
Orthoptera Community Dynamics and Conservation in a Natura 2000 Site (Greece): The Role of Beta Diversity




Abstract
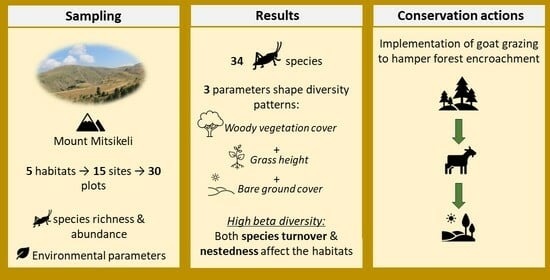
:
1. Introduction
2. Materials and Methods
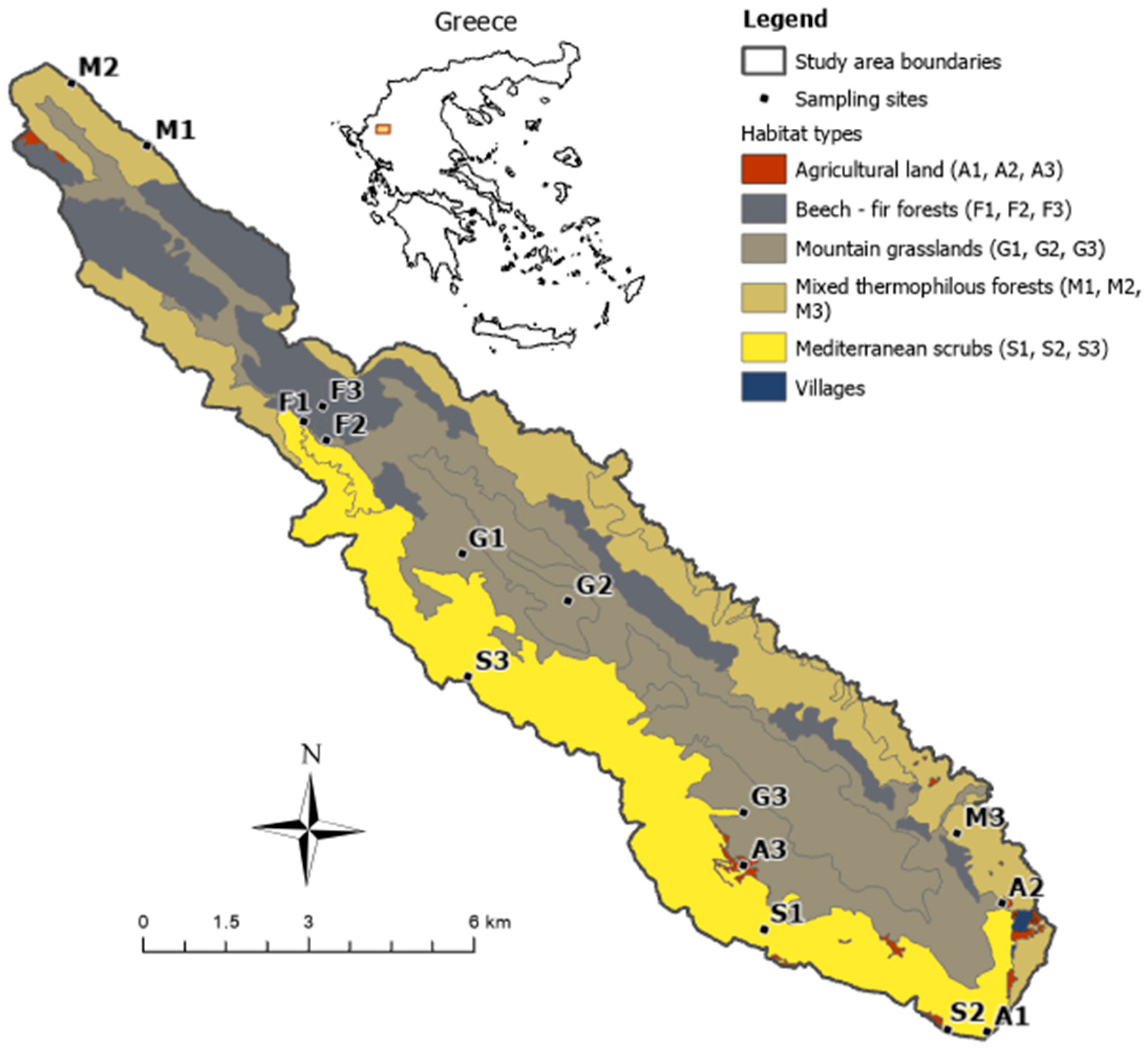
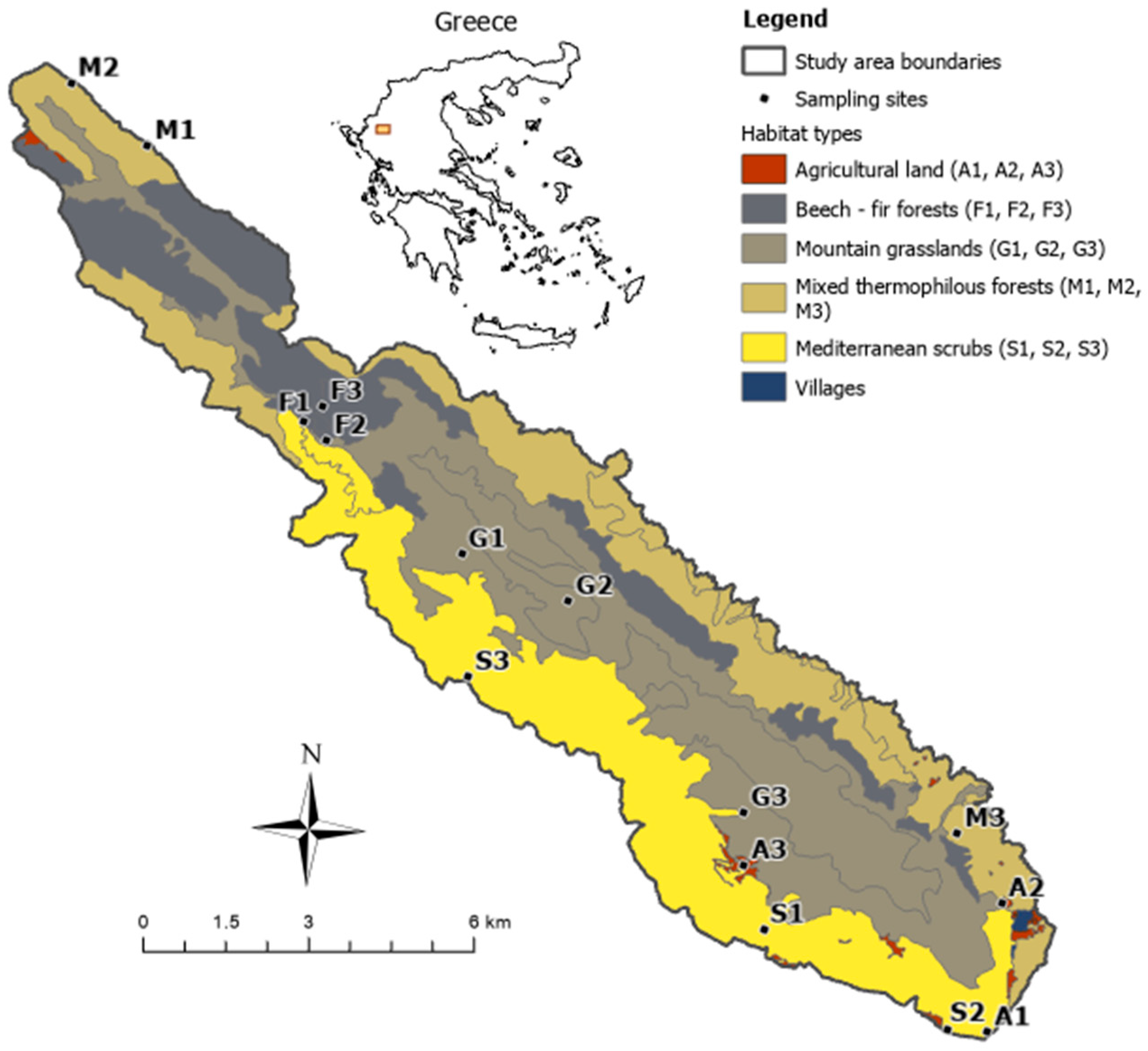
2.1. Study Area and Site Selection
2.2. Orthoptera Sampling
2.3. Environmental Factors
2.4. Data Analysis
2.4.1. Sampling Efficiency and Alpha Diversity
2.4.2. Community Composition and Structure
2.4.3. Beta Diversity and Its Partitions
3. Results
3.1. Species Richness
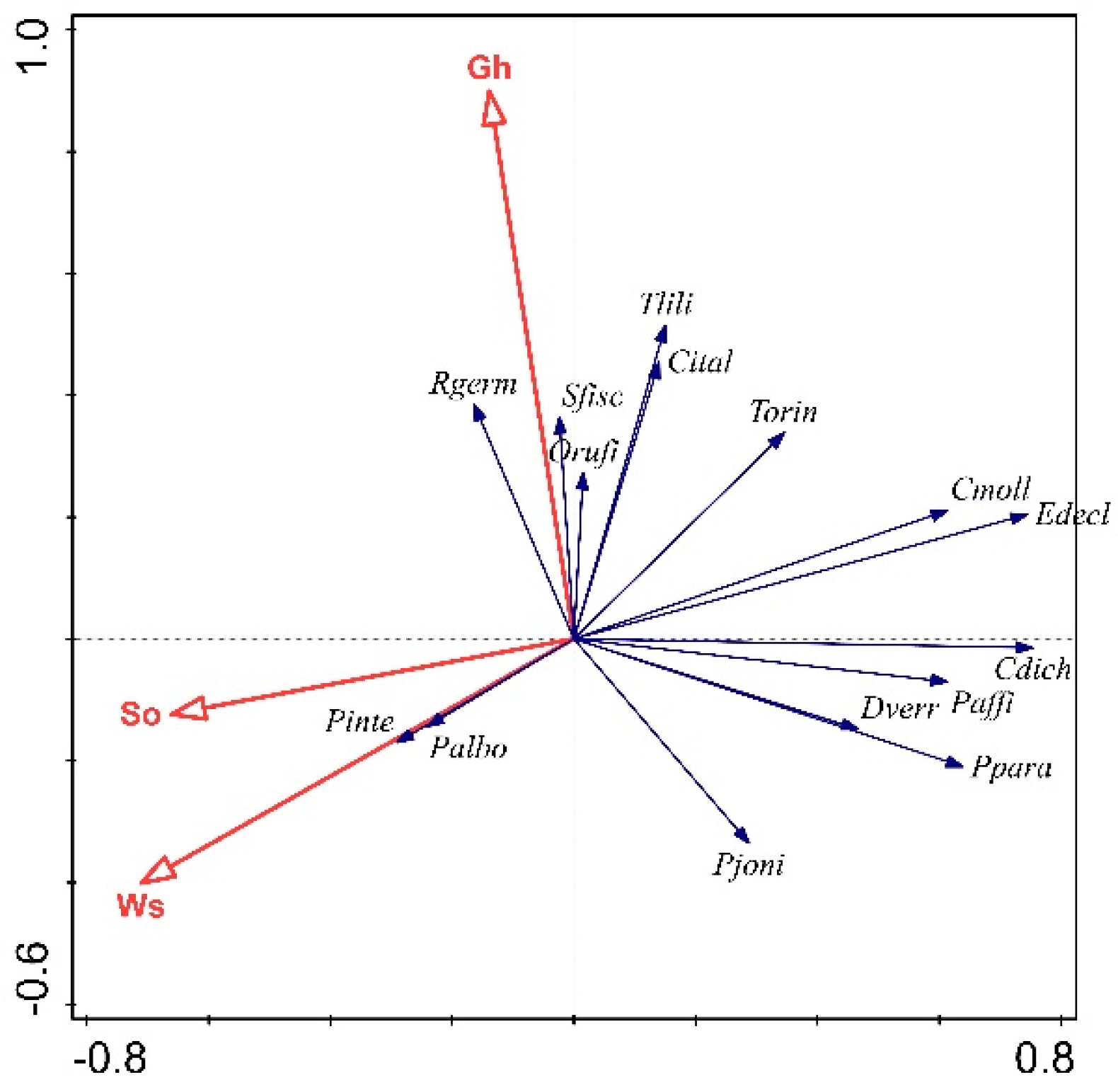
3.2. Environmental Factors Shaping Community Composition
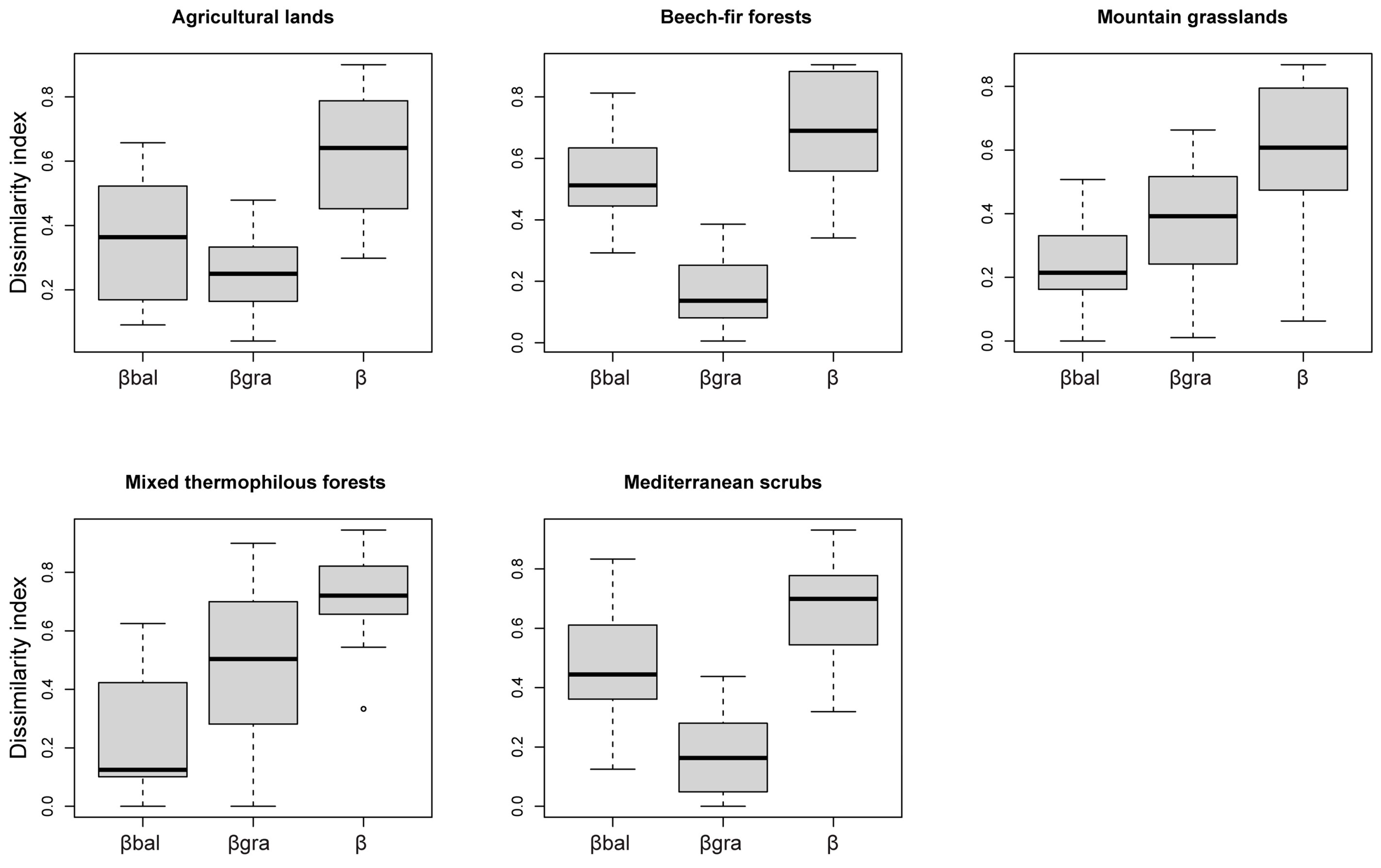
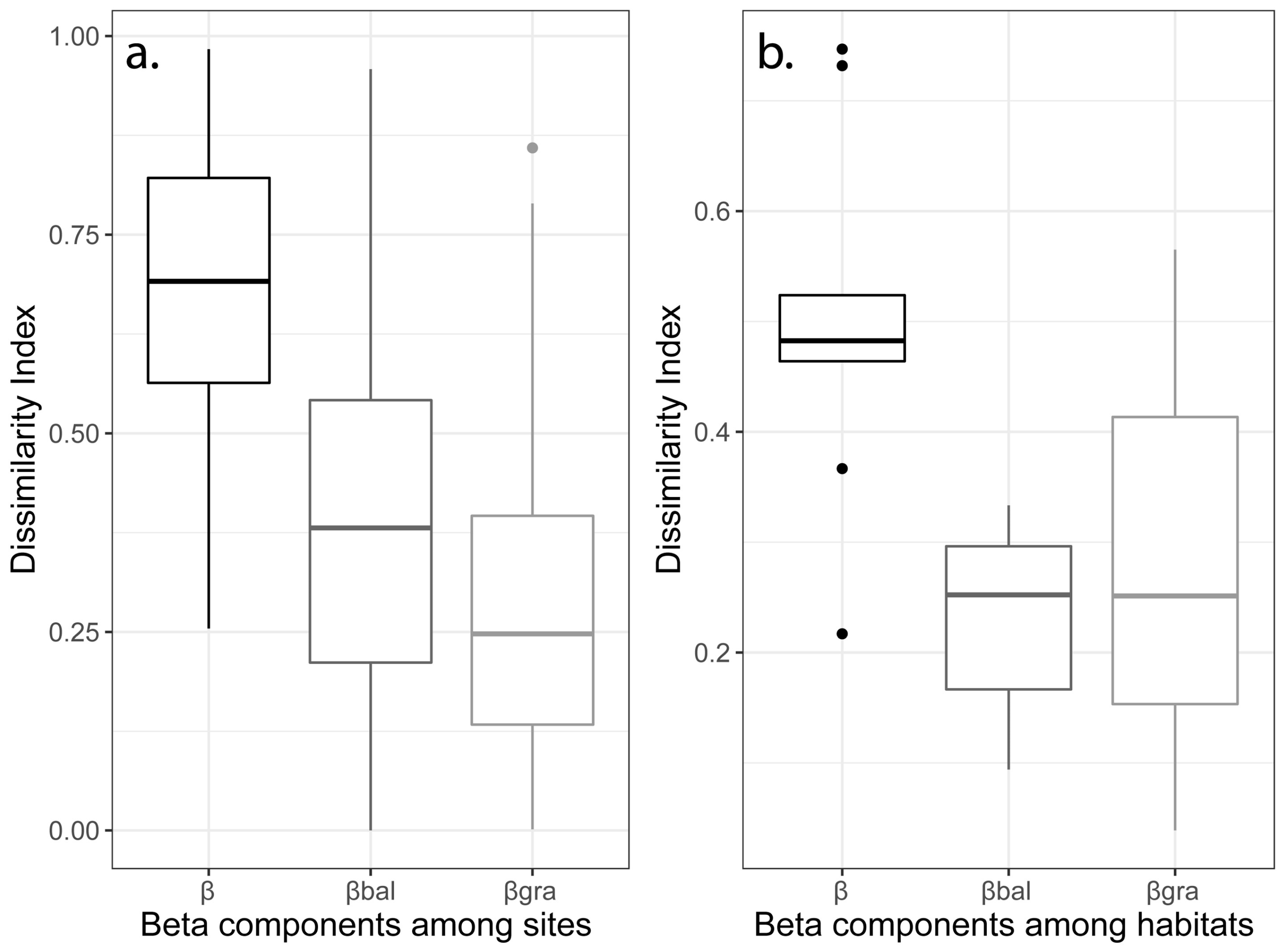
3.3. Beta Diversity and Its Partitions
4. Discussion
4.1. Species Richness and Conservation Value of Mt Mitsikeli
4.2. Environmental Factors Shaping Community Composition
4.3. Beta Diversity and the Role of Environmental Variables
4.4. Spatial Dynamics of Beta Diversity
4.5. Conservation Implications
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat Type | Habitat Code | Habitat Name | Cover (%) |
|---|---|---|---|
| Mountain grasslands | 4090 | Endemic oro-Mediterranean heath with gorse | 35 |
| 5210 | Arborescent matorral with Juniperus spp. | ||
| Mediterranean scrub | 5330 | Thermo-Mediterranean and pre-desert scrub | 25 |
| Mixed thermophilous forest | 9280 | Quercus frainetto woods | 23 |
| 91M0 | Pannonian balkanic turkey oak–sessile oak forest | ||
| 9250 | Forest of Carpinus orientalis and Ostrya carpinifolia and mixed thermophilous forest | ||
| Beech–fir forest | 9150 | Medio-European limestone beech forest of the Cephalanthero-Fagion | 16 |
| 9270 | Hellenic beech forest with Abies borisii-regis | ||
| Agricultural land | 1050 | Pure non-irrigated arable land | 1 |
| 1051 | Mixed non-irrigable arable land | ||
| 1056 | Pure permanently irrigated land | ||
| 1057 | Mixed permanently irrigated land |
Appendix B
| Plot | Habitat Type | Ground Coverage (%) | Vegetation | Grazing | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| A (m) | H | W | St | G | So | Gh (cm) | F | I | ||
| A1a | Agricultural land (A) | 973 | 1 | 0 | 15 | 75 | 10 | 70 | 40 | 2 |
| A1b | 984 | 1 | 0 | 25 | 60 | 15 | 40 | 0 | 2 | |
| A2a | 1039 | 2 | 15 | 0 | 85 | 0 | 35 | 0 | 1 | |
| A2b | 1059 | 2 | 10 | 0 | 90 | 0 | 45 | 0 | 2 | |
| A3a | 1180 | 1 | 30 | 25 | 20 | 20 | 45 | 5 | 2 | |
| A3b | 1175 | 1 | 0 | 20 | 60 | 20 | 15 | 0 | 3 | |
| F1a | Beech–fir forest (F) | 1327 | 1 | 5 | 20 | 65 | 10 | 55 | 0 | 1 |
| F1b | 1324 | 1 | 0 | 10 | 90 | 0 | 50 | 0 | 1 | |
| F2a | 1314 | 1 | 15 | 20 | 50 | 15 | 35 | 0 | 1 | |
| F2b | 1307 | 2 | 50 | 25 | 15 | 10 | 30 | 0 | 1 | |
| F3a | 1139 | 2 | 0 | 0 | 90 | 10 | 15 | 0 | 2 | |
| F3b | 1146 | 2 | 5 | 5 | 85 | 5 | 25 | 25 | 2 | |
| G1a | Mountain grasslands (G) | 1272 | 1 | 20 | 10 | 70 | 0 | 15 | 0 | 2 |
| G1b | 1273 | 1 | 0 | 15 | 85 | 0 | 20 | 0 | 2 | |
| G2a | 1380 | 1 | 0 | 0 | 90 | 10 | 30 | 0 | 1 | |
| G2b | 1366 | 1 | 0 | 0 | 90 | 10 | 30 | 0 | 1 | |
| G3a | 1344 | 1 | 25 | 5 | 50 | 20 | 10 | 0 | 3 | |
| G3b | 1330 | 2 | 0 | 5 | 85 | 10 | 30 | 0 | 2 | |
| M1a | Mixed thermophilous forest (M) | 1071 | 2 | 45 | 15 | 40 | 0 | 30 | 20 | 1 |
| M1b | 1070 | 2 | 20 | 10 | 55 | 15 | 30 | 0 | 1 | |
| M2a | 1000 | 3 | 15 | 0 | 85 | 0 | 60 | 150 | 1 | |
| M2b | 1007 | 3 | 20 | 5 | 65 | 10 | 25 | 300 | 1 | |
| M3a | 1035 | 1 | 20 | 10 | 70 | 0 | 45 | 0 | 1 | |
| M3b | 1057 | 1 | 30 | 0 | 65 | 5 | 40 | 0 | 1 | |
| S1a | Mediterranean scrub (S) | 1178 | 1 | 20 | 30 | 40 | 10 | 30 | 0 | 2 |
| S1b | 1170 | 1 | 0 | 40 | 45 | 15 | 15 | 25 | 2 | |
| S2a | 934 | 1 | 15 | 40 | 25 | 20 | 30 | 0 | 2 | |
| S2b | 932 | 1 | 0 | 40 | 40 | 20 | 20 | 0 | 2 | |
| S3a | 896 | 2 | 0 | 10 | 75 | 15 | 20 | 15 | 1 | |
| S3b | 830 | 2 | 15 | 40 | 30 | 15 | 15 | 0 | 1 | |
Appendix C
| Site level (100 × 100 m) | |||||
| Bray–Curtis dissimilarity index (βbc) | df | S.S. | R2 | F | p |
| Tree cover (>2 m) | 1 | 1.01 | 0.13 | 4.31 | 0.002 |
| Woody vegetation cover | 1 | 0.23 | 0.03 | 0.98 | 0.477 |
| High shrub cover (0.5–2 m) | 1 | 0.34 | 0.04 | 1.44 | 0.158 |
| Low shrub cover (<0.5 m) | 1 | 0.28 | 0.04 | 1.07 | 0.366 |
| Abundance gradient (βbc.gra) | |||||
| Tree cover (>2 m) | 1 | 0.39 | 0.21 | 8.06 | 0.007 |
| Woody vegetation cover | 1 | 0.07 | 0.04 | 1.51 | 0.302 |
| Cover of tall shrubs (0.5–2 m) | 1 | 0.12 | 0.07 | 2.55 | 0.127 |
| Cover of short shrubs (<0.5 m) | 1 | 0.12 | 0.07 | 1.99 | 0.224 |
| Balanced variation in abundance (βbc.bal) | |||||
| Tree cover (>2 m) | 1 | 0.31 | 0.09 | 2.64 | 0.064 |
| Woody vegetation cover | 1 | −0.03 | −0.01 | −0.27 | 0.951 |
| High shrub cover (0.5–2 m) | 1 | 0.12 | 0.03 | 1.00 | 0.443 |
| Low shrub cover (<0.5 m) | 1 | 0.10 | 0.03 | 0.82 | 0.545 |
| Plot level (10 × 10 m) | |||||
| Bray–Curtis dissimilarity index (βbc) | df | S.S. | R2 | F | p |
| Woody vegetation cover | 1 | 0.39 | 0.05 | 1.75 | 0.083 |
| Rockiness | 1 | 0.34 | 0.04 | 1.54 | 0.130 |
| Grass/herb cover | 1 | 0.63 | 0.08 | 2.84 | 0.004 |
| Bare ground | 1 | 0.26 | 0.03 | 1.17 | 0.310 |
| Grass/herb height | 1 | 0.30 | 0.04 | 1.36 | 0.216 |
| Flower heads | 1 | 0.26 | 0.03 | 1.15 | 0.321 |
| Elevation | 1 | 0.56 | 0.07 | 2.49 | 0.013 |
| Abundance gradient (βbc.gra) | |||||
| Woody vegetation cover | 1 | 0.23 | 0.13 | 4.09 | 0.047 |
| Rockiness | 1 | −0.05 | −0.03 | −0.85 | 0.958 |
| Grass/herb cover | 1 | 0.06 | 0.03 | 1.06 | 0.366 |
| Bare ground | 1 | 0.20 | 0.11 | 3.56 | 0.075 |
| Grass/herb height | 1 | 0.08 | 0.05 | 1.50 | 0.295 |
| Flower heads | 1 | 0.09 | 0.05 | 1.53 | 0.269 |
| Elevation | 1 | −0.01 | −0.01 | −0.18 | 0.834 |
| Balanced variation in abundance (βbc.bal) | |||||
| Woody vegetation cover | 1 | 0.10 | 0.03 | 1.14 | 0.387 |
| Rockiness | 1 | 0.25 | 0.07 | 2.72 | 0.056 |
| Grass/herb cover | 1 | 0.31 | 0.09 | 3.38 | 0.037 |
| Bare ground | 1 | −0.02 | −0.01 | −0.27 | 0.958 |
| Grass/herb height | 1 | 0.07 | 0.02 | 0.73 | 0.567 |
| Flower heads | 1 | 0.09 | 0.03 | 1.01 | 0.394 |
| Elevation | 1 | 0.65 | 0.19 | 7.17 | 0.001 |
References
- Fartmann, T.; Krämer, B.; Stelzner, F.; Poniatowski, D. Orthoptera as ecological indicators for succession in steppe grassland. Ecol. Indic. 2012, 20, 337–344. [Google Scholar] [CrossRef]
- Gandar, M. The dynamics and trophic ecology of grasshoppers (Acridoidea) in a South African savanna. Trophic ecology of grasshoppers in South African savanna. Oecologia 1982, 54, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Ryszkowski, L.; Karg, J.; Margarit, G.; Paoletti, M.; Zlotin, R. Above-ground insect biomass in agricultural landscapes of Europe. Landsc. Ecol. Agroecosystems 1993, 71–82. [Google Scholar]
- Samways, M. Conservation biology of Orthoptera. Bionomics of Grasshoppers, Katydids, and Their Kin; CABI: Oxford, UK, 1997; pp. 481–496. [Google Scholar]
- Belovsky, G.E.; Slade, J. The role of vertebrate and invertebrate predators in a grasshopper community. Oikos 1993, 68, 193–201. [Google Scholar] [CrossRef]
- Parr, S.J.; Naveso, M.Á.; Yarar, M. Habitat and potential prey surrounding lesser kestrel Falco naumanni colonies in central Turkey. Biol. Conserv. 1997, 79, 309–312. [Google Scholar] [CrossRef]
- Curry, J.P. Grassland Invertebrates: Ecology, Influence on Soil Fertility and Effects on Plant Growth; Springer Science & Business Media: Cham, Switzerland, 1993; p. 446. [Google Scholar]
- Aleksanov, V.V.; Karmazina, I.O.; Ruchin, A.B.; Esin, M.N.; Lukiyanov, S.V.; Lobachev, E.A.; Artaev, O.N.; Ryzhov, M.K. Diversity and Biology of Terrestrial Orthopteroids (Insecta) in the Republic of Mordovia (Russia). Diversity 2023, 15, 803. [Google Scholar] [CrossRef]
- Báldi, A.; Kisbenedek, T. Orthopteran assemblages as indicators of grassland naturalness in Hungary. Agric. Ecosyst. Environ. 1997, 66, 121–129. [Google Scholar] [CrossRef]
- Kruess, A.; Tscharntke, T. Grazing intensity and the diversity of grasshoppers, butterflies, and trap-nesting bees and wasps. Conserv. Biol. 2002, 16, 1570–1580. [Google Scholar] [CrossRef]
- Marini, L.; Bommarco, R.; Fontana, P.; Battisti, A. Disentangling effects of habitat diversity and area on orthopteran species with contrasting mobility. Biol. Conserv. 2010, 143, 2164–2171. [Google Scholar] [CrossRef]
- Marini, L.; Fontana, P.; Battisti, A.; Gaston, K.J. Response of orthopteran diversity to abandonment of semi-natural meadows. Agric. Ecosyst. Environ. 2009, 132, 232–236. [Google Scholar] [CrossRef]
- Schirmel, J.; Mantilla-Contreras, J.; Blindow, I.; Fartmann, T. Impacts of succession and grass encroachment on heathland Orthoptera. J. Insect Conserv. 2011, 15, 633–642. [Google Scholar] [CrossRef]
- Gardiner, T.; Dover, J. Is microclimate important for Orthoptera in open landscapes? J. Insect Conserv. 2008, 12, 705–709. [Google Scholar] [CrossRef]
- Kati, V.; Dufrêne, M.; Legakis, A.; Grill, A.; Lebrun, P. Conservation management for Orthoptera in the Dadia reserve, Greece. Biol. Conserv. 2004, 115, 33–44. [Google Scholar] [CrossRef]
- Theuerkauf, J.; Rouys, S. Do Orthoptera need human land use in Central Europe? The role of habitat patch size and linear corridors in the Białowieża Forest, Poland. Biodivers. Conserv. 2006, 15, 1497–1508. [Google Scholar] [CrossRef]
- Bazelet, C.S.; Samways, M.J. Grasshopper assemblage response to conservation ecological networks in a timber plantation matrix. Agric. Ecosyst. Environ. 2011, 144, 124–129. [Google Scholar] [CrossRef]
- Bazelet, C.S.; Samways, M.J. Identifying grasshopper bioindicators for habitat quality assessment of ecological networks. Ecol. Indic. 2011, 11, 1259–1269. [Google Scholar] [CrossRef]
- Kovács-Hostyánszki, A.; Kőrösi, Á.; Orci, K.M.; Batáry, P.; Báldi, A. Set-aside promotes insect and plant diversity in a Central European country. Agric. Ecosyst. Environ. 2011, 141, 296–301. [Google Scholar] [CrossRef]
- Marini, L.; Fontana, P.; Scotton, M.; Klimek, S. Vascular plant and Orthoptera diversity in relation to grassland management and landscape composition in the European Alps. J. Appl. Ecol. 2008, 45, 361–370. [Google Scholar] [CrossRef]
- Steck, C.E.; Bürgi, M.; Bolliger, J.; Kienast, F.; Lehmann, A.; Gonseth, Y. Conservation of grasshopper diversity in a changing environment. Biol. Conserv. 2007, 138, 360–370. [Google Scholar] [CrossRef]
- Kenyeres, Z.; Rácz, I.A.; Varga, Z. Endemism hot spots, core areas and disjunctions in European Orthoptera. Acta Zoologica Cracoviensia-Series B: Invertebrata. 2009, 52, 189–211. [Google Scholar] [CrossRef]
- Hochkirch, A.; Nieto, A.; Criado, M. European Red List of Grasshoppers, Crickets and Bush-Crickets; Publications Office of the European Union: Luxembourg, 2016; p. 95. [Google Scholar]
- Kati, V.; Willemse, L. Database of Endemic and/or Threatened Orthoptera Species in the Natura 2000 Sites of Greece; V1; Mendeley Data: Amsterdam, The Netherlands, 2020. [Google Scholar] [CrossRef]
- Willemse, L.; Kleukers, R.; Odé, B. The Grasshoppers of Greece; EIS Kenniscentrum Insecten: Leiden, The Netherlands, 2018; p. 440. [Google Scholar]
- Kati, V.; Zografou, K.; Tzirkalli, E.; Chitos, T.; Willemse, L. Butterfly and grasshopper diversity patterns in humid Mediterranean grasslands: The roles of disturbance and environmental factors. J. Insect Conserv. 2012, 16, 807–818. [Google Scholar] [CrossRef]
- Zografou, K.; Sfenthourakis, S.; Pullin, A.; Kati, V. On the surrogate value of red-listed butterflies for butterflies and grasshoppers: A case study in Grammos site of Natura 2000, Greece. J. Insect Conserv. 2009, 13, 505–514. [Google Scholar] [CrossRef]
- Zografou, K.; Wilson, R.J.; Halley, J.M.; Tzirkalli, E.; Kati, V. How are arthopod communities structured and why are they so diverse? Answers from Mediterranean mountains using hierarchical additive partitioning. Biodivers. Conserv. 2017, 26, 1333–1351. [Google Scholar] [CrossRef]
- Legendre, P.; Borcard, D.; Peres-Neto, P.R. Analyzing beta diversity: Partitioning the spatial variation of community composition data. Ecol. Monogr. 2005, 75, 435–450. [Google Scholar] [CrossRef]
- Legendre, P.; De Cáceres, M. Beta diversity as the variance of community data: Dissimilarity coefficients and partitioning. Ecol. Lett. 2013, 16, 951–963. [Google Scholar] [CrossRef] [PubMed]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning abundance-based multiple-site dissimilarity into components: Balanced variation in abundance and abundance gradients. Methods Ecol. Evol. 2017, 8, 799–808. [Google Scholar] [CrossRef]
- Podani, J.; Schmera, D. A new conceptual and methodological framework for exploring and explaining pattern in presence—Absence data. Oikos 2011, 120, 1625–1638. [Google Scholar] [CrossRef]
- Baselga, A. The relationship between species replacement, dissimilarity derived from nestedness, and nestedness. Glob. Ecol. Biogeogr. 2012, 21, 1223–1232. [Google Scholar] [CrossRef]
- Pärtel, M.; Bennett, J.A.; Zobel, M. Macroecology of biodiversity: Disentangling local and regional effects. New Phytol. 2016, 211, 404–410. [Google Scholar] [CrossRef]
- Moreno, C.E.; Calderón-Patrón, J.M.; Arroyo-Rodríguez, V.; Barragán, F.; Escobar, F.; Gómez-Ortiz, Y.; Martín-Regalado, N.; Martínez-Falcón, A.P.; Martínez-Morales, M.Á.; Mendoza, E.; et al. Measuring biodiversity in the Anthropocene: A simple guide to helpful methods. Biodivers. Conserv. 2017, 26, 2993–2998. [Google Scholar] [CrossRef]
- Pitta, E.; Zografou, K.; Poursanidis, D.; Chatzaki, M. Effects of climate on spider beta diversity across different Mediterranean habitat types. Biodivers. Conserv. 2019, 28, 3971–3988. [Google Scholar] [CrossRef]
- Nekola, J.C.; White, P.S. The distance decay of similarity in biogeography and ecology. J. Biogeogr. 1999, 26, 867–878. [Google Scholar] [CrossRef]
- Grzywacz, B.; Heller, K.-G.; Chobanov, D.P.; Warchałowska-Śliwa, E. Conventional and molecular chromosome study in the European genus Parnassiana Zeuner, 1941 (Orthoptera, Tettigoniinae, Platycleidini). Folia Biol. 2017, 65, 1–8. [Google Scholar] [CrossRef]
- Pamperis, L.N. The Butterflies of Greece; Editions Pamperis; Hellenic Society for the Protection of Nature: Athens, Greece, 2009; p. 766. [Google Scholar]
- Pamperis, L. The Butterflies of Greece; An update of distribution maps, plates and diagrams 3.3, in Map 3.4, in Chart 4.15, and in Chart 4.16. Editions Pamperis; Hellenic Society for the Protection of Nature: Athens, Greece, 2022. [Google Scholar]
- Guido, M.; Gianelle, D. Distribution patterns of four Orthoptera species in relation to microhabitat heterogeneity in an ecotonal area. Acta Oecologica 2001, 22, 175–185. [Google Scholar] [CrossRef]
- Malfatti, E.; Ferreira, P.M.A.; Utz, L.R.P. Eukaryotic Communities in Bromeliad Phytotelmata: How Do They Respond to Altitudinal Differences? Diversity 2020, 12, 326. [Google Scholar] [CrossRef]
- Gerasimidis, A.; Korakis, G. Contribution to the study of the flora of Mount Mitsikeli, NW Greece. Fl Medit 2009, 19, 161–184. [Google Scholar]
- NOA. National Observatory of Athens. Available online: https://www.meteo.gr/index.cfm (accessed on 20 November 2023).
- Soulis, N. The Climate of Ipiros (In Greek). Ioannina. Greece. 1994, 216. [Google Scholar]
- Sarika, M.; Christopoulou, A.; Zikos, A.; Kazanis, D.; Bazos, I. Plant communities and habitat types in the protected area of Lake Pamvotis (Epirus, Northwestern Greece). Mediterr. Bot. 2020, 41, 129–148. [Google Scholar] [CrossRef]
- Mountrakis, D. The Geology of Greece; University Press Studio: Thessaloniki, Greece, 1985; p. 208. (In Greek) [Google Scholar]
- EIONET. Report on Progress and Implementation (Article 17-Habitats Directive). 2019. Available online: https://cdr.eionet.europa.eu/gr/eu/art17/envxrm90g/ (accessed on 20 November 2023).
- Petridou, M.; Benson, J.F.; Gimenez, O.; Kati, V. Spatiotemporal Patterns of Wolves, and Sympatric Predators and Prey Relative to Human Disturbance in Northwestern Greece. Diversity 2023, 15, 184. [Google Scholar] [CrossRef]
- Willemse, F. Fauna Graeciae Ⅰ: Catalogue of the Orthoptera of Greece. Hellenic Zoological Society. 1984, 275. [Google Scholar]
- Turker, A.U.; Gurel, E. Common mullein (Verbascum thapsus L.): Recent advances in research. Phytother. Res. 2005, 19, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Magurran, A.E. Measuring Biological Diversity; Wiley: Hoboken, NJ, USA, 2013. [Google Scholar]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Ter Braak, C.; Šmilauer, P. Canoco 5: Software for Multivariate Data Exploration, Testing and Summarization; (Version 5.0); Microcomputer Power: Ithaca, NY, USA, 2012. [Google Scholar]
- Baselga, A. Separating the two components of abundance-based dissimilarity: Balanced changes in abundance vs. abundance gradients. Methods Ecol. Evol. 2013, 4, 552–557. [Google Scholar] [CrossRef]
- Keil, P.; Schweiger, O.; Kühn, I.; Kunin, W.E.; Kuussaari, M.; Settele, J.; Henle, K.; Brotons, L.; Pe’er, G.; Lengyel, S. Patterns of beta diversity in Europe: The role of climate, land cover and distance across scales. J. Biogeogr. 2012, 39, 1473–1486. [Google Scholar] [CrossRef]
- Dinno, A. dunn.test: Dunn’s Test of Multiple Comparisons Using Rank Sums. R Package Version 1.3.5. 2017. Available online: https://CRAN.R-project.org/package=dunn.test (accessed on 15 March 2022).
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. Vegan: Community Ecology Package, R Package Version 2.6-4. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 15 March 2022).
- Borg, I.; Groenen, P. Modern Multidimensional Scaling. Theory and Applications; Springer: Cham, Switzerland, 1997. [Google Scholar]
- Gómez-Rodríguez, C.; Baselga, A. Variation among European beetle taxa in patterns of distance decay of similarity suggests a major role of dispersal processes. Ecography 2018, 41, 1825–1834. [Google Scholar] [CrossRef]
- Kati, V.; Poirazidis, K.; Dufrêne, M.; Halley, J.M.; Korakis, G.; Schindler, S.; Dimopoulos, P. Towards the use of ecological heterogeneity to design reserve networks: A case study from Dadia National Park, Greece. Biodivers. Conserv. 2010, 19, 1585–1597. [Google Scholar] [CrossRef]
- Tzirkalli, E.; Kadis, C.; Halley, J.M.; Vogiatzakis, I.; Wilson, R.J.; Zografou, K.; Antoniou, A.; Tsintides, T.; Makris, C.; Kati, V. Conservation ecology of butterflies on Cyprus in the context of Natura 2000. Biodivers. Conserv. 2019, 28, 1759–1782. [Google Scholar] [CrossRef]
- Neilly, H.; Jones, H.; Schwarzkopf, L. Ants drive invertebrate community response to cattle grazing. Agric. Ecosyst. Environ. 2020, 290, 106742. [Google Scholar] [CrossRef]
- Schaffers, A.P.; Raemakers, I.P.; Sýkora, K.V.; ter Braak, C.J.F. Arthropod Assemblages are Best Predicted by Plant Species Composition. Ecology 2008, 89, 782–794. [Google Scholar] [CrossRef] [PubMed]
- Willemse, L.; Tilmans, J.; Kotitsa, N.; Trichas, A.; Heller, K.G.; Chobanov, D.; Odé, B. A review of Eupholidoptera (Orthoptera, Tettigoniidae) from Crete, Gavdos, Gavdopoula, and Andikithira. ZooKeys 2023, 1151, 67–158. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, T.; Pye, M.; Field, R.; Hill, J. The Influence of Sward Height and Vegetation Composition in Determining the Habitat Preferences of Three Chorthippus Species (Orthoptera: Acrididae) in Chelmsford, Essex, UK. J. Orthoptera Res. 2002, 11, 207–213. [Google Scholar] [CrossRef]
- Theron, K.J.; Pryke, J.S.; Samways, M.J. Identifying managerial legacies within conservation corridors using remote sensing and grasshoppers as bioindicators. Ecol. Appl. 2022, 32, e02496. [Google Scholar] [CrossRef] [PubMed]
- Clark, E.J. Studies in the Ecology of British Grasshoppers. Trans. R. Entomol. Soc. Lond. 1948, 99, 173–222. [Google Scholar] [CrossRef]
- Wingerden, W.v.; Kreveld, A.v.; Bongers, W. Analysis of species composition and abundance of grasshoppers (Orth., Acrididae) in natural and fertilized grasslands. J. Appl. Entomol. 1992, 113, 138–152. [Google Scholar] [CrossRef]
- Humbert, J.-Y.; Delley, S.; Arlettaz, R. Grassland intensification dramatically impacts grasshoppers: Experimental evidence for direct and indirect effects of fertilisation and irrigation. Agric. Ecosyst. Environ. 2021, 314, 107412. [Google Scholar] [CrossRef]
- Gardiner, T.; Hassall, M. Does microclimate affect grasshopper populations after cutting of hay in improved grassland? J. Insect Conserv. 2009, 13, 97–102. [Google Scholar] [CrossRef]
- Palmer, M.W. Species-area curves and the geometry of nature. In Scaling Biodiversity, Ecological Reviews; Cambridge University Press: Cambridge, UK, 2007; pp. 15–31. [Google Scholar] [CrossRef]
- Ibanez, S.; Bison, M.; Lavorel, S.; Moretti, M. Herbivore species identity mediates interspecific competition between plants. Community Ecol. 2013, 14, 41–47. [Google Scholar] [CrossRef]
- Ritchie, J.M. Biology of grasshoppers. In Bulletin of Entomological Research; Chapman, R.F., Joern, A., Eds.; Cambridge University Press: Cambridge, UK, 1991; Volume 81, pp. 221–222. [Google Scholar] [CrossRef]
- Cardoso, P.; Barton, P.S.; Birkhofer, K.; Chichorro, F.; Deacon, C.; Fartmann, T.; Fukushima, C.S.; Gaigher, R.; Habel, J.C.; Hallmann, C.A.; et al. Scientists’ warning to humanity on insect extinctions. Biol. Conserv. 2020, 242, 108426. [Google Scholar] [CrossRef]
- Chase, J.M.; Leibold, M.A. Ecological Niches: Linking Classical and Contemporary Approaches; University of Chicago Press: Chicago, Ilinois, USA, 2009. [Google Scholar]
- Cottenie, K. Integrating environmental and spatial processes in ecological community dynamics. Ecol. Lett. 2005, 8, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, T. Grazing and Orthoptera: A review. J. Orthoptera Res. 2018, 27, 3–11. [Google Scholar] [CrossRef]
- Bascompte, J.; Rodríguez, M.Á. Habitat patchiness and plant species richness. Ecol. Lett. 2001, 4, 417–420. [Google Scholar] [CrossRef]
- Fartmann, T.; Behrens, M.; Loritz, H. Orthopteran communities in the conifer-broadleaved woodland zone of the Russian Far East. Eur. J. Entomol. 2008, 105, 673–680. [Google Scholar] [CrossRef]
- Jauregui, B.M.; Rosa-Garcia, R.; Garcia, U.; WallisDeVries, M.F.; Osoro, K.; Celaya, R. Effects of stocking density and breed of goats on vegetation and grasshopper occurrence in heathlands. Agric. Ecosyst. Environ. 2008, 123, 219–224. [Google Scholar] [CrossRef]
- Bazelet, C.S.; Gardiner, T. Orthoptera response to grazing: An introduction to the special issue. J. Orthoptera Res. 2018, 27. [Google Scholar] [CrossRef]
- Zografou, K.; Adamidis, G.C.; Grill, A.; Kati, V.; Wilson, R.J.; Halley, J.M. Who flies first?—Habitat-specific phenological shifts of butterflies and orthopterans in the light of climate change: A case study from the south-east Mediterranean. Ecol. Entomol. 2015, 40, 562–574. [Google Scholar] [CrossRef]
- Anthelme, F.; Grossi, J.-L.; Brun, J.-J.; Didier, L. Consequences of green alder expansion on vegetation changes and arthropod communities removal in the northern French Alps. For. Ecol. Manag. 2001, 145, 57–65. [Google Scholar] [CrossRef]
- Dennis, P.; Young, M.; Gordon, I. Distribution and abundance of small insects and arachnids in relation to structural heterogeneity of grazed, indigenous grasslands. Ecol. Entomol. 1998, 23, 253–264. [Google Scholar] [CrossRef]
- Condit, R.; Pitman, N.; Leigh, E.G.; Chave, J.; Terborgh, J.; Foster, R.B.; Núñez, P.; Aguilar, S.; Valencia, R.; Villa, G.; et al. Beta-Diversity in Tropical Forest Trees. Science 2002, 295, 666–669. [Google Scholar] [CrossRef]
- Hubbell, S.P. A Unified Theory of Biodiversity and Biogeography; Princeton University Press: Princeton, NJ, USA, 2001. [Google Scholar]
- SDF. Natura 2000-Standard Data Form. GR2130008 Oros Mitsikeli. Available online: https://natura2000.eea.europa.eu/natura2000/SDF.aspx?site=GR2130008 (accessed on 1 December 2023).
- Slancarova, J.; Bartonova, A.; Zapletal, M.; Kotilinek, M.; Faltynek Fric, Z.; Micevski, N.; Kati, V.; Konvicka, M. Life History Traits Reflect Changes in Mediterranean Butterfly Communities Due to Forest Encroachment. PLoS ONE 2016, 11, e0152026. [Google Scholar] [CrossRef] [PubMed]
- Zakkak, S.; Chatzaki, M.; Karamalis, N.; Kati, V. Spiders in the context of agricultural land abandonment in Greek Mountains: Species responses, community structure and the need to preserve traditional agricultural landscapes. J. Insect Conserv. 2014, 18, 599–611. [Google Scholar] [CrossRef]
- Zakkak, S.; Kakalis, E.; Radović, A.; Halley, J.M.; Kati, V. The impact of forest encroachment after agricultural land abandonment on passerine bird communities: The case of Greece. J. Nat. Conserv. 2014, 22, 157–165. [Google Scholar] [CrossRef]
- Zakkak, S.; Radovic, A.; Panitsa, M.; Vassilev, K.; Shuka, L.; Kuttner, M.; Schindler, S.; Kati, V. Vegetation patterns along agricultural land abandonment in the Balkans. J. Veg. Sci. 2018, 29, 877–886. [Google Scholar] [CrossRef]

| Abbreviation | CAELIFERA ACRIDIDAE | Abbreviation | ENSIFERA TETTIGONIIDAE |
|---|---|---|---|
| Aunga | Acrida ungarica + | Dalbi | Decticus albifrons |
| Astre | Aiolopus streppens + | Dverr | Decticus verrucivorus |
| Amicr | Arcyptera microptera | Eschm | Eupholidoptera schmidti |
| Cital | Calliptamus italicus | Lpunc | Leptophyes punctatissima |
| Cborn | Chorthippus bornhalmi | Pfemo | Pholidoptera femorata |
| Cdich | Chorthippus dichrous | Paffi | Platycleis affinis |
| Cmoll | Chorthippus mollis | Palbo | Platycleis albopunctata |
| Dbrev | Dociostaurus brevicollis | Pinte | Platycleis intermedia |
| Edecl | Euchorthippus declivus | Pjoni | Poecilimon jonicus |
| Lmigr | Locusta migratoria + | Rgerm | Rhacocleis germanica |
| Ocaer | Oedipoda caerulescens | Shell | Saga hellenica + |
| Ogerm | Oedipoda germanica | Ssepi | Sepiana sepium |
| Orufi | Omocestus rufipes | Torin | Tessellana orina |
| Pcalo | Paracaloptenus caloptenoides *+ | Tviri | Tettigonia viridissima |
| Pgior | Pezotettix giornae | Tlili | Tylopsis lilifolia |
| Ppara | Pseudochorthippus parallelus | Voblo | Vichetia oblongicollis + |
| Sfisc | Stenobothrus fischeri | GRYLLIDAE | |
| Gcamp | Gryllus campestris |
| Parameter | Agricultural Land | Beech–Fir Forest | Mountain Grasslands | Mixed Thermophilous Forest | Mediterranean Scrub | |
|---|---|---|---|---|---|---|
| Site level (Av. Val.) | T | 16.7 | 80 | 16.7 | 43.3 | 33.3 |
| Hs | 25 | 8.3 | 18.3 | 13.3 | 15.7 | |
| Ls | 6.7 | 5 | 1.7 | 6.7 | 1.7 | |
| Ws | 48.3 | 93.3 | 36.7 | 63.3 | 50.7 | |
| Plot level (Av. Val.) | A | 1068.3 | 1259.5 | 1327.5 | 1040 | 990 |
| H | 1.3 | 1.5 | 1.2 | 2 | 1.3 | |
| W | 9.2 | 12.5 | 7.5 | 25 | 8.3 | |
| St | 14.2 | 13.3 | 6.7 | 6.7 | 33.3 | |
| G | 56 | 66.7 | 78.3 | 63.3 | 43.3 | |
| So | 10 | 7.5 | 8.3 | 5 | 16.7 | |
| Gh | 41.7 | 35 | 22.5 | 38.3 | 21.7 | |
| F | 7.5 | 4.2 | 0 | 78.3 | 6.7 | |
| I | 2 | 1.3 | 1.8 | 1 | 1.7 | |
| Diversity indices | S | 15 | 18 | 11 | 15 | 14 |
| E | 6 | 8 | 6 | 6 | 3 | |
| Smean | 6.33 ± 1.75 | 7.67 ± 2.07 | 6.17 ± 0.98 | 5.33 ± 2.58 | 6.17 ± 1.17 | |
| H | 1.9 | 2.3 | 1.7 | 1.9 | 2.2 | |
| Hmean | 1.54 ± 0.20 | 1.94 ± 0.15 | 1.48 ± 0.08 | 1.53 ± 0.30 | 1.79 ± 0.06 | |
| J | 0.7 | 0.8 | 0.7 | 0.7 | 0.8 | |
| N | 456 | 216 | 1126 | 415 | 264 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stefanidis, A.; Zografou, K.; Tzortzakaki, O.; Kati, V. Orthoptera Community Dynamics and Conservation in a Natura 2000 Site (Greece): The Role of Beta Diversity. Diversity 2024, 16, 11. https://doi.org/10.3390/d16010011
Stefanidis A, Zografou K, Tzortzakaki O, Kati V. Orthoptera Community Dynamics and Conservation in a Natura 2000 Site (Greece): The Role of Beta Diversity. Diversity. 2024; 16(1):11. https://doi.org/10.3390/d16010011
Chicago/Turabian StyleStefanidis, Apostolis, Konstantina Zografou, Olga Tzortzakaki, and Vassiliki Kati. 2024. "Orthoptera Community Dynamics and Conservation in a Natura 2000 Site (Greece): The Role of Beta Diversity" Diversity 16, no. 1: 11. https://doi.org/10.3390/d16010011
APA StyleStefanidis, A., Zografou, K., Tzortzakaki, O., & Kati, V. (2024). Orthoptera Community Dynamics and Conservation in a Natura 2000 Site (Greece): The Role of Beta Diversity. Diversity, 16(1), 11. https://doi.org/10.3390/d16010011