
Exploring Morphological Population Variability: Host Plant and Habitat Dependency in the Protected Moth Gortyna borelii (Lepidoptera, Noctuidae)
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Habitat Description
2.2. Plant Measurement
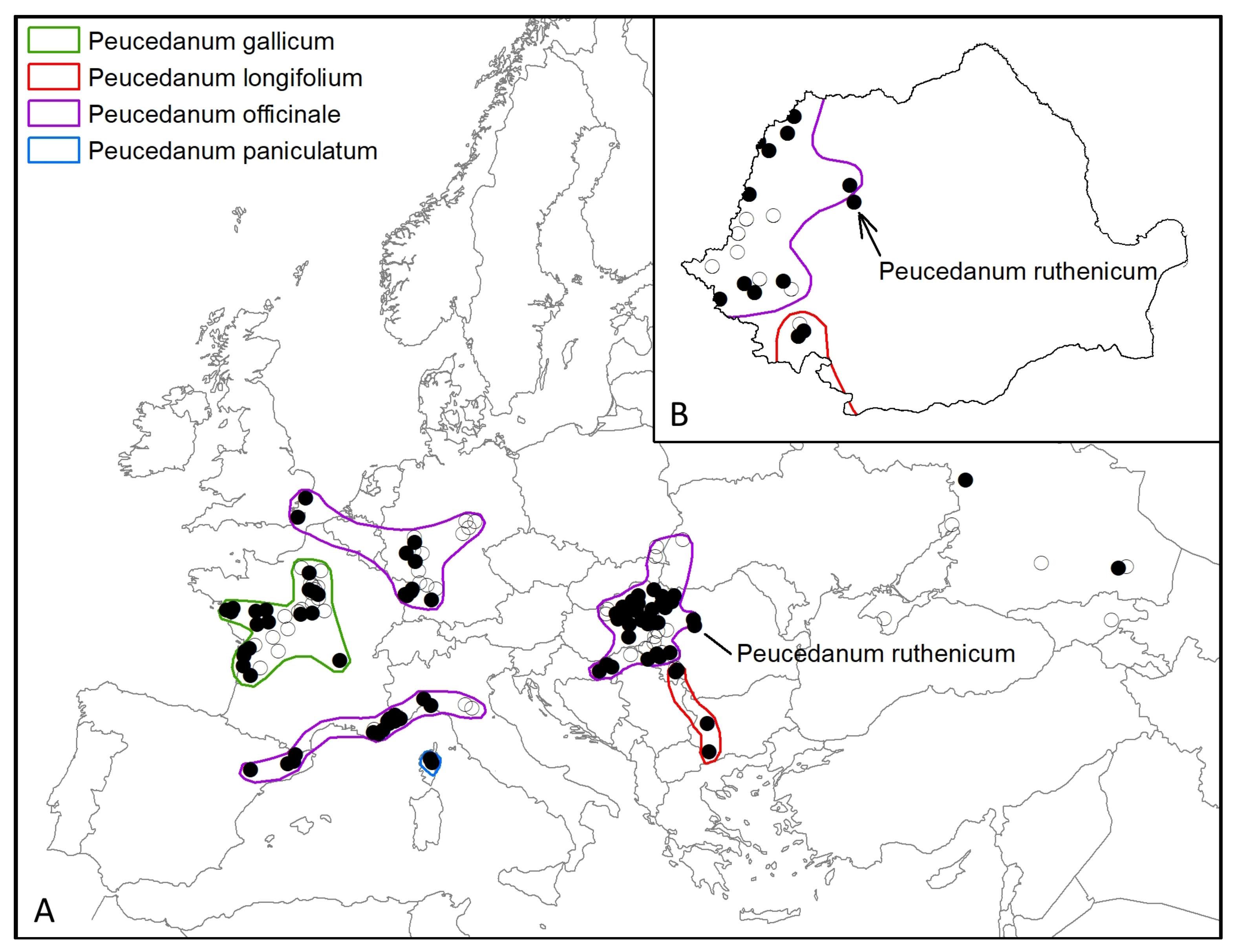
2.3. Distribution Analysis of G. borelii Populations Correlated with the Peucedanum Species
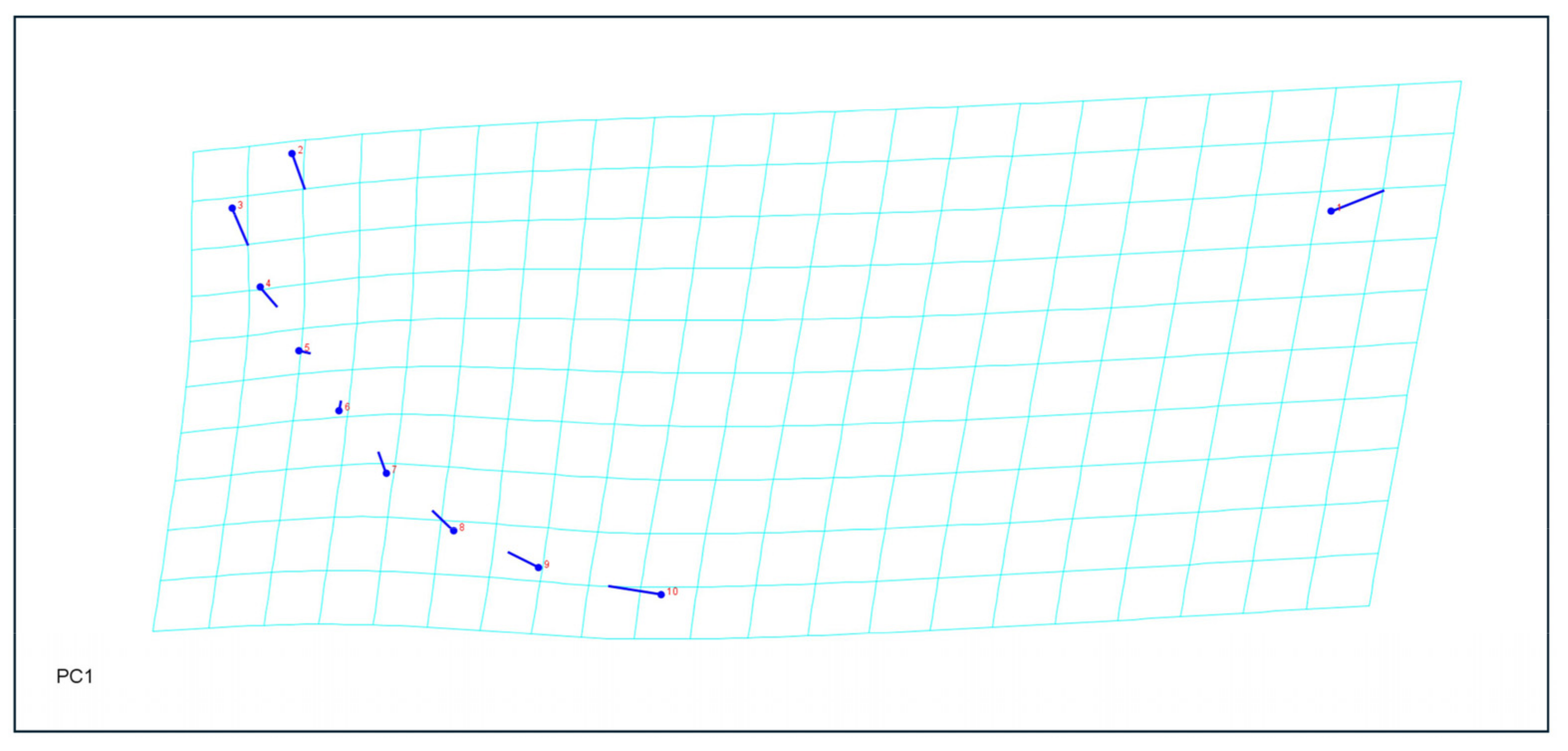
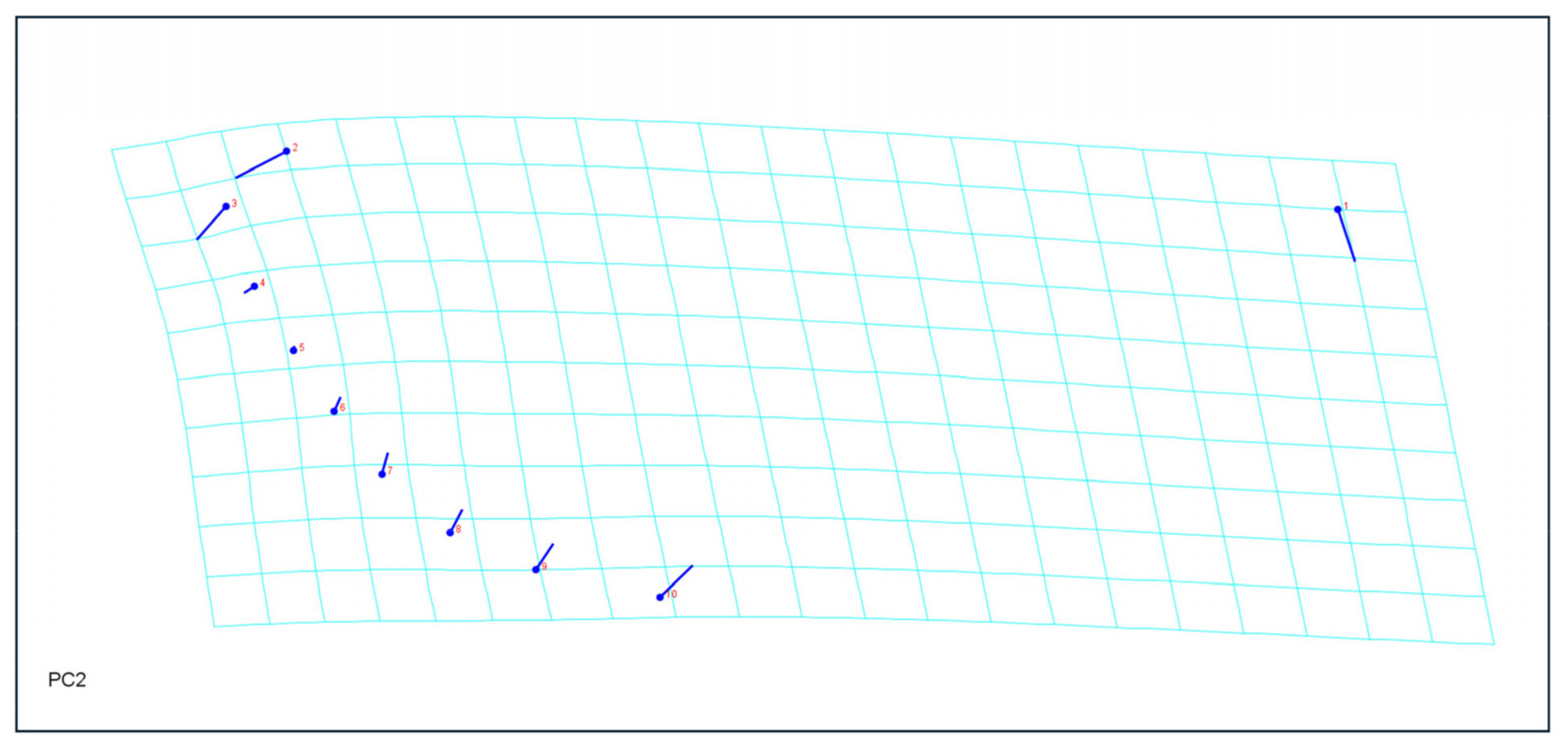
2.4. Morphometric Analyses
3. Results
3.1. Study Sites with New Population Identified in Romania
3.2. Host Plant Analyses
3.3. Distribution Analysis of G. borelii Populations Correlated with the Peucedanum Species
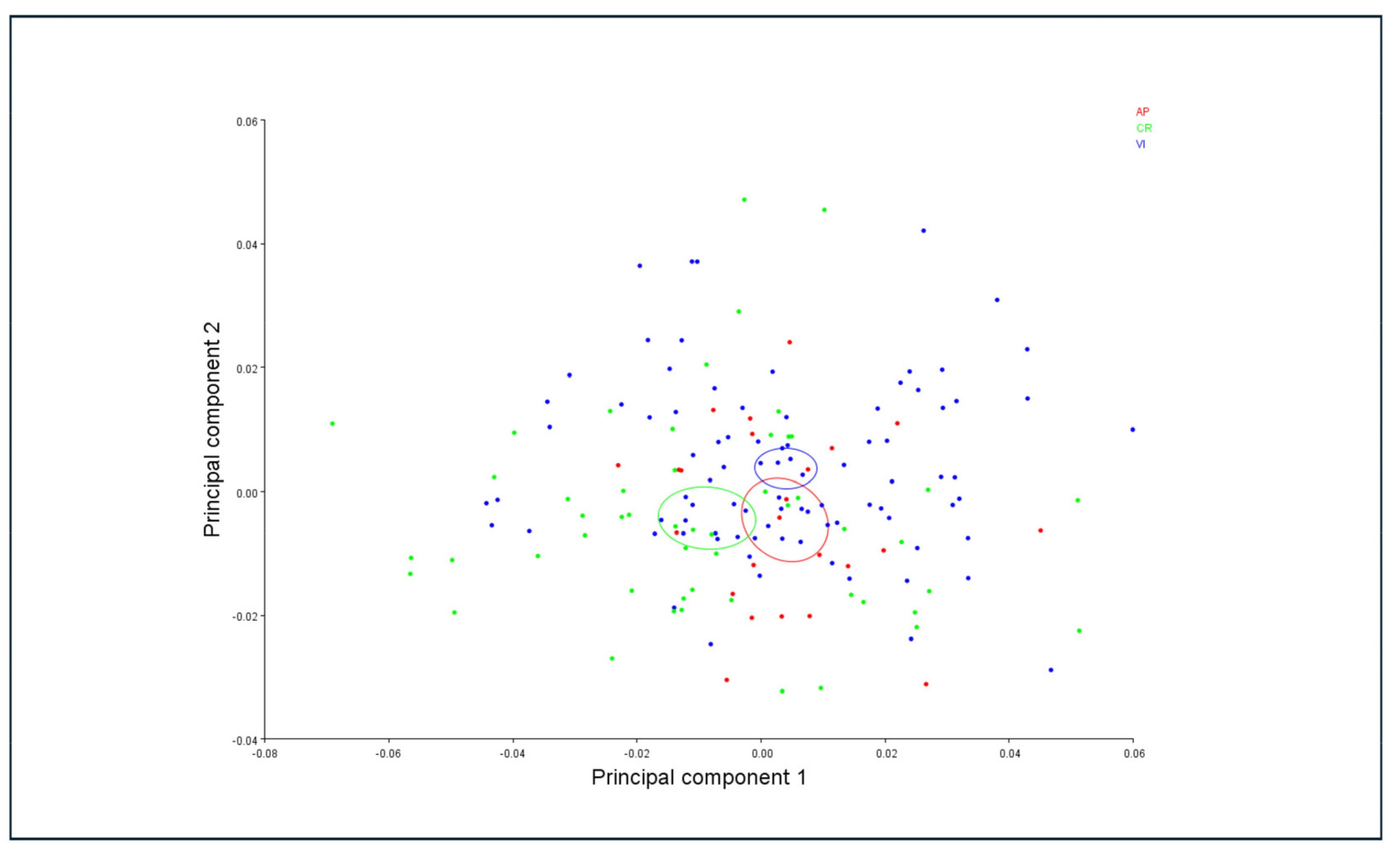
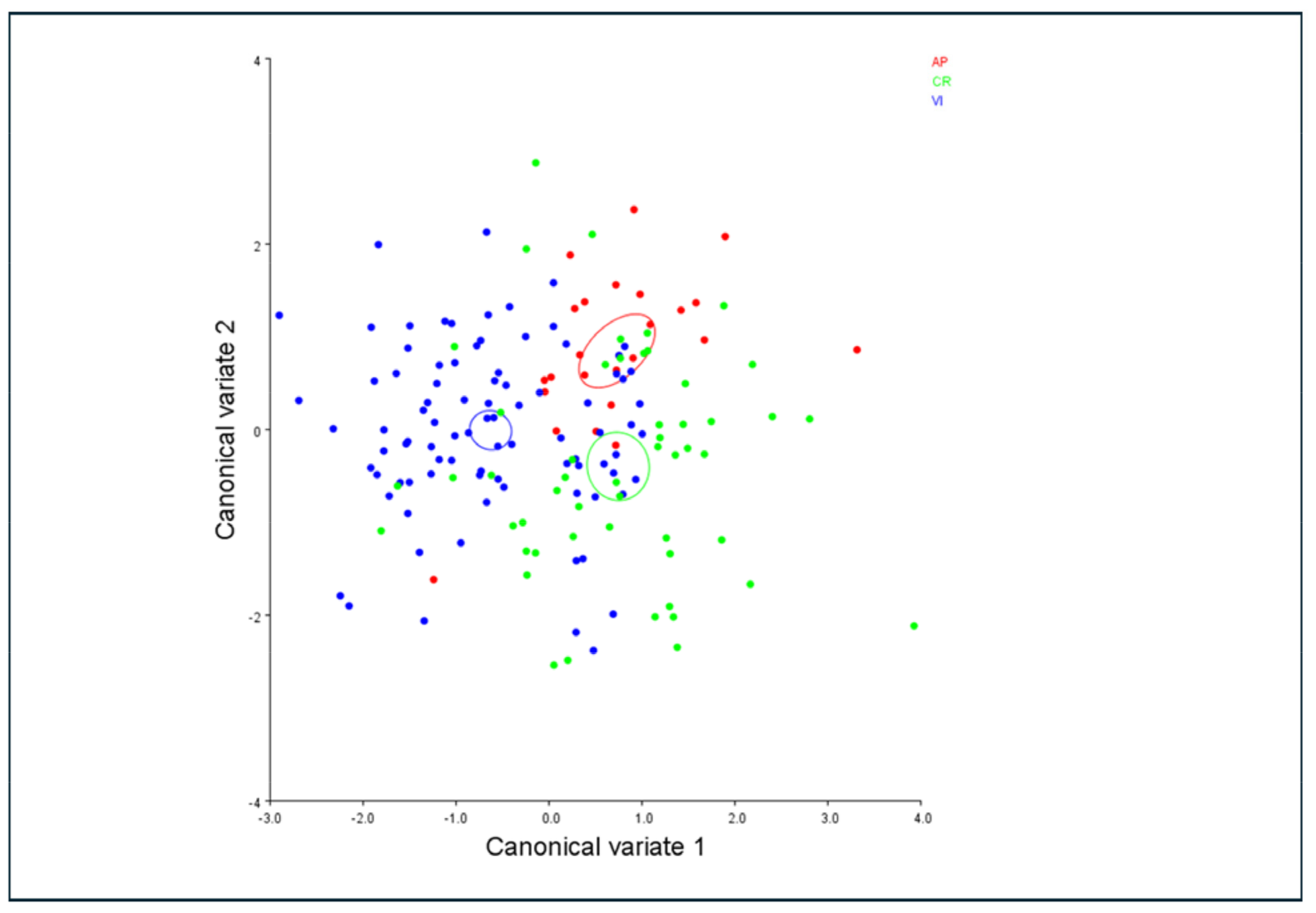
3.4. Morphometric Analysis
4. Discussion
4.1. Correlation between the Different Species of Peucedanum and the Distribution of G. borelii Populations, with Implications in Intraspecific Diversification
4.2. Morphological and Morphometric Differences in Relation to the Host Plant
4.3. The Newly Discovered Population of G. borelii and Its Importance for the Conservation of the Species
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Powell, S.K.; Spence, J.; Bharathi, M.; Gatehouse, A.J.; Gatehouse, R.M.A. Immunohistochemical and developmental studies to elucidate the mechanism of action of the snowdrop lectin on the rice brown planthopper, Nilaparvata lugens (Stal). J. Insect. Physiol. 1998, 44, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Menken, S.B.; Boomsma, J.J.; Van Nieukerken, E.J. Large-scale evolutionary patterns of host plant associations in the lepidoptera. Evolution 2010, 64, 1098–1119. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.A.; Zhang, X. The evolution of coevolution in the study of species interactions. Evolution 2021, 75, 1594–1606. [Google Scholar] [CrossRef] [PubMed]
- Forister, L.M.; Dyer, A.L.; Singer, S.M.; Stireman, O.J.; Lill, T.J. Revisiting the evolution of ecological specialization, with emphasison insect–plant interactions. Ecology 2012, 93, 981–999. [Google Scholar] [CrossRef] [PubMed]
- Hardy, N.B.; Kaczvinsky, C.; Bird, G.; Normark, B.B. Whatwe don’t know about diet-breadth evolution in herbivorous insects. Annu. Rev. Ecol. Evol. Syst. 2020, 51, 103–122. [Google Scholar] [CrossRef]
- Braga, P.M.; Janz, N. Host repertoires and changing insect–plant interactions. Ecol. Entomol. 2021, 46, 1241–1253. [Google Scholar] [CrossRef]
- Rákosy, L.; Corduneanu, C.; Crisan, A.; Dinca, V.; Kovács, S.; Stanescu, M.; Szekely, L. Red List of Lepidoptera of Romania; Rakosy, L., Ed.; Presa Universitara Clujeană: Cluj-Napoca, Romania, 2021; p. 187. ISBN 978-606-37-1126-8. [Google Scholar]
- Zilli, A.; Ronkay, L.; Fibiger, M. Noctuidae Europaeae: Apameini; Entomological Press: Faringdon, UK, 2005. [Google Scholar]
- Baranyi, T.; Varga, Z. Gortyna borelii (Pierret, 1837)—Nagy Szikibagolylepke. In The Living Heritage of the Pannonian Region—The Natura 2000 Network (in Hungarian); Varga, Z., Ed.; Dél-Nyírség-Bihari Tájvédelmi és Kulturális Értékõrzõ Egyesület: Debrecen, Hungary, 2014. [Google Scholar]
- Bator, D.; Guilloton, J.A. Contribution à la cartographie de Gortyna borelii (Pierret, 1837) en France. Alexanor 2015, 27, 7–87. [Google Scholar]
- Kovács, L. The Macrolepidoptera characteristic to our sandz district. Ann. Hist. Nat. Musei. Natl. Hung. 1955, 6, 327–342. [Google Scholar]
- König, F. Beiträge zur Kenntnis der Lebensweise von Hydraecia leucographa Bkh. Folia Entomol. Hung. 1959, 12, 481–493. [Google Scholar]
- König, F. Erfolgreiche Eizuchten von Hydroecia (Hydraecia) leucographa Bkh. Entomol. Z. 1960, 70, 69–75. [Google Scholar]
- Ippolito, R.; Parenzan, P. Contributo alla conoscenza della Gortyna Ochs. Europe. Entomol. 1978, 14, 159–202. [Google Scholar]
- Goater, B. Noctuidae (partim). In The Moths and Butterflies of Great Britain and Ireland; Heath, J., Emmet, A.M., Eds.; Harley Books: London, UK, 1983; Volume 10, pp. 36–413. [Google Scholar]
- Steiner, A. Bemerkungen über Gortyna borelii (Pierret, 1837) in Südwestdeutschland (Lepidoptera: Noctuidae). Entomol. Z. Mit Insektenbörse 1985, 95, 161–176. [Google Scholar]
- Steiner, A. Gortyna borelii. In Die Schmetterlinge Baden-Württembergs 7 Nachtfalter; Ebert, G., Ed.; V.E Ulmer Verlag: Stuttgart, Germany, 1998; pp. 79–86. [Google Scholar]
- Gyulai, P. Notes on the distribution of Gortyna borelii lunata in the Carpathian Basin. Nota Lepid. 1987, 10, 54–60. [Google Scholar]
- Gibson, C. The conservation of Gortyna borelii lunata Freyer (Lep.:Noctuidae). Entomol. Rec. J. Var. 2000, 112, 1–5. [Google Scholar]
- Hill, J.; Ringwood, Z.; Rouse, T. Distribution and status of Gortyna borelii Pierret ssp. lunata Freyer (Lep.: Noctuidae) in south-east England. Entomol. Rec. J. Var. 2002, 114, 49–53. [Google Scholar]
- Ringwood, Z.; Gardiner, T.; Steiner, A.; Hill, J. Comparison of factors influencing the habitat characteristics of Gortyna borelii (Noctuidae) and its larval food plant Peucedanum officinale in Britain and Germany. Nota Lepid. 2002, 25, 23–38. [Google Scholar]
- Ringwood, Z.; Hill, J.; Gibson, C. Conservation management of Gortyna borelii lunata (Lepidoptera: Noctuidae) in the United Kingdom. J. Insect Conserv. 2004, 8, 173–183. [Google Scholar] [CrossRef]
- Biewald, G. Gortyna borelii lunata Freyer, 1838. In Das Europäische Schutzgebietssystem Natura 2000. Ökologie und Verbreitung von Arten der FFH-Richtlinie in Deutschland. Band 3: Arten der EU-Osterweiterung; Schriftenreihe für Landschaftspflege und Naturschutz; Petersen, B., Ellwanger, G., Eds.; Landwirtschaftsverlag: Bonn, Germany, 2005; Volume 69, pp. 128–138. [Google Scholar]
- Sum, S. Gortyna borelii Pierret, 1837 (Nagy szikibagoly). In Natura 2000 Species and Habitats in Hungary; Haraszthy, L., Ed.; Pro Vértes Közalapitvány: Csákvár, Hungary, 2014; pp. 351–359. (In Hungarian) [Google Scholar]
- Berquier, C.; Mothiron, P.M.-C. Andrei-Ruiz. Bilan synthétique sur la connaissance de la Noctuelle des Peucédans en France Gortyna borelii (Pierret, 1827). Avancement des connaissances sur la répartition, l’écologie et l’état de conservation de Gorytna borelii en Corse (Lepidoptera Noctuidae Noctuinae Apameini). Alexanor 2016, 27, 95–112. [Google Scholar]
- Toussaint, E.F.A.; Condamine, F.L.; Kergoat, G.J.; Capdevielle-Dulac, C.; Barbut, J.; Silvain, J.F.; Le Ru, B.P. Palaeoenvironmental Shifts Drove the Adaptive Radiation of a Noctuid Stemborer Tribe (Lepidoptera, Noctuidae, Apameini) in the Miocene. PLoS ONE 2012, 7, e41377. [Google Scholar] [CrossRef]
- Ringwood, Z.K.; Hill, J.; Gibson, C. Observations on the ovipositing strategy of Gortyna borelii Pierret, 1837 (Lepidoptera, Noctuidae) in a British population. Acta Zool. Acad. Sci. Hung. 2002, 48, 89–99. [Google Scholar]
- Cini, A.; Bordoni, A.; Ghisolfi, G.; Lazzaro, L.; Platania, L.; Pasquali, L.; Negroni, R.; Benetello, F.; Coppi, A.; Zanichelli, F.; et al. Host plant selection and differential survival on two Aristolochia, L. species in an insular population of Zerynthia cassandra. J. Insect Conserv. 2019, 23, 239–246. [Google Scholar] [CrossRef]
- García-Barros, E.; Fartmann, T. Butterfly oviposition: Sites, behaviour and modes. In Ecology of Butterflies in Europe; Settele, J., Shreeve, T., Konvička, M., van Dick, H., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 29–42. [Google Scholar]
- Gripenberg, S.; Mayhew, J.P.; Parnell, M.; Roslin, T. A meta-analysis of preference–performance relationships in phytophagous insects. Ecol. Lett. 2010, 13, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Regier, J.C.; Mitter, C.; Kristensen, N.P.; Davis, D.R.; Van Nieukerken, E.J.; Rota, J.; Simonsen, J.T.; Mitter, K.T.; Kawahara, A.Y.; Yen, S.-H.; et al. A molecular phylogeny for the oldest (nonditrysian) lineages of extant L epidoptera, with implications for classification, comparative morphology and life-history evolution. Syst. Entomol. 2015, 40, 671–704. [Google Scholar] [CrossRef]
- Diamond, S.E.; Blair, C.P.; Abrahamson, W.G. Testing the nutrition hypothesis for the adaptive nature of insect galls: Does a non-adapted herbivore perform better in galls? Ecol. Entomol. 2008, 33, 385–393. [Google Scholar] [CrossRef]
- Tooker, J.F.; De Moraes, C.M. A gall-inducing caterpillar species increases essential fatty acid content of its host plant without concomitant increases in phytohormone levels. Mol. Plant Microbe Interact. 2009, 22, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Giron, D.; Huguet, E.; Stone, N.G.; Body, M. Insect-induced effects on plants and possible effectors used by galling and leaf-mining insects to manipulate their host-plant. J. Insect Physiol. 2016, 84, 70–89. [Google Scholar] [CrossRef] [PubMed]
- Connor, E.F.; Taverner, M.P. The Evolution and Adaptive Significance of the Leaf-Mining Habit. Oikos 1997, 79, 6–25. [Google Scholar] [CrossRef]
- Price, P.W.; Fernandes, G.W.; Warring, G.L. Adaptive nature of insect galls. Environ. Entomol. 1987, 16, 15–24. [Google Scholar] [CrossRef]
- Denno, R.F.; McClure, M.S.; Ott, J.R. Interspecific interactions in phytophagous insects: Competition reexamined and resurrected. Annu. Rev. Entomol. 1995, 40, 297–331. [Google Scholar] [CrossRef]
- Tooker, J.F.; Giron, D. The Evolution of Endophagy in Herbivorous Insects. Front. Plant Sci. 2020, 11, 581816. [Google Scholar] [CrossRef]
- Nosil, P.; Vines, T.H.; Funk, D.J. Perspective: Reproductive isolation caused by natural selection against immigrants from divergent habitats. Evolution 2005, 59, 705–719. [Google Scholar] [CrossRef] [PubMed]
- Nosil, P.; Harmon, L.J.; Seehausen, O. Ecological explanations for (incomplete) speciation. Trends Ecol. Evol. 2009, 24, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Fiteni, E.; Durand, K.; Gimenez, S.; Meagher, R.L., Jr.; Legeai, F.; Kergoat, G.J.; Nègre, N.; d’Alençon, E.; Nam, K. Host-plant adaptation as a driver of incipient speciation in the fall armyworm (Spodoptera frugiperda). BMC Ecol. Evol. 2022, 22, 133. [Google Scholar] [CrossRef] [PubMed]
- Gloss, D.A.; Abbot, P.; Whiteman, K.N. How interactions with plant chemicals shape insect genomes. Curr. Opin. Insect. Sci. 2019, 36, 149–156. [Google Scholar] [CrossRef]
- Simon, C.-J.; d’ Alencon, E.; Guy, E.; Jacquin-Joly, E.; Jaquiéry, J.; Nouhaud, P.; Peccoud, J.; Sugio, A.; Streiff, R. Genomics of adaptation to host-plants in herbivorous insects. Brief. Funct. Genom. 2015, 14, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Fischer, H.M.; Wheat, C.W.; Heckel, D.G.; Vogel, H. Evolutionary origins of a novel host plant detoxification gene in butterflies. Mol. Biol. Evol. 2008, 25, 809–820. [Google Scholar] [CrossRef]
- Kulmuni, J.; Wurm, Y.; Pamilo, P. Comparative genomics of chemosensory protein genes reveals rapid evolution and positive selection in ant-specific duplicates. Heredity 2013, 110, 538–547. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Skepticism towards Santa Rosalia, or why are there so few kinds of animals? Evolution 1981, 35, 124–138. [Google Scholar] [CrossRef]
- Gavrilets, S. Models of Speciation: Where Are We Now? J. Hered. 2014, 105, 743–755. [Google Scholar] [CrossRef]
- Tessnow, A.E.; Raszick, T.J.; Porter, P.; Sword, G.A. Patterns of genomic and allochronicstrain divergence in the fall armyworm, Spodoptera frugiperda (J.E. Smith). Ecol. Evol. 2022, 12, e8706. [Google Scholar] [CrossRef]
- Johannesson, K. Parallel speciation: A key to sympatric divergence. Trends Ecol. Evol. 2001, 16, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Turelli, M.; Barton, H.N.; Coyne, A.J. Theory and speciation. Trends Ecol. Evol. 2001, 16, 330–343. [Google Scholar] [CrossRef] [PubMed]
- Drès, M.; Mallet, J. Host races in plant-feeding insects and their importance in sympatric speciation. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2002, 357, 471–492. [Google Scholar] [CrossRef] [PubMed]
- Feder, J.L.; Roethele, F.B.; Filchak, K.; Niedbalski, J.; Romero-Sever-son, J. Evidence for inversion polymorphism related tosympatric host race formation in the apple maggot fly, Rhagoletis pomonella. Genetics 2003, 163, 939–953. [Google Scholar] [CrossRef]
- Peccoud, J.; Ollivier, A.; Plantegenest, M.; Simon, J.C. A continuum of genetic divergence from sympatric host races to species in the pea aphid complex. Proc. Natl. Acad. Sci. USA 2009, 106, 7495–7500. [Google Scholar] [CrossRef] [PubMed]
- Berlocher, S.H.; Feder, J.L. Sympatric speciation in phytophagous insects: Moving beyond controversy? Annu. Rev. Entomol. 2002, 47, 773–815. [Google Scholar] [CrossRef]
- Feder, J.L.; Xie, X.; Rull, J.; Velez, S.; Forbes, A.; Leung, B.; Dambroski, H.; Filchak, K.E.; Aluja, M. Mayr, Dobzhansky, and Bush and the complexities of sympatric speciation in Rhagoletis. Proc. Natl. Acad. Sci. USA 2005, 102, 6573–6580. [Google Scholar] [CrossRef]
- Feder, J.L.; Forbes, A.A. Habitat Avoidance and Speciation for Phytophagous Insect Specialists. Funct. Ecol. 2007, 21, 585–597. [Google Scholar] [CrossRef]
- Forbes, A.A.; Fisher, J.; Feder, J.L. Habitat avoidance: Overlooking an important aspect of host-specific mating and sympatric speciation? Evolution 2005, 59, 1552–1559. [Google Scholar]
- Forbes, A.A.; Powell, T.H.Q.; Stelinski, L.L.; Smith, J.J.; Feder, J.L. Sequential sympatric speciation across trophic levels. Science 2009, 323, 776–779. [Google Scholar] [CrossRef]
- Lin, A.P.; Chan, P.W.; Cai, L.; Dankowicz, E.; Gilbert, K.; Pierce, N.; Felton, G. The links between plant volatiles and host plant specialization of herbivores. Res. Sq. 2022, 2–38. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Cornell, H.V.; Hochberg, M.E. Predators, parasitoids, and pathogens as mortality agents in phytophagous insect populations. Ecology 1997, 78, 2145–2152. [Google Scholar] [CrossRef]
- Stone, G.N.; Schönrogge, K. The adaptive significance of insect gall morphology. Trends Ecol. Evol. 2003, 18, 512–522. [Google Scholar] [CrossRef]
- Body, M.; Burlat, V.; Giron, D. Hypermetamorphosis in a leaf-miner allows insects to cope with a confined nutritional space. Arthropod-Plant Interact. 2015, 9, 75–84. [Google Scholar] [CrossRef]
- Gutzwiller, F.; Dedeine, F.; Kaiser, W.; Giron, D.; Lopez-Vaamonde, C. Correlation between the green-island phenotype and Wolbachia infections during the evolutionary diversification of Gracillariidae leaf-mining moths. Ecol. Evol. 2015, 5, 4049–4062. [Google Scholar] [CrossRef]
- Rákosy, L. Die Noctuiden Rumäniens (Lepidoptera: Noctuidae); Stapfia 46: Linz, Austria, 1996; p. 648. [Google Scholar]
- Bookstein, F.L. Size and shape spaces for Landmark data in two dimensions. Stat. Sci. 1986, 1, 181–242. [Google Scholar] [CrossRef]
- Zelditch, M.L.; Swiderski, D.L.; Sheets, H.D.; Fink, W.L. Geometric Morphometrics for Biologists: A Primer; Elsevier Academic Press: Cambridge, MA, USA, 2004; p. 444. [Google Scholar]
- Baranyi, T.; Korompai, T.; Józsa Á, C.s.; Kozma, P. Gortyna borelii lunata (Freyer, 1838). In Natura 2000 Species Studies I; Varga, Z., Ed.; Dél-Nyírség Bihari Tájvédelmi és Kulturális Értékırzı Egyesület: Debrecen, Hungary, 2006; pp. 3–69. [Google Scholar]
- Grimaldi, D. The co-radiations of pollinating insects and angiosperms in the Cretaceous. Ann. Mo. Bot. Gard. 1999, 86, 373–406. [Google Scholar] [CrossRef]
- Janz, N. Ehrlich and Raven revisited: Mechanisms underlying codiversification of plants and enemies. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 71–89. [Google Scholar] [CrossRef]
- Joy, J.B. Symbiosis catalyses niche expansion and diversification. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122820. [Google Scholar] [CrossRef]
- Darwell, C.T.; Fox, K.A.; Althoff, D.M. The roles of geography and founder effects in promoting host-associated differentiation in the generalist bogus yucca moth Prodoxus decipiens. J. Evol. Biol. 2014, 27, 2706–2718. [Google Scholar] [CrossRef]
- Gavrilets, S. Fitness Landscapes and the Origin of Species (MPB-41); Princeton University Press: Princeton, NJ, USA, 2004; Volume 41. [Google Scholar]
- Bereczki, J.; Póliska, S.; Váradi, A.; Tóth, J.P. Incipient sympatric speciation via host race formation in Phengaris arion (Lepidoptera: Lycaenidae). Org. Divers. Evol. 2020, 20, 63–76. [Google Scholar] [CrossRef]
- Durand, K.; Yainna, S.; Nam, K. Incipient speciation between host-plant strains in the fall armyworm. BMC Ecol. Evol. 2022, 22, 52. [Google Scholar] [CrossRef]
- Zinetti, F.; Dapporto, L.; Vovlas, A.; Chelazzi, G.; Bonelli, S.; Balletto, E.; Ciofi, C. When the Rule Becomes the Exception. No Evidence of Gene Flow between Two Zerynthia Cryptic Butterflies Suggests the Emergence of a New Model Group. PLoS ONE 2013, 8, e65746. [Google Scholar] [CrossRef]
- Burban, C.; Rocha, S.; Leblois, R.; Rossi, J.P.; Sauné, L.; Branco, M.; Kerdelhué, C. From sympatry to parapatry: A rapid change in the spatial context of incipient allochronic speciation. Evol. Ecol. 2020, 34, 101–121. [Google Scholar] [CrossRef]
- Nishida, R.; Fukami, H. Ecological adaptation of an Aristolochiaceae-feeding swallowtail butterfly, Atrophaneura alcinous, to aristolochic acids. J. Chem. Ecol. 1989, 15, 2549–2563. [Google Scholar] [CrossRef]
- Descimon, H.; Renon, C. Mélanisme et facteurs climatiques: I—Étude biométrique de la variation de Melanargia galathea (Linné) (Lepidoptera Satyridae) en France. Arch. Zool. Exp. Gén. 1975, 116, 255–292. [Google Scholar]
- Luebke, H.J.; Scriber, J.M.; Yandell, B.S. Use of Multivariate Discriminant Analysis of male wing morphometrics to delineate a hybrid zone for Papilio glaucus glacus and P. g. canadensis in Wisconsin. Am. Midl. Nat. 1988, 119, 366–379. [Google Scholar] [CrossRef]
- Winding, J.J.; Rintamäki, P.T.; Cassel, A.; Nylin, S. How useful is fluctuating asymmetry in conservation biology: Asymmetry in rare and abundant Coenonympha butterflies. J. Insect Conserv. 2001, 4, 253–261. [Google Scholar] [CrossRef]
- Cespedes, A.; Penz, C.M.; Devries, P.J. Cruising the rain forest floor: Butterfly wing shape evolution and gliding in ground effect. J. Anim. Ecol. 2014, 84, 808–816. [Google Scholar] [CrossRef]
- Habel, J.C.; Vila, R.; Vodă, R.; Husemann, M.; Schmitt, T.; Dapporto, L. Differentiation in the marbled white butterfly species complex driven by multiple evolutionary forces. J. Biogeogr. 2014, 44, 433–445. [Google Scholar] [CrossRef]
- Lemic, D.; Viric Gasparic, H.; Majcenic, P.; Pajač Živković, I.; Bjeliš, M.; Suazo, M.J.; Correa, M.; Hernández, J.; Benítez, H.A. Wing Shape Variation between Terrestrial and Coastal Populations of the Invasive Box Tree Moth, Cydalima perspectalis, in Croatia. Animals 2023, 13, 3044. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.A.; Sitar, C.; Rákosy, L. Non-invasive methods for morphometric analyses of lepidopteran wings. Entomol. Rom. 2020, 24, 25–28. [Google Scholar] [CrossRef]
- Forsman, A.; Polic, D.; Sunde, J.; Betzholtz, P.E.; Franzén, M. Variable colour patterns indicate multidimensional, intraspecific trait variation and ecological generalization in moths. Ecography 2020, 43, 823–833. [Google Scholar] [CrossRef]
- Ministerul Mediului, Apelor și Pădurilor. MMAP Supune Dezbaterii Publice Planul de Management al Sitului de Importanță Comunitară ROȘCI0238 Suatu-Cojocna-Crairat. Available online: https://mmediu.ro/articol/mmap-supune-dezbaterii-publice-planul-de-management-al-sitului-de-importanta-comunitara-rosci0238-suatu-cojocna-crairat/1147 (accessed on 17 January 2024).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Min. | Median | Max. | Average | |
|---|---|---|---|---|
| Apahida | 5 | 10 | 17 | 10.23 |
| Viisoara | 2.5 | 3.75 | 13 | 4.28 |
| Cefa | 6 | 9 | 14 | 9.33 |
| Location | Average Wing Length | Average Wing Width |
|---|---|---|
| Apahida | 24.03958333 | 12.66208333 |
| Viișoara | 20.70883721 | 10.75348837 |
| Cefa | 24.90826923 | 13.27403846 |
| PC | % Variance | Cumulative % |
|---|---|---|
| 1 | 44.355 | 44.355 |
| 2 | 19.81 | 64.165 |
| AP | CR | |
|---|---|---|
| CR | 0.0266 | |
| VI | 0.0158 | 0.0004 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rákosy, L.; Martin, M.A.; Sitar, G.M.; Crișan, A.; Sitar, C. Exploring Morphological Population Variability: Host Plant and Habitat Dependency in the Protected Moth Gortyna borelii (Lepidoptera, Noctuidae). Diversity 2024, 16, 227. https://doi.org/10.3390/d16040227
Rákosy L, Martin MA, Sitar GM, Crișan A, Sitar C. Exploring Morphological Population Variability: Host Plant and Habitat Dependency in the Protected Moth Gortyna borelii (Lepidoptera, Noctuidae). Diversity. 2024; 16(4):227. https://doi.org/10.3390/d16040227
Chicago/Turabian StyleRákosy, László, Mihai Alexandru Martin, Geanina Magdalena Sitar, Andrei Crișan, and Cristian Sitar. 2024. "Exploring Morphological Population Variability: Host Plant and Habitat Dependency in the Protected Moth Gortyna borelii (Lepidoptera, Noctuidae)" Diversity 16, no. 4: 227. https://doi.org/10.3390/d16040227