Abstract
While native grass and shrub restoration is often promoted for bird conservation, evaluations are necessary to understand the effects of habitat change on songbird vital rates and migratory stopover use. We evaluated the effects of native vegetation restoration on the species richness, productivity, and capture rate of breeding and fall migrating songbirds at the Shaker Village of Pleasant Hill, in central Kentucky, USA. From 2009 to 2022, we conducted constant effort mist netting, resulting in 8418 captures of 110 bird species. In 2012–2013, we implemented management in the study area, which began as an old field, to restore native grasses, forbs, and shrubs. Effects on the vital rates of breeding species included higher Spizella pusilla (field sparrow) and Spinus tristis (American goldfinch) productivity post restoration. Restoration of native vegetation also led to significantly higher fall capture rates for pooled cohorts of both long-distance and medium-distance migrants. Our results demonstrate that the restoration and management of native vegetation can enhance stopover habitats for fall migrating songbirds including neotropical migrants and the field sparrow, a common bird in steep decline. We recommend prioritizing areas for native grass and shrub restoration that initially lack structural heterogeneity (a well-developed grass component with diverse forbs and interspersed shrubs).
1. Introduction
Grassland and shrubland birds are declining in eastern North America [1,2,3]. Many grasslands and shrublands contain exotic flora, and native vegetation restoration is often recommended to improve the habitat quality for songbirds [4,5]. Initiatives to restore native vegetation in grassland and early successional landcover types have gained momentum in recent decades in the USA through multistate partnerships and federal cost-share programs funded through the U.S. Farm Bill [6,7,8]. However, restorations are costly and if benefiting bird populations is the primary goal, evaluations are necessary to understand the effects of vegetation manipulations on breeding and migrating birds [9,10].
The effects of native vegetation restoration and management recommendations for the North American gamebird, Colinus virginianus (Northern Bobwhite), are well-documented [11,12,13]. The literature also supports the benefits of native vegetation management for North American pollinators including management recommendations for Danaus plexippus (Monarch butterfly) [14,15,16,17]. Landscape-scale auditory surveys to evaluate the effects of federal (U.S.) cost-share conservation practices (including native vegetation restoration) on breeding birds have reported mixed results [18,19,20]. Meanwhile, studies of the non-breeding bird use of managed grassland habitats have primarily focused on winter songbird abundance, rather than migration stopovers [21,22,23].
Prior to our study, reports on the effects of localized native vegetation restoration on the vital rates and migratory stopover use of songbirds in North America have been sparse. However, native grass restoration and invasive plant removal increased the capture rate for several grassland species in Georgia [24]. Furthermore, a study evaluating the fitness of fall migrants stopping over in a site dominated by non-native Lonicera spp. (honeysuckle) in Pennsylvania concluded that the habitat quality was poor at that site for most species [25]. A study of native grassland restoration supported by cost-share conservation programs in Maryland reported that, when properly executed, management benefitted suitable habitats for Ammodramus savannarum (grasshopper sparrow) [26].
Several North American studies have identified the negative effects of exotic shrubs on songbirds and support the impetus to replace exotic shrubs with native species. Borgmann and Rodewald [27] found that Turdus migratorius (American robin) and Cardinalis cardinalis (Northern cardinal) nests in exotic shrub species Lonicera spp. and Rosa multiflora (multi-flora rose) had reduced nesting success and were twice as likely to be depredated. Research by Smith et al. [28] in New York found that the fruits of native shrubs were of greater nutritional value to fall migrating songbirds than the fruits of invasive shrubs. Narango et al. [29] found that exotic plants reduced the habitat suitability for breeding Poecile carolinensis (Carolina chickadees) by reducing insect food abundance. In contrast, one study in New England reported that Mniotilta varia (black-and-white warbler), Gray catbird, Empidonax alnorum (alder flycatcher), and Setophaga petechia (yellow warbler) increased with the cover of non-native woody plants and hesitated to recommend plant species manipulations for shrubland birds [2]. Furthermore, a meta-analysis by Nelson et al. [9] highlighted the mixed results of studies on the effects of invasive plants on birds and recommended that managers eradicating invasive plants to improve bird habitat should monitor the impacts locally to determine the benefits.
Efforts to restore native vegetation in early successional areas occurred within focal areas in Kentucky during 2008–2022, for the most part due to the Kentucky Northern Bobwhite Conservation Initiative and federal (U.S.) cost-share conservation programs [30]. The Kentucky Department of Fish and Wildlife Resources (KDFWR) designated the Shaker Village of Pleasant Hill (SVPH) in Mercer County, Kentucky, as a Quail Focal Area in 2008. Subsequently, the KDFWR and SVPH restored 405 ha of native grassland and shrubland habitat in the area over the next decade [30]. The KDFWR initiated a songbird banding station at Shaker Village during the spring of 2009 prior to the restoration of native vegetation at the site. Songbird banding occurred through 2022 during the nesting and fall migration seasons at this site to collect data before, during, and after typical management actions that occur during native grass and shrub restorations. The primary objective of this project was to evaluate the effects of common management practices for native vegetation restoration on nesting and migrating songbirds. Although we were interested in all landbirds that use grassland/shrubland, we were particularly interested in the Spizella pusilla (field sparrow) because it is a species of greatest conservation need in Kentucky’s State Wildlife Action Plan [31] and a Partners in Flight Common Bird in Steep Decline [4].
2. Materials and Methods
2.1. Study Site
The Shaker Village of Pleasant Hill is a National Historic Landmark in Mercer County, Kentucky, USA (37° 48.977 N, 84° 44.419 W). The study site is situated on the natural area owned and managed by the SVPH and comprised 8.1 ha. It began as an old field containing open areas dominated by non-native cool season grasses with patches of shrubs and a wooded edge (Figures S1 and S2). The site is surrounded by hayfields, pasture, and native warm-season grass (NWSG) fields as well as conventional row-crops (soybeans and corn) and free-range cattle operations.
2.2. Vegetation Management
Mist netting began in 2009 to obtain baseline data prior to the restoration of native vegetation at the study site. In late fall 2012, efforts to convert the area to native vegetation began with the removal of non-native and undesirable trees and shrubs. We removed non-native species Maclura pomifera (Osage orange) and Lonicera maackii (bush honeysuckle) and a prolific native species, Robinia pseudo-acacia (black locust) by using a chainsaw and a skid-steer with a mulcher attachment (Figure S3). We then mowed the area to make planting of NWSG possible. To preserve the vegetative structure for mist netting efforts post-conversion, we spared several small patches of desirable native trees and shrubs, mostly comprised of young Juglans nigra (black walnut), Fraxinus americana (white ash), and Juniperus virginiana (eastern red cedar).
We planted 560 native shrubs and small trees in late March 2013. We targeted shrub plantings in areas where shrub cover was recorded in 2009–2012 to bring back a similar structure to what was there previously. We planted shrubs and trees with mesh tree guards in groves of 25–100 to allow for herbicide treatment and the planting of NWSG in the remainder of the area (Figure S4). Species planted included Cornus drummondii (rough-leaved dogwood), Crataegus phaenopyrum (Washington hawthorn), Prunus americana (American plum), Cercis canadensis (eastern redbud), and Rhus glabra (smooth sumac).
Cool season grasses (CSGs) present at the site pre-restoration included Schedonorus arundinaceus (tall fescue) and Poa pratensis (Kentucky bluegrass). To control CSGs, in April 2013, we employed a licensed herbicide applicator to apply 2.24 kg ha−1 glyphosate and 0.42 kg ha−1 imazapic, followed by 2.24 kg ha−1 glyphosate two weeks later. We added a non-ionic surfactant as per the label instructions for imazapic. NWSG planting took place in May 2013 using a tractor and seed drill (Figures S5 and S6). We planted a native short-grass mixture including Elymus virginicus (Virginia wild rye), Bouteloua curtipendula (sideoats grama), and Schizachyrium scoparium (little bluestem). Forbs included in the planting mix were Rudbeckia hirta (black-eyed Susan), Echinacea purpurea (purple coneflower), Chamaecrista fasciculata (partridge pea), and Heliopsis helianthoides (false sunflower). We backpack-sprayed areas inaccessible to large equipment due to native shrub plots and later hand-seeded grasses in the same areas.
Follow-up management of non-native/invasive shrubs occurred in spring 2014. We used triclopyr with a non-ionic surfactant for the spot control of undesirable woody plants (Osage orange and black locust) to avoid encroachment on native vegetation. We implemented prescribed burning in the area on 27 February 2019 (Figure S7). In April 2021, we disked small portions of the field to promote the growth of annuals and diverse vegetation structure.
2.3. Constant Effort Mist Netting
Monitoring for migratory stopover use is difficult using traditional auditory or visual surveys, but standardized mist netting has been shown to be a useful tool for monitoring populations [32]. We conducted constant effort mist netting during both the nesting and fall migration seasons of 2009–2022. During the nesting season, we followed the most current Monitoring Avian Productivity and Survivorship (MAPS) protocol set forth by the Institute of Bird Populations [33,34]. During fall migration, we used a protocol identical to MAPS, except where noted below.
During the breeding season, we opened 10 mist nets (32 mm mesh) at sunrise for four hours. Mist nets were dispersed throughout the study area and remained in the same locations for the duration of the study. We operated the station on eight dates, at least six days apart, in each breeding season between 20 May and 9 August. For migration banding, we opened nets 30 min prior to sunrise and operated for four hours. During the migration season, we operated the station on seven to nine dates, at least six days apart between August 30 and November 3. We checked the nets every hour, except during July and August, when we checked the nets every 40 min to avoid heat stress in the birds. We did not operate during wind speeds greater than 23 km per hour or during rain. On occasion, the nets were closed early due to inclement weather. In such cases, we carefully recorded the effort data for use in the computation of the capture rate.
We banded all captured birds, except for hummingbirds (Archilochus spp.), with U.S. Geological Survey aluminum leg bands. We recorded the morphological measurements and plumage characteristics including breeding condition, wing chord, mass, fat, extent of flight feather and body molt, and degree of feather wear. When possible, we aged and sexed birds using plumage characteristics (including the presence or absence of molt limits) and/or the degree of skull pneumatization [35].
2.4. Vegetation Monitoring
We measured vegetation variables at the study area on three occasions, three to four years apart, at 13 points determined by a randomly placed 75 m grid. Monitoring occurred in fall (September–October), and eight variables were measured or recorded at each point. We determined the dominant stand size class of vegetation within 15 m of the plot center by choosing one of the following categories: grassland (a field with little to no woody component); seedling (stands dominated by trees between 2.5 and 7.6 cm diameter at breast height [DBH]); sapling (stands dominated by trees between 7.6 and 12.7 cm DBH); poletimber (stands dominated by trees between 12.7 and 27.9 cm DBH); or sawtimber (stands dominated by trees >28 cm DBH) [36].
We measured the basal area using a 10-factor prism in plots and recorded the dominant tree species (any trees larger than 12.7 cm DBH occurring within 15 m of the point center) by listing the three most common species present. We tallied the snag presence by recording the number of snags within 15 m of the point and measured the small stem density by walking with arms outstretched from the plot center 15 m in each of the four cardinal directions and counting the total number of small trees (<2.5 cm DBH and at least 0.30 m tall) under or touching the arms of the observer. We also recorded the “reach” of the observer or the width of the transect (measured fingertip to fingertip prior to the survey and used this to calculate the sample area). We then listed the small stem species by noting the three most common species encountered during the small stem strip transects [36].
We counted all of the larger shrubs and small trees (2.5–12.7 cm DBH) within 15 m of the point to estimate the larger shrub and small tree density (Figure S8). We also listed the three most common larger shrub and small tree species. We then estimated the vegetative composition of the plot within a 15 m radius by estimating the percentage of cover type in each of nine categories from eye level looking down. Categories included NWSG, CSG, forbs, shrubs, trees, water, rock, bare ground, and leaf litter [37]. For consistency, two of same three rotating observers conducted the vegetation monitoring for the duration of the project.
2.5. Data Preparation and Statistical Analysis
We only included a species in the productivity analyses if we had at least 100 captures for that species. Capture-mark-recapture (CMR) survival analyses included species that had at least a capture rate of 3.9 captures per year and a recapture rate of 0.4 recaptures per year for marginally precise estimates, and 14.9 captures and 1.1 recaptures per year for precise estimates [38]. We calculated the capture rate as the number of captures per hour that a net was open. For most resident species and some migrating species, no seasonal drop-off in captures was evident, suggesting that the sampled population was mostly local, late to migrate, or that the separation of local breeders from migrating transients was not possible. Hence, we assumed that some species were capturable on all dates. For others, we used the first or last date we captured them to determine capturability.
We conducted all statistical tests using the program R (R v. 4.2.2, R Core Team, Vienna, Austria) and performed all CMR analyses using Program MARK (Program MARK v. 10.1, Colorado State University, Fort Collins, CO, USA) and the R package, RMark (RMark v. 3.0.0, National Marine Fisheries Service, Seattle, WA, USA). We created survival models for each species and tested the goodness of fit using RMark’s “release.gof” function to determine whether all individuals had the same probability of recapture and survival. We created Cormack–Jolly–Seber models [39] to best explain the capture histories from 2009 to 2022 at SVPH. We only included individuals that were aged as adults at initial capture in the modeling due to young birds having lower survival and recapturability [40].
We created reproductive indices for each species including field sparrows by calculating the ratio of adults and juveniles captured for each year [41]. We included the fall data in the productivity analyses for Spinus tristis (American goldfinch) because they are late breeders, with young often fledging from July to September [42]. We assigned each species a migration strategy and winter location (Table 1) based on the information available from Birds of the World species accounts [43]. We calculated the fall capture rates for each group by determining the captures per net hour for each year. We excluded same year recapture records to avoid overestimating the capture rates. We also used Wilcoxon rank sum tests to assess whether the productivity and capture rates differed after the restoration of the old field using a significance p-value cutoff of 0.05. We modeled the species and migration strategy groups that had capture rates and reproductive indices that significantly differed before and after the restoration.

Table 1.
Reproductive indices (young/adult), migration strategy, and fall capture rates (birds/net hour) of species captured at the Shaker Village of Pleasant Hill during 2009–2022. Total captures include adult and immature birds.
We calculated the species richness during the breeding season and fall migration for each year of banding. We used Wilcoxon rank sum tests to compare the species richness of captured birds before and after the restoration. Weather data were compiled using Kentucky Mesonet [44]. Monthly averages were pulled directly from Mesonet and compiled for the months of May–Oct for temperature and precipitation.
We tested the productivity and capture rates for normality using Shapiro–Wilk tests with a p-value cutoff of 0.05 [45,46]. If the capture rate distribution was normal, we placed the capture rates in linear models to assess how the weather and vegetation factors influenced the change in the migration strategy capture rates. If the distribution was non-normal, we created generalized linear models to explain which covariates most influenced the capture rates. The covariates for these models included precipitation, temperature, restoration status, and vegetation sampling. We selected the best model based on the corrected Akaike information criterion (AICc) [47,48].
3. Results
During the 2009–2022 breeding seasons, we operated the station for 4400 net hours on 110 dates, resulting in 3516 captures (2603 new bands, 767 recaptures, and 146 unbanded). We captured sixty-eight species during the breeding season, though eight species were assumed to be transients and are not known to breed in the area. During the 2009–2022 fall migration seasons, we operated the station for 4220 net hours on 108 dates, resulting in 4902 captures (4135 new bands, 606 recaptures, and 161 unbanded) of 90 species.
The American goldfinch productivity was significantly higher after the restoration of native vegetation (p = 0.009 mean = 0.75 immature/adult). Field sparrows also exhibited significantly higher productivity post restoration (p = 0.01 mean = 1.0 immature/adult; Figure 1). The long-distance and medium-distance migrant fall capture rates were significantly higher after the conversion of the old field (p = 0.006 mean = 0.4 captures/net hour and p = 0.04 mean = 0.2 captures/net hour, respectively; Figure 2). When the capture rates were combined across all seasons for each year, only the long-distance migration species capture rates were significantly higher after the restoration (p = 0.009 mean = 0.5 captures/net hour). Capture rates for single species did not differ during fall before and after the conversion (Table 1), while the survival estimates (φ) varied by species (Table 2). Species richness did not significantly change as a result of the restoration.
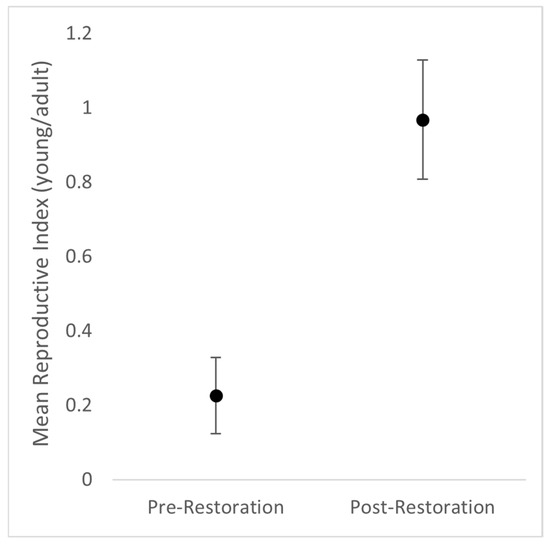
Figure 1.
The productivity of field sparrows at the Shaker Village of Pleasant Hill in Mercer County, KY before and after the restoration of the old field.
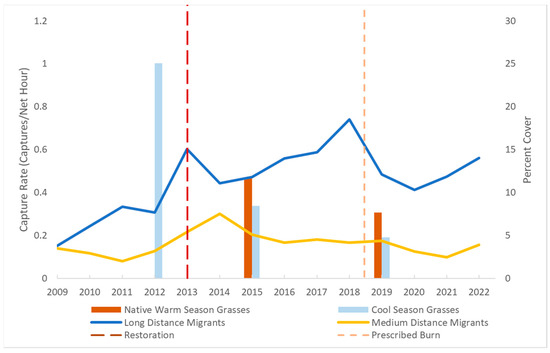
Figure 2.
The fall capture rates for long-distance and medium-distance migrants at Shaker Village of Pleasant Hill in Mercer County, KY 2009–2022. The left axis is the bird capture rate, and the right axis is the percent cover of native warm season or cool season grasses. Vertical lines indicate the restoration of native vegetation in the area and prescribed fire.
Table 2.
The capture-mark-recapture survival estimates for species captured at Shaker Village of Pleasant Hill during 2009–2022.
All of the generalized linear models for the pooled fall capture rates and the combined seasons shared the same covariates in their best fit model (Table S1). This covariate was the interaction between NWSG and CSG. Similarly, reproductive indices for field sparrows and American goldfinches were also best explained by the interaction of NWSG and CSG (Tables S2 and S3, AICc = −213.6 Wi = 1, AICc = −222.4 Wi = 1, respectively). We found that the percentage of NWSG significantly increased from 2012 to 2019 (p = 0.006 mean = 6.5% coverage). Conversely, the percentage of CSG significantly decreased during the same period (p = 0.002 mean = 12.8% coverage) (Figure 2).
4. Discussion
The impact on breeding birds was more subtle than expected, but we did find positive effects after native vegetation restoration including higher field sparrow productivity (Figure 1). Field sparrow productivity at our site started at 0.272 ± 0.102 young/adult and significantly increased to 0.892 ± 0.16 post restoration. The Institute for Bird Populations reported a rangewide mean reproductive index of 0.27 young/adult for field sparrows for 1992–2006 (Figure 3) [49]. Thus, it seems that the restoration led to a notable and valuable increase in productivity for this species. The American goldfinch productivity also increased post restoration. There is a paucity of productivity information in the literature for American goldfinches because their young generally fledge after the typical summer banding season. Nonetheless, Port and Schottler [50] reported American goldfinch preferences for areas with high forb diversity. Although our vegetation monitoring did not account for forb diversity, we successfully planted several species of forbs that produced edible seeds including black-eyed Susan and purple coneflower. Because American goldfinches are seed specialists [42], we surmise that increased small seed availability boosted their productivity.
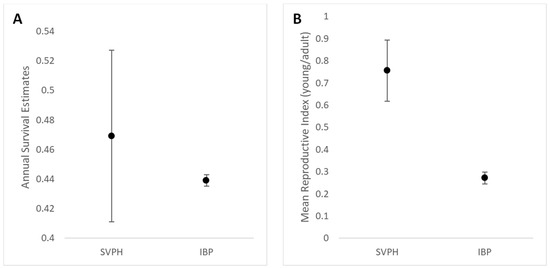
Figure 3.
The estimated annual survival for field sparrows was similar between Shaker Village and the Institute for Bird Populations (A). Field sparrows at Shaker Village of Pleasant Hill had a higher mean reproductive index compared to that reported in the study by the Institute for Bird Populations (B).
The breeding season capture rates and productivity did not change significantly for species other than field sparrows and American goldfinches as a result of native vegetation restoration, and neither did species richness. In general, we were surprised to find this old field to be an area of productive, abundant breeding birds prior to restoration. The eradication of CSG and other non-native plants may not be necessary in all circumstances if supporting the productivity of breeding birds is the objective, and if the existing habitat conditions already support a diverse structure. We believe that in part, what allowed our study site to support a diversity of bird species pre-conversion was its interspersed shrub-grass structure. Woody vegetation typically adds bird diversity to grassy fields, allowing species that generally nest in shrubs and edges as well as generalists to co-exist with grassland species [18]. Given the initial abundance of breeding birds at our site, we recommend particularly prioritizing grass-shrub restoration in areas that lack initial structural heterogeneity (a well-developed grass component with diverse forbs and interspersed shrubs).
The bird conservation community has placed increased emphasis on the conservation of the full life cycle of birds in recent years [3]. While native grass and shrub restoration is often promoted for bird conservation, the value of resulting habitats during migration has rarely been assessed. We found the strongest and most beneficial effects of the native conversion during the fall migration season. A decline in the coverage of CSG and an increase in the coverage of native grasses corresponding to an increase in the long-distance migrant capture rates can be seen in Figure 2. Long-distance migrants included large samples for neotropical migrants, Leiothlypis peregrina (Tennessee warbler), Icteria virens (yellow-breasted chat), Dumetella carolinensis (gray catbird), Passerina cyanea (indigo bunting), and Geothlypis trichas (common yellowthroat), and the capture rates for the cohort including these species more than doubled on average after the conversion. Medium-distance migrants with high capture rates included Melospiza georgiana (swamp sparrow), Zonotrichia albicollis (white-throated sparrow), Troglodytes aedon (house wren), Setophaga palmarum (palm warbler), and Melospiza lincolnii (Lincoln’s sparrow), and this group also showed a significantly higher capture rate after restoration. Our results demonstrate that the restoration and management of native vegetation can enhance stopover habitats for fall migrating songbirds including neotropical migrants.
Many neotropical migrants are declining [4]. Over the course of our study, the Tennessee warbler was the most frequently captured neotropical migrant outside of its breeding range. This is not surprising since they are a fairly common migrant in the area [51]. Tennessee warblers are primarily insectivorous, although they may consume some fruit during migratory stopovers [52]. While not the focus of their study, Morris et al. [53] also noted high capture rates of Tennessee warblers in an area of invasive shrub removal and native shrub plantings in Michigan. In our study, while the capture rates of Tennessee warblers were not significantly different after the conversion, neither were any other species analyzed separately (in fall). Nonetheless, the capture rates for pooled cohorts of medium- and long-distance migrants were significantly higher, demonstrating that it was not any single fall migrant species that increased post restoration, instead, many species increased.
Capture rates are affected by more than local habitat change. The abundance of migratory birds is no doubt influenced by broader population trends. The North American Breeding Bird Survey reported a trend of −2.6% change per year for field sparrows in Kentucky, and −2.3% change per year in the United States, with high credibility during the time period of our study [54]. Thus, the positive responses we found for this species during the same time frame cannot be attributed to larger population trends.
Due to our methodology, the weather conditions were fairly stable on days when netting activity occurred, but overall seasonal weather patterns may also affect the capture rates [55]. We used modeling to assess whether our results were affected more so by seasonal weather patterns than habitat change, and found that the interaction between NWSG and CSG best explained the changes we observed in the fall capture rate for migratory cohorts and productivity for field sparrows (Tables S2 and S4). Nonetheless, we believe that seasonal weather patterns may explain some of the annual variation that can be seen in Figure 2 and Figure 4.
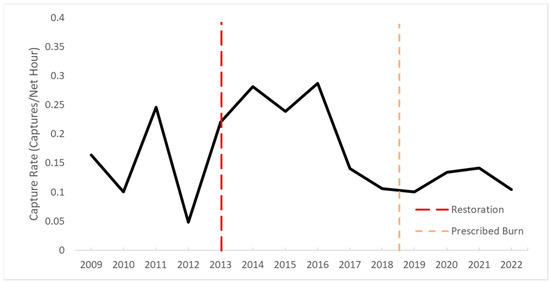
Figure 4.
The fall capture rate of field sparrows (n = 657) at Shaker Village of Pleasant Hill in Mercer County, KY 2009–2022. Vertical lines indicate the restoration of native vegetation in the area and prescribed fire.
Studies often focus on changes in habitat structure when it comes to native plant conversions and their effects on birds [13,26]. However, we attempted to produce a similar structure with native vegetation that mirrored the baseline vegetative structure. Evidently we had some success, as supported by the lack of significant structural differences found in the vegetation monitoring data, despite the change in species composition. No doubt, some small-scale structural change occurred as a result of restoring NWSGs (bunchgrass species) where CSGs (sod-forming species) previously occurred. Still, the increase in numbers of neotropical migrants and boost in productivity we observed for select species may have been caused in part by increased food availability [29]. Previous studies in the eastern U.S. have found the abundance and diversity of herbivorous insects to be higher in areas with greater native plant cover [56,57]. The fruits from native plants have also been found to be important and may be preferred by fall migrants [28,58]. Furthermore, grass and forb seed availability generally peaks 2–3 years following native restoration due to young perennial plants producing an abundance of seeds, which tapers off as they age [59]. This may have resulted in a peak in the capture rates for seed-eating species including field sparrows and several medium-distance migrants in the years immediately following the conversion (Figure 2 and Figure 3).
Continued disturbance is the key to encouraging native grasses to prevail in restored areas. Managers typically conduct prescribed burning to promote NWSGs and discourage cool-season grasses. Burn regimes of two to three years are recommended [13]; however, our study represents a real-world example of how burns often occur at longer intervals due to weather, staff limitations, etc. Due to sample size, we did not conduct analyses to assess the effects of the prescribed burn on the vital rates or capture rates. However, there has been an increase in Solidago spp. (goldenrod) and Rubus spp. (blackberries) and a decrease in grasses at this study area since the conversion, and future management efforts may attempt to encourage more grasses. We surmise that the decrease in grasses may be leading to the decrease in the fall capture rates for some species. For instance, the capture rates of field sparrows during the fall increased after the conversion of the old field but declined in 2017 (Figure 4). More study is needed to understand how to best prolong the positive effects that we documented in the initial years after the restoration.
Disturbance can temporarily displace birds, which may result in concerns over negative effects, especially when disturbance occurs during the nesting season [60]. Based on recent studies, we did not have enough data to compare survival before and after the conversion [38]. However, we were able to estimate the adult apparent survival for the duration of the study period with good precision for five species (Table 2). Our survival estimate for field sparrows (φ = 0.469, SE= 0.058) was comparable to the rangewide mean (φ = 0.439) reported by the Institute for Bird Populations [49]. However, we also obtained precise estimates for common yellowthroat, gray catbird, and yellow-breasted chat, which were low in comparison to the rangewide means [49]. We surmise that our survival estimates for these species, which prefer thick cover with a woody component, were affected by the low capture rates that occurred during years with restoration-related disturbance. We focused on management efforts outside of the nesting season to avoid a loss in productivity, but the capture rates dropped temporarily for these species for a few years post-conversion, as Rubus spp. and other thicket forming vegetation were reduced briefly by the management. More study is needed to understand the effects of similar management on survival and emigration, particularly for species with a narrow range of habitat preferences. Of course, other factors are at play when it comes to adult survival, especially for neotropical migrants. Factors hindering the survival of common yellowthroats, gray catbirds, and yellow-breasted chats in our area may occur on stopover sites elsewhere or the winter grounds. Such factors may have impacts reaching far beyond our local population. For instance, regional analyses estimate that gray catbirds have the lowest survival within our bird conservation region (BCR) in comparison to other North American BCRs [61].
Despite difficulties in assessing the effects of management on the survival for thicket-loving species, we did observe positive effects shortly after the restoration. The season with the most habitat disturbance resulted in higher-than-average productivity for field sparrows, and the second highest peak for long-distance migrant capture rates. Furthermore, the long-term effects of management were likely beneficial by promoting ephemeral shrubland habitats, even for species that experienced short-term drops in capture rate. Along these lines, Akresh et al. [60] found that the short-term effects of shrubland management on Setophaga discolor (prairie warblers) were minimal and allowed second year birds to colonize territories in disturbed areas.
Although we restored native grass in our study area, the area contained a significant woody component. We had very small sample sizes for species that prefer more open grasslands, many of which are of conservation concern (e.g., grasshopper sparrow, Centronyx henslowii [Henslow’s sparrow], Sturnella magna [eastern meadowlark], Dolichonyx oryzivorus [bobolink], and Spiza americana [dickcissel]). Although several studies have provided management recommendations for these species [18,26,50], more study is needed to understand how native vegetation restoration affects the migratory stopover use and vital rates for these species, especially within our region.
The size of our study area was small (8.1 ha) but representative of many other similarly managed areas in the region. Given that our study site was bordered by a row crop field, in a sense, our study mimics a private land setting where a less-productive area has been set aside for wildlife habitat. However, it is important to note that this study site was also located on the edge of a larger area where a great deal of habitat management occurred during the study period. Over 400 ha was managed for native early successional vegetation on the preserve during 2009–2022, with most of the restoration efforts taking place from 2009 to 2014. Thus, the changes we observed in the capture rates, especially in fall, may have been bolstered by changes occurring on the larger landscape. Nonetheless, the management conducted on our study site was representative of the management occurring in the larger area, and hence, the landscape effect should not contradict our conclusions.
Initiatives to restore native vegetation for pollinators, grassland birds, northern bobwhite, and grassland biodiversity continually help to fuel more restorations of varying scales in the USA. In general, our study shows that native grass and shrub restorations can support a wide variety of bird species. The restoration of native vegetation at our study site boosted productivity for select species and the capture rate for fall migrants using the area as a stopover habitat. As conservation biologists grapple with how to support the full life cycle of migratory songbirds and manage public and private areas to maximize benefit, it seems that the restoration of native grass and shrub vegetation is worthy of effort, especially to support stopover habitats.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d16100615/s1. Table S1: Model selection using AICc to determine the best fit model to explain the changes in capture rates for migration strategy cohorts during the breeding and fall migration season. Table S2: Model selection using AICc to determine the best fit model to explain the changes in field sparrow productivity. Table S3: Model selection using AICc to determine the best fit model to explain the changes in American goldfinch productivity. Table S4: Model selection using AICc to determine the best fit model to explain the changes in field sparrow capture rates for the breeding and fall migration seasons. Figures S1–S8: Photographs showing the study site before, during, and after the native vegetation restoration.
Author Contributions
Conceptualization, K.G.S. and D.P.; Resources, K.G.S., B.L. and D.P.; Methodology, K.G.S.; Data analysis and figures, M.D.P.; Fieldwork, K.G.S., B.L., M.D.P., L.K.B., J.M., D.S., D.P., J.O.B., L.T. and L.S.B.; Writing—original draft, K.G.S., B.L. and M.D.P.; Writing—review and editing, K.G.S., B.L., M.D.P., L.K.B., J.M., D.S., D.P., J.O.B., L.T. and L.S.B.; Funding acquisition, K.G.S. and B.L.; Data curation, K.G.S., B.L., M.D.P., D.S. and L.T.; Project administration, K.G.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Pittman-Robertson Wildlife Restoration Program and Shaker Village of Pleasant Hill.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data from this project are available upon an approved data-sharing agreement with the Kentucky Department of Fish and Wildlife Resources.
Acknowledgments
We thank Shaker Village of Pleasant Hill for their support of this project. We also acknowledge Ben Bowman, Kasie Bradley, Bree Furfey, Erin Harper, Kaitlyn Kelly, Tonya Mammone, Meghan Raley, Michaela Rogers, Scott Slankard, Angel Smallwood, Caleb Switzer, and Shawchyi Vorisek for their work on this project. The graphical abstract artwork was produced by Devon Stivers. The findings and conclusions in this article are those of the authors and do not necessarily represent the views of the United States Fish and Wildlife Service.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Brawn, J.D.; Robinson, S.K.; Thompson, F.R., III. The role of disturbance in the ecology and conservation of birds. Annu. Rev. Ecol. Syst. 2001, 32, 251–276. [Google Scholar] [CrossRef]
- Schlossberg, S.; King, D.I.; Chandler, R.B.; Mazzei, B.A. Regional syntheses of habitat relationships in shrubland birds. J. Wildl. Manag. 2010, 74, 1513–1522. [Google Scholar]
- Rosenberg, K.V.; Dokter, A.M.; Blancher, P.J.; Sauer, J.R.; Smith, A.C.; Smith, P.A.; Stanton, J.C.; Panjabi, A.; Helft, L.; Parr, M.; et al. Decline of the North American avifauna. Science 2019, 366, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, K.V.; Kennedy, J.A.; Dettmers, R.; Ford, R.P.; Reynolds, D.; Alexander, J.D.; Beardmore, C.J.; Blancher, P.J.; Bogart, R.E.; Butcher, G.S.; et al. Partners in Flight Landbird Conservation Plan: 2016 Revision for Canada and Continental United States. Available online: https://partnersinflight.org/resources/the-plan/ (accessed on 30 August 2024).
- Shaffer, J.A.; DeLong, J.P. The effects of management practices on grassland birds—An introduction to North American grasslands and the practices used to manage grasslands and grassland birds (ver. 1.1, March 2022). In The Effects of Management Practices on Grassland Birds; Johnson, D.H., Igl, L.D., Shaffer, J.A., DeLong, J.P., Eds.; Professional Paper 1842; U.S. Geological Survey: Reston, VA, USA, 2019. [Google Scholar] [CrossRef]
- U.S. North American Bird Conservation Initiative Committee (NABCI). 2014 Farm Bill Field Guide to Fish and Wildlife Conservation; U.S. North American Bird Conservation Initiative (NABCI): Washington, DC, USA, 2015. [Google Scholar]
- Estes, D.; Brock, M.; Homoya, M.; Dattilo, A. A Guide to the Grasslands of the Mid-South. 2016. Available online: https://www.segrasslands.org/guide-to-the-grasslands-of-the-midsouth (accessed on 30 August 2024).
- National Bobwhite and Grassland Initiative (NBGI). Bobwhite Almanac: State of the Bobwhite 2022; NBGI Technical Committee Publication: Clemson, SC, USA, 2022. [Google Scholar]
- Nelson, S.B.; Coon, J.J.; Duchardt, C.J.; Fischer, J.D.; Halsey, S.J.; Kranz, A.J.; Parker, C.M.; Schneider, S.C.; Swartz, T.M.; Miller, J.R. Patterns and mechanisms of invasive plant impacts on North American birds: A systematic review. Biol. Invasions 2017, 19, 1547–1563. [Google Scholar] [CrossRef]
- Keyser, P.D.; Buehler, D.A.; Hedges, K.; Hodges, J.; Lituma, C.; Loncarich, F.; Martin, J.A. Eastern grasslands: Conservation challenges and opportunities on private lands. Wildl. Soc. Bull. 2019, 43, 382–390. [Google Scholar] [CrossRef]
- Washburn, B.E.; Barnes, T.G.; Sole, J.D. Improving Northern Bobwhite Habitat by converting tall fescue fields to native warm-season grasses. Wildl. Soc. Bull. 2000, 28, 97–104. [Google Scholar]
- Gruchy, J.P.; Harper, C.A. Effects of field management practices on Northern Bobwhite habitat. J. Southeast. Assoc. Fish Wildl. Agencies 2014, 1, 133–141. [Google Scholar]
- Sinnott, E.A.; Thompson, F.R.; Weegman, M.D.; Thompson, T.R. Northern Bobwhite juvenile survival is greater in native grasslands managed with fire and grazing and lower in non-native field borders and strip crop fields. Ornithol. Appl. 2022, 124, duab057. [Google Scholar] [CrossRef]
- Baskett, C.A.; Emery, S.A.; Rudgers, J.A. Pollinator visits to threatened species are restored following invasive plant removal. Int. J. Plant Sci. 2011, 172, 411–422. [Google Scholar] [CrossRef]
- Lukens, L.; Kasten, K.; Stenoien, C.; Cariveau, A.; Caldwell, W.; Oberhauser, K. Monarch habitat in conservation grasslands. Front. Ecol. Evol. 2020, 8, 13. [Google Scholar] [CrossRef]
- Sexton, A.; Emery, S.M. Grassland restorations improve pollinator communities: A meta-analysis. J. Insect Conserv. 2020, 24, 719–726. [Google Scholar] [CrossRef]
- Beneduci, Z.J.; Scott, D.A.; Byrd, S.M.; Swab, R.M. We Built It; Did They Come? Pollinator Diversity and Community Structure in a Post-Mining Prairie Restoration Project. Ecol. Restor. 2023, 41, 180–188. [Google Scholar] [CrossRef]
- Dykes, S.A. Effectiveness of native grassland restoration in restoring grassland bird communities in Tennessee. Master’s Thesis, University of Tennessee-Knoxville, Knoxville, TN, USA, 2005. [Google Scholar]
- Yeiser, J.; Morgan, J.J.; Baxley, D.; Chandler, R.B.; Martin, J.A. Private land conservation has landscape-scale benefits for wildlife in agroecosystems. J. Appl. Ecol. 2018, 55, 1930–1939. [Google Scholar] [CrossRef]
- Lituma, C.M.; Buehler, D.A. Cost-share conservation practices have mixed effects on priority grassland and shrubland breeding bird occupancy in the Central Hardwoods Bird Conservation Region, USA. Biol. Conserv. 2020, 244, 108510. [Google Scholar] [CrossRef]
- Baldwin, H.Q.; Grace, J.B.; Barrow, W.C.; Rohwer, F.C. Habitat relationships of birds overwintering in a managed coastal prairie. Wilson J. Ornithol. 2007, 119, 189–197. [Google Scholar] [CrossRef]
- Blank, P.J.; Parks, J.R.; Dively, G.P. Wintering bird response to fall mowing of herbaceous buffers. Wilson J. Ornithol. 2011, 123, 59–64. [Google Scholar] [CrossRef]
- Evans, K.O.; Burger, L.W.; Riffell, S.; Smith, M.D.; Twedt, D.J.; Wilson, R.R.; Vorisek, S.; Rideout, C.; Heyden, K. Avian response to conservation buffers in agricultural landscapes during winter. Wildl. Soc. Bull. 2014, 38, 257–264. [Google Scholar] [CrossRef]
- Stumpf, K.; Muise, C. Increasing capture rates of grassland birds over thirteen years indicates successful restoration. Ga. J. Sci. 2023, 81, 1. [Google Scholar]
- Smith, R.J.; Hatch, M.L. Stopover ecology of fall migrating landbirds at an inland stopover site in northeastern Pennsylvania dominated by nonnative vegetation. Wilson J. Ornithol. 2020, 132, 398–409. [Google Scholar] [CrossRef]
- Gill, D.E.; Blank, P.; Parks, J.; Guerard, J.B.; Lohr, B.; Schwartzman, E.; Gruber, J.G.; Dodge, G.; Rewa, C.A.; Sears, H.F. Plants and breeding bird response on managed conservation reserve program grassland in Maryland. Wildl. Soc. Bull. 2006, 34, 944–956. [Google Scholar] [CrossRef]
- Borgmann, K.L.; Rodewald, A.D. Nest predation in an urbanizing landscape: The role of exotic shrubs. Ecol. Appl. 2004, 14, 1757–1765. [Google Scholar] [CrossRef]
- Smith, S.B.; DeSando, S.A.; Pagano, T. The value of native and invasive fruit-bearing shrubs for migrating songbirds. Northeast. Nat. 2013, 20, 171–184. [Google Scholar] [CrossRef]
- Narango, D.L.; Tallamy, D.W.; Marra, P.P. Native plants improve breeding and foraging habitat for an insectivorous bird. Biol. Conserv. 2017, 213, 42–50. [Google Scholar] [CrossRef]
- Morgan, J.; Robinson, B. Road to Recovery: Five Year Benchmark Report. 2015. Available online: https://fw.ky.gov/Documents/quailbenchmarkreport2015.pdf (accessed on 30 August 2024).
- Kentucky Department of Fish and Wildlife Resources (KDFWR). Kentucky’s State Wildlife Action Plan. 2023. Available online: https://fw.ky.gov/WAP/Documents/2023_SWAP_PublicComment_AR02.pdf (accessed on 30 August 2024).
- Dunn, E.H.; Hussell, D.J.T.; Adams, R.J. Monitoring songbird population change with autumn mist netting. J. Wildl. Manag. 1997, 61, 389–396. [Google Scholar] [CrossRef]
- DeSante, D.F.; Burton, F.R.; Velez, P.; Froehlich, D.; Kaschube, D. MAPS Manual: 2009 Protocol; The Institute for Bird Populations: Point Reyes Station, CA, USA, 2009. [Google Scholar]
- DeSante, D.F.; Burton, K.M.; Kaschube, D.; Velez, P.; Froehlich, D.; Albert, S. MAPS Manual: 2022 Protocol; The Institute for Bird Populations: Petaluma, CA, USA, 2022. [Google Scholar]
- Pyle, P. Identification Guide to North American Birds; Part I; Slate Creek Press: Bolinas CA, USA, 1997. [Google Scholar]
- Husch, B.; Beers, T.W.; Kershaw, J.A., Jr. Forest Mensuration, 4th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2003. [Google Scholar]
- Sampling Vegetation Attributes; Technical Reference 1734-4; Bureau of Land Management: Denver, CO, USA, 1999.
- Kaschube, D.R.; Saracco, J.F.; Ray, C.; Godwin, C.M.; Foster, K.R.; Pyle, P. Minimum capture-recapture rates and years of banding station operations to obtain reliable adult annual survival estimates. J. Field Ornithol. 2022, 93, 7. [Google Scholar] [CrossRef]
- Lebreton, J.D.; Burnham, K.P.; Clobert, J.; Anderson, D.R. Modeling survival and testing biological hypotheses using marked animals: A unified approach with case studies. Ecol. Monogr. 1992, 62, 67–118. [Google Scholar] [CrossRef]
- Beauchamp, G. Annual apparent survival across species is lower in juvenile than adult birds but has similar ecological correlates. Ibis 2023, 165, 448–457. [Google Scholar] [CrossRef]
- DeSante, D.F.; Kaschube, D.R. The Monitoring Avian Productivity and Survivorship (MAPS) Program 2004, 2005, and 2006 Report. Bird Popul. 2009, 9, 86–169. [Google Scholar]
- McGraw, K.J.; Middleton, A.L. American Goldfinch (Spinus tristis). In Birds of the World, version 1.0; Rodewald, P.G., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Billerman, S.M.; Keeney, B.K.; Rodewald, P.G.; Schulenberg, T.S. (Eds.) Birds of the World; Cornell Laboratory of Ornithology: Ithaca, NY, USA, 2022; Available online: https://birdsoftheworld.org/bow/home (accessed on 30 August 2024).
- Western Kentucky University. Kentucky Mesonet. 2024. Available online: https://www.kymesonet.org/about.html (accessed on 7 January 2024).
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Mendes, M.; Pala, A. Type I error rate and power of three normality tests. Pak. J. Inf. Technol. 2003, 2, 135–139. [Google Scholar]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Sigiura, N. Further analysis of the data by Akaike’s information criterion and the finite corrections. Commun. Stat. 1978, 7, 13–26. [Google Scholar] [CrossRef]
- DeSante, D.F.; Kaschube, D.R.; Saracco, J.F.; Vital Rates of North American Landbirds. The Institute for Bird Populations. 2015. Available online: https://www.VitalRatesOfNorthAmericanLandbirds.org (accessed on 29 January 2024).
- Port, J.; Schlottler, S. The effect of floristic composition and bird communities in a set of four grassland reconstruction types. Ecol. Restor. 2017, 35, 112–119. [Google Scholar] [CrossRef]
- Palmer-Ball, B.L. Annotated Checklist of the Bird of Kentucky; Kentucky Ornithological Society: Louisville, KY, USA, 2019. [Google Scholar]
- Bayley, N.J.; Rosenberg, K.V.; Gomez, C.; Hobson, K.A. Habitat choice shapes the spring stopover behaviour of a Nearctic-Neotropical migratory songbird. J. Ornithol. 2019, 160, 377–388. [Google Scholar] [CrossRef]
- Morris, S.R.; Andrijevic, A.S.; Sullivan, R.; Keith, R.S.; Keith, B.S.; Sheets, H.D. An 18-year study of migration and stopover ecology of Tennessee Warblers in Kalamazoo County Michigan. Wilson J. Ornithol. 2013, 125, 70–78. [Google Scholar] [CrossRef]
- Sauer, J.R.; Link, W.A.; Hines, J.E. The North American Breeding Bird Survey, Analysis Results 1966–2022; U.S. Geological Survey: Kearneysville, WV, USA, 2020. [CrossRef]
- Berthold, P. Use of mist nets for monitoring landbird fall population trends, and comparison with other methods. Stud. Avian Biol. 2004, 29, 112–115. [Google Scholar]
- Burghardt, K.T.; Tallamy, D.W.; Philips, C.; Shropshire, K.J. Non-native plants reduce abundance, richness, and host specialization in lepidopteran communities. Ecosphere 2010, 1, 11. [Google Scholar] [CrossRef]
- Burghardt, K.T.; Tallamy, D.W. Plant origin asymmetrically impacts feeding guilds and life stages driving community structure of herbivorous arthropods. Divers. Distrib. 2013, 19, 553–1565. [Google Scholar] [CrossRef]
- Gallinat, A.S.; Primack, R.; Lloyd-Evans, T.L. Can invasive species replace native species as a resource for birds under climate change? A case study on bird-fruit interactions. Biol. Conserv. 2019, 241, 108268. [Google Scholar] [CrossRef]
- Houseal, G.A. Tallgrass Prairie Center’s Native Seed Production Manual. 2007. Available online: https://scholarworks.uni.edu/facbook/102 (accessed on 25 August 2024).
- Akresh, M.E.; King, D.I.; Brooks, R.T. Demographic response of a shrubland bird to habitat creation, succession and disturbance in a dynamic landscape. For. Ecol. Manag. 2015, 336, 72–80. [Google Scholar] [CrossRef]
- Institute for Bird Populations (IBP). MAPS Data Exploration Tool: Processed MAPS Results. Version 1.0. 2023. Available online: https://ibp-maps-data-exploration-tool.org (accessed on 24 January 2024).




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).