Abstract
Geographic variation in diet can indicate if species have unique ecological roles or feeding strategies in different environments. Bats have a unique nocturnal ecological niche and a diverse diet, making them an ideal model for studying changes in feeding. Some bats have adapted to reduced food resources caused by environmental changes and human disturbances by expanding their dietary ecological niche. As one of the typical species whose dietary niche has expanded from invertebrates (insects) to vertebrates (birds), the feeding habits of the great evening bat (Ia io) have attracted much attention from researchers. Unfortunately, little is known about how the diet of Ia io varies in different regions. Therefore, in this study, the diversity and composition of the bird prey of the great evening bat from two populations were examined using cytochrome C oxidase subunit I (COI) amplicon sequencing. The results showed that the great evening bat in both populations preyed on a variety of passerine birds, with the Phylloscopidae being the dominant family. Although most birds preyed upon by these two populations are the same, there are significant differences in predation on Cettiidae between the two populations. Both populations prey on migratory birds more than resident birds. Both populations of bats can prey on finch birds weighing more than 15 g, but they preyed on more birds weighing less than 10 g. The results of this study provide an important addition to the study of changes in bat diets under the conditions of expanding dietary ecological niches.
1. Introduction
Habitat choice is critical for animal survival, and animal diets change depending on geography and habitat [1]. Species occupy vast geographic areas with different environments by altering their intraspecific diet or foraging behavior [2]. Intraspecific differences in diet or foraging behavior among geographic populations may be due to differences in abiotic factors such as temperature and different external factors such as habitat type, prey availability, and interspecific competition [3]. It has been shown that there are significant differences in the feeding habits of insectivorous bats in different geographic populations [4]. Clare found that the diets of little brown bats (Myotis lucifugus) in Canada differ significantly even when separated by only a few hundred kilometers [5]. And, there are differences in insect predation for the big brown bat (Eptesicus fuscus) in Indiana and Illinois [6], particularly for some island-distributed populations that differ in morphology and feeding habits compared to mainland populations [5,7]. For example, the subspecies of Hoary bat (Lasiurus cinereus) on the Galapagos Islands differ significantly from the mainland subspecies in their feeding habits. Information on the resource use and trophic dynamics of predators is important for understanding their role in ecosystem functioning and predicting population-level responses to environmental change.
Optimal foraging theory states that predators will maximize their energy intake at an evolutionary scale by turning to abundant, unexploited resources to gain fitness [8]. Most bats feed primarily on insects, but also prey on some small vertebrate animals, including frogs, birds, and rodents [9]. Bat predation on birds, a rare phenomenon in nature, is a typical case of expansion of the feeding ecological niche from invertebrates (insects) to vertebrates (birds) [10,11]. Rare ecological niche expansion may also occur in different geographic populations, and we know little about whether there are differences in the prey that expand in the diet of different geographic populations. Populations in different geographic regions may experience different prey availability, or individuals may selectively seek out prey that is more nutritious or energetically beneficial under different environmental conditions, resulting in geographic variation in diet [5]. Bats are nocturnal mammals with a wide range of feeding habits, and there are more than 1400 species of bats worldwide, accounting for one-fifth of the world’s mammals [12]. The Chiroptera occupy a unique nocturnal ecological niche and have a wide range of food habits, making them an ideal taxon for studying changes in the food habits of different populations [13]. Bat predation of birds was previously little known, and even among carnivorous bats predation of birds is rare [10]. So far, only nine species of carnivorous bats have been reported to prey on birds, namely Vampyrum spectrum, Chrotopterus auritus, Trachops cirrhosus, Megaderma lyra, Macroderma gigas, Nycteris grandis, Nyctalus lasiopterus, Nyctalus aviator, and Ia io [14,15,16]. Out of them, only a few known carnivorous bats have been found to have predatory behavior towards birds in flight, such as Nyctalus lasiopterus and Ia io [10,17].
The great evening bat is a cave-dwelling species endemic to Southeast Asia and is one of the largest aerial foragers among the Chiroptera [18]. This species is mainly distributed in China in Jiangsu, Guangdong, Sichuan, Hubei, Guizhou, and Yunnan provinces [11,19]. They feed mainly on insects, and as their dietary niche expands, they can utilize more food resources [20]. Thabah (2007) first reported that Ia io distributed in India and China feed on birds [10]. This study found residual bird feathers in the guano, confirming that the great evening bat is a bird-eating bat, and based on the feather color identification, it is hypothesized that the bird preyed upon by Ia io is likely to be the Phylloscopus affinis. The successful predation of birds by bird-eating bats and their use of different bird species are necessarily influenced by ecological selection [20]. Studies related to the food habits of different populations of species reveal the food needs of wild animals, and their utilization patterns, and help researchers determine the appropriate habitat for the species and how to manage it effectively.
According to the previous literature [21,22,23], we hypothesize that there are differences in avian prey taken by different geographic populations of Ia io. We predict that using high-throughput sequencing techniques to measure Ia io feeding in the two locations will differ in the following ways: (1) bird prey species composition; (2) bird prey diversity.
2. Materials and Methods
2.1. Study Site
This study was conducted at two sites, Fei Long Cave, Xingyi City, Guizhou Province, China (24°58.426′ N, 104°52.687′ E) and a bat cave in Longfeng Village, Qinghua Township, Weishan, Dali, Yunnan Province, China (25°1′40.98″ N, 100°12′32.43″ E). The study site in Guizhou is a mountainous area with elevations ranging from 1500 m to 2200 m; the study site in Yunnan has an elevation of 1878 m. The study area in Guizhou has a subtropical humid monsoon climate with well-developed karst landscapes at this site, and the territory has a warm and humid climate, with an average annual temperature of about 15 °C and an annual rainfall of between 1000 and 1400 mm [24]. The study site in Yunnan has a low-latitude highland monsoon climate with small temperature differences between the four seasons, divided into dry and rainy seasons [25].
2.2. Fecal Collection
In this study, Guizhou guano samples were collected from October to November, and Yunnan guano samples were collected from 28 to 31 October. Mist nets were used to capture the bats at the entrance of the cave when they returned from foraging (20:00–7:00). Sampling was completed on alternate days to reduce disturbance to the bat cave. Each bat was placed individually in a clean and disinfected cotton bag for 30–60 min or until it defecated (less than two hours). Fecal pellets were collected and stored in 2 mL lyophilized tubes containing anhydrous ethanol. The samples were refrigerated for a short period (0–4 °C) before being transported to the laboratory for dry ice preservation. A total of 17 samples were collected from bat individuals in Guizhou (excluding pregnant individuals to avoid interfering with them and affecting the fetus); a total of 14 samples were collected from individuals in Yunnan (excluding pregnant individuals). After the guano collection, the bats were released into the habitat. All collected samples were transported back to the laboratory using dry ice and stored in a −80 °C refrigerator for subsequent experiments.
2.3. DNA Extraction and PCR Amplification
Before extraction, all guano pellets from individual bats were homogenized using a grinder (TL2010S, DHS Life Sciences Ltd., Beijing, China). The guano mixture was sampled at 150–180 mg and DNA from the prey was extracted using the QIAamp DNA Guano Mini Kit (Qiagen, Crawley, UK), modified accordingly according to the manufacturer’s instructions. To aid adequate solubilization, the first step of thermal suspension at 70 °C was increased from 5 to 20 min, with a final elution in 60 uL of AE buffer.
In the bird DNA analysis, COI fragments up to 380 bp long were amplified using bird-specific primer pairs COIPreyFW (CGAGCAGARCTAGGCCAACC) and COIPreyRW (GCAGGCGGTTTATGTTTTTTGTGATTGCTG) [26]. Both forward and reverse primers were sequence tagged with adapters, pads, and connectors. A unique barcode sequence was appended to the meta-labeled primers to allow for the multiplexing of the samples. PCR amplification was then performed using ABI GeneAmp 9700, and all PCRs were performed following the protocol in Pastor-beviá et al. (2014) [17]. Each PCR sequence included a pair of PCR blanks. Each sample was mixed in triplicate after PCR amplification. All PCR products were visualized using 2% agarose gel electrophoresis, then purified with the AxyPrep DNA Gel Extraction Kit (Axygen Science, Union City, CA, USA) and quantified with QuantiFluorTM-ST (Promega, Madison, WI, USA) according to the manufacturer’s protocol. Purified products were mixed under equimolar conditions and paired-end sequencing (2 × 300) was performed on the Illumina MiSeq platform (Illumina Inc., Sandiego, CA, USA) as described in the standard protocol of Majorbio Biopharmaceutical Biotechnology Co. in Shanghai, China (https://www.majorbio.com/) (accessed on 26 July 2022).
2.4. Sequence Analysis and Taxonomic Identification
The raw sequences were quality-filtered using Trimomatic [27] and merged with Flash [28]. Valid sequences were obtained after quality processing, and individual sequences were classified and excluded using Usearch [29]. At a 97% similarity threshold, the remaining sequences were clustered into MOTUs (molecular operational taxonomic units) using Usearch, while removing chimeras [29]. The identification of prey took place by removing MOTUs with <1% of the total sequences of birds in each sample to eliminate possible errors and low abundance sequences. The representative sequences of each MOTU were identified taxonomically based on a similarity threshold of >98% using the BOLD database and via BLAST comparison in the GenBank database [30], where MOTUs that did not match the reference sequence or did not meet the taxonomic criteria were considered unidentified.
2.5. Statistical Analysis
The residence type and body mass of the above-identified bird species were obtained from The Avifauna of Guizhou [31], Fauna Sinica: Aves: Passeriformes (Vols. 10, 13, 14) [32,33,34] and A Checklist on the Classification and Distribution of the Birds of China [35]. In order to investigate the migratory patterns of bird prey, we classified them as migratory birds (M, including S—summer visitor, W—winter visitor, and P—passing bird) and resident birds (R) following Gong et al. (2021) [36]. In order to study the selection of bird prey size by bats, we grouped the average body mass of each bird species into three categories (small size, <10 g; medium size, 10–15 g; and large size, >15 g) following Ibáñez et al. (2016) [37]. In the vegan of R 4.2.3 [38], the Shannon index [39] was used to obtain information on the diversity of predated birds in different regions, and they were tested for differences using the Wilcoxon rank-sum test. The differences in the food composition of different populations of the great evening bat were tested using the Kruskal–Wallis H Test. The Wilcoxon rank-sum test was used to test for differences in bird migration types and body mass in the great evening bat diets. The dietary ecological niche width of different bat geographic populations was quantified by calculating the Levins’ BA index [40].
3. Result
3.1. Differences in Bird Species Diversity among Different Populations of the Great Evening Bat
It was found that the great evening bat in the Guizhou population preyed on 17 species of passerine birds in 4 families (Phylloscopidae, Muscicapidae, Cettiidae, Zosteropidae; Table 1); while the Yunnan population preyed on 13 species of passerine birds belonging to 6 families (i.e., Phylloscopidae, Muscicapidae, Motacillidae, Locustellidae, Cettiidae, Zosteropidae; Table 2).

Table 1.
Bird prey identified in the diet of the Guizhou population, China. The frequency indicates the number of occurrences of the identified species in all samples. Migration patterns are divided into migratory birds (M, including S-summering birds, W-wintering birds, and P-travelers) and resident birds (R). In order to investigate the migratory patterns of bird prey, we classified them as migratory birds (M, including S—summer visitor, W—winter visitor, and P—passing bird) and resident birds (R) following [36]. Body mass is the average weight of males and females of each bird species.
Table 2.
Bird prey identified in the diet of the Yunnan population, China. The frequency indicates the number of occurrences of the identified species in all samples. Migration patterns are divided into migratory birds (M, including S-summering birds, W-wintering birds, and P-travelers) and resident birds (R). In order to investigate the migratory patterns of bird prey, we classified them as migratory birds (M, including S—summer visitor, W—winter visitor, and P—passing bird) and resident birds (R) following [36]. Body mass is the average weight of males and females of each bird species.
Phylloscopidae was the dominant family of bird prey preyed upon by the great evening bat in both regions (45% of prey items in Yunnan and 39% of prey items in Guizhou), representing the highest bird species diversity in the great evening bats diet, followed by Muscicapidae (27% of prey items in Yunnan and 30% of prey items in Guizhou). The remaining four families in the diet of the Yunnan population, Motacillidae, Cettiidae, Sylviidae, and Locustellidae, comprised a total of 27% of all prey items, including only one bird species, respectively (Figure 1, Table 1 and Table 2). The two remaining families in the diet of the Guizhou population, Cettiidae and Zosteropidae, together accounted for 31% of all prey items. The percentage frequency of occurrence (%FOO) and relative sequence abundance (%RRA) of bird prey in great evening bat feeding showed similar results at the family level (Figure 2 and Figure 3). The food composition of the different geographic populations differed significantly in Cettiidae (H = 12.479; p < 0.001, Figure 4). The diversity of birds preyed upon by Guizhou bat populations and Yunnan bat populations did not differ significantly (Z = 1.111, p = 0.279; Figure 5).
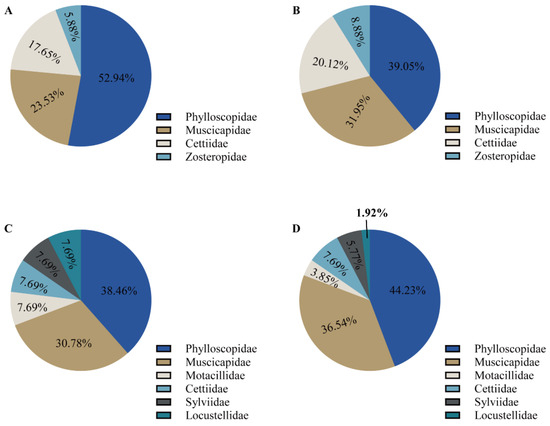
Figure 1.
Prey composition and diversity in the diet of the great evening bat. (A) Percent occurrence of bird at the family level in Guizhou, China. (B) Percent occurrence of bird prey items at the family level in Guizhou, China. (C) Percent occurrence of bird at the family level in Yunnan, China. (D) Percent occurrence of bird prey items at the family level in Yunnan, China.
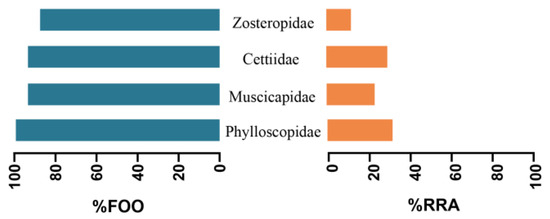
Figure 2.
Percentage frequency of occurrence (FOO) and relative read abundance (RRA) of bird prey families preyed upon by the Guizhou bat population, China.
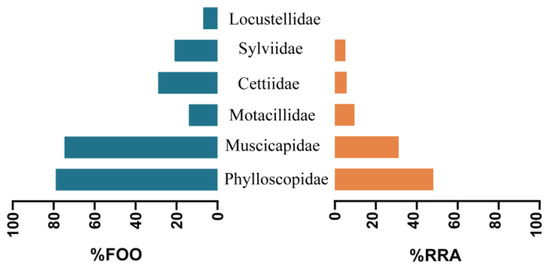
Figure 3.
Percentage frequency of occurrence (FOO) and relative read abundance (RRA) of bird prey families preyed upon by the Yunnan bat population, China.
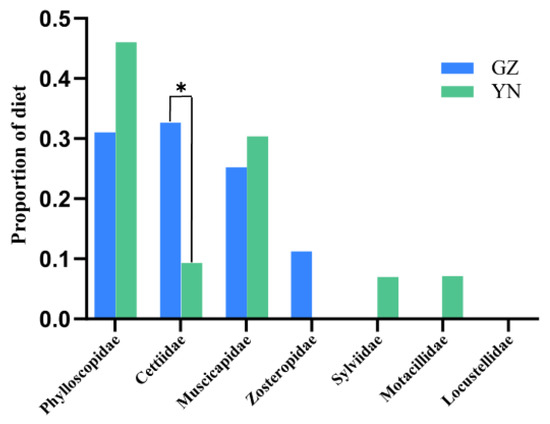
Figure 4.
Comparison of bird predation at the family level between the Guizhou and Yunnan bat populations, China. (* represent 0.01< p ≤ 0.05).
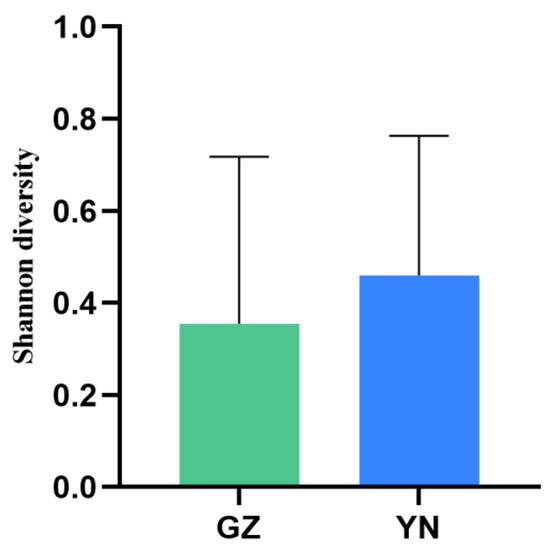
Figure 5.
Comparison of the diversity of predatory birds between Guizhou and Yunnan bat populations, China.
3.2. Selection of Bird Prey Types by Different Populations of the Great Evening Bat
The majority of avian prey items (64%) preyed on by the Guizhou population were migratory birds, and four species (24%): Phylloscopus yunnanensis, Myiomela leucura, Horornis fortipes, an Abroscopus albogularis were resident birds, accounting for 36% of all avian prey items (Figure 6a). The Guizhou populations prey on significantly more migratory birds than resident birds (Z = 4.173, p < 0.001). The majority of avian prey items (90%, excluding three prey items for which no data were available) preyed on by the Yunnan population were migratory birds, and two species (17%): Saxicola ferreus and Phyllergates cucullatus were resident birds, accounting for 10% of all bird prey items (Figure 6b). The Yunnan population also prey on significantly more migratory birds than resident birds (Z = 4.231, p < 0.001).
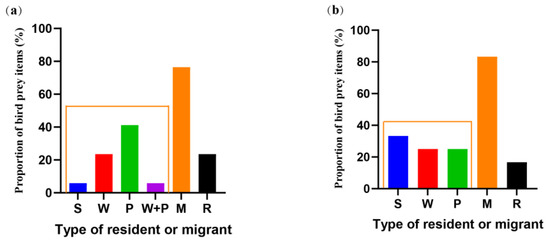
Figure 6.
The proportion of bird prey migration patterns. (a) Autumn in Guizhou. (b) Autumn in Yunnan. Migration patterns are divided into migratory birds (M, including S-summer migratory birds, W-winter migratory birds, and P-travelers) and resident birds (R).
The average body mass of the 17 prey species in Guizhou (excluding six species without data) was 10.00 ± 3.86 g, ranging from 3.5 g for P. yunnanensis to 18.5 g for C. calliope. The average body mass of identified prey items in Guizhou was 10.49 ± 3.89 g (Figure 7a). Compared to birds weighing more than 15 g, Guizhou bat populations preyed on more birds weighing less than 10 g (Z = 4.785, p < 0.001) and on birds weighing 10–15 g (Z = 4.691, p < 0.001). The average body mass of the 13 prey species in Yunnan (excluding two species without data) was 11.45 ± 4.95 g, ranging from 7.0 g for P. cucullatus to 21.5 g for Anthus hodgsoni. The average body mass of identified prey items in Yunnan was 10.41 ± 5.06 g (Figure 7b). Yunnan bat populations prey significantly more on birds weighing less than 10 g than on birds weighing 10–15 g (Z = 3.528, p < 0.001) and on birds weighing more than 15 g (Z = 2.650, p = 0.009).
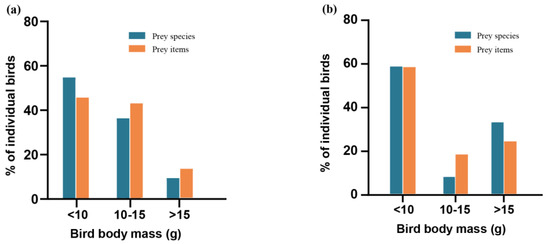
Figure 7.
Proportional distribution of bird prey species and prey entry weight. In order to study the selection of bird prey size by bats, we grouped the average body mass of each bird species into three categories (small size, <10 g; medium size, 10–15 g; and large size, >15 g) following Ibáñez et al. (2016). (a) Guizhou; (b) Yunnan.
The results of the ecological niche width index calculation showed the niche width of Yunnan and Guizhou populations represented by Levins’ BA index were 0.631 and 0.535, respectively.
4. Discussion
In this study, the differences in bird prey composition and diversity in the diets of two geographic populations of Ia io were examined. The results showed that: (1) Both populations preyed on a wide range of passerine birds, with Phylloscopidae dominating at the family level. Even though most birds preyed upon by these two populations were the same, there were significant differences in predation on Cettiidae between the two populations. This result is consistent with our first prediction. (2) The diversity of birds in the diet of the Yunnan population is slightly higher than that of the Guizhou population, but there is no significant difference between them, which is inconsistent with our second prediction. (3) Both populations preyed mostly on migratory birds, with resident birds in the minority. And, they are capable of hunting birds weighing more than 15 g, but preyed on more birds weighing less than 10 g.
The choice of prey species is influenced by a number of factors such as the existence of prey patches, prey availability and abundance, and the energy demand associated with prey capture and handling [41]. According to the previous literature [21,22,23], we hypothesized that differences in habitat would result in the heterogeneity of bird prey in the diet. Our results indicated that both populations preyed primarily on migratory birds, and the composition of bird prey eaten by individuals from Guizhou and Yunnan was not significantly different. About 200 species of birds are currently nocturnal and overlap with the activity of bats, increasing their likelihood of being prey [42]. Although migratory bird species diversity in both study sites was not investigated in this study, the existing literature shows that the number of bird resources in Yunnan is more than that in Guizhou. There are 275 species of nocturnal migratory birds in Yunnan province belonging to 15 orders and 37 families, accounting for 71.4%, 42.0%, and 30.6% of the recorded orders, families, and species of birds in Yunnan [43]. The wild bird resources in Guizhou province are 18 orders, 56 families, 216 genera, 450 species, and 100 subspecies of birds [44]. However, the two study locations are only 650 km apart, and many migratory birds can travel at speeds of around 100 km/day [45], which may allow for consistency in seasonal migratory bird prey abundance and availability in the two study areas when studies are conducted in the fall. In addition, due to the limited size of the sampling area and the limited sample size, it was likely that not all migratory bird species that the great evening bat preys on in the study area were covered.
Although the difference in diversity of bird prey between the two bat populations was not statistically significant, we still found that the bird prey diversity of low-latitude Yunnan bats was higher than that of high-latitude Guizhou bats, and there are significant differences in predation on Cettiidae between the two populations. Habitat selection is critical for bats, and diets may vary depending on geography and habitat [23]. This may be due to differences in habitat type and latitude in the study locations. The study site in Guizhou has a subtropical humid monsoon climate with well-developed karst landscapes [24], whereas the study site in Yunnan has a low-latitude highland monsoon climate [25]. Climate, landscape, and especially latitude are thought to have a huge impact on prey diversity [46]. A case study comparing the dietary diversity of lesser short-tailed bat (Mystacina tuberculata) populations at different latitudes confirmed that the population at lower latitudes had a higher prey diversity and feeding niche than that of the population at higher latitudes [47]. Our results were consistent with this, showing niche breadth and a number of prey species that decreased with latitude.
Ever since I. io expanded their diet from insects to birds [10], they have been preying mainly on small migratory birds. We found that I. io consumed 17 species of seven families from Passeriformes with a body mass of 7–18.5 g. Optimal foraging theory suggests that predators choose the size of their prey according to their body size [48]. In this study, there were no significant differences in body size and frequency of echolocation calls between the bats from the two locations (unpublished data). And, previous work from our group has shown that I. io body mass did not influence prey choice and predation richness on birds, suggesting I. io is an opportunistic carnivorous predator [36]. Because of the high energy cost of hunting migratory birds, becoming a generalist is beneficial for bird-eating bats. In addition, I. io has been shown to prey on birds that fly at higher altitudes in the fall, which may be one reason why both populations tend to prey on birds weighing less than 15 g in the passerine birds [49]. Furthermore, it is unclear whether migratory birds are captured during migration or while resting at the study site, and follow-up studies are needed to verify this.
In conclusion, we did not find significant differences in the composition of bird prey in their diets between the two populations of I. io with relatively small geographic distances. While we need to take these results with a grain of caution, again, opportunistic hunting must be most beneficial for a species like the great evening bat, which is an energy-intensive predator hunting fast-flying migratory birds. Future studies combining information on prey availability and abundance at broader spatial and temporal scales are needed to test the geographical differences in the diet composition of the great evening bat.
Author Contributions
Conceptualization, J.F.; methodology, T.J.; software, Y.L. (Yu Liu); validation, Y.L. (Yu Liu), H.W. and L.G.; formal analysis, Y.L. (Yingying Liu); resources, H.W.; data curation, L.G.; writing—original draft preparation, Y.L. (Yu Liu); writing—review and editing, Y.L. (Yu Liu); visualization, Y.L. (Yu Liu); supervision, H.W.; project administration, H.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Natural Science Foundation of China (Grant No. 32171489).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data is unavailable due to privacy.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Stephens, D.W.; Krebs, J.R. Foraging Theory; Princeton University Press: Princeton, NJ, USA, 1986; Volume 1. [Google Scholar]
- Maucieri, D.; Barclay, R.M. Consumption of spiders by the little brown bat (Myotis lucifugus) and the long-eared myotis (Myotis evotis) in the Rocky Mountains of Alberta, Canada. Can. J. Zool. 2021, 99, 221–226. [Google Scholar] [CrossRef]
- Kaupas, L.A.; Barclay, R.M. Temperature-dependent consumption of spiders by little brown bats (Myotis lucifugus), but not northern long-eared bats (Myotis septentrionalis), in northern Canada. Can. J. Zool. 2018, 96, 261–268. [Google Scholar] [CrossRef]
- Aspetsberger, F.; Brandsen, D.; Jacobs, D.S. Geographic variation in the morphology, echolocation and diet of the little free-tailed bat, Chaerephon pumilus(Molossidae). Afr. Zool. 2003, 38, 245–254. [Google Scholar] [CrossRef]
- Clare, E.L.; Symondson, W.O.; Broders, H.; Fabianek, F.; Fraser, E.E.; MacKenzie, A.; Boughen, A.; Hamilton, R.; Willis, C.K.; Martinez-Nuñez, F. The diet of Myotis lucifugus across Canada: Assessing foraging quality and diet variability. Mol. Ecol. 2014, 23, 3618–3632. [Google Scholar] [CrossRef]
- Whitaker, J.O., Jr. Food of the big brown bat Eptesicus fuscus from maternity colonies in Indiana and Illinois. Am. Midl. Nat. 1995, 134, 346–360. [Google Scholar] [CrossRef]
- Leite, T.; Batista, A.; Lima, F.; Barbosa, J.; Mather, J. Geographic variability of Octopus insularis diet: From oceanic island to continental populations. Aquat. Biol. 2016, 25, 17–27. [Google Scholar] [CrossRef]
- Popa-Lisseanu, A.G.; Delgado-Huertas, A.; Forero, M.G.; Rodríguez, A.; Arlettaz, R.; Ibáñez, C. Bats’ Conquest of a Formidable Foraging Niche: The Myriads of Nocturnally Migrating Songbirds. PLoS ONE 2007, 2, e205. [Google Scholar] [CrossRef] [PubMed]
- Norberg, U.M.; Rayner, J.M.V. Ecological morphology and flight in bats (Mammalia; Chiroptera): Wing adaptations, flight performance, foraging strategy and echolocation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1987, 316, 335–427. [Google Scholar] [CrossRef]
- Thabah, A.; Li, G.; Wang, Y.; Liang, B.; Hu, K.; Zhang, S.; Jones, G. Diet, Echolocation Calls, and Phylogenetic Affinities of the Great Evening Bat (Ia io; Vespertilionidae): Another Carnivorous Bat. J. Mammal 2007, 88, 728–735. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, Q.; Tan, L.; Shen, Q.; Chen, Z.; Gong, Y.; Xiang, Z.; Zhang, L. Ia io found in Guangdong Province. Chin. J. Zool. 2013, 2, 287–291. [Google Scholar]
- Zukal, J. Handbook of the Mammals of the World. J. Vertebr. Biol. 2020, 69, E2003. [Google Scholar] [CrossRef]
- Ingala, M.R.; Simmons, N.B.; Wultsch, C.; Krampis, K.; Speer, K.A.; Perkins, S.L. Comparing Microbiome Sampling Methods in a Wild Mammal: Fecal and Intestinal Samples Record Different Signals of Host Ecology, Evolution. Front. Microbiol. 2018, 9, 803. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez, C.; Juste, J.; García-Mudarra, J.L.; Agirre-Mendi, P.T. Bat predation on nocturnally migrating birds. Proc. Natl. Acad. Sci. USA 2001, 98, 9700–9702. [Google Scholar] [CrossRef] [PubMed]
- Gual-Suárez, F.; Medellín, R.A. We eat meat: A review of carnivory in bats. Mammal Rev. 2021, 51, 540–558. [Google Scholar] [CrossRef]
- Fukui, D.; Dewa, H.; Katsuta, S.; Sato, A. Bird predation by the birdlike noctule in Japan. J. Mammal 2013, 94, 657–661. [Google Scholar] [CrossRef][Green Version]
- Norberg, U.M.; Fenton, M.B. Carnivorous bats? Biol. J. Linn. Soc. 1988, 33, 383–394. [Google Scholar] [CrossRef]
- Osgood, W.H.; Allen, G.M. The Mammals of China and Mongolia. Part 2 of Vol. XI of Natural History of Central Asia. J. Mammal 1941, 22, 206–208. [Google Scholar] [CrossRef]
- Csorba, G. The distribution of the great evening bat Ia io in the Indo-Malayan region. Myotis 1998, 36, 197–201. [Google Scholar]
- Shi, B.; Wang, Y.; Gong, L.; Chang, Y.; Liu, T.; Zhao, X.; Lin, A.; Feng, J.; Jiang, T. Correlation of skull morphology and bite force in a bird-eating bat (Ia io; Vespertilionidae). Front. Zool. 2020, 17, 1–14. [Google Scholar] [CrossRef]
- Davis, N.E.; Forsyth, D.M.; Triggs, B.; Pascoe, C.; Benshemesh, J.; Robley, A.; Lawrence, J.; Ritchie, E.G.; Nimmo, D.G.; Lumsden, L.F. Interspecific and geographic variation in the diets of sympatric carnivores: Dingoes/wild dogs and red foxes in south-eastern Australia. PLoS ONE 2015, 10, e0120975. [Google Scholar] [CrossRef]
- Scherer, R.; Doll, A.; Rea, L.; Christ, A.; Stricker, C.; Witteveen, B.; Kline, T.; Kurle, C.; Wunder, M. Stable isotope values in pup vibrissae reveal geographic variation in diets of gestating Steller sea lions Eumetopias jubatus. Mar. Ecol. Prog. Ser. 2015, 527, 261–274. [Google Scholar] [CrossRef]
- Law, B.; Gonsalves, L.; Chidel, M.; McConville, A. When bat eats bat: Diet and roosts of the greater broad-nosed bat (Scoteanax rueppellii) across different regions and habitats. Aust. Mammal. 2023. [Google Scholar] [CrossRef]
- Xie, J.; He, G.; He, T. Influence of climatic factors on soil types and distribution in Guizhou. J. Zhejiang Agric. Sci. 2015, 56, 510–514. [Google Scholar]
- An, G.; Guo, Z.; Ye, P. Climatic Changes and Impacts on Water Quality of Erhai Lake in Dali Area, Yunnan Province over the Period from 1989 to 2019. Geoscience 2022, 36, 406. [Google Scholar]
- Pastor-Beviá, D.; Ibáñez, C.; García-Mudarra, J.L.; Juste, J. A Molecular Approach to the Study of Avian DNA in Bat Faeces. Acta Chiropterolog. 2014, 16, 451–460. [Google Scholar] [CrossRef][Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Aizpurua, O.; Budinski, I.; Georgiakakis, P.; Gopalakrishnan, S.; Ibañez, C.; Mata, V.; Rebelo, H.; Russo, D.; Szodoray-Parádi, F.; Zhelyazkova, V.; et al. Agriculture shapes the trophic niche of a bat preying on multiple pest arthropods across Europe: Evidence from DNA metabarcoding. Mol. Ecol. 2018, 27, 815–825. [Google Scholar] [CrossRef]
- Wu, Z.; Chen, K. The Avifauna of Guizhou; Guizhou People’s Publishing House: Guiyang, China, 1986. [Google Scholar]
- Cheng, T.; Long, Z.; Lu, T. Fauna Sinica: Aves. Volume 10 Passeriformes, Muscicapidae I. Turdinae; Science Press: Beijing, China, 1995. [Google Scholar]
- Fu, T.; Song, Y.; Gao, W. Fauna Sinica, Aves Volume 14 Passeriformes, Ploceidae and Fringillidae; Science Press: Beijing, China, 1998. [Google Scholar]
- Cheng, T.; Long, Z.; Zheng, B. Fauna Sinica: Aves, Volume 13, Passeriformes, Paridae and Zosteropidae; Science Press: Beijing, China, 1982. [Google Scholar]
- Zheng, G. A Checklist on the Classification and Distribution of the Birds of China; Geological Publishing House: Bath, UK, 2005. [Google Scholar]
- Gong, L.; Shi, B.; Wu, H.; Feng, J.; Jiang, T. Who’s for dinner? Bird prey diversity and choice in the great evening bat, Ia io. Ecol. Evol. 2021, 11, 8400–8409. [Google Scholar] [CrossRef]
- Ibáñez, C.; Popa-Lisseanu, A.G.; Pastor-Beviá, D.; García-Mudarra, J.L.; Juste, J. Concealed by darkness: Interactions between predatory bats and nocturnally migrating songbirds illuminated by DNA sequencing. Mol. Ecol. 2016, 25, 5254–5263. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Simpson, G. Package ‘vegan’ J. Community Ecol. 2019, 2. [Google Scholar]
- Allen, B.; Kon, M.; Bar-Yam, Y. A New Phylogenetic Diversity Measure Generalizing the Shannon Index and Its Application to Phyllostomid Bats. Am. Nat. 2009, 174, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Razgour, O.; Clare, E.L.; Zeale, M.R.K.; Hanmer, J.; Schnell, I.B.; Rasmussen, M.; Gilbert, T.P.; Jones, G. High-throughput sequencing offers insight into mechanisms of resource partitioning in cryptic bat species. Ecol. Evol. 2011, 1, 556–570. [Google Scholar] [CrossRef] [PubMed]
- Stephens, D.W.; Brown, J.S.; Ydenberg, R.C. Foraging: Behavior and Ecology; University of Chicago Press: Chicago, IL, USA, 2007. [Google Scholar]
- Fenton, M.B.; Fleming, T.H. Ecological Interactions between Bats and Nocturnal Birds. Biotropica 1976, 8, 104–110. [Google Scholar] [CrossRef]
- Bingxue, Z. The Study on Nocturnal Migrating Birds in Yunnan Province; Yunnan University: Kunming, China, 2015. [Google Scholar]
- Wang, H.; Wang, Y.; Xiang, Z.; Li, Q.; Li, R. The present condition of the bird resource in Guizhou. Environ. Prot. Technol. 2011, 17, 5–8+13. [Google Scholar]
- Schmaljohann, H. Proximate mechanisms affecting seasonal differences in migration speed of avian species. Sci. Rep. 2018, 8, 4106. [Google Scholar] [CrossRef]
- Clough, Y.; Putra, D.D.; Pitopang, R.; Tscharntke, T. Local and landscape factors determine functional bird diversity in Indonesian cacao agroforestry. Biol. Conserv: England. 2009, 142, 1032–1041. [Google Scholar] [CrossRef]
- Czenze, Z.J.; Tucker, J.L.; Clare, E.L.; Littlefair, J.E.; Hemprich-Bennett, D.; Oliveira, H.F.M.; Brigham, R.M.; Hickey, A.J.R.; Parsons, S. Spatiotemporal and demographic variation in the diet of New Zealand lesser short-tailed bats (Mystacina tuberculata). Ecol. Evol. 2018, 8, 7599–7610. [Google Scholar] [CrossRef]
- Krebs, J.R.; Stephens, D.W. Foraging Theory; Princeton University Press: Princeton, NJ, USA, 2019. [Google Scholar]
- Wang, Z.; Gong, L.; Huang, Z.; Geng, Y.; Zhang, W.; Si, M.; Wu, H.; Feng, J.; Jiang, T. Linking changes in individual specialization and population niche of space use across seasons in the great evening bat (Ia io). Mov. Ecol. 2023, 11, 32. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).