
Dissimilarity among Species and Higher Taxa of Amphibians in a Hotspot of Biodiversity and Endemism in the Neotropics


 ,
,
Abstract
1. Introduction
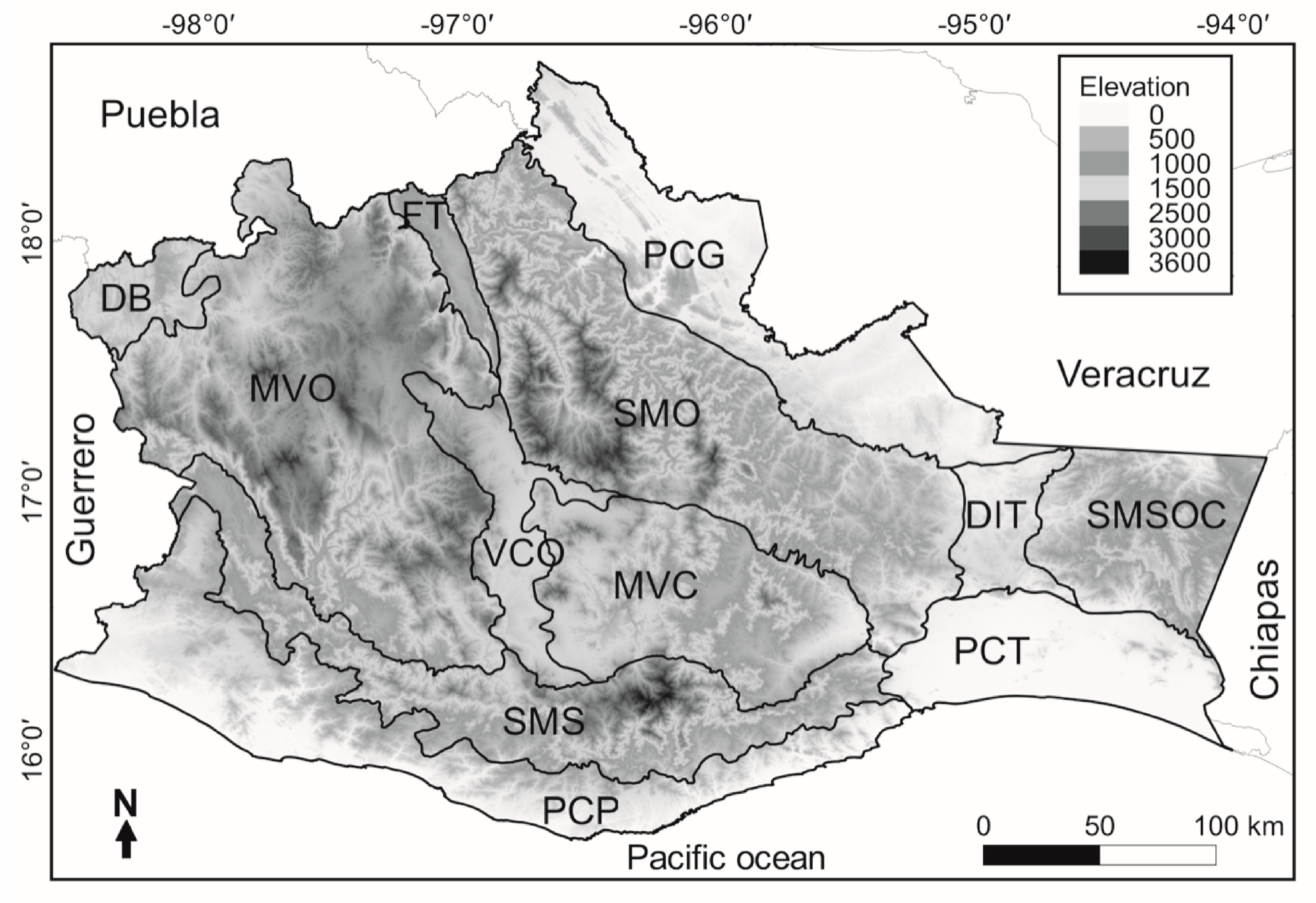
2. Materials and Methods
3. Results
3.1. Species Richness and Endemism
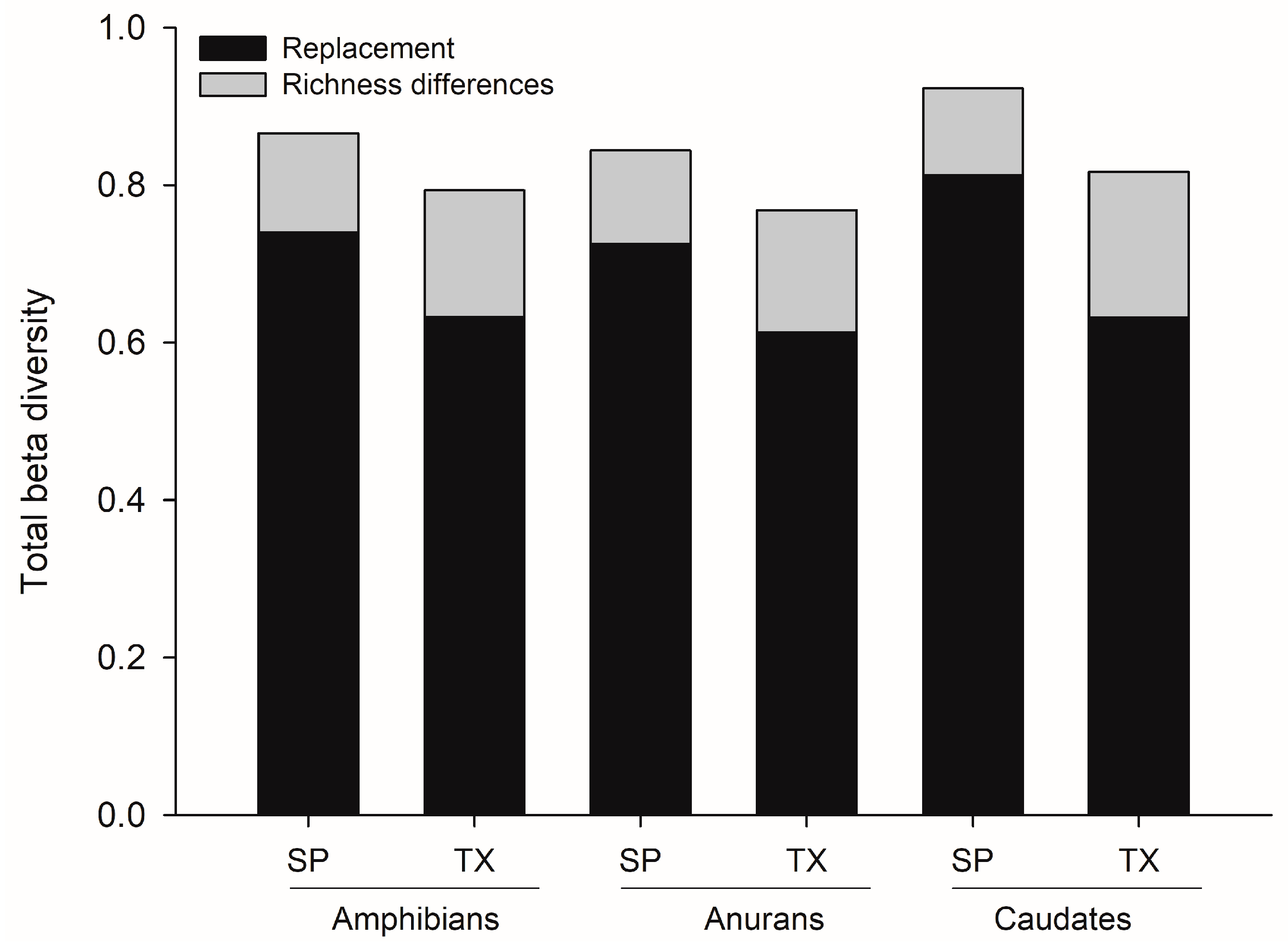
3.2. Total Beta Diversity of Amphibian Species at the State Level
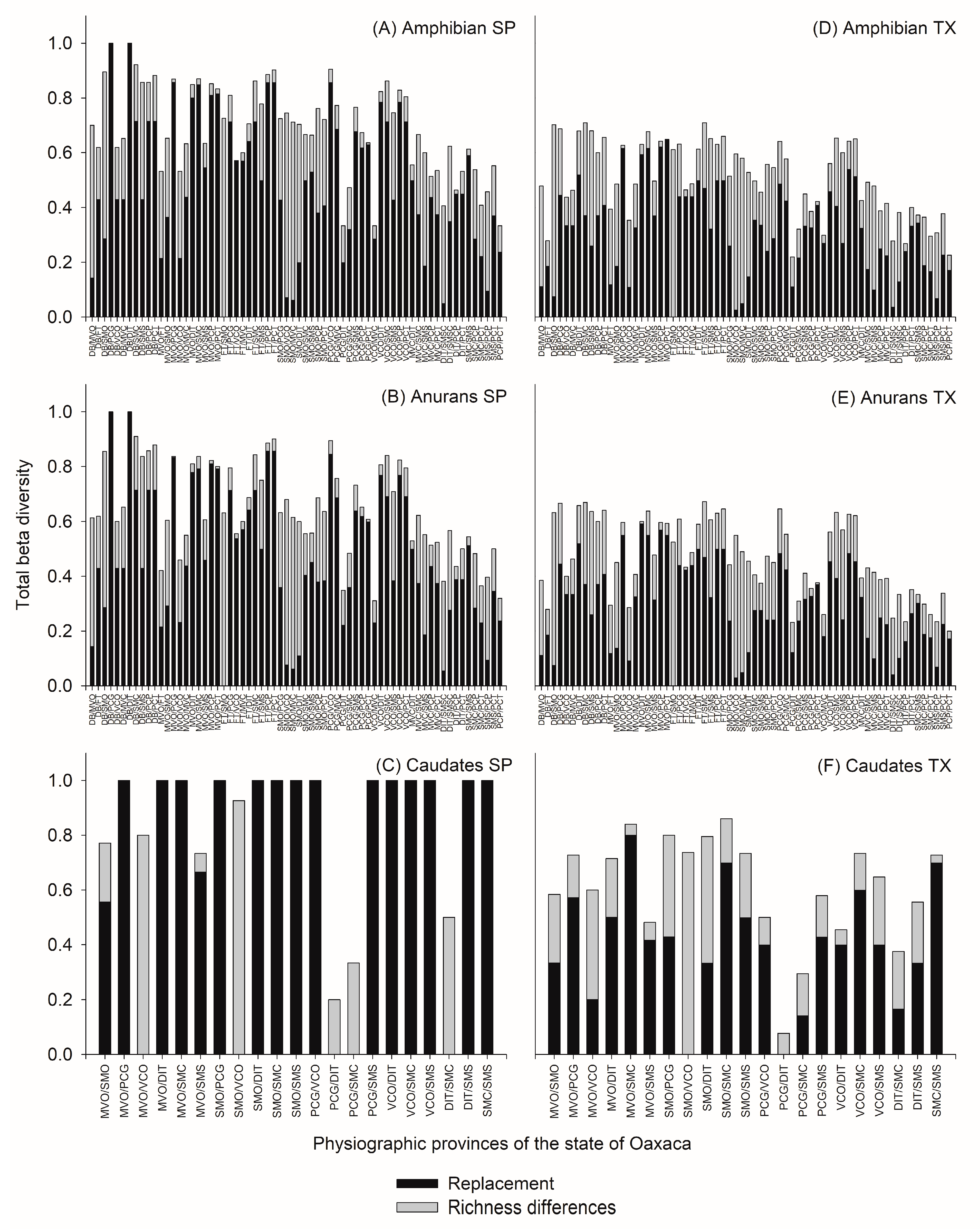
3.3. Beta Diversity of Amphibians between Pairwise of Physiographic Subprovinces
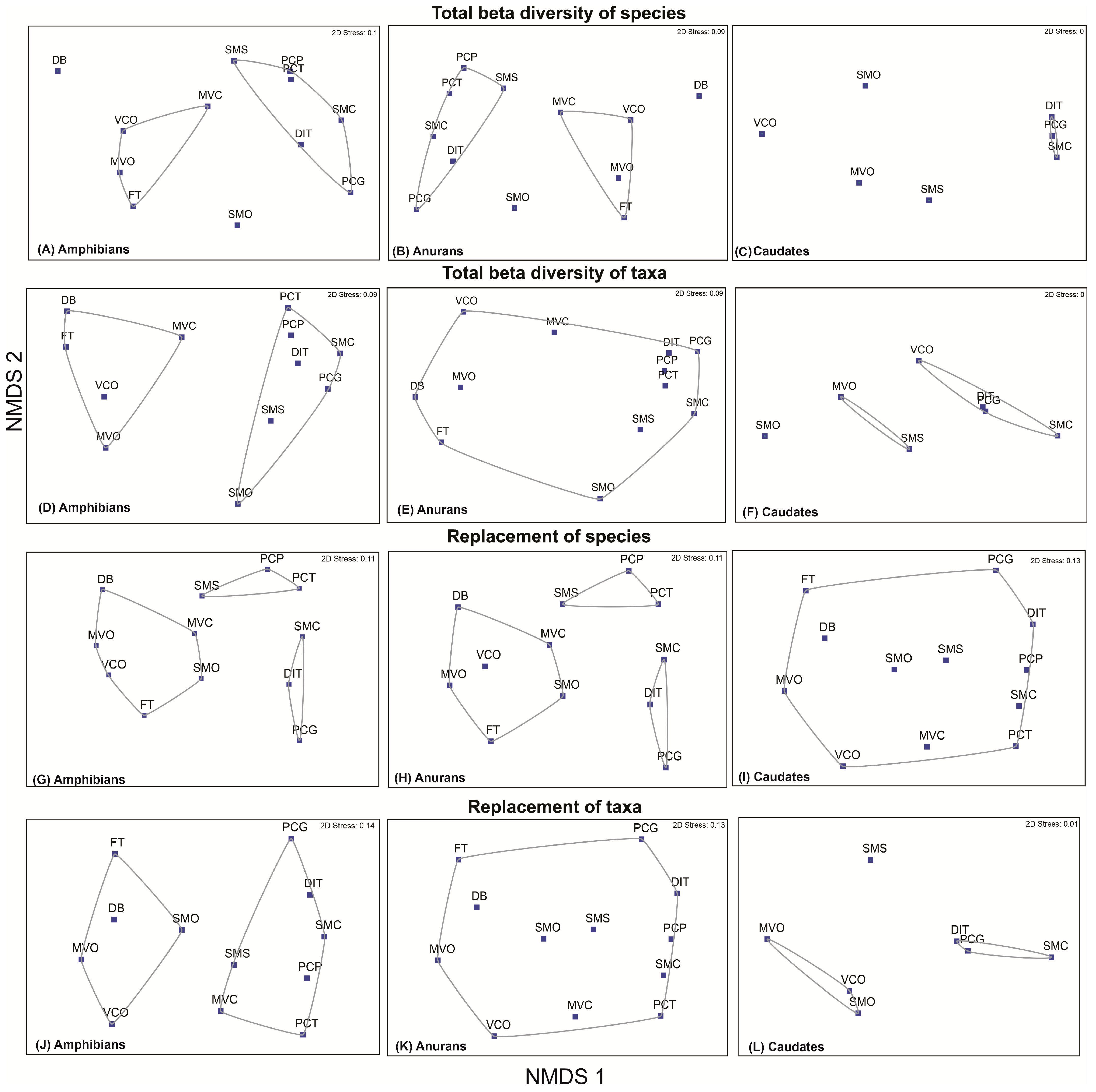
3.4. Determination of Beta Diversity of Species and Higher Taxa
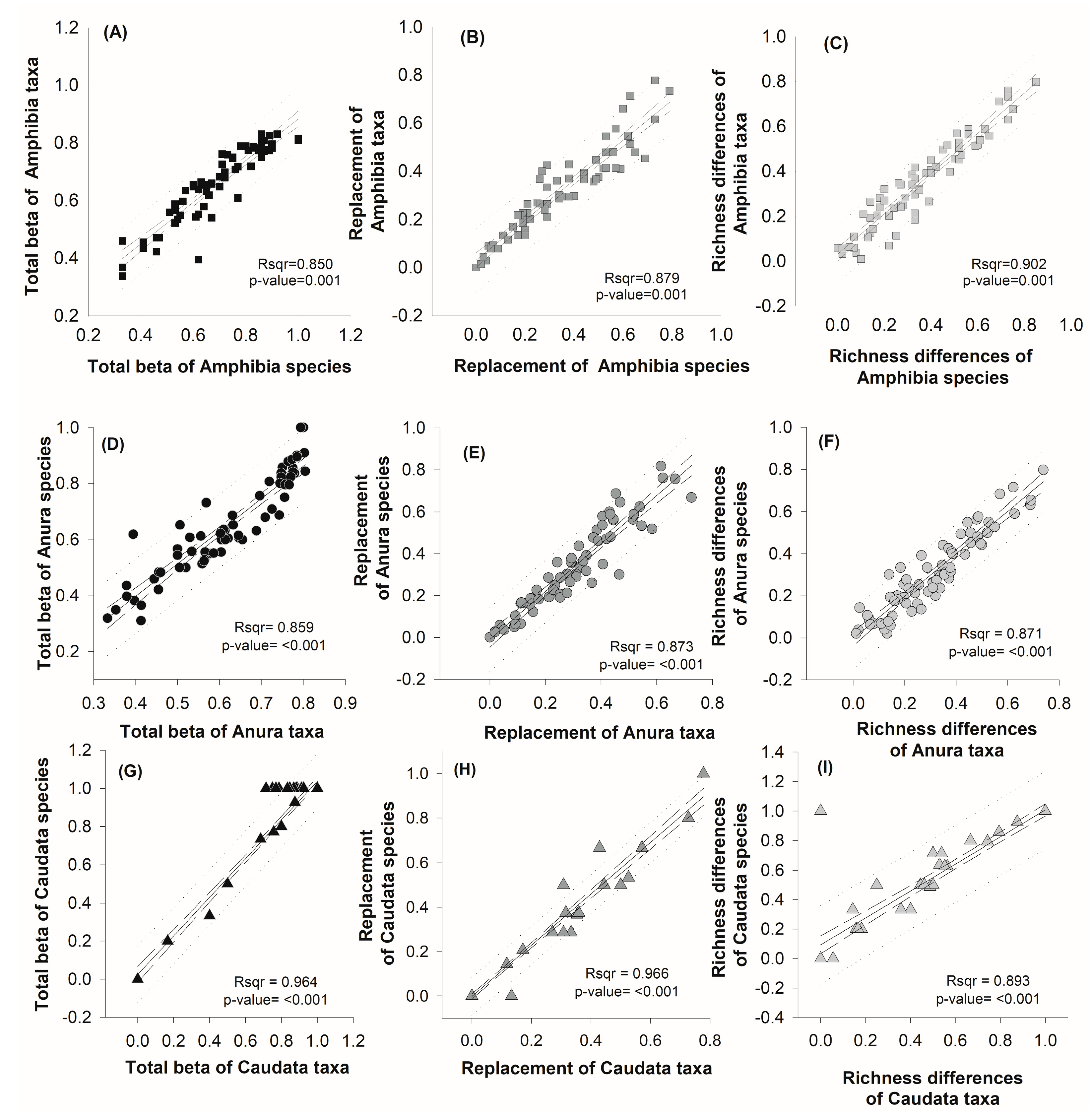
3.5. Relationship between Beta Diversity of Species and Higher Taxa of amphibians from Oaxaca
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ramírez-Bautista, A.; Torres-Hernández, L.A.; Cruz-Elizalde, R.; Berriozabal-Islas, C.; Hernández-Salinas, U.; Wilson, L.D.; Johson, J.D.; Porras, L.W.; Balderas-Valdivia, C.J.; Gonzalez-Hernández, A.J.X.; et al. An updated list of the Mexican herpetofauna: With a summary of historical and contemporary studies. ZooKeys 2023, 1166, 287–306. [Google Scholar] [CrossRef]
- Mata-Silva, V.; García-Padilla, E.; Rocha, A.; Desantis, D.L.; Johnson, J.D.; Ramírez-Bautista, A.; Wilson, L.D. A reexamination of the herpetofauna of Oaxaca, Mexico: Composition update, physiographic distribution, and conservation commentary. Zootaxa 2021, 4996, 201–252. [Google Scholar] [CrossRef]
- Parra-Olea, G.; Flores-Villela, O.; Mendoza-Almeralla, C. Biodiversity of amphibians in Mexico. Rev. Mex. Biodivers. 2014, 85, 460–466. [Google Scholar] [CrossRef]
- Wake, D.; Vredenburg, V.T. Are we in the midst of the sixth mass extinction? A view from the world of amphibians. Proc. Natl. Acad. Sci. USA 2008, 115, 11466–11473. [Google Scholar] [CrossRef] [PubMed]
- García-Bañuelos, P.; Rovito, S.M.; Pineda, E. Representation of Threatened Biodiversity in Protected Areas and Identification of Complementary Areas for Their Conservation: Plethodontid Salamanders in Mexico. Trop. Conserv. Sci. 2019, 12, 1940082919834156. [Google Scholar] [CrossRef]
- Lips, K.R.; Burrowes, P.A.; Mendelson, J.R.; Parra Olea, G. Amphibian population declines in latin America: A synthesis. Biotropica 2005, 37, 222–226. [Google Scholar] [CrossRef]
- Koleff, P.; Soberón, J.; Arita, H.; Dávila, P.; Flores-Villela, O.; Halffter, G.; Lira-Noriega, A.; Moreno, C.E.; Moreno, E.; Munguía, M.; et al. Patrones de diversidad espacial en grupos selectos de especies. In Capital Natural de México, Vol. I: Conocimiento Actual de la Biodiversidad; Ciudad de México: México, Spain, 2008; pp. 323–364. [Google Scholar]
- Ochoa-Ochoa, L.M.; Munguía, M.; Lira-Noriega, A.; Sánchez-Cordero, V.; Flores-Villela, O.; Navarro-Sigüenza, A.; Rodríguez, P. Spatial scale and β-diversity of terrestrial vertebrates in Mexico. Rev. Mex. Biodivers. 2014, 85, 918–930. [Google Scholar] [CrossRef]
- Rodríguez, P.; Ochoa-Ochoa, L.M.; Munguía, M.; Sánchez-Cordero, V.; Navarro-Sigüenza, A.; Flores-Villela, O.; Nakamura, M. Environmental heterogeneity explains coarse-scale β-diversity of terrestrial vertebrates in Mexico. PLoS ONE 2019, 14, e0210890. [Google Scholar] [CrossRef]
- Baselga, A.; Gómez-Rodríguez, C.; Lobo, J.M. Historical legacies in world amphibian diversity revealed by the turnover and nestedness components of beta diversity. PLoS ONE 2012, 7, e32341. [Google Scholar] [CrossRef] [PubMed]
- Mata-Silva, V.; Johnson, J.D.; Wilson, D.; García-Padilla, E. The herpetofauna of Oaxaca, Mexico: Composition, physiographic distribution, and conservation status. Mesoamerican Herpetol. 2015, 2, 6–62. [Google Scholar]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Baselga, A. The relationship between species replacement, dissimilarity derived from nestedness, and nestedness. Glob. Ecol. Biogeogr. 2012, 21, 1223–1232. [Google Scholar] [CrossRef]
- Carvalho, J.C.; Cardoso, P.; Gomes, P. Determining the relative roles of species replacement and species richness differences in generating beta-diversity patterns. Glob. Ecol. Biogeogr. 2012, 21, 760–771. [Google Scholar] [CrossRef]
- Carvalho, J.C.; Cardoso, P.; Borges, P.A.V.; Schmera, D.; Podani, J. Measuring fractions of beta diversity and their relationships to nestedness: A theoretical and empirical comparison of novel approaches. Oikos 2013, 122, 825–834. [Google Scholar] [CrossRef]
- Baselga, A.; Leprieur, F. Comparing methods to separate components of beta diversity. Methods Ecol. Evol. 2015, 6, 1069–1079. [Google Scholar] [CrossRef]
- Cardoso, P.; Rigal, F.; Carvalho, J.C. BAT—Biodiversity Assessment Tools, an R package for the measurement and estimation of alpha and beta taxon, phylogenetic and functional diversity. Methods Ecol. Evol. 2015, 6, 232–236. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. Computer Software v4.03. ISBN 3-900051-07-0. 2018. Available online: https://www.r-project.org/ (accessed on 27 March 2021).
- Bacaro, G.; Ricotta, C.; Mazzoleni, S. Measuring beta-diversity from taxonomic similarity. J. Veg. Sci. 2007, 18, 793–798. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Primer: User Manual/Tutoriall; Primer-E Ltd.: Plymouth, UK, 2015. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. 2019. Available online: https://www.iucnredlist.org/resources/summary-statistics.2019 (accessed on 20 January 2021).
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Rosenzweig, M.L. Species Diversity in Space and Time; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Gaston, K.J. Global Patterns in Biodiversity. Nature 2000, 405, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Casas-Andreu, G. Amphibians and reptiles of the Marías Islands and other adjacent islands off the coast of Nayarit, Mexico. Aspects about its biogeography and conservation. An. Del Inst. De Biol. Ser. Zool. 1992, 63, 95–112, (Spanish version). [Google Scholar]
- Calderón-Patrón, J.M. Island Biogeography: The Case of the Mexican Herpetofauna. Master’s Thesis, Universidad Autónoma del Estado de Hidalgo, México, Spain, 2007. (Spanish version). [Google Scholar]
- Bell, K.E.; Donnelly, M.A. Influence of forest fragmentation on community structure of frogs and lizards in northeastern Costa Rica. Conserv. Biol. 2006, 20, 1750–1760. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Guzmán, E.; Reynoso, V.H. Amphibian and reptile communities of rainforest fragments: Minimum patch size to support high richness and abundance. Biodivers. Conserv. 2012, 21, 3243–3265. [Google Scholar] [CrossRef]
- Russildi, G.; Arroyo-Rodríguez, V.; Hernández-Ordóñez, O.; Pineda, E.; Reynoso, V.H. Species- and community-level responses to habitat spatial changes in fragmented rainforests: Assessing compensatory dynamics in amphibians and reptiles. Biodivers. Conserv. 2016, 25, 375–392. [Google Scholar] [CrossRef]
- Vitt, L.J.; Caldwell, J.P. Herpetology: An Introductory Biology of Amphibians and Reptiles, 4th ed.; Elsevier Inc.: Norman, OK, USA, 2014. [Google Scholar]
- Shi, X.; Zhao, Y.; Chen, C.H.; Ren, P.; Zeng, D.; Wu, L.; Ding, P. Beta-diversity partitioning: Methods, applications and perspectives. Biodivers. Sci. 2017, 25, 464–480. [Google Scholar]
- Calderón-Patrón, J.M.; Moreno, C.E.; Pineda-López, R.; Sánchez-Rojas, G.; Zuria, I. Vertebrate dissimilarity due to turnover and richness differences in a highly beta-diverse region: The role of spatial grain size, dispersal ability and distance. PLoS ONE 2014, 8, e82905. [Google Scholar] [CrossRef]
- Calderón-Patrón, J.M.; Goyenechea, I.; Ortiz-Pulido, R.; Castillo-Cerón, J.; Manríquez, N.; Ramírez-Bautista, A.; Rojas-Martínez, A.E.; Sánchez-Rojas, G.; Zuria, I.; Moreno, C.E. Beta diversity in a highly heterogeneous area: Disentangling species and taxonomic dissimilarity for terrestrial vertebrates. PLoS ONE 2016, 11, e0160438. [Google Scholar] [CrossRef]
- Serrano, J.M.; Berlanga-Robles, C.A.; Ruíz Luna, A. High amphibian diversity related to unexpected environmental values in a biogeographic transitional area in north western Mexico. Contrib. Zool. 2014, 83, 151–166. [Google Scholar] [CrossRef]
- Juárez-Ramírez, C.M.; Aguilar-López, J.L.; Pineda, E. Protected natural areas and the conservation of amphibians in a highly transformed mountainous region in Mexico. Herpetol. Conserv. Biol. 2016, 11, 19–28. [Google Scholar]
- Aguilar-López, J.L.; Ortiz Lozada, L.; Pelayo-Martínez, J.; Mota-Vargas, C.; Alarcón-Villegas, L.E.; Demeneghi-Calatayud, A.P. Diversity and conservation of amphibians and reptiles in a private protected area of a highly transformed region in southern Veracruz, Mexico. Acta Zool. Mex. 2020, 36, 1–14, (Spanish version). [Google Scholar] [CrossRef]
- Calderón-Patrón, J.M.; Moreno, C.E.; Zuria, I. Diversidad beta: Medio siglo de avances. Rev. Mex. Biodivers. 2012, 83, 879–891. [Google Scholar] [CrossRef]
- Dalmolin, D.A.; Tozetti, A.M.; Pereira, M.J.R. Taxonomic and functional anuran beta diversity of a subtropical metacommunity respond differentially to environmental and spatial predictors. PLoS ONE 2019, 14, e0214902. [Google Scholar] [CrossRef] [PubMed]
- Alroy, J. Current extinction rates of reptiles and amphibians. Proc. Natl. Acad. Sci. USA 2015, 112, 13003–13008. [Google Scholar] [CrossRef] [PubMed]
- McCallum, M.L. Amphibian decline or extinction? Current declines dwarf background extinction rate. J. Herpetol. 2007, 41, 483–491. [Google Scholar] [CrossRef]
- SEMARNAT. Secretaria de Medio Ambiente y Recursos Naturales. In Mexican Official Standard NOM-059-SEMARNAT-2010, Environmental Protection–Mexican Native Species of Wild Flora and Fauna–Risk Categories and Specifications for Inclusion, Exclusion or Change–List of Species at Risk; Diario Oficial de la Federación 14 Noviembre: México, Spain, 2019; (Spanish version). [Google Scholar]
- Monroy-Gamboa, A.G.; Sánchez-Cordero, V.; Briones-Salas, M.; Lira-Sadee, R.; Maass-Moreno, J.M. Representativeness of vegetation types in different conservation initiatives in Oaxaca, Mexico. Bosque 2015, 36, 199–210, (Spanish version). [Google Scholar] [CrossRef][Green Version]
- Ramírez-Toro, W.; Torres-Miranda, A.; González-Rodríguez, A.; Ruíz-Sánchez, E.; Luna-Vega, I.; Oyama, K. A multicriteria analysis for prioritizing areas for conservation of oaks (Fagaceae: Quercus) in Oaxaca, Southern Mexico. Trop. Conserv. Sci. 2017, 10, 1940082917714227. [Google Scholar] [CrossRef]
- García-Mendoza, A. Integration of floristic knowledge of the State. In Biodiversity of Oaxaca; García-Mendoza, A., Ordoñez, M., Briones-Salas, M., Eds.; Instituto de Biología, UNAM-Fondo Oaxaqueño para la Conservación: México, Spain, 2004; pp. 305–325, (Spanish version). [Google Scholar]
- Contreras-Medina, R.; Luna-Vega, I. Species richness, endemism and conservation of Mexican gymnosperms. Biodivers. Conserv. 2007, 16, 1803–1821. [Google Scholar] [CrossRef]
- Lips, K.R.; Mendelson, J.R.; Muñoz-Alonso, A.; Canseco-Márquez, L.; Mulcahy, D.G. Amphibian population declines in montane southern Mexico: Resurveys of historical localities. Biol. Conserv. 2004, 119, 555–564. [Google Scholar] [CrossRef]
- Wake, D.B.; Papenfuss, T.J.; Lynch, J.F. Distribution of salamanders along elevational transects in Mexico and Guatemala. Tulane Publ. Zool. Bot. Suppl. Publ. 1991, 1, 303–319. [Google Scholar]
- Rovito, S.M.; Parra-Olea, G.; Vásquez-Almazán, C.R.; Papenfuss, T.J.; Wake, D.B. Dramatic declines in neotropical salamander populations are an important part of the global amphibian crisis. Proc. Natl. Acad. Sci. USA 2009, 106, 3231–3236. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Area | Species Richness | Endemic Species to Mexico and Oaxaca | ||||||
|---|---|---|---|---|---|---|---|---|
| Subprovinces | km2 | S Amphibia | S Anura | S Caudata | S Caecilians | Amphibia Mx/Oax | Anura Mx/Oax | Caudata Mx/Oax |
| DB | 1788.17 | 7 | 7 | 0 | 0 | 5/0 | 5/0 | 0/0 |
| MVO | 21,262.73 | 33 | 24 | 9 | 0 | 27/13 | 18/5 | 9/8 |
| FT | 1134.21 | 14 | 14 | 0 | 0 | 7/0 | 7/0 | 0/0 |
| SMO | 17,519.96 | 88 | 62 | 26 | 0 | 62/43 | 36/19 | 26/24 |
| PCG | 7975.92 | 28 | 25 | 3 | 0 | 4/1 | 2/1 | 2/0 |
| VCO | 2267.42 | 14 | 13 | 1 | 0 | 11/2 | 10/1 | 1/1 |
| MVC | 6662.62 | 16 | 16 | 0 | 0 | 8/1 | 8/1 | 0/0 |
| DIT | 2114.12 | 20 | 18 | 2 | 0 | 3/0 | 1/0 | 2/0 |
| SMC | 5816.08 | 44 | 37 | 6 | 1 | 11/01 | 7/0 | 4/1 |
| SMS | 12,350.15 | 49 | 42 | 6 | 1 | 32/12 | 25/6 | 7/6 |
| PCP | 9262.06 | 21 | 21 | 0 | 0 | 7/0 | 7/0 | 0/0 |
| PCT | 4298.77 | 27 | 26 | 0 | 1 | 6/1 | 6/1 | 0/0 |
| No. Subprovinces | No Endemics | Mexico Endemics | Oaxaca Endemics | Total |
|---|---|---|---|---|
| 1 | 10 | 23 | 48 | 81 |
| 2 | 12 | 1 | 10 | 23 |
| 3 | 5 | 8 | 2 | 15 |
| 4 | 4 | 1 | 0 | 5 |
| 5 | 4 | 3 | 0 | 7 |
| 6 | 3 | 2 | 0 | 5 |
| 7 | 4 | 3 | 0 | 7 |
| 8 | 1 | 1 | 0 | 2 |
| 9 | 1 | 0 | 0 | 1 |
| 10 | 3 | 0 | 0 | 3 |
| Total | 47 | 42 | 60 | 149 |
| SUBPROV | JAN | FEB | MAR | APR | MAY | JUNE | JULY | AUG | SEP | OCT | NOV | DEC | ANNUAL |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DB | 19.3/4 | 20.9/3 | 23.4/6 | 24.9/28 | 25.4/75 | 24.4/156 | 23.3/132 | 23.2/145 | 22.8/161 | 22.0/45 | 20.5/6 | 19.4/1 | 22.5/762 |
| MVO | 17.2/6 | 18.7/4 | 20.9/8 | 22.4/20 | 23.2/76 | 22.3/155 | 21.4/113 | 21.5/103 | 21.1/149 | 20.0/52 | 18.4/13 | 17.3/3 | 20.4/702 |
| FT | 21.3/3 | 22.6/3 | 25.5/5 | 27.9/12 | 28.5/33 | 27.8/104 | 26.1/113 | 26.3/84 | 26.1/101 | 24.9/30 | 22.9/8 | 21.7/4 | 25.1/500 |
| SMO | 13.5/14 | 14.9/10 | 16.8/13 | 17.3/33 | 17.9/54 | 16.7/171 | 15.9/171 | 16.0/152 | 15.8/178 | 14.9/88 | 13.9/45 | 14.1/22 | 15.6/951 |
| PCG | 21.4/37 | 22.3/41 | 24.4/38 | 27.2/36 | 28.4/91 | 28.0/402 | 26.6/516 | 27.2/377 | 26.7/448 | 21.2/225 | 23.2/81 | 21.9/56 | 24.9/2348 |
| VCO | 17.5/2 | 19.0/5 | 21.1/10 | 22.7/32 | 22.9/71 | 22.1/161 | 21.1/109 | 21.2/107 | 20.9/126 | 19.9/41 | 18.5/9 | 17.6/3 | 20.4/676 |
| MVC | 20.0/2 | 21.2/3 | 23.1/6 | 24.9/21 | 25.6/54 | 24.7/113 | 24.0/83 | 24.1/81 | 23.6/101 | 22.8/32 | 21.5/7 | 20.4/2 | 23.0/505 |
| DIT | 22.3/25 | 23.1/20 | 24.7/23 | 26.5/21 | 27.5/56 | 26.5/212 | 25.6/292 | 25.9/287 | 25.2/295 | 24.4/142 | 23.4/47 | 22.6/30 | 25.0/1450 |
| SMC | 21.8/107 | 22.9/56 | 24.7/34 | 26.4/39 | 27.3/80 | 26.3/273 | 25.6/302 | 25.9/351 | 25.5/462 | 24.4/284 | 23.1/154 | 22.2/116 | 25.0/2258 |
| SMS | 13.2/5 | 13.8/21 | 14.5/18 | 14.9/39 | 14.9/124 | 14.2/258 | 13.9/204 | 13.9/232 | 13.6/238 | 13.8/93 | 13.5/19 | 13.4/8 | 14.0/1259 |
| PCP | 26.1/3 | 26.5/7 | 27.2/2 | 27.8/8 | 28.6/65 | 28.1/163 | 28.0/138 | 28.2/249 | 27.9/217 | 27.2/83 | 26.4/8 | 26.0/3 | 27.3/946 |
| PCT | 24.8/4 | 25.6/4 | 27.1/3 | 29.0/6 | 29.6/55 | 28.6/201 | 28.3/132 | 28.7/160 | 27.7/237 | 27.1/58 | 26.1/14 | 25.2/5 | 27.3/879 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calderón-Patrón, J.M.; Peña-Joya, K.E.; Téllez-López, J.; Canales-Gómez, E.P. Dissimilarity among Species and Higher Taxa of Amphibians in a Hotspot of Biodiversity and Endemism in the Neotropics. Diversity 2024, 16, 224. https://doi.org/10.3390/d16040224
Calderón-Patrón JM, Peña-Joya KE, Téllez-López J, Canales-Gómez EP. Dissimilarity among Species and Higher Taxa of Amphibians in a Hotspot of Biodiversity and Endemism in the Neotropics. Diversity. 2024; 16(4):224. https://doi.org/10.3390/d16040224
Chicago/Turabian StyleCalderón-Patrón, Jaime Manuel, Karen Elizabeth Peña-Joya, Jorge Téllez-López, and Eréndira Patricia Canales-Gómez. 2024. "Dissimilarity among Species and Higher Taxa of Amphibians in a Hotspot of Biodiversity and Endemism in the Neotropics" Diversity 16, no. 4: 224. https://doi.org/10.3390/d16040224
APA StyleCalderón-Patrón, J. M., Peña-Joya, K. E., Téllez-López, J., & Canales-Gómez, E. P. (2024). Dissimilarity among Species and Higher Taxa of Amphibians in a Hotspot of Biodiversity and Endemism in the Neotropics. Diversity, 16(4), 224. https://doi.org/10.3390/d16040224