
The Effects of Native Grass and Shrub Restoration on Nesting and Fall Migrant Songbirds in Central Kentucky

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Vegetation Management
2.3. Constant Effort Mist Netting
2.4. Vegetation Monitoring
2.5. Data Preparation and Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brawn, J.D.; Robinson, S.K.; Thompson, F.R., III. The role of disturbance in the ecology and conservation of birds. Annu. Rev. Ecol. Syst. 2001, 32, 251–276. [Google Scholar] [CrossRef]
- Schlossberg, S.; King, D.I.; Chandler, R.B.; Mazzei, B.A. Regional syntheses of habitat relationships in shrubland birds. J. Wildl. Manag. 2010, 74, 1513–1522. [Google Scholar]
- Rosenberg, K.V.; Dokter, A.M.; Blancher, P.J.; Sauer, J.R.; Smith, A.C.; Smith, P.A.; Stanton, J.C.; Panjabi, A.; Helft, L.; Parr, M.; et al. Decline of the North American avifauna. Science 2019, 366, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, K.V.; Kennedy, J.A.; Dettmers, R.; Ford, R.P.; Reynolds, D.; Alexander, J.D.; Beardmore, C.J.; Blancher, P.J.; Bogart, R.E.; Butcher, G.S.; et al. Partners in Flight Landbird Conservation Plan: 2016 Revision for Canada and Continental United States. Available online: https://partnersinflight.org/resources/the-plan/ (accessed on 30 August 2024).
- Shaffer, J.A.; DeLong, J.P. The effects of management practices on grassland birds—An introduction to North American grasslands and the practices used to manage grasslands and grassland birds (ver. 1.1, March 2022). In The Effects of Management Practices on Grassland Birds; Johnson, D.H., Igl, L.D., Shaffer, J.A., DeLong, J.P., Eds.; Professional Paper 1842; U.S. Geological Survey: Reston, VA, USA, 2019. [Google Scholar] [CrossRef]
- U.S. North American Bird Conservation Initiative Committee (NABCI). 2014 Farm Bill Field Guide to Fish and Wildlife Conservation; U.S. North American Bird Conservation Initiative (NABCI): Washington, DC, USA, 2015. [Google Scholar]
- Estes, D.; Brock, M.; Homoya, M.; Dattilo, A. A Guide to the Grasslands of the Mid-South. 2016. Available online: https://www.segrasslands.org/guide-to-the-grasslands-of-the-midsouth (accessed on 30 August 2024).
- National Bobwhite and Grassland Initiative (NBGI). Bobwhite Almanac: State of the Bobwhite 2022; NBGI Technical Committee Publication: Clemson, SC, USA, 2022. [Google Scholar]
- Nelson, S.B.; Coon, J.J.; Duchardt, C.J.; Fischer, J.D.; Halsey, S.J.; Kranz, A.J.; Parker, C.M.; Schneider, S.C.; Swartz, T.M.; Miller, J.R. Patterns and mechanisms of invasive plant impacts on North American birds: A systematic review. Biol. Invasions 2017, 19, 1547–1563. [Google Scholar] [CrossRef]
- Keyser, P.D.; Buehler, D.A.; Hedges, K.; Hodges, J.; Lituma, C.; Loncarich, F.; Martin, J.A. Eastern grasslands: Conservation challenges and opportunities on private lands. Wildl. Soc. Bull. 2019, 43, 382–390. [Google Scholar] [CrossRef]
- Washburn, B.E.; Barnes, T.G.; Sole, J.D. Improving Northern Bobwhite Habitat by converting tall fescue fields to native warm-season grasses. Wildl. Soc. Bull. 2000, 28, 97–104. [Google Scholar]
- Gruchy, J.P.; Harper, C.A. Effects of field management practices on Northern Bobwhite habitat. J. Southeast. Assoc. Fish Wildl. Agencies 2014, 1, 133–141. [Google Scholar]
- Sinnott, E.A.; Thompson, F.R.; Weegman, M.D.; Thompson, T.R. Northern Bobwhite juvenile survival is greater in native grasslands managed with fire and grazing and lower in non-native field borders and strip crop fields. Ornithol. Appl. 2022, 124, duab057. [Google Scholar] [CrossRef]
- Baskett, C.A.; Emery, S.A.; Rudgers, J.A. Pollinator visits to threatened species are restored following invasive plant removal. Int. J. Plant Sci. 2011, 172, 411–422. [Google Scholar] [CrossRef]
- Lukens, L.; Kasten, K.; Stenoien, C.; Cariveau, A.; Caldwell, W.; Oberhauser, K. Monarch habitat in conservation grasslands. Front. Ecol. Evol. 2020, 8, 13. [Google Scholar] [CrossRef]
- Sexton, A.; Emery, S.M. Grassland restorations improve pollinator communities: A meta-analysis. J. Insect Conserv. 2020, 24, 719–726. [Google Scholar] [CrossRef]
- Beneduci, Z.J.; Scott, D.A.; Byrd, S.M.; Swab, R.M. We Built It; Did They Come? Pollinator Diversity and Community Structure in a Post-Mining Prairie Restoration Project. Ecol. Restor. 2023, 41, 180–188. [Google Scholar] [CrossRef]
- Dykes, S.A. Effectiveness of native grassland restoration in restoring grassland bird communities in Tennessee. Master’s Thesis, University of Tennessee-Knoxville, Knoxville, TN, USA, 2005. [Google Scholar]
- Yeiser, J.; Morgan, J.J.; Baxley, D.; Chandler, R.B.; Martin, J.A. Private land conservation has landscape-scale benefits for wildlife in agroecosystems. J. Appl. Ecol. 2018, 55, 1930–1939. [Google Scholar] [CrossRef]
- Lituma, C.M.; Buehler, D.A. Cost-share conservation practices have mixed effects on priority grassland and shrubland breeding bird occupancy in the Central Hardwoods Bird Conservation Region, USA. Biol. Conserv. 2020, 244, 108510. [Google Scholar] [CrossRef]
- Baldwin, H.Q.; Grace, J.B.; Barrow, W.C.; Rohwer, F.C. Habitat relationships of birds overwintering in a managed coastal prairie. Wilson J. Ornithol. 2007, 119, 189–197. [Google Scholar] [CrossRef]
- Blank, P.J.; Parks, J.R.; Dively, G.P. Wintering bird response to fall mowing of herbaceous buffers. Wilson J. Ornithol. 2011, 123, 59–64. [Google Scholar] [CrossRef]
- Evans, K.O.; Burger, L.W.; Riffell, S.; Smith, M.D.; Twedt, D.J.; Wilson, R.R.; Vorisek, S.; Rideout, C.; Heyden, K. Avian response to conservation buffers in agricultural landscapes during winter. Wildl. Soc. Bull. 2014, 38, 257–264. [Google Scholar] [CrossRef]
- Stumpf, K.; Muise, C. Increasing capture rates of grassland birds over thirteen years indicates successful restoration. Ga. J. Sci. 2023, 81, 1. [Google Scholar]
- Smith, R.J.; Hatch, M.L. Stopover ecology of fall migrating landbirds at an inland stopover site in northeastern Pennsylvania dominated by nonnative vegetation. Wilson J. Ornithol. 2020, 132, 398–409. [Google Scholar] [CrossRef]
- Gill, D.E.; Blank, P.; Parks, J.; Guerard, J.B.; Lohr, B.; Schwartzman, E.; Gruber, J.G.; Dodge, G.; Rewa, C.A.; Sears, H.F. Plants and breeding bird response on managed conservation reserve program grassland in Maryland. Wildl. Soc. Bull. 2006, 34, 944–956. [Google Scholar] [CrossRef]
- Borgmann, K.L.; Rodewald, A.D. Nest predation in an urbanizing landscape: The role of exotic shrubs. Ecol. Appl. 2004, 14, 1757–1765. [Google Scholar] [CrossRef]
- Smith, S.B.; DeSando, S.A.; Pagano, T. The value of native and invasive fruit-bearing shrubs for migrating songbirds. Northeast. Nat. 2013, 20, 171–184. [Google Scholar] [CrossRef]
- Narango, D.L.; Tallamy, D.W.; Marra, P.P. Native plants improve breeding and foraging habitat for an insectivorous bird. Biol. Conserv. 2017, 213, 42–50. [Google Scholar] [CrossRef]
- Morgan, J.; Robinson, B. Road to Recovery: Five Year Benchmark Report. 2015. Available online: https://fw.ky.gov/Documents/quailbenchmarkreport2015.pdf (accessed on 30 August 2024).
- Kentucky Department of Fish and Wildlife Resources (KDFWR). Kentucky’s State Wildlife Action Plan. 2023. Available online: https://fw.ky.gov/WAP/Documents/2023_SWAP_PublicComment_AR02.pdf (accessed on 30 August 2024).
- Dunn, E.H.; Hussell, D.J.T.; Adams, R.J. Monitoring songbird population change with autumn mist netting. J. Wildl. Manag. 1997, 61, 389–396. [Google Scholar] [CrossRef]
- DeSante, D.F.; Burton, F.R.; Velez, P.; Froehlich, D.; Kaschube, D. MAPS Manual: 2009 Protocol; The Institute for Bird Populations: Point Reyes Station, CA, USA, 2009. [Google Scholar]
- DeSante, D.F.; Burton, K.M.; Kaschube, D.; Velez, P.; Froehlich, D.; Albert, S. MAPS Manual: 2022 Protocol; The Institute for Bird Populations: Petaluma, CA, USA, 2022. [Google Scholar]
- Pyle, P. Identification Guide to North American Birds; Part I; Slate Creek Press: Bolinas CA, USA, 1997. [Google Scholar]
- Husch, B.; Beers, T.W.; Kershaw, J.A., Jr. Forest Mensuration, 4th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2003. [Google Scholar]
- Sampling Vegetation Attributes; Technical Reference 1734-4; Bureau of Land Management: Denver, CO, USA, 1999.
- Kaschube, D.R.; Saracco, J.F.; Ray, C.; Godwin, C.M.; Foster, K.R.; Pyle, P. Minimum capture-recapture rates and years of banding station operations to obtain reliable adult annual survival estimates. J. Field Ornithol. 2022, 93, 7. [Google Scholar] [CrossRef]
- Lebreton, J.D.; Burnham, K.P.; Clobert, J.; Anderson, D.R. Modeling survival and testing biological hypotheses using marked animals: A unified approach with case studies. Ecol. Monogr. 1992, 62, 67–118. [Google Scholar] [CrossRef]
- Beauchamp, G. Annual apparent survival across species is lower in juvenile than adult birds but has similar ecological correlates. Ibis 2023, 165, 448–457. [Google Scholar] [CrossRef]
- DeSante, D.F.; Kaschube, D.R. The Monitoring Avian Productivity and Survivorship (MAPS) Program 2004, 2005, and 2006 Report. Bird Popul. 2009, 9, 86–169. [Google Scholar]
- McGraw, K.J.; Middleton, A.L. American Goldfinch (Spinus tristis). In Birds of the World, version 1.0; Rodewald, P.G., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Billerman, S.M.; Keeney, B.K.; Rodewald, P.G.; Schulenberg, T.S. (Eds.) Birds of the World; Cornell Laboratory of Ornithology: Ithaca, NY, USA, 2022; Available online: https://birdsoftheworld.org/bow/home (accessed on 30 August 2024).
- Western Kentucky University. Kentucky Mesonet. 2024. Available online: https://www.kymesonet.org/about.html (accessed on 7 January 2024).
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Mendes, M.; Pala, A. Type I error rate and power of three normality tests. Pak. J. Inf. Technol. 2003, 2, 135–139. [Google Scholar]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Sigiura, N. Further analysis of the data by Akaike’s information criterion and the finite corrections. Commun. Stat. 1978, 7, 13–26. [Google Scholar] [CrossRef]
- DeSante, D.F.; Kaschube, D.R.; Saracco, J.F.; Vital Rates of North American Landbirds. The Institute for Bird Populations. 2015. Available online: https://www.VitalRatesOfNorthAmericanLandbirds.org (accessed on 29 January 2024).
- Port, J.; Schlottler, S. The effect of floristic composition and bird communities in a set of four grassland reconstruction types. Ecol. Restor. 2017, 35, 112–119. [Google Scholar] [CrossRef]
- Palmer-Ball, B.L. Annotated Checklist of the Bird of Kentucky; Kentucky Ornithological Society: Louisville, KY, USA, 2019. [Google Scholar]
- Bayley, N.J.; Rosenberg, K.V.; Gomez, C.; Hobson, K.A. Habitat choice shapes the spring stopover behaviour of a Nearctic-Neotropical migratory songbird. J. Ornithol. 2019, 160, 377–388. [Google Scholar] [CrossRef]
- Morris, S.R.; Andrijevic, A.S.; Sullivan, R.; Keith, R.S.; Keith, B.S.; Sheets, H.D. An 18-year study of migration and stopover ecology of Tennessee Warblers in Kalamazoo County Michigan. Wilson J. Ornithol. 2013, 125, 70–78. [Google Scholar] [CrossRef]
- Sauer, J.R.; Link, W.A.; Hines, J.E. The North American Breeding Bird Survey, Analysis Results 1966–2022; U.S. Geological Survey: Kearneysville, WV, USA, 2020. [CrossRef]
- Berthold, P. Use of mist nets for monitoring landbird fall population trends, and comparison with other methods. Stud. Avian Biol. 2004, 29, 112–115. [Google Scholar]
- Burghardt, K.T.; Tallamy, D.W.; Philips, C.; Shropshire, K.J. Non-native plants reduce abundance, richness, and host specialization in lepidopteran communities. Ecosphere 2010, 1, 11. [Google Scholar] [CrossRef]
- Burghardt, K.T.; Tallamy, D.W. Plant origin asymmetrically impacts feeding guilds and life stages driving community structure of herbivorous arthropods. Divers. Distrib. 2013, 19, 553–1565. [Google Scholar] [CrossRef]
- Gallinat, A.S.; Primack, R.; Lloyd-Evans, T.L. Can invasive species replace native species as a resource for birds under climate change? A case study on bird-fruit interactions. Biol. Conserv. 2019, 241, 108268. [Google Scholar] [CrossRef]
- Houseal, G.A. Tallgrass Prairie Center’s Native Seed Production Manual. 2007. Available online: https://scholarworks.uni.edu/facbook/102 (accessed on 25 August 2024).
- Akresh, M.E.; King, D.I.; Brooks, R.T. Demographic response of a shrubland bird to habitat creation, succession and disturbance in a dynamic landscape. For. Ecol. Manag. 2015, 336, 72–80. [Google Scholar] [CrossRef]
- Institute for Bird Populations (IBP). MAPS Data Exploration Tool: Processed MAPS Results. Version 1.0. 2023. Available online: https://ibp-maps-data-exploration-tool.org (accessed on 24 January 2024).



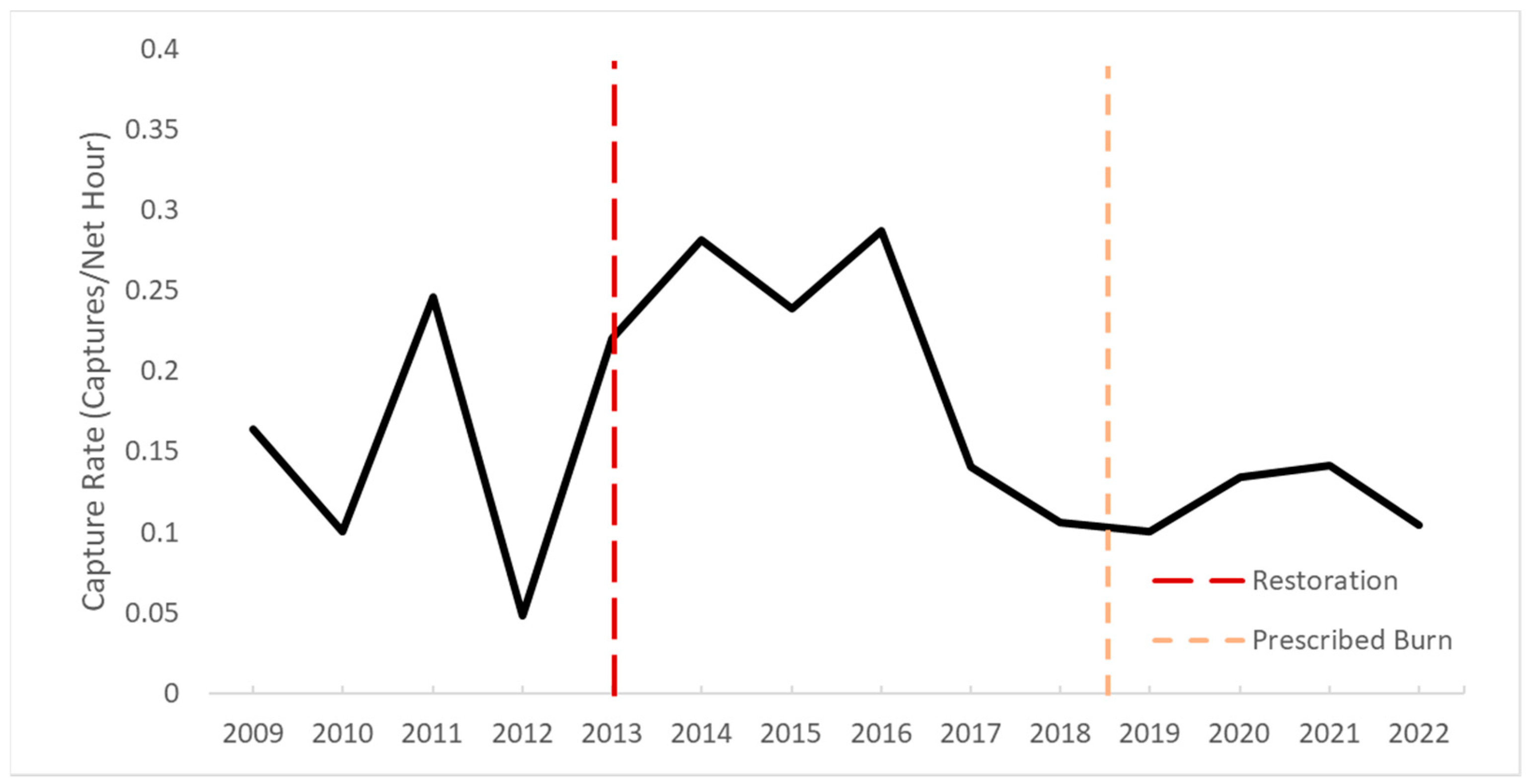

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pre-Restoration | Post-Restoration | |||||
|---|---|---|---|---|---|---|
| Species | Migration Strategy | Total Captures | Reproductive Index | Capture Rate | Reproductive Index | Capture Rate |
| American goldfinch * | Short distance | 651 | 0.155 ± 0.043 ‡ | 0.026 ± 0.02 | 0.718 ± 0.177 ‡ | 0.046 ± 0.02 |
| Common yellowthroat * | Long distance | 839 | 0.169 ± 0.037 | 0.026 ± 0.019 | 0.451 ± 0.076 | 0.08 ± 0.019 |
| Field sparrow * | Short distance | 1213 | 0.272 ± 0.102 ‡ | 0.045 ± 0.043 | 0.892 ± 0.16 ‡ | 0.095 ± 0.023 |
| Gray catbird * | Long distance | 475 | 0.877 ± 0.259 | 0.006 ± 0.005 | 0.5 ± 0.123 | 0.012 ± 0.004 |
| Indigo bunting * | Long distance | 841 | 0.071 ± 0.038 | 0.009 ± 0.063 | 0.063 ± 0.026 | 0.047 ± 0.026 |
| Magnolia warbler † | Long distance | 167 | - | 0.016 ± 0.009 | - | 0.024 ± 0.008 |
| Northern cardinal * | Resident | 393 | 0.5 ± 0.217 | 0.012 ± 0.011 | 0.355 ± 0.166 | 0.004 ± 0.006 |
| Song sparrow * | Short distance | 684 | 1.657 ± 0.366 | 0.052 ± 0.007 | 1.352 ± 1.202 | 0.064 ± 0.014 |
| Swamp sparrow † | Medium distance | 137 | - | 0.004 ± 0.007 | - | 0.016 ± 0.007 |
| Tennessee warbler † | Long distance | 302 | - | 0.018 ± 0.024 | - | 0.04 ± 0.02 |
| White-throated sparrow † | Medium distance | 115 | - | 0.012 ± 0.008 | - | 0.017 ± 0.005 |
| Yellow-breasted chat * | Long distance | 306 | 0.088 ± 0.049 | 0.001 ± 0.002 | 0.094 ± 0.025 | 0.003 ± 0.002 |
| Species | Precision | Migration Strategy | φ | φ se | φ lcl | φ ucl |
|---|---|---|---|---|---|---|
| American goldfinch | Marginal | Short distance | 0.346 | 0.076 | 0.214 | 0.506 |
| Common yellowthroat | Good | Long distance | 0.274 | 0.053 | 0.183 | 0.388 |
| Field sparrow | Good | Short distance | 0.469 | 0.058 | 0.359 | 0.583 |
| Gray catbird | Good | Long distance | 0.236 | 0.070 | 0.126 | 0.397 |
| Indigo bunting | Good | Long distance | 0.382 | 0.061 | 0.271 | 0.507 |
| Northern cardinal | Marginal | Resident | 0.581 | 0.076 | 0.428 | 0.719 |
| Song sparrow | Marginal | Short distance | 0.215 | 0.131 | 0.057 | 0.556 |
| Yellow-breasted chat | Good | Long distance | 0.282 | 0.073 | 0.161 | 0.444 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slankard, K.G.; Leffew, B.; Patton, M.D.; Baird, L.K.; Miller, J.; Stoelb, D.; Pelly, D.; Barnard, J.O.; Taylor, L.; Burford, L.S. The Effects of Native Grass and Shrub Restoration on Nesting and Fall Migrant Songbirds in Central Kentucky. Diversity 2024, 16, 615. https://doi.org/10.3390/d16100615
Slankard KG, Leffew B, Patton MD, Baird LK, Miller J, Stoelb D, Pelly D, Barnard JO, Taylor L, Burford LS. The Effects of Native Grass and Shrub Restoration on Nesting and Fall Migrant Songbirds in Central Kentucky. Diversity. 2024; 16(10):615. https://doi.org/10.3390/d16100615
Chicago/Turabian StyleSlankard, Kate G., Benjamin Leffew, Michael D. Patton, Laura K. Baird, Jessica Miller, Daniel Stoelb, Don Pelly, James O. Barnard, Loren Taylor, and Laura S. Burford. 2024. "The Effects of Native Grass and Shrub Restoration on Nesting and Fall Migrant Songbirds in Central Kentucky" Diversity 16, no. 10: 615. https://doi.org/10.3390/d16100615
APA StyleSlankard, K. G., Leffew, B., Patton, M. D., Baird, L. K., Miller, J., Stoelb, D., Pelly, D., Barnard, J. O., Taylor, L., & Burford, L. S. (2024). The Effects of Native Grass and Shrub Restoration on Nesting and Fall Migrant Songbirds in Central Kentucky. Diversity, 16(10), 615. https://doi.org/10.3390/d16100615