
Soil–Plant–Pollinator Relationships in Urban Grass and Meadow Habitats: Competing Benefits and Demands of Tall Flowering Plants on Soil and Pollinator Diversity


Abstract
1. Introduction
2. Materials and Methods
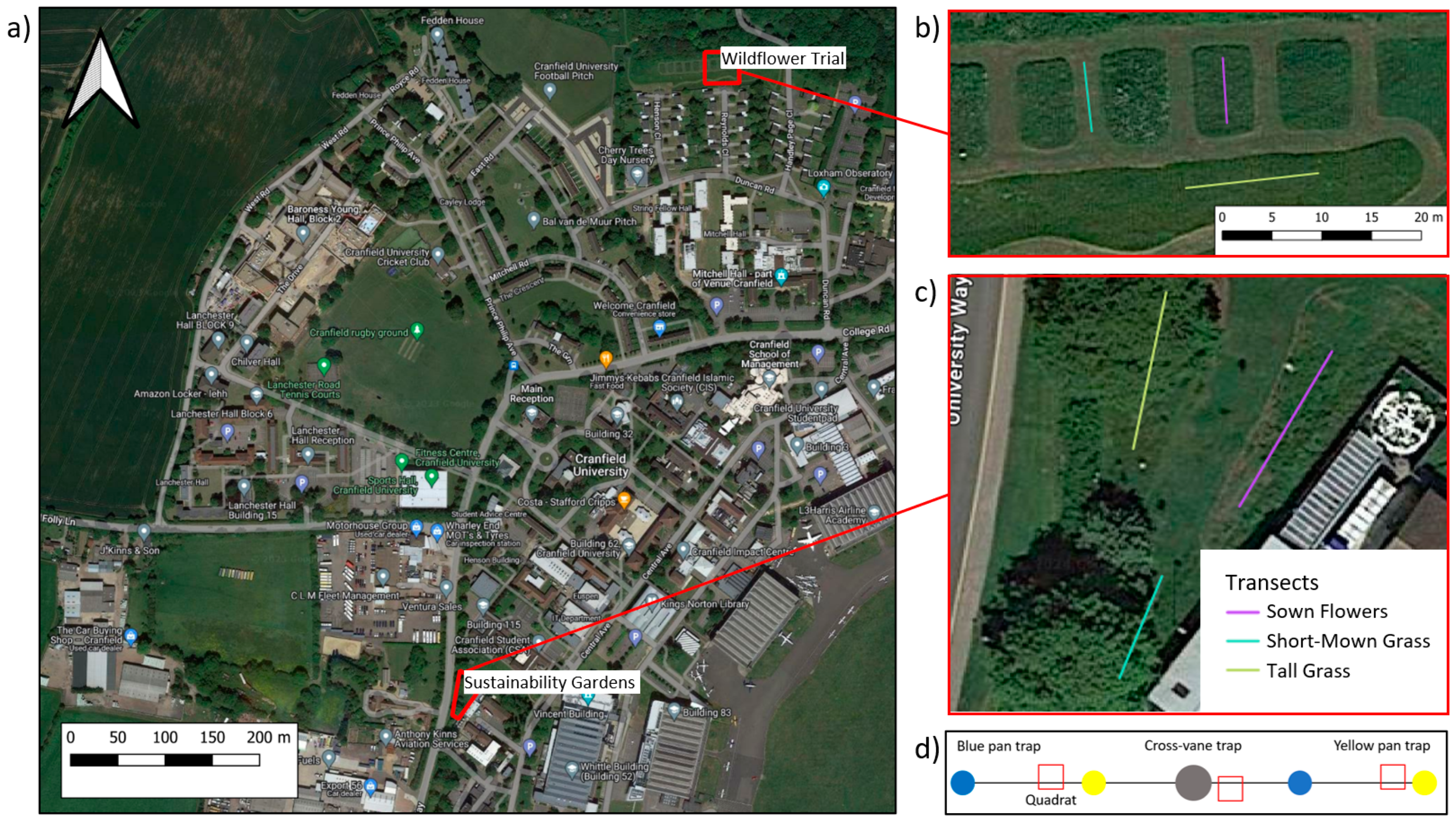
2.1. Study Area and Field Design
2.2. Aboveground Invertebrates
2.3. Soil and Plant Analyses
2.4. Statistical Analyses
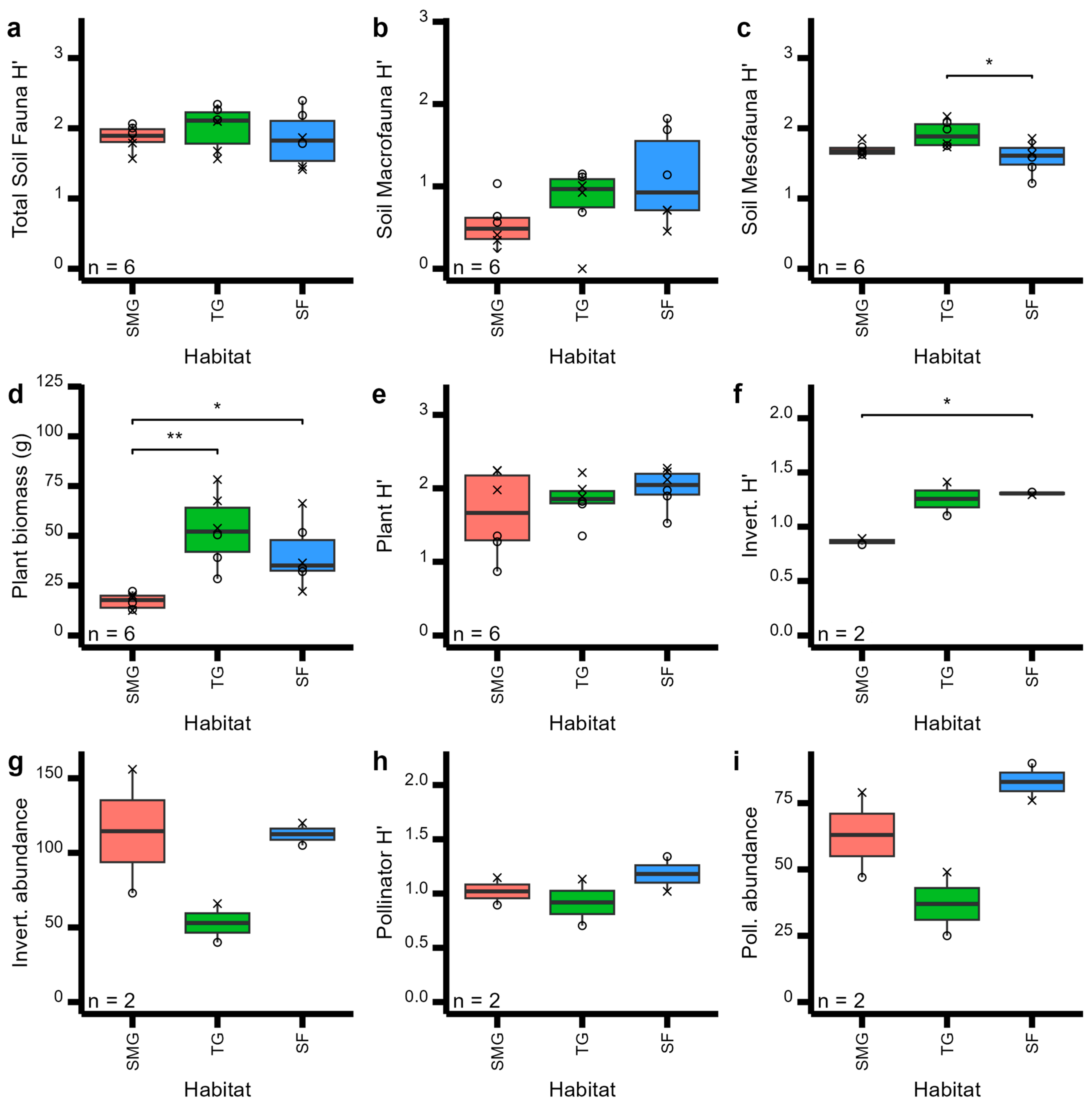
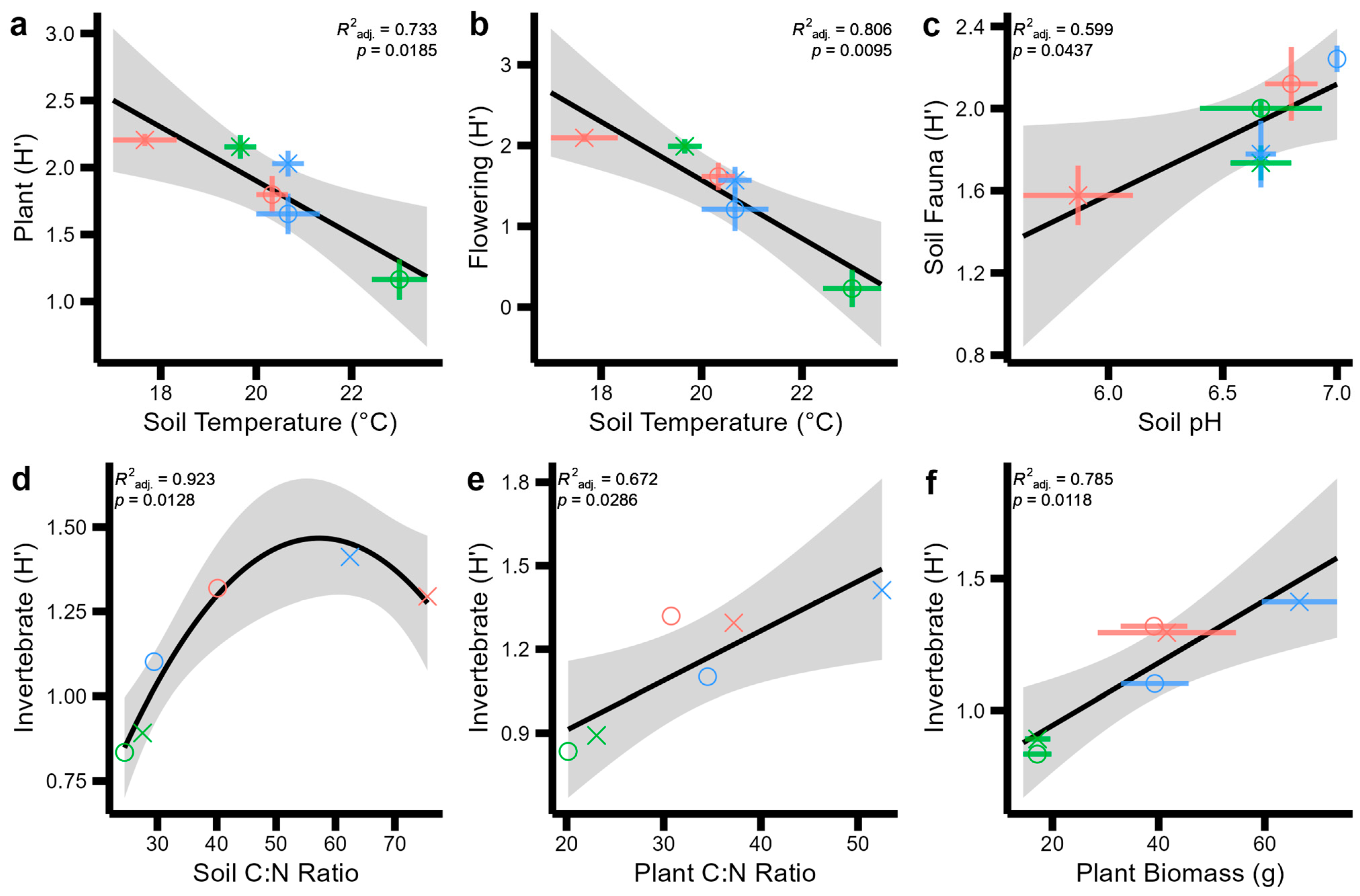
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPBES. Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; Brondizio, E.S., Settele, J., Díaz, S., Ngo, H.T., Eds.; IPBES Secretariat: Bonn, Germany, 2019; 1148p. [Google Scholar] [CrossRef]
- Mace, G.M.; Norris, K.; Fitter, A.H. Biodiversity and Ecosystem Services: A Multilayered Relationship. Trends Ecol. Evol. 2012, 27, 19–26. [Google Scholar] [CrossRef]
- Mori, A. Advancing Nature—Based Approaches to address the Biodiversity and Climate Emergency. Ecol. Lett. Viewp. 2020, 23, 1729–1732. [Google Scholar] [CrossRef]
- United Nations Environment Programme (UNEP) UN Decade on Restoration. Available online: http://www.decadeonrestoration.org/node (accessed on 18 March 2024).
- Newbold, T.; Hudson, L.N.; Hill, S.L.L.; Contu, S.; Lysenko, I.; Senior, R.A.; Börger, L.; Bennett, D.J.; Choimes, A.; Collen, B.; et al. Global Effects of Land Use on Local Terrestrial Biodiversity. Nature 2015, 520, 45–50. [Google Scholar] [CrossRef]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global Consequences of Land Use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, J.; Buchholz, S.; Theodorou, P. The Degree of Urbanisation Reduces Wild Bee and Butterfly Diversity and Alters the Patterns of Flower-Visitation in Urban Dry Grasslands. Sci. Rep. 2023, 13, 2702. [Google Scholar] [CrossRef] [PubMed]
- Garibaldi, L.A.; Gomez Carella, D.S.; Nabaes Jodar, D.N.; Smith, M.R.; Timberlake, T.P.; Myers, S.S. Exploring Connections between Pollinator Health and Human Health. Philos. Trans. R. Soc. B Biol. Sci. 2022, 377, 20210158. [Google Scholar] [CrossRef]
- Bates, A.J.; Sadler, J.P.; Fairbrass, A.J.; Falk, S.J.; Hale, J.D.; Matthews, T.J. Changing Bee and Hoverfly Pollinator Assemblages along an Urban-Rural Gradient. PLoS ONE 2011, 6, e23459. [Google Scholar] [CrossRef]
- Fortel, L.; Henry, M.; Guilbaud, L.; Guirao, A.L.; Kuhlmann, M.; Mouret, H.; Rollin, O.; Vaissière, B.E. Decreasing Abundance, Increasing Diversity and Changing Structure of the Wild Bee Community (Hymenoptera: Anthophila) along an Urbanization Gradient. PLoS ONE 2014, 9, e104679. [Google Scholar] [CrossRef] [PubMed]
- Graffigna, S.; González-Vaquero, R.A.; Torretta, J.P.; Marrero, H.J. Importance of Urban Green Areas’ Connectivity for the Conservation of Pollinators. Urban Ecosyst. 2023, 27, 417–426. [Google Scholar] [CrossRef]
- Theodorou, P.; Albig, K.; Radzevičiūtė, R.; Settele, J.; Schweiger, O.; Murray, T.E.; Paxton, R.J. The Structure of Flower Visitor Networks in Relation to Pollination across an Agricultural to Urban Gradient. Funct. Ecol. 2017, 31, 838–847. [Google Scholar] [CrossRef]
- Theodorou, P.; Radzevičiūtė, R.; Lentendu, G.; Kahnt, B.; Husemann, M.; Bleidorn, C.; Settele, J.; Schweiger, O.; Grosse, I.; Wubet, T.; et al. Urban Areas as Hotspots for Bees and Pollination but not a Panacea for all Insects. Nat. Commun. 2020, 11, 576. [Google Scholar] [CrossRef] [PubMed]
- Dobbs, C.; Nitschke, C.R.; Kendal, D. Global Drivers and Tradeoffs of Three Urban Vegetation Ecosystem Services. PLoS ONE 2014, 9, e113000. [Google Scholar] [CrossRef] [PubMed]
- Drillet, Z.; Fung, T.K.; Leong, R.A.T.; Sachidhanandam, U.; Edwards, P.; Richards, D. Urban Vegetation Types Are not Perceived Equally in Providing Ecosystem Services and Disservices. Sustainability 2020, 12, 2076. [Google Scholar] [CrossRef]
- Faucon, M.-P.; Houben, D.; Lambers, H. Plant Functional Traits: Soil and Ecosystem Services. Trends Plant Sci. 2017, 22, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Lavorel, S. Plant Functional Effects on Ecosystem Services. J. Ecol. 2013, 101, 4–8. [Google Scholar] [CrossRef]
- Llodrà-Llabrés, J.; Cariñanos, P. Enhancing Pollination Ecosystem Service in Urban Green Areas: An Opportunity for the Conservation of Pollinators. Urban For. Urban Green. 2022, 74, 127621. [Google Scholar] [CrossRef]
- Griffiths-Lee, J.; Nicholls, E.; Goulson, D. Sown Mini-Meadows Increase Pollinator Diversity in Gardens. J. Insect Conserv. 2022, 26, 299–314. [Google Scholar] [CrossRef]
- Cohen, H.; Philpott, S.M.; Liere, H.; Lin, B.B.; Jha, S. The Relationship between Pollinator Community and Pollination Services is Mediated by Floral Abundance in Urban Landscapes. Urban Ecosyst. 2021, 24, 275–290. [Google Scholar] [CrossRef]
- Norton, B.A.; Bending, G.D.; Clark, R.; Corstanje, R.; Dunnett, N.; Evans, K.L.; Grafius, D.R.; Gravestock, E.; Grice, S.M.; Harris, J.A.; et al. Urban Meadows as an Alternative to Short Mown Grassland: Effects of Composition and Height on Biodiversity. Ecol. Appl. 2019, 29, e01946. [Google Scholar] [CrossRef]
- Lerman, S.B.; Contosta, A.R.; Milam, J.; Bang, C. To Mow or to Mow Less: Lawn Mowing Frequency Affects Bee Abundance and Diversity in Suburban Yards. Biol. Conserv. 2018, 221, 160–174. [Google Scholar] [CrossRef]
- Roguz, K.; Chiliński, M.; Roguz, A.; Zych, M. Pollination of Urban Meadows—Plant Reproductive Success and Urban-Related Factors Influencing Frequency of Pollinators Visits. Urban For. Urban Green. 2023, 84, 127944. [Google Scholar] [CrossRef]
- Hussain, R.I.; Walcher, R.; Vogel, N.; Krautzer, B.; Rasran, L.; Frank, T. Effectiveness of Flowers Strips on Insect’s Restoration in Intensive Grassland. Agric. Ecosyst. Environ. 2023, 348, 108436. [Google Scholar] [CrossRef]
- Sehrt, M.; Bossdorf, O.; Freitag, M.; Bucharova, A. Less is More! Rapid Increase in Plant Species Richness after Reduced Mowing in Urban Grasslands. Basic Appl. Ecol. 2020, 42, 47–53. [Google Scholar] [CrossRef]
- Watson, C.J.; Carignan-Guillemette, L.; Turcotte, C.; Maire, V.; Proulx, R. Ecological and Economic Benefits of Low-Intensity Urban Lawn Management. J. Appl. Ecol. 2020, 57, 436–446. [Google Scholar] [CrossRef]
- Piqueray, J.; Gilliaux, V.; Decruyenaere, V.; Cornelis, J.-T.; Uyttenbroeck, R.; Mahy, G. Management of Grassland-like Wildflower Strips Sown on Nutrient-Rich Arable Soils: The Role of Grass Density and Mowing Regime. Environ. Manag. 2019, 63, 647–657. [Google Scholar] [CrossRef]
- Barber, N.A.; Soper Gorden, N.L. How Do Belowground Organisms Influence Plant–Pollinator Interactions? J. Plant Ecol. 2015, 8, 1–11. [Google Scholar] [CrossRef]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setälä, H.; van der Putten, W.H.; Wall, D.H. Ecological Linkages Between Aboveground and Belowground Biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef] [PubMed]
- Blaydes, H.; Potts, S.G.; Whyatt, J.D.; Armstrong, A. Opportunities to Enhance Pollinator Biodiversity in Solar Parks. Renew. Sustain. Energy Rev. 2021, 145, 111065. [Google Scholar] [CrossRef]
- Hyvönen, T.; Huusela, E.; Kuussaari, M.; Niemi, M.; Uusitalo, R.; Nuutinen, V. Aboveground and Belowground Biodiversity Responses to Seed Mixtures and Mowing in a Long-Term Set-Aside Experiment. Agric. Ecosyst. Environ. 2021, 322, 107656. [Google Scholar] [CrossRef]
- McCravy, K.W. A Review of Sampling and Monitoring Methods for Beneficial Arthropods in Agroecosystems. Insects 2018, 9, 170. [Google Scholar] [CrossRef]
- Popic, T.J.; Davila, Y.C.; Wardle, G.M. Evaluation of Common Methods for Sampling Invertebrate Pollinator Assemblages: Net Sampling Out-Perform Pan Traps. PLoS ONE 2013, 8, e66665. [Google Scholar] [CrossRef] [PubMed]
- Broussard, M.; Rao, S.; Stephen, W.P.; White, L. Native Bees, Honeybees, and Pollination in Oregon Cranberries. HortScience 2011, 46, 885–888. [Google Scholar] [CrossRef]
- Hall, M.A.; Reboud, E.L. High sampling effectiveness for Non-Bee Pollinators Using Vane Traps in Both Open and Wooded Habitats. bioRxiv 2019, 6, 556498. [Google Scholar] [CrossRef]
- Prayogo, C.; Sholehuddin, N.; Putra, E.; Rachmawati, R. Soil Macrofauna Diversity and Structure under Different Management of Pine-Coffee Agroforestry System. J. Degrad. Min. Lands Manag. 2019, 6, 1727–1736. [Google Scholar] [CrossRef]
- Smith, J.; Potts, S.; Eggleton, P. Evaluating the Efficiency of Sampling Methods in Assessing Soil Macrofauna Communities in Arable Systems. Eur. J. Soil Biol. 2008, 44, 271–276. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Straube, D.; Scheu, S. Efficiency of Two Widespread Non-Destructive Extraction Methods under Dry Soil Conditions for Different Ecological Earthworm Groups. Eur. J. Soil Biol. 2008, 44, 141–145. [Google Scholar] [CrossRef]
- Iannone, B.V.; Umek, L.G.; Wise, D.H.; Heneghan, L. A Simple, Safe, and Effective Sampling Technique for Investigating Earthworm Communities in Woodland Soils: Implications for Citizen Science. Naar 2012, 32, 283–292. [Google Scholar] [CrossRef]
- Roper, W.R.; Robarge, W.P.; Osmond, D.L.; Heitman, J.L. Comparing Four Methods of Measuring Soil Organic Matter in North Carolina Soils. Soil Sci. Soc. Am. J. 2019, 83, 466–474. [Google Scholar] [CrossRef]
- Heuck, C.; Weig, A.; Spohn, M. Soil Microbial Biomass C:N:P Stoichiometry and Microbial Use of Organic Phosphorus. Soil Biol. Biochem. 2015, 85, 119–129. [Google Scholar] [CrossRef]
- Chandra, K.; Rizvi, A.; Acharya, S.; Raghunathan, C. Soil Fauna of India Manual; ResearchGate: Berlin, Germany, 2019; ISBN 978-81-8171-523-4. [Google Scholar]
- Edwards, C.A. The Assessment of Populations of Soil-Inhabiting Invertebrates. Agric. Ecosyst. Environ. 1991, 34, 145–176. [Google Scholar] [CrossRef]
- Paul, B.K.; Vanlauwe, B.; Hoogmoed, M.; Hurisso, T.T.; Ndabamenye, T.; Terano, Y.; Six, J.; Ayuke, F.O.; Pulleman, M.M. Exclusion of Soil Macrofauna Did not Affect Soil Quality but Increased Crop Yields in a Sub-Humid Tropical Maize-Based System. Agric. Ecosyst. Environ. 2015, 208, 75–85. [Google Scholar] [CrossRef]
- Crotty, F.; Shepherd, M. A Key to the Soil Mites of Britain and Ireland. Available online: https://www.fscbiodiversity.uk/idsignpost/key-soil-mites-britain-and-ireland (accessed on 18 March 2024).
- Tan, H.; Barret, M.; Mooij, M.J.; Rice, O.; Morrissey, J.P.; Dobson, A.; Griffiths, B.; O’Gara, F. Long-Term Phosphorus Fertilisation Increased the Diversity of the Total Bacterial Community and the phoD Phosphorus Mineraliser Group in Pasture Soils. Biol Fertil Soils 2013, 49, 661–672. [Google Scholar] [CrossRef]
- R Core Team R: The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 3 June 2024).
- Wickham, H.; François, R.; Henry, L.; Müller, K.; Vaughan, D. A Grammar of Data Manipulation. Available online: https://dplyr.tidyverse.org/ (accessed on 3 June 2024).
- An Introduction to Reshape2—Reshaping Data Easily with the Reshape2 R Package.—Seananderson.ca. Available online: https://seananderson.ca/2013/10/19/reshape/ (accessed on 3 June 2024).
- Create Elegant Data Visualisations Using the Grammar of Graphics. Available online: https://ggplot2.tidyverse.org/ (accessed on 3 June 2024).
- Ahlmann-Eltze, P.C. Indrajeet Ggsignif Package. Available online: https://cran.r-project.org/web/packages/ggsignif/vignettes/intro.html (accessed on 3 June 2024).
- Tresch, S.; Frey, D.; Bayon, R.-C.L.; Mäder, P.; Stehle, B.; Fliessbach, A.; Moretti, M. Direct and Indirect Effects of Urban Gardening on Aboveground and Belowground Diversity Influencing Soil Multifunctionality. Sci. Rep. 2019, 9, 9769. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Peng, S.; Chen, X.; Chen, H.Y.H. Plant Diversity Increases the Abundance and Diversity of Soil Fauna: A Meta-Analysis. Geoderma 2022, 411, 115694. [Google Scholar] [CrossRef]
- Bednar, Z.; Vaupel, A.; Blümel, S.; Herwig, N.; Hommel, B.; Haberlah-Korr, V.; Beule, L. Earthworm and Soil Microbial Communities in Flower Strip Mixtures. Plant Soil 2023, 492, 209–227. [Google Scholar] [CrossRef]
- Kozlov, M.V.; Zverev, V. Insecticide Application Did Not Reveal Any Impact of Herbivory on Plant Roots in Boreal Forests. Appl. Soil Ecol. 2022, 178, 104554. [Google Scholar] [CrossRef]
- Graves, R.A.; Pearson, S.M.; Turner, M.G. Species Richness Alone Does not Predict Cultural Ecosystem Service Value. Proc. Natl. Acad. Sci. USA 2017, 114, 3774–3779. [Google Scholar] [CrossRef]
- Southon, G.E.; Jorgensen, A.; Dunnett, N.; Hoyle, H.; Evans, K.L. Biodiverse Perennial Meadows Have Aesthetic Value and Increase Residents’ Perceptions of Site Quality in Urban Green-Space. Landsc. Urban Plan. 2017, 158, 105–118. [Google Scholar] [CrossRef]
- Humbert, J.-Y.; Pellet, J.; Buri, P.; Arlettaz, R. Does Delaying the First Mowing Date Benefit Biodiversity in Meadowland? Environ. Evid. 2012, 1, 9. [Google Scholar] [CrossRef]
- Raubenheimer, D.; Simpson, S.J.; Mayntz, D. Nutrition, Ecology and Nutritional Ecology: Toward an Integrated Framework. Funct. Ecol. 2009, 23, 4–16. [Google Scholar] [CrossRef]
- Gaston, K.J.; Ávila-Jiménez, M.L.; Edmondson, J.L. Review: Managing Urban Ecosystems for Goods and Services. J. Appl. Ecol. 2013, 50, 830–840. [Google Scholar] [CrossRef]
- Oelmann, Y.; Wilcke, W.; Temperton, V.M.; Buchmann, N.; Roscher, C.; Schumacher, J.; Schulze, E.-D.; Weisser, W.W. Soil and Plant Nitrogen Pools as Related to Plant Diversity in an Experimental Grassland. Soil Sci. Soc. Am. J. 2007, 71, 720–729. [Google Scholar] [CrossRef]
- Rojas-Botero, S.; Dietzel, S.; Kollmann, J.; Teixeira, L.H. Towards a Functional Understanding of Rehabilitated Urban Road Verge Grasslands: Effects of Planting Year, Site Conditions, and Landscape Factors. Flora 2023, 309, 152417. [Google Scholar] [CrossRef]
- Zaninotto, V.; Fauviau, A.; Dajoz, I. Diversity of Greenspace Design and Management Impacts Pollinator Communities in a Densely Urbanized Landscape: The City of PARIS, France. Urban Ecosyst. 2023, 26, 503–515. [Google Scholar] [CrossRef]
- Vega, K.A.; Küffer, C. Promoting Wildflower Biodiversity in Dense and Green Cities: The Important Role of Small Vegetation Patches. Urban For. Urban Green. 2021, 62, 127165. [Google Scholar] [CrossRef]
- Wenzel, A.; Grass, I.; Belavadi, V.V.; Tscharntke, T. How Urbanization is Driving Pollinator Diversity and Pollination—A Systematic Review. Biol. Conserv. 2020, 241, 108321. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Short-Mown Grass | Tall Grass | Sown Flowers |
|---|---|---|---|
| Vegetation Height (cm) Site: ns; HT: ***. | 10.75 ± 4.66 | 75.50 ± 45.46 | 84.50 ± 13.69 |
| Plant C:N ratio Site: ns; HT: **** | 21.62 ± 1.60 | 43.49 ± 9.83 | 33.98 ± 3.53 |
| Soil C:N ratio Site: **; HT: ** | 25.99 ± 1.66 | 45.94 ± 18.07 | 57.81 ± 19.36 |
| Soil microbial C:N ratio (0–5 cm) Site: ns; HT: ns | 13.11 ± 0.35 | 13.51 ± 1.79 | 13.45 ± 0.97 |
| Soil microbial C:N ratio (0–20 cm) Site: ns; HT: ns | 12.19 ± 0.79 | 12.19 ± 1.30 | 11.92 ± 0.98 |
| Soil temperature (°C) Site: *; HT: ns | 21.34 ± 1.97 | 20.67 ± 0.82 | 19.00 ± 1.67 |
| Soil pH Site: *; HT: ns | 6.67 ± 0.33 | 6.84 ± 0.19 | 6.34 ± 0.59 |
| Soil moisture (%) Site: ns; HT: ns | 0.36 ± 0.05 | 0.30 ± 0.03 | 0.33 ± 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rust, W.; Sotkewicz, M.; Li, Z.; Mercer, T.; Johnston, A.S. Soil–Plant–Pollinator Relationships in Urban Grass and Meadow Habitats: Competing Benefits and Demands of Tall Flowering Plants on Soil and Pollinator Diversity. Diversity 2024, 16, 354. https://doi.org/10.3390/d16060354
Rust W, Sotkewicz M, Li Z, Mercer T, Johnston AS. Soil–Plant–Pollinator Relationships in Urban Grass and Meadow Habitats: Competing Benefits and Demands of Tall Flowering Plants on Soil and Pollinator Diversity. Diversity. 2024; 16(6):354. https://doi.org/10.3390/d16060354
Chicago/Turabian StyleRust, Will, Madison Sotkewicz, Zhaoxing Li, Theresa Mercer, and Alice S. Johnston. 2024. "Soil–Plant–Pollinator Relationships in Urban Grass and Meadow Habitats: Competing Benefits and Demands of Tall Flowering Plants on Soil and Pollinator Diversity" Diversity 16, no. 6: 354. https://doi.org/10.3390/d16060354
APA StyleRust, W., Sotkewicz, M., Li, Z., Mercer, T., & Johnston, A. S. (2024). Soil–Plant–Pollinator Relationships in Urban Grass and Meadow Habitats: Competing Benefits and Demands of Tall Flowering Plants on Soil and Pollinator Diversity. Diversity, 16(6), 354. https://doi.org/10.3390/d16060354