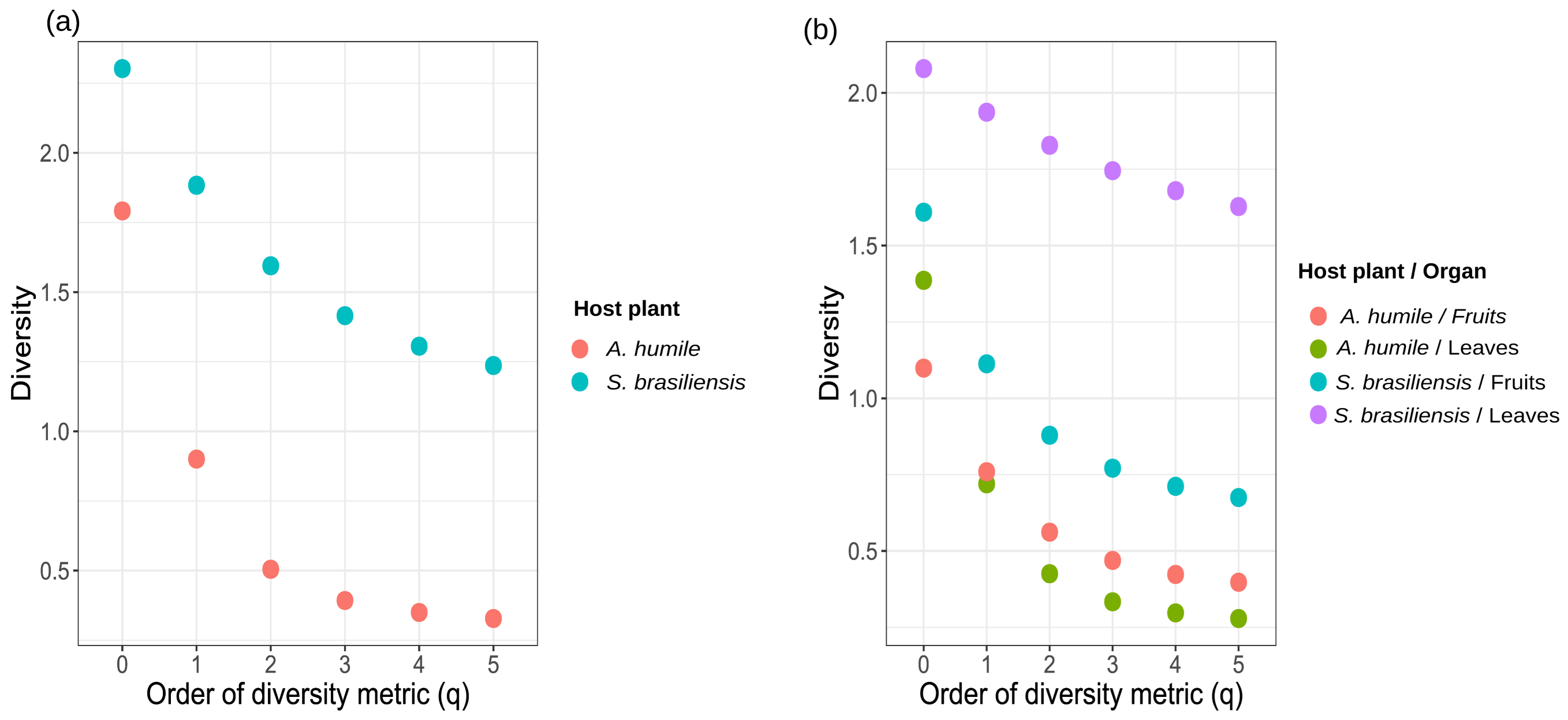
As for the indices presented by the Hill series (q), S. brasiliensis showed higher values for all of them, including the indices that give more weight to the most abundant species (q = 2, 3, 4, and 5) (Figure 5a). Numerically, the values obtained for the different indices in S. brasiliensis were q = 0, S’ = 2.30; q = 1, H’ = 1.88; q = 2, 1/D’ = 1.59; q = 3, Gini–Simpson index = 1.41; q = 4, Tsallis index (HCDT) = 1.302; q = 5, Renyi index = 1.23. The values obtained for A. humile were q = 0, S = 1.79; q = 1, H’ = 0.90; q = 2, 1/D’ = 0.50; q = 3, Gini–Simpson index = 0.39; q = 4, Tsallis index (HCDT) = 0.34; q = 5, Renyi index = 0.32. For the different organs analyzed from both hosts, the highest values of the Hill series were found in leaves of S. brasiliensis (q = 0, S = 2.07; q = 1, H’ = 1.93; q = 2, 1/D’ = 1.82; q = 3, Gini–Simpson index = 1.74; q = 4, Tsallis index HCDT = 1.67; q = 5, Renyi index = 1.62) while the lowest values were observed in the leaves of A. humile (q = 0, S = 1.386294; q = 1, H’ = 0.72; q = 2, 1/D’ = 0.42; q = 3, Gini–Simpson index = 0.33; q = 4, Tsallis index HCDT = 0.29; q = 5, Renyi index = 0.27). Thus, these findings suggest that the yeast community in S. brasiliensis is perhaps two-to-three times more diverse than the present A. humile (Figure 5b).
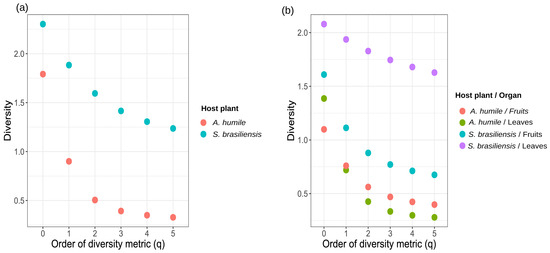
Figure 5.
Alpha diversity profiles for the community of cultivable yeast associated with aerial parts (fruits and leaves) of Anacardium humile and Sabicea brasiliensis shown by the Hill series (q). The alpha diversity metric is shown on the X-axis. The Y-axis shows the diversity value obtained on the X-axis for each diversity metric used. (a) = Diversity in plant host (Anacardium humile and Sabicea brasiliensis), (b) = Diversity in aerial parts (fruits and leaves). The circles show the hostesse/organ. q = 0, species richness; q = 1, Shannon–Wiener index (H’); q = 2, Simpson dominance (1/D’); q = 3, Gini–Simpson index; q = 4, Tsallis index (HCDT); q = 5, Renyi index. Figure created in software R version 4.1.1.
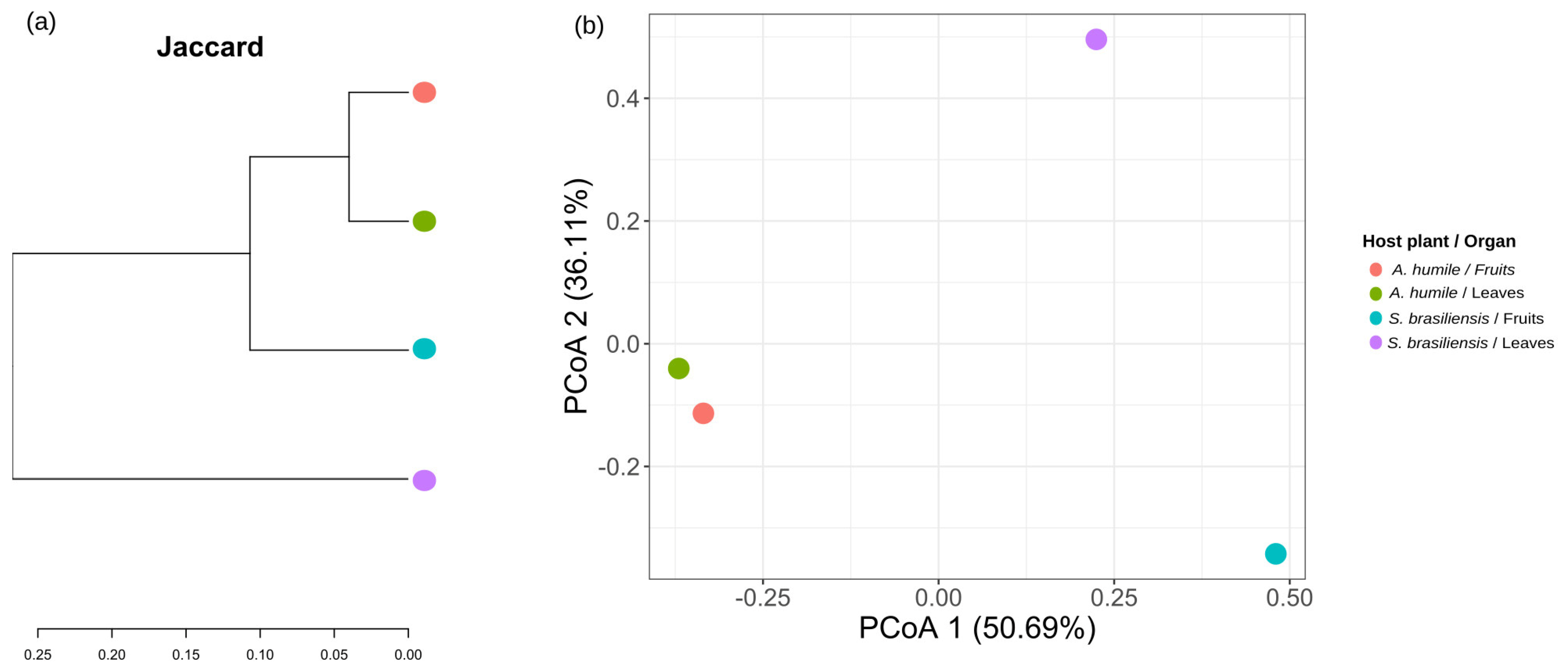
The similarity values obtained from the Jaccard distance, comparing the different types of organs analyzed between the host plants, show that the yeast community is more similar between the different organs of the same host species (Figure 6a). Principal coordinate analysis (PCoA), based on the Jaccard distance matrix, shows that yeast communities present in fruits of both host species and leaves of A. humile are distributed and spaced along the X axis (PCoA1) (Figure 6b). The yeast community associated with the leaves of S. brasiliensis is distant along the Y axis (PCoA2). The smallest distance value is found between the community present in the leaves and fruits of A. humile when compared with each other.
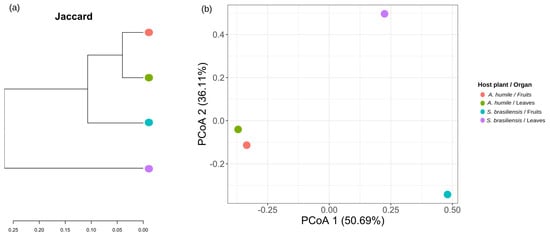
Figure 6.
Similarity ordinations are shown by the Jaccard distance of the yeast community associated with the fruits and leaves of Anacardium humile and Sabicea brasiliensis. (a) Jaccard similarity dendrogram, generated from a binary matrix (absence/presence), of the yeast community. (b) Principal coordinate analysis (PCoA) obtained from the Jaccard distance matrix. The percentage of variation explained by the plotted principal coordinates is indicated on the X and Y axes. The points in both Figures refer to the host species and the type of organ analyzed. The green ellipse in (b) refers to the host A. humile, while the red ellipse refers to the host S. brasiliensis. Figure created in software R version 4.1.1.
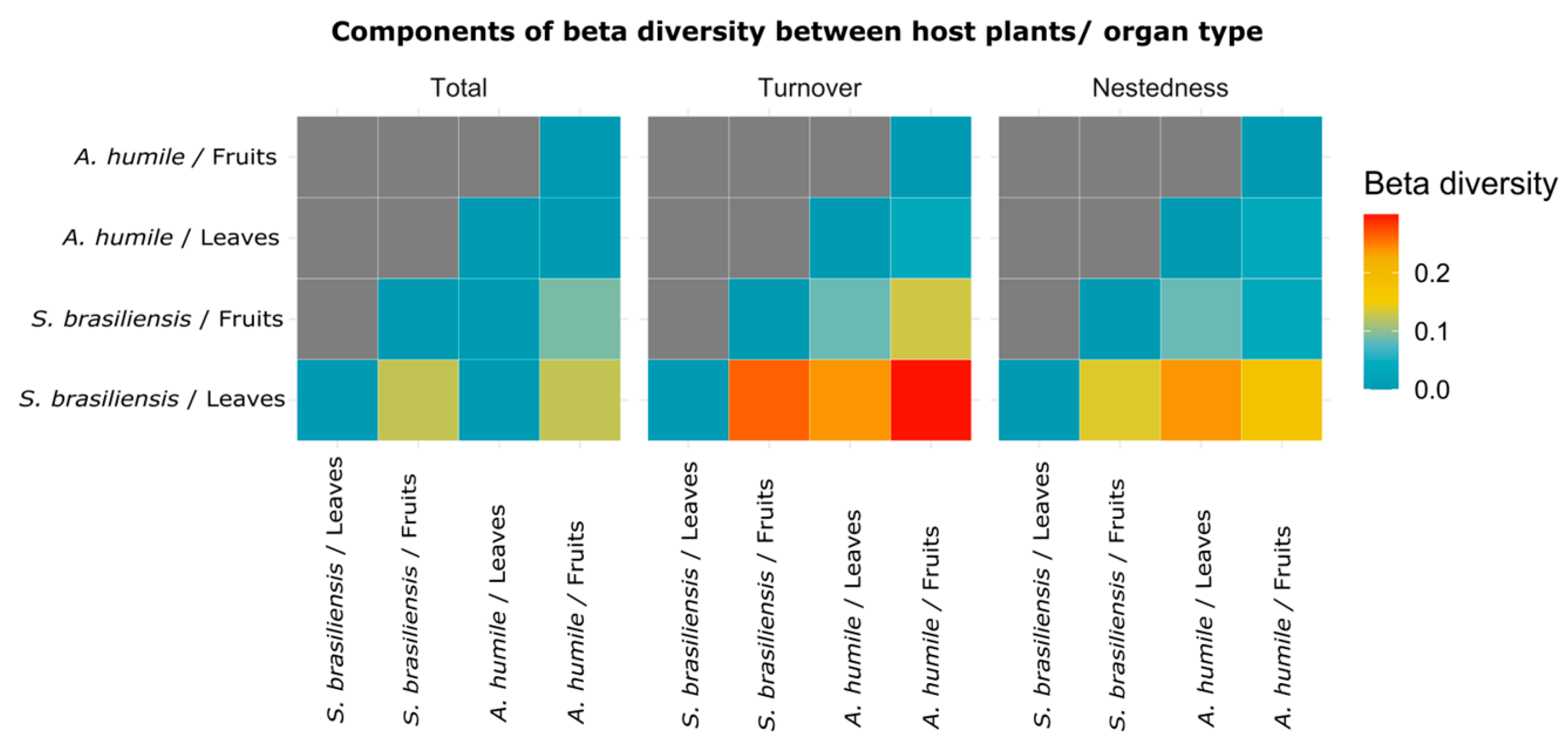
The pairwise comparison generated by the Sorensen beta diversity index points out the main components responsible for the dissimilarities (total = total dissimilarity value; turnover = species change/species gain; and nestedness = species loss/replacement) found in the community of yeasts associated with fruits and leaves of A. humile and S. brasiliensis (Figure 7). The highest values were shown by turnover, where the highest value is observed when the community present in S. brasiliensis fruits was compared with the community present in A. humile leaves. This finding suggests that the main factor responsible for the dissimilarities of the communities analyzed here is the gain in the number of species that occur in a host and its organs (S. brasiliensis) when compared with the species that occur in the other host and its organs (A. humile).
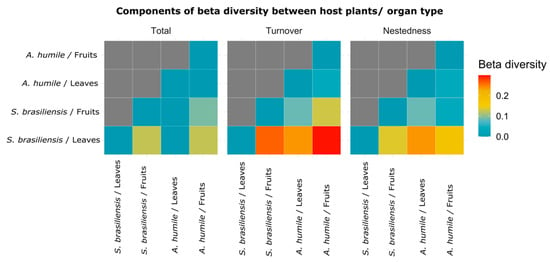
Figure 7.
Heatmap visualization showing the pairwise comparison obtained from the Sorenson index and its components (total, turnover, and nestedness) of the yeast community associated with leaves and fruits of A. humile and S. brasiliensis. The X-axis and Y-axis show the host species and organ types analyzed. The values obtained in each of the components of the Sorensen index is shown in the color scale: blue= lowest; red= highest. Figure created in software R version 4.1.1.
4. Discussion
There are still only a small number of studies on yeasts associated with native plants of this Brazilian savannah, so our work brings contributions to the knowledge about the occurrence and diversity of yeasts associated with leaves and fruits of A. humile and S. brasiliensis.
The first parameter evaluated was the population density of yeasts that occur in leaves and fruits of A. humile and S. brasiliensis. In general, population density values close to those found here (Table 2) have already been reported for some plant species in the Cerrado [18,19,35], but may be considered low when compared with other studies [19,36]. The density of microorganisms that occur in a given habitat is influenced by biotic and abiotic factors [14,15,37]. For example, the density values found here were higher in the fruits than in the leaves in both hosts and this may have been determined by the characteristics and nutrient availability of the fruits concerning the leaves as the fruits provide excellent habitats for yeasts due to its low pH, availability of micro and macronutrients, and active vectors that visit organs and plant tissues [14].
Most of the species found here in the fruits and leaves of S. brasiliensis and A. humile belong to the phylum Ascomycota (84.3%), with the classes Saccharomycetes dominant in the fruits and Dothideomycetes in the leaves. In some plant species from the Cerrado, ascomycetes represented more than 80% of the isolates found [20] with the predominance of the class Saccharomycetes in fruits [19] and Dothideomycetes in leaves [38]. The low occurrence of yeasts belonging to the phylum Basidiomycota in the fruits and leaves of both hosts can be explained by the fact that yeasts of this phylum usually colonize the soil [39], where they are the dominant group [40], or colonize the phylloplane of plant species, but occur in lower abundance [36,41]. In addition, we must consider the ecological filter effect exerted by the host in concert with environmental factors, as microbial communities are determined by this set of factors [42,43]. Thus, we can suggest that the influence of host-specific selective forces, such as available carbohydrates, creates conditions that determine the compositions of the communities of microorganisms found in each host [44].
Aureobasidium spp. was dominant in the leaves and fruits of A. humile. This genus comprises a complex of highly adaptative polymorphic yeast species with great phenotypic plasticity [45] and which are often reported as the main epiphytes of several plant species [46,47]. Furthermore, Aureobasidium spp. are known to produce a wide range of volatile organic compounds with fungistatic action [48,49], which could explain their dominance in the organs of A. humile. In the leaves and fruits of S. brasiliensis, the dominant species belong to the genera Candida, Aureobasidium, Wickerhamiella, and Rhodotorula. Species of the genera Candida and Rhodotorula are frequently reported in studies on the diversity of yeasts that occur in fruits [19,41,48,50,51,52] and leaves [35,53,54]. It is only A. humile that has had part of its mycobiota recently studied by our team using samples from another region of the Brazilian Cerrado [19], where Candida easanensis, Debaryomyces fabryi, D. nepalensis, Hanseniaspora meyeri, H. opuntiae, Meyerozyma caribbica and Rhynchogastrema complexa were reported. All of these species are different from the ones we found in the present work. This indicates that environmental resources and conditions have an important effect on the assembly of host plant mycobiota. The more samples you collect from the same plant species in different regions of the Cerrado, the more yeast species are isolated.
Among the yeast species shared between organs and hosts, only Aureobasidium and H. zeae occur in both hosts. The presence of yeasts of the same species colonizing different plant species from the same geographic region has already been reported in the Cerrado [19] and is perhaps related to vectors that act by dispersing these microorganisms and contributing directly to the structuring taxonomy of the assemblage of microorganisms that colonize plant tissues of different species [55,56]. For example, Wickerhamiella dianesei f.a., sp. nov., a new species of yeast isolated from leaves of Sabicea brasiliensis (Rubiaceae), was isolated from the flower of Byrsonima crassifolia (Malpighiaceae) in Brazil and from the stingless bee, Trigona fulviventris, collected from an Asteraceae flower in Costa Rica [57]. The report of these yeasts associated with stingless bees (Trigona fulviventris) reinforces the claim that these insects can be vectors of yeast between leaves, nectaries, flowers, and the fruits of the different plants visited by them. It should also be noted that the number of shared species is smaller than the number of species exclusive to each host plant, demonstrating the host’s role in filtering yeast species.
The alpha diversity metrics showed differences in species richness and diversity between hosts and tissue types, while the beta diversity metrics showed the main differences between these communities. In S. brasiliensis, the highest values were observed for the indices that measure species richness and diversity compared with A. humile (alpha diversity). Turnover suggests that the factor linked to the differences between communities associated with different hosts and their organs was the gain of species from one host concerning another. Similar to density, the richness and diversity of yeast species in a given niche are also influenced by numerous deterministic factors (nutrient availability, dispersal limitations, host genetics, environmental filtration, etc.) [41,58,59,60,61]. Some of the factors that may explain this difference in the number of species isolated among the hosts and in the organs analyzed here are: (1) the environmental filtering process (for example, the difference between the thickness and composition of the fruit peel, as well as the nutrients present in organs analyzed, organs may require some microorganisms to develop strategies to colonize them) [62]; (2) the genotype of the host plant, which can act by selecting the composition of the microbial assemblage [1].
The Cerrado presents an extreme abundance of endemic species of native plants and is recognized as a global biodiversity hotspot [63]. Our data indicate that Cerrado plants can also be considered an important habitat for yeast diversity and may potentially include new and/or previously unreported species, such as some of the yeasts that were isolated from leaves of a host that has since been described as the new species Wickerhamiella dianesei f.a., sp. nov. [57].
5. Conclusions
The plants sampled proved to be substrates with the potential to isolate yeasts of industrial interest, as studies with ecological, taxonomic, and biotechnological purposes of yeasts in Brazil are still scarce, especially for the Cerrado biome. The most frequent genera were Aureobasidium, Candida, Rhodotorula, and Wickerhamiella, with the genus Candida showing the greatest diversity of species. The Wickerhamiella isolates made possible the subsequent description of a new species within this genus. Our data reveal the yeast species that inhabit different Cerrado plant organs and indicate that these hosts can be considered an important habitat for yeast diversity, including new and/or previously unreported species in this biome.
Author Contributions
Conceptualization, E.M.S. and H.M.M.d.V.; methodology, E.M.S. and H.M.M.d.V.; software, J.B.A.d.R.; validation, E.M.S.; H.M.M.d.V. and J.B.A.d.R.; formal analysis, E.M.S.; H.M.M.d.V.; J.B.A.d.R. and L.G.F.C.; investigation, E.M.S.; resources, H.M.M.d.V.; data curation, E.M.S.; J.B.A.d.R. and L.G.F.C.; writing—original draft preparation, E.M.S.; H.M.M.d.V.; J.B.A.d.R. and L.G.F.C.; writing—review and editing, H.M.M.d.V. and J.B.A.d.R.; visualization, H.M.M.d.V.; supervision, H.M.M.d.V.; project administration, H.M.M.d.V.; funding acquisition, H.M.M.d.V. All authors have read and agreed to the published version of the manuscript.
Funding
Federal District Research Support Foundation (FAPDF)–Proc. SEI 00193.00000147/2019-61.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The nucleotide sequences generated from the yeasts isolated in this study were deposited in GenBank under the accession numbers OP912408-OP912460.
Acknowledgments
Financial support from the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brasil (CAPES)—Financing Code 001.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Whipps, J.M.; Hand, P.; Pink, D.; Bending, G.D. Phyllosphere microbiology with special reference to diversity and plant genotype. J. Appl. Microbiol. 2008, 105, 1744–1755. [Google Scholar] [CrossRef]
- Lindow, S.E.; Brandl, M.T. Microbiology of the phyllosphere. Appl. Environ. Microbiol. 2003, 69, 1875–1883. [Google Scholar] [CrossRef]
- Proença, D.N.; Grass, G.; Morais, P.V. Understanding pine wilt disease: Roles of the pine endophytic bacteria and of the bacteria carried by the disease-causing pinewood nematode. Microbiol. Open 2016, 6, e00415. [Google Scholar] [CrossRef]
- Sivakumar, N.; Sathishkumar, R.; Selvakumar, G.; Shyamkumar, R.; Arjunekumar, K. Phyllospheric Microbiomes: Diversity, Ecological Significance, and Biotechnological Applications. In Plant Microbiomes for Sustainable Agriculture—Sustainable Development and Biodiversity; Yadav, A., Singh, J., Rastegari, A., Yadav, N., Eds.; Springer: Berlin/Heidelberg, Germany, 2020; Volume 25, pp. 113–172. [Google Scholar] [CrossRef]
- Micci, A.; Zhang, Q.; Chang, X.; Kingsley, K.; Park, L.; Chiaranunt, P.; Strickland, R.; Velazquez, F.; Lindert, S.; Elmore, M.; et al. Histochemical Evidence for Nitrogen-Transfer Endosymbiosis in Non-Photosynthetic Cells of Leaves and Inflorescence Bracts of Angiosperms. Biology 2022, 11, 876. [Google Scholar] [CrossRef] [PubMed]
- Vermote, L.; Verce, M.; Mozzi, F.; De Vuyst, L.; Weckx, S. Microbiomes Associated With the Surfaces of Northern Argentinian Fruits Show a Wide Species Diversity. Front. Microbiol. 2022, 13, 872281. [Google Scholar] [CrossRef] [PubMed]
- Nasanit, R.; Tangwong-O-Thai, A.; Tantirungkij, M.; Limtong, S. The assessment of epiphytic yeast diversity in sugarcane phyllosphere in Thailand by culture-independent method. Fungal Biol. 2015, 119, 1145–1157. [Google Scholar] [CrossRef]
- Nasanit, R.; Jaibangyang, S.; Tantirungkij, M.; Limtong, S. Yeast diversity and novel yeast D1/D2 sequences from corn phylloplane obtained by a culture-independent approach. Antonie van Leeuwenhoek 2016, 109, 1615–1634. [Google Scholar] [CrossRef]
- Khunnamwong, P.; Jindamorakot, S.; Limtong, S. Endophytic yeast diversity in leaf tissue of rice, corn and sugarcane cultivated in Thailand assessed by a culture-dependent approach. Fungal Biol. 2018, 122, 785–799. [Google Scholar] [CrossRef] [PubMed]
- Into, P.; Pontes, A.; Sampaio, J.P.; Limtong, S. Yeast Diversity Associated with the Phylloplane of Corn Plants Cultivated in Thailand. Microorganisms 2020, 8, 80. [Google Scholar] [CrossRef]
- Boundy-Mills, K.L.; Glantschnig, E.; Roberts, I.N.; Yurkov, A.; Casaregola, S.; Daniel, H.M.; Groenewald, M.; Turchetti, B. Yeast culture collections in the twenty-first century: New opportunities and challenges. Yeast 2016, 33, 243–260. [Google Scholar] [CrossRef]
- Zhang, T.; Yao, Y.F. Endophytic fungal communities associated with vascular plants in the high Arctic zone are highly diverse and host-plant specific. PLoS ONE 2015, 10, e0130051. [Google Scholar] [CrossRef] [PubMed]
- Sipiczki, M. Overwintering of Vineyard Yeasts: Survival of Interacting Yeast Communities in Grapes Mummified on Vines. Front. Microbiol. 2016, 7, 212. [Google Scholar] [CrossRef] [PubMed]
- Vadkertiová, R.; Molnárová, J.; Vránová, D.; Sláviková, E. Yeasts and yeast-like organisms associated with fruits and blossoms of different fruit trees. Can. J. Microbiol. 2012, 58, 1344–1352. [Google Scholar] [CrossRef] [PubMed]
- Dabassa Koricha, A.; Han, D.Y.; Bacha, K.; Bai, F.Y. Occurrence and Molecular Identification of Wild Yeasts from Jimma Zone, South West Ethiopia. Microorganisms 2019, 7, 633. [Google Scholar] [CrossRef]
- Oliveira-Filho, A.; Ratter, J. A study of the origin of central Brazilian forests by the analysis of plant species distribution patterns. Edinb. J. Bot. 1995, 52, 141–194. [Google Scholar] [CrossRef]
- Silva, J.M.C.; Bates, J.M. Biogeographic patterns and conservation in the South American Cerrado: A tropical savanna hotspot. BioScience 2002, 52, 225–233. [Google Scholar] [CrossRef]
- Coelho, L.G.F.; Moreira, G.A.M.; Bomfim, C.A.; Vale, H.M.M. Yeast Communities Structure in Fruits of Native plant species of Brazilian Cerrado. Rev. Biol. Neotrop. 2020, 17, 35–46. [Google Scholar] [CrossRef]
- Vale, H.M.M.; Reis, J.B.A.; Oliveira, M.; Moreira, G.A.M.; Bomfim, C.A. Yeasts in native fruits from Brazilian neotropical savannah: Occurrence, diversity and enzymatic potential. Biota Neotrop. 2021, 21. [Google Scholar] [CrossRef]
- Dos Reis, J.B.A.; do Vale, H.M.M.; Lorenzi, A.S. Insights into taxonomic diversity and bioprospecting potential of Cerrado endophytic fungi: A review exploring an unique Brazilian biome and methodological limitations. World J. Microbiol. Biotechnol. 2022, 38, 202. [Google Scholar] [CrossRef]
- Ferreira, P.R.B.; Mendes, C.S.O.; Rodrigues, C.G.; Rocha, J.C.M.; Royo, V.D.A.; Valério, H.M.; Oliveira, D.A.D. Antibacterial activity tannin-rich fraction fromleaves of Anacardium humile. Ciência Rural 2012, 42, 1861–1864. [Google Scholar] [CrossRef]
- Batista, J.C.; Santin, S.M.d.O.; Schuquel, I.T.A.; de Arruda, L.L.M.; Bersani-Amado, C.A.; de Oliveira, C.M.A.; Kato, L.; Ferreira, H.D.; da Silva, C.C. Chemical constituents and evaluation of antioxidant and anti-inflammatory activities of roots of Sabicea brasiliensis wernh (Rubiaceae). Quim. Nova 2014, 37, 638–642. [Google Scholar] [CrossRef]
- Royo, V.A.; Mercadante-Simões, M.O.; Ribeiro, L.M.; Oliveira, D.A.; Aguiar, M.M.; Costa, E.R.; Ferreira, P.R. Anatomy, Histochemistry, and Antifungal Activity of Anacardium humile (Anacardiaceae) Leaf. Microsc. Microanal. 2015, 21, 1549–1561. [Google Scholar] [CrossRef]
- Costa, T.R.; Francisco, A.F.; Cardoso, F.F.; Moreira-Dill, L.S.; Fernandes, C.A.H.; Gomes, A.A.S.; Guimarães, C.L.S.; Marcussi, S.; Pereira, P.S.; Oliveira, H.C.; et al. Gallic acid anti-myotoxic activity and mechanism of action, a snake venom phospholipase A2 toxin inhibitor, isolated from the medicinal plant Anacardium humile. Int. J. Biol. Macromol. 2021, 185, 494–512. [Google Scholar] [CrossRef]
- Lima Júnior, J.P.; Franco, R.R.; Saraiva, A.L.; Moraes, I.B.; Espindola, F.S. Anacardium humile St. Hil as a novel source of antioxidant, antiglycation and α-amylase inhibitors molecules with potential for management of oxidative stress and diabetes. J. Ethnopharmacol. 2021, 268, 113667. [Google Scholar] [CrossRef]
- Makimura, K.; Murayama, S.Y.; Yamaguchi, H. Detection of a wide range of medically important fungi by the polymerase chain reaction. J. Med. Microbiol. 1994, 40, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Yarrow, D. Methods for the isolation, maintenance and identification of yeasts. In The Yeasts—A Taxonomic Study; Kurtzman, C.P., Fell, J.W., Eds.; Elsevier Science Publishers: Amsterdam, The Netherlands, 1998; pp. 77–100. [Google Scholar]
- Libkind, D.; Brizzio, S.; Ruffini, A.; Gadanho, M.; Van Broock, M.; Sampaio, J.P. Molecular characterization of carotenogenic yeasts from aquatic environments in Patagonia, Argentina. Antonie van Leeuwenhoek 2003, 84, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P.; Robnett, C.J. Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie van Leeuwenhoek 1998, 73, 331–371. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Pielou, E.C. The Interpretation of Ecological Data: A Primer on Classification and Ordination; John Wiley and Sons: New York, NY, USA, 1984. [Google Scholar]
- Hill, M.O. Diversity and Evenness: A Unifying Notation and Its Consequences. Ecology 1973, 54, 427–432. [Google Scholar] [CrossRef]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef]
- Baselga, A.; Orme, C.D.L. betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Sperandio, E.M.; do Vale, H.M.M.; Moreira, G.A.M. Yeasts from native Brazilian Cerrado plants: Occurrence, diversity and use in the biocontrol of citrus green mould. Fungal Biol. 2015, 119, 984–993. [Google Scholar] [CrossRef]
- Landell, M.F.; Mautone, J.N.; Valente, P. Biodiversity of yeasts associated to bromeliads in Itapuã Park, Viamão—RS. Biociências 2006, 14, 144–149. [Google Scholar]
- Dik, A.J.; Fokkema, N.J.; van Pelt, J.A. Influence of climatic and nutritional factors on yeast population dynamics in the phyllosphere of wheat. Microb. Ecol. 1992, 23, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Noriler, S.A.; Savi, D.C.; Aluizio, R.; Palácio-Cortes, A.M.; Possiede, Y.M.; Glienke, C. Bioprospecting and Structure of Fungal Endophyte Communities Found in the Brazilian Biomes, Pantanal, and Cerrado. Front. Microbiol. 2018, 9, 1526. [Google Scholar] [CrossRef]
- Yurkov, A.M. Yeasts of the soil—Obscure but precious. Yeast 2018, 35, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Lei, Y.; Wang, C.; Wei, Y.; Wang, C.; Sun, Y. Patterns of yeast diversity distribution and its drivers in rhizosphere soil of Hami melon orchards in different regions of Xinjiang. BMC Microbiol. 2021, 21, 170. [Google Scholar] [CrossRef]
- Grondin, E.; Sing, S.C.A.; Caro, Y.; Raherimandimby, M.; Randrianierenana, A.L.; James, S.; Nueno-Palop, C.; François, J.M.; Petit, T. A comparative study on the potential of epiphytic yeasts isolated from tropical fruits to produce flavoring compounds. Int. J. Food Microbiol. 2015, 203, 101–108. [Google Scholar] [CrossRef]
- Dang, H.; Lovell, C.R. Microbial Surface Colonization and Biofilm Development in Marine Environments. Microbiol. Mol. Biol. Rev. 2015, 80, 91–138. [Google Scholar] [CrossRef]
- Huang, Z.; Hou, D.; Zhou, R.; Zeng, S.; Xing, C.; Wei, D.; Deng, X.; Yu, L.; Wang, H.; Deng, Z.; et al. Environmental Water and Sediment Microbial Communities Shape Intestine Microbiota for Host Health: The Central Dogma in an Anthropogenic Aquaculture Ecosystem. Front. Microbiol. 2021, 12, 772149. [Google Scholar] [CrossRef]
- Singh, D.K.; Sharma, V.K.; Kumar, J.; Mishra, A.; Verma, S.K.; Sieber, T.N.; Kharwar, R.N. Diversity of endophytic mycobiota of tropical tree Tectona grandis Linn.f.: Spatiotemporal and tissue type effects. Sci. Rep. 2017, 7, 3745. [Google Scholar] [CrossRef]
- Zalar, P.; Gostincar, C.; de Hoog, G.S.; Ursic, V.; Sudhadham, M.; Gunde-Cimerman, N. Redefinition of Aureobasidium pullulans and its varieties. Stud. Mycol. 2008, 61, 21–38. [Google Scholar] [CrossRef]
- Di Francesco, A.; Zajc, J.; Gunde-Cimerman, N.; Aprea, E.; Gasperi, F.; Placì, N.; Caruso, F.; Baraldi, E. Bioactivity of volatile organic compounds by Aureobasidium species against gray mold of tomato and table grape. World J. Microbiol. Biotechnol. 2020, 36, 171. [Google Scholar] [CrossRef]
- Bozoudi, D.; Tsaltas, D. The Multiple and Versatile Roles of Aureobasidium pullulans in the Vitivinicultural Sector. Fermentation 2018, 4, 85. [Google Scholar] [CrossRef]
- Kucharska, K.; Wachowska, U.; Czaplicki, S. Wheat phyllosphere yeasts degrade propiconazole. BMC Microbiol. 2020, 20, 242. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, A.; Ugolini, L.; Lazzeri, L.; Mari, M. Production of volatile organic compounds by Aureobasidium pullulans as a potential mechanism action against postharvest fruit pathogens. Biol. Control 2015, 81, 8–14. [Google Scholar] [CrossRef]
- Morais, P.B.; Martins, M.B.; Klaczko, L.B.; Mendonça-Hagler, L.C.; Hagler, A.N. Yeast succession in the Amazon fruit Parahancornia amapa as resource partitioning among Drosophila spp. Appl. Environ. Microbiol. 1995, 61, 4251–4257. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Castrillón, M.; Usman, L.M.; Silva-Bedoya, L.M.; Osorio-Cadavid, E. Dominant yeasts associated to mango (Mangifera indica) and rose apple (Syzygium malaccense) fruit pulps investigated by culture-based methods. An. Acad. Bras. Cienc. 2019, 91, e20190052. [Google Scholar] [CrossRef]
- Vegas, C.; Zavaleta, A.I.; Canales, P.E.; Esteve-Zarzoso, B. Yeasts Associated with Various Amazonian Native Fruits. Pol. J. Microbiol. 2020, 69, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Shiraishi, K.; Oku, M.; Kawaguchi, K.; Uchida, D.; Yurimoto, H.; Sakai, Y. Yeast nitrogen utilization in the phyllosphere during plant lifespan under regulation of autophagy. Sci. Rep. 2015, 5, 9719. [Google Scholar] [CrossRef]
- Limtong, S.; Into, P.; Attarat, P. Biocontrol of Rice Seedling Rot Disease Caused by Curvularia lunata and Helminthosporium oryzae by Epiphytic Yeasts from Plant Leaves. Microorganisms 2020, 8, 647. [Google Scholar] [CrossRef]
- Belisle, M.; Peay, K.G.; Fukami, T. Flowers as islands: Spatial distribution of nectar-inhabiting microfungi among plants of Mimulus aurantiacus, a hummingbird-pollinated shrub. Microb. Ecol. 2012, 63, 711–718. [Google Scholar] [CrossRef]
- Mittelbach, M.; Yurkov, A.M.; Nocentini, D.; Nepi, M.; Weigend, M.; Begerow, D. Nectar sugars and bird visitation define a floral niche for basidiomycetous yeast on the Canary Islands. BMC Ecol. 2015, 15, 2. [Google Scholar] [CrossRef]
- Lachance, M.-A.; Vale, H.M.M.; Sperandio, E.M.; Carvalho, A.O.S.; Santos, A.R.O.; Grondin, C.; Jacques, N.; Casaregola, S.; Rosa, C.A. Wickerhamiella dianesei f.a., sp. nov. and Wickerhamiella kurtzmanii f.a., sp. nov., two yeast species isolated from plants and insects. Int. J. Syst. Evol. Microbiol. 2018, 68, 3351–3355. [Google Scholar] [CrossRef] [PubMed]
- Canto, A.; Herrera, C.M.; Rodriguez, R. Nectar-living yeasts of a tropical host plant community: Diversity and effects on community-wide floral nectar traits. PeerJ 2017, 5, e3517. [Google Scholar] [CrossRef] [PubMed]
- Ricks, K.D.; Koide, R.T. The role of inoculum dispersal and plant species identity in the assembly of leaf endophytic fungal communities. PLoS ONE 2019, 14, e0219832. [Google Scholar] [CrossRef]
- Ettinger, C.L.; Vann, L.E.; Eisen, J.A. Global Diversity and Biogeography of the Zostera marina Mycobiome. Appl. Environ. Microbiol. 2021, 87, e0279520. [Google Scholar] [CrossRef] [PubMed]
- Malassigné, S.; Minard, G.; Vallon, L.; Martin, E.; Valiente Moro, C.; Luis, P. Diversity and Functions of Yeast Communities Associated with Insects. Microorganisms 2021, 9, 1552. [Google Scholar] [CrossRef]
- Tournas, V.H.; Katsoudas, E. Mould and yeast flora in fresh berries, grapes and citrus fruits. Int. J. Food Microbiol. 2005, 105, 11–17. [Google Scholar] [CrossRef]
- Damasco, G.; Fontes, C.; Françoso, R.; Haidar, R. The Cerrado Biome: A Forgotten Biodiversity Hotspot. Front. Young Minds 2018, 6, 22. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).