

Unrecognised Ant Megadiversity in the Australian Monsoonal Tropics III: The Meranoplus ajax Forel Complex



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
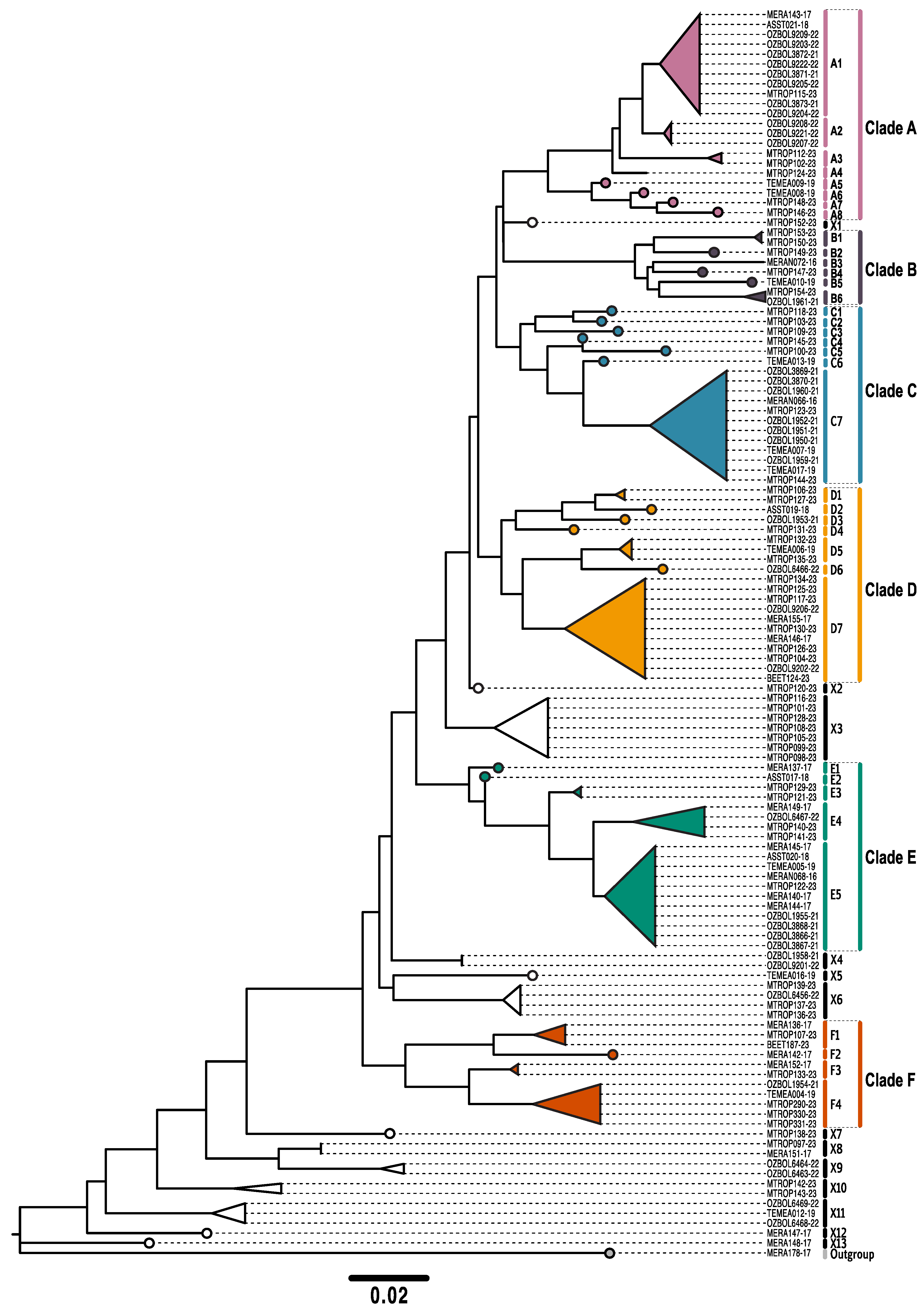
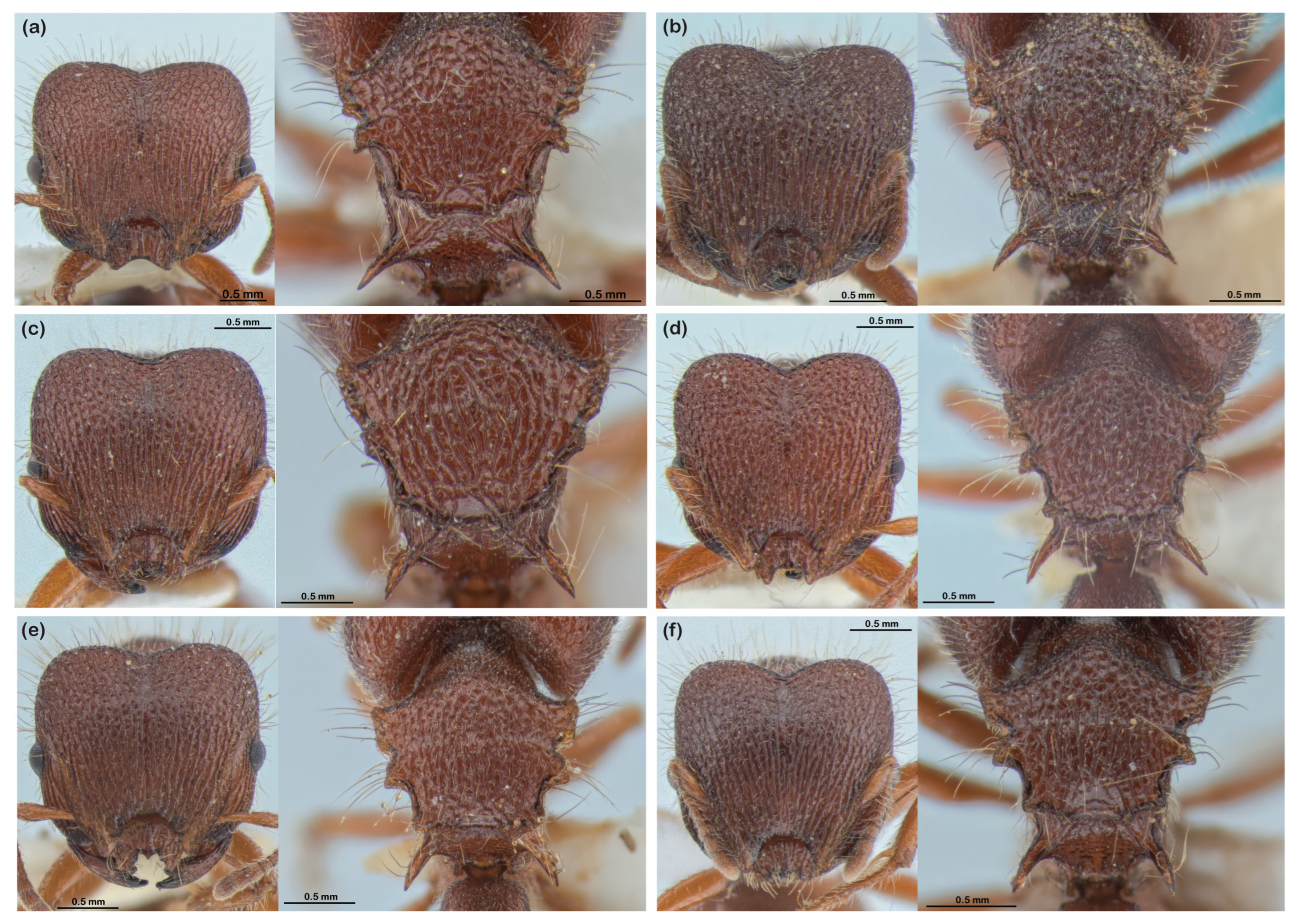
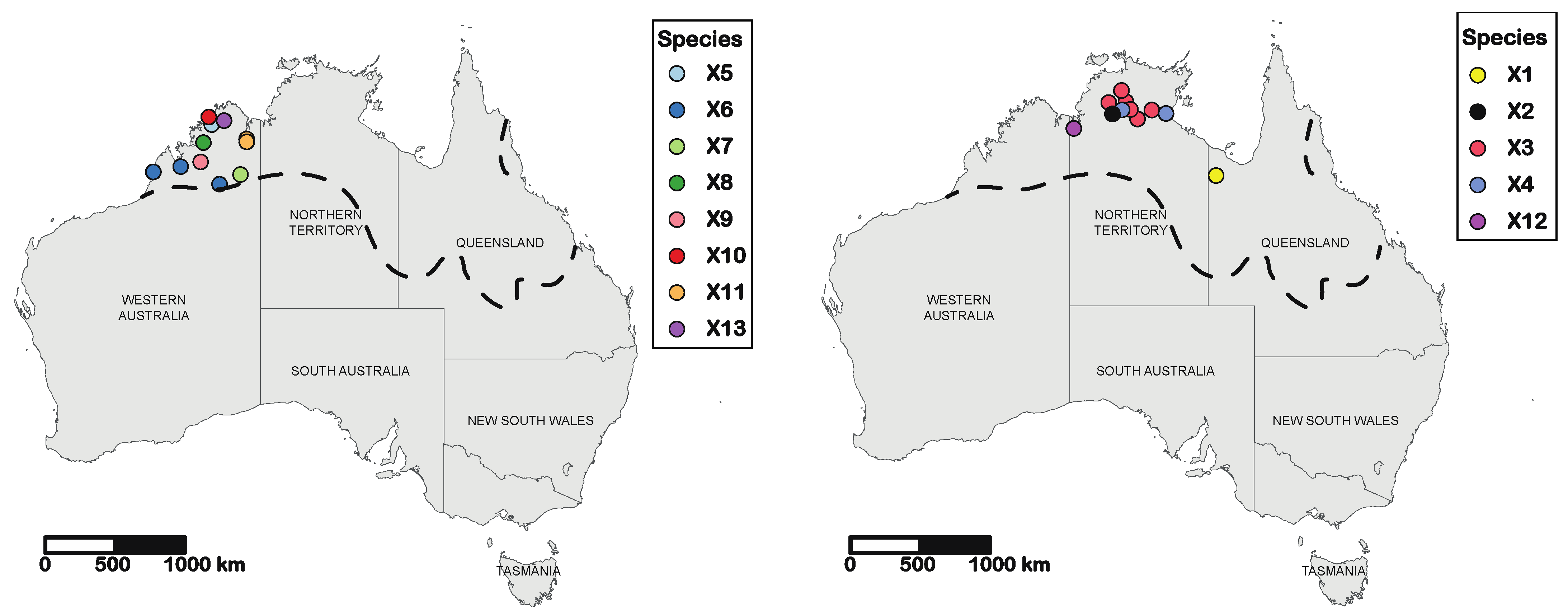
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kass, J.M.; Guénard, B.; Dudley, K.L.; Jenkins, C.N.; Azuma, F.; Fisher, B.L.; Parr, C.L.; Gibb, H.; Longino, J.Y.; Ward, P.S.; et al. The global distribution of known and undiscovered ant biodiversity. Sci. Adv. 2022, 8, eabp9908. [Google Scholar] [CrossRef] [PubMed]
- Andersen, A. The Ants of Northern Australia: A Guide to the Monsoonal Fauna; CSIRO Publishing: Collingwood, VIC, Australia, 2000; ISBN 0643066039. [Google Scholar]
- Oberprieler, S.K.; Andersen, A.N.; Moritz, C.C. Ants in Australia’s monsoonal tropics: CO1 barcoding reveals extensive unrecognised diversity. Diversity 2018, 10, 36. [Google Scholar] [CrossRef]
- Andersen, A.N.; Vasconcelos, H.L. Historical biogeography shapes functional ecology: Inter-continental contrasts in responses of savanna ant communities to stress and disturbance. J. Biogeogr. 2022, 49, 590–599. [Google Scholar] [CrossRef]
- Heterick, B.H.; Castalanelli, M.; Shattuck, S.O. Revision of the ant genus Melophorus (Hymenoptera, Formicidae). ZooKeys 2017, 700, 1–420. [Google Scholar] [CrossRef] [PubMed]
- Heterick, B.E. Revision of the Australian ants of the genus Monomorium (Hymenoptera: Formicidae). Invertebr. Syst. 2001, 15, 353–459. [Google Scholar] [CrossRef]
- Andersen, A.N.; Hoffmann, B.D.; Oberprieler, S.K. Megadiversity in the ant genus Melophorus: The M. rufoniger Heterick, Castalanelli and Shattuck species group in the top end of Australia’s Northern Territory. Diversity 2020, 12, 386. [Google Scholar] [CrossRef]
- Andersen, A.N.; Brassard, F.; Hoffmann, B.D. Unrecognised ant megadiversity in monsoonal Australia: The Melophorus hirsutipes Heterick, Castalanelli & Shattuck group. Diversity 2023, 15, 892. [Google Scholar]
- Andersen, A.N.; Brassard, F.; Hoffmann, B.D. Ant megadiversity in monsoonal Australia: Diversity and distribution in the hyperdiverse Monomorium nigrius Forel group. Diversity 2022, 14, 46. [Google Scholar] [CrossRef]
- Andersen, A.N.; Hoffmann, B.D.; Brassard, F. Unrecognised ant megadiversity in monsoonal Australia: The Tetramorium spininode Bolton group in the Northern Territory. Diversity 2023, 15, 476. [Google Scholar] [CrossRef]
- Andersen, A.N. A systematic overview of Australian species of the myrmicine ant genus Meranoplus F. Smith, 1893 (Hymenoptera: Formicidae). Myrmecol. Nachrichten 2006, 8, 157–170. [Google Scholar]
- Schödl, S. Revision of Australian Meranoplus: The Meranoplus diversus group. In Advances in Ant (Hymenoptera: Formicidae) Systematics: Homage to E. O. Wilson-50 Years of Contributions; Snelling, R.R., Fisher, B.L., Ward, P.S., Eds.; The American Entomological Institute: Gainesville, FL, USA, 2007; Volume 80, pp. 370–424. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.A.; Fisher, B.L.; Hebert, P.D.N. DNA barcoding for effective biodiversity assessment of a hyperdiverse arthropod group: The ants of Madagascar. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 1825–1834. [Google Scholar] [CrossRef] [PubMed]
- Schlick-Steiner, B.C.; Steiner, F.M.; Moder, K.; Seifert, B.; Sanetra, M.; Dyreson, E.; Stauffer, C.; Christian, E. A multidisciplinary approach reveals cryptic diversity in Western Palearctic Tetramorium ants (Hymenoptera: Formicidae). Mol. Phylogenet. Evol. 2006, 40, 259–273. [Google Scholar] [CrossRef] [PubMed]
- Bickford, D.; Lohman, D.J.; Sodhi, N.S.; Ng, P.K.L.; Meier, R.; Winker, K.; Krista, K.; Ingram, K.K.; Das, I. Cryptic species as a window on diversity and conservation. Tree 2007, 22, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Andrew, M.H. Granivory of the annual grass Sorghum intrans by the harvester ant Meranoplus sp. in tropical Australia. Biotropica 1986, 18, 344–349. [Google Scholar] [CrossRef]
- Andersen, A.N.; Azcárate, F.M.; Cowie, I.D. Seed selection by an exceptionally rich community of harvester ants in the Australian seasonal tropics. J. Anim. Ecol. 2000, 69, 975–984. [Google Scholar] [CrossRef]
- Andersen, A.N.; Hoffmann, B.D.; Oberprieler, S. Diversity and biogeography of a species-rich ant fauna of the Australian seasonal tropics. Insect Sci. 2018, 25, 519–526. [Google Scholar] [CrossRef]
- Feitosa, R.M.; Camacho, G.P.; Silva, T.S.; Ulysséa, M.A.; Ladino, N.; Oliveira, A.M.; Albuquerque, E.Z.; Schmidt, F.A.; Ribas, C.R.; Nogueira, A.; et al. Ants of Brazil: An overview based on 50 years of diversity studies. Syst. Biodivers. 2020, 20, 1–27. [Google Scholar] [CrossRef]
- Andrade-Silva, J.; Baccaro, F.B.; Prado, L.P.; Guénard, B.; Warren, D.L.; Kass, J.M.; Evan, P.; Economo, E.P.; Silva, R.R. A large-scale assessment of ant diversity across the Brazilian Amazon Basin: Integrating geographic, ecological and morphological drivers of sampling bias. Ecography 2022, 2022, e06295. [Google Scholar] [CrossRef]
- Hanisch, P.E.; Lavinia, P.D.; Suarez, A.V.; Lijtmaer, D.A.; Leponce, M.; Paris, C.I.; Tubaro, P.L. Mind the gap! Integrating taxonomic approaches to assess ant diversity at the southern extreme of the Atlantic Forest. Ecol. Evol. 2017, 7, 10451–10466. [Google Scholar] [CrossRef]
- Brühl, C.A.; Gunsalam, G.; Linsenmair, K.E. Stratification of ants (Hymenoptera, Formicidae) in a primary rain forest in Sabah, Borneo. J. Trop. Ecol. 1998, 14, 285–297. [Google Scholar] [CrossRef]
- Wilkie, K.T.R.; Mertl, A.L.; Traniello, J.F.A. Species diversity and distribution patterns of the ants of Amazonian Ecuador. PLoS ONE 2010, 5, e13146. [Google Scholar]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andersen, A.N.; Brassard, F.; Hoffmann, B.D. Unrecognised Ant Megadiversity in the Australian Monsoonal Tropics III: The Meranoplus ajax Forel Complex. Diversity 2024, 16, 126. https://doi.org/10.3390/d16020126
Andersen AN, Brassard F, Hoffmann BD. Unrecognised Ant Megadiversity in the Australian Monsoonal Tropics III: The Meranoplus ajax Forel Complex. Diversity. 2024; 16(2):126. https://doi.org/10.3390/d16020126
Chicago/Turabian StyleAndersen, Alan N., François Brassard, and Benjamin D. Hoffmann. 2024. "Unrecognised Ant Megadiversity in the Australian Monsoonal Tropics III: The Meranoplus ajax Forel Complex" Diversity 16, no. 2: 126. https://doi.org/10.3390/d16020126
APA StyleAndersen, A. N., Brassard, F., & Hoffmann, B. D. (2024). Unrecognised Ant Megadiversity in the Australian Monsoonal Tropics III: The Meranoplus ajax Forel Complex. Diversity, 16(2), 126. https://doi.org/10.3390/d16020126