Abstract
Limosilactobacillus pontis (L. pontis) is a species of lactic acid bacteria (LAB) found in various products, including fermented milk, sourdough, and broiler chickens’ gastrointestinal tracts. However, the evolutionary strategies and genomic features of the species remain unknown, which limits its application. In this study, whole-genome sequencing was carried out to investigate the genomic characteristics and evolutionary strategies of L. pontis, combined with a comparative genomic approach, providing the necessary theoretical basis for its application. The genomic sequences in this study included three published genomic sequences and two strains isolated from fermented milk in our lab in Inner Mongolia, China. The mean genome size of the five L. pontis strains was 1.70 Mb, and the mean DNA G + C content was 53.06%. L. pontis was a species with higher G + C content in LAB. The phylogenetic evolutionary tree for the core genes showed that the strains from the same sources were aggregated into a cluster. There were some differences in the genomic information and phylogenetic relationships amongst L. pontis from different sources. An analysis of the annotation results identified differences in the functional genes, carbohydrate-active enzymes, and bacteriocins amongst different isolated strains, which were related to the environment. L. pontis was rich in glycosyltransferases, especially in strains isolated from fermented milk, indicating that they had higher sugar synthesis abilities. It is worth mentioning that only the L. pontis strains isolated in our laboratory identified the bacteriocin operon, which may increase its application potential. The genomic characteristics and evolutionary process of L. pontis were analyzed by comparative genomics, and this study explored the differences in the functional genes amongst the strains, aiming to provide new insights into the genetic characteristics and niche adaptations of L. pontis.
1. Introduction
Limosilactobacillus pontis (L. pontis) used to belong to Lactobacillus and is commonly found in fermenting animal and plant materials, such as sourdough, fermented milk, broiler chickens’ gastrointestinal tracts, and vinegar [1]. It should be mentioned that because of the extreme phenotypic, ecological, and genotypic diversity among the species within the genus Lactobacillus, it has been reclassified into 25 genera, including the amended genera Lactobacillus and Limosilactobacillus and 23 others [2]. Limosilactobacillus includes L. pontis, Limosilactobacillus reuteri, and Limosilactobacillus fermentum, among others. Being generally recognized as safe [3], strains of this species have been widely applied in various foods’ fermentation [4]. Kiran Shazadi [5] found that L. reuteri (MT180537) had a significant bactericidal effect on Enterococcus faecalis (MW051601) in the pharmaceutical industry; the antimicrobial compounds or bacteriocin-like substances (BLIS) produced by these strains could be used in the preparation of ointments for intravaginal application against aerobic vaginitis (AV). Regarding L. reuteri, a member of the lactic acid bacteria (LAB) [6], a broad-spectrum antimicrobial substance is produced by this species during the fermentation of glycerol, called reuterin [7], and the secretion conditions include an appropriate temperature, weakly acidic conditions, and an anaerobic environment. It has been verified by experiments that reuterin is a mixture of monomers, hydrated monomers, and cyclic dimers of L-hydroxypropanal [8]. Reuterin has a large antibacterial range and can inhibit the growth of some Gram-positive and Gram-negative bacteria, fungi, and molds [9]. As it is in the same genus as L. reuteri, L. pontis may have potential bacteriocin-producing characteristics as well. Although L. pontis has been recorded from a wide range of sources, it has been the subject of few studies and is worthy of further exploration.
Comparative genomic studies of strains within the same species provide insights into genetic features, including gene modifications, gains, or losses, that facilitate the evolution and adaptation of strains to specific environmental niches [10]. In recent years, along with the development of rapid throughput DNA sequencing technologies, genome studies have become more feasible and affordable, and genomic data have become publicly available for numerous organisms [11]. However, the publicly available genomic data for L. pontis are very limited. Because pure cultures of L. pontis are not easy to obtain, there is little research on it. L. pontis is phylogenetically close to Limosilactobacillus vaginalis, Limosilactobacillus oris, and L. reuteri [1]. At present, compared with L. pontis, research on L. reuteri is favored in the field of Limosilactobacillus. L. reuteri can be isolated from the intestines of pigs, humans, and rodents. An investigation of L. reuteri from different sources has defined a phylogenetic tree based on core genomes that are distributed on different evolutionary branches, with habitats as clusters. This indicates that L. reuteri with different genetic backgrounds can selectively colonize and adapt to specific habitats, and this specific ability is significantly associated with the acid and bile salt tolerance and cell adhesion abilities of L. reuteri [12]. Most L. reuteri have good intestinal colonization and bacteriocin secretion abilities and have been widely used in daily life [13,14]. However, our understanding of L. pontis’ genomic characteristics is poor, which limits its application; comparative genomics could be used to reveal its genomic characteristics.
We obtained two strains of L. pontis from fermented milk through the Lactic Acid Bacteria Collection Center (LABCC) of Inner Mongolia Agricultural University: L. pontis IMAU10341 and L. pontis IMAU10345. This study analyzed the genome sequences of L. pontis IMAU10341 and L. pontis IMAU10345 using Illumina Novaseq 6000 genome sequencing. We downloaded the type strain L. pontis DSM8475T and three strains published by the NCBI (http://www.ncbi.nlm.nih.gov, accessed on 7 November 2023): L. pontis LP475, L. pontis UMB0683, and L. pontis MAG.251. We used a comparative genomics approach to define the pangenome, core genome, and unique genes of isolates from sourdough, fermented milk, and the broiler chicken gastrointestinal tract; assess the genetic diversity and gain insights into the distinguishing features; compare the numbers of functional genes and carbohydrate-active enzymes (CAZy); and explore the expression of bacteriocin genes. The aim of the current work was to carry out comparative genomics analyses of L. pontis, with more strains isolated from different niches, to explore its genetic diversity and the potential for host adaptation in this species.
2. Materials and Methods
2.1. Strains
The two strains evaluated in this study were L. pontis IMU10341 and L. pontis IMU10345 from the LABCC of Inner Mongolia Agricultural University; these strains had been isolated separately from two types of fermented milk collected in Dongwuzhumuqin Banner, Xilingol League, Inner Mongolia, China (N: 45°30′34.56″ E: 116°58′19.74″) [15]. The genomes of the remaining four L. pontis and 11 closely related strains were downloaded from the NCBI Refseq (National Coalition Building Institute, https://www.ncbi.nlm.nih.gov/refseq/, accessed on 7 November 2023) database. The strain information is listed in Table 1.

Table 1.
Genomic information for six strains of L. pontis and 11 closely related strains.
2.2. Main Instruments and Reagents
Genomic DNA extraction kits (Beijing Tiangen Biochemical Technology Co., Ltd., Beijing, China), MRS medium (Qingdao Haibo Biotechnology Co., Ltd., Qingdao, China), an electric heating constant-temperature incubator (Beijing Yiheng Technology Co., Ltd., Beijing, China), and an ultra-micro UV spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) [16].
2.3. Strain Culture and Genomic DNA Extraction
For each strain, an appropriate amount of freeze-dried bacterial powder was inoculated into 5 mL of MRS liquid medium and incubated anaerobically at 37 °C in the incubator for 24 h. Then, a 2% volume of inoculum was added to 15 mL of MRS liquid medium. The culture was incubated anaerobically for 24 h at 37 °C and the resulting bacteria collected by centrifugation [16]. A genomic DNA extraction kit was used to extract the genomic DNA, following the manufacturer’s instructions, and the purity and quality of the DNA samples were checked with an ultra-micro ultraviolet spectrophotometer [17].
2.4. Genome Sequencing and Assembly
Sample quality was determined after the genomic DNA was extracted. Once the concentration and purity met the sequencing requirements, the Illumina Novaseq 6000 sequencing platform was used to complete whole-genome sequencing [18]. Moreover, 150 bp was selected to construct paired-end (PE) sequencing libraries and the average coverage of high-quality data was about 500× [19]. The quality of the obtained raw data was evaluated. After low-quality reads were filtered, adapters and primers were removed, the high-quality reads’ sequences were obtained, and then the high-quality reads were spliced using the software SOAP denovo v2.04 (http://soap.genomics.org.cn/soapdenovo.html, accessed on 2 August 2021). Moreover, appropriate kmer values were selected [20]. Local inner gaps were then filled and single-base errors corrected using GapCloser v1.12 (http://soap.genomics.org.cn/soapdenovo.html, accessed on 3 August 2021) to finally complete the assembly of the genomes [21].
2.5. Comparative Genomics Analysis
2.5.1. Average Nucleotide Identity (ANI) Value Calculation
In this study, ANI values were calculated using FastANI (https://github.com/ParBLiSS/FastANI, accessed on 13 November 2023) according to Goris et al. [22,23]. TBtools (https://github.com/CJ-Chen/TBtools/releases, accessed on 13 November 2023) was used to draw an ANI heatmap [24].
2.5.2. Construction of Core Gene Set and Accessory Gene Set Analysis
After gene prediction based on the Prokka (v1.1.1) [25] software, the Roary (v3.13.0) [26] software was used to identify the core gene set and the accessory gene set, among which core genes were identified based on the principle that the amino acid similarity in the encoded protein was greater than 95%. A core gene is a gene that exists in the genomes of all strains; among them, a gene that appears in only one strain is a unique gene, and the unique genes from all of the strains represented the sum of the unique genes [27].
2.5.3. Phylogenetic Tree Construction
We used the core gene set obtained by Roary to construct a phylogenetic tree on the ITOL online website (https://itol.embl.de/, accessed on 14 November 2023) and explore the phylogenetic relationships among the L. pontis isolates [28].
2.5.4. Functional Gene Annotation
The genome sequences of all five L. pontis strains were annotated by uploading each sequence file to the Rapid Annotation using Subsystem Technology (RAST) web server (https://rast.nmpdr.org/rast.cgi, accessed on 17 November 2023) for genomic annotation. Protein coding sequences were predicted and categorized based on the SEED subsystem (most prominently FIGfams) in RAST [29].
2.5.5. CAZy Annotation
Carbohydrate-active enzymes were annotated for the genome sequences of the five L. pontis strains and compared and annotated through CAZy (http://www.cazy.org/, accessed on 22 November 2023). According to the results, the detailed carbohydrate-active enzyme family was searched on the CAZy webpage [30].
2.5.6. Bacteriocin
BAGEL4 (http://bagel4.molgenrug.nl/index.php, accessed on 27 November 2023) is an online database that facilitates the mining and visualization of ribosome-synthesized and post-translationally modified peptides and bacteriocin-producing gene clusters in the prokaryotic genome. On the BAGEL4 web server (accessed on 27 November 2023), a DNA nucleotide sequence was used as an input file [31].
2.6. Uploading Strain Genome
The assembled genomic data of the two strains (L. pontis IMAU10341 and L. pontis IMAU10345) were uploaded to the SRA database in the NCBI. The whole-genome shotgun sequences of strains IMAU10341 and IMAU10345 were deposited at DDBJ/EMBL/GenBank under the accession numbers JAIUWJ000000000 and JAIUWK000000000, respectively.
3. Results
3.1. General Genomic Characteristics
The G + C content of L. pontis UMB0683 was 48.70%, and the genome size was 1.87 Mb, which does not match the characteristics of the L. pontis genome and requires further analysis. Except for L. pontis UMB0683, the genome sizes of all strains of L. pontis ranged from 1.60 to 1.77 Mb, the average size was 1.78 Mb, and the average G + C content was 53.06%. The size of the assembled genome of L. pontis IMAU10341 was 1.77 Mb, encoding 1787 genes, and the G + C content was 52.49%; the size of the assembled genome of L. pontis MAU10345 was 1.76 Mb, encoding 1786 genes, and the G + C content was 52.49%. In addition, the number of tRNAs in each genome of L. pontis was predicted, and the average number of tRNAs was 51 (Table 1).
3.2. ANI Analysis
The ANI is the gold standard for species identification, with ANI values of 95–96% being the boundary for species demarcation and an ANI value greater than 95% theoretically being considered to indicate the same species [22].
In this study, the ANI analysis was first performed on the whole-genome sequences of all strains in Table 1 and heat maps were drawn (Figure 1). The ANI values of these genomes were determined through pairwise comparison at the 95% threshold to further identify their species [22]. All strains clustered into two clusters: L. oris DSM4864T, Limosilactobacillus antri DSM16041T, Limosilactobacillus panis DSM6035T, L. reuteri DSM20016T, and L. vaginalis DSM5837T clustered in one large branch with L. pontis, indicating that they were more closely related to L. pontis; L. pontis IMAU10341, L. pontis IMAU10345, L. pontis LP475, and L. pontis MAG.251 clustered in one small branch with the type strain L. pontis DSM8475T (Figure 1). The ANI value of L. pontis DSM8475T was greater than 97%. In contrast, L. pontis UMB0683 was on the other small branch, and the ANI value with L. pontis DSM8475T was only 84% (less than 95%). In addition, the genomic characteristics of L. pontis UMB0683, downloaded from the NCBI, were also quite different from those of the other five strains. Thus, L. pontis UMB0683 is not a strain from this species and was excluded from further analyses. Then, the other five strains were analyzed, specifically L. pontis IMAU10341, L. pontis IMAU10345, L. pontis LP475, L. pontis DSM8475T, and L. pontis MAG.251.
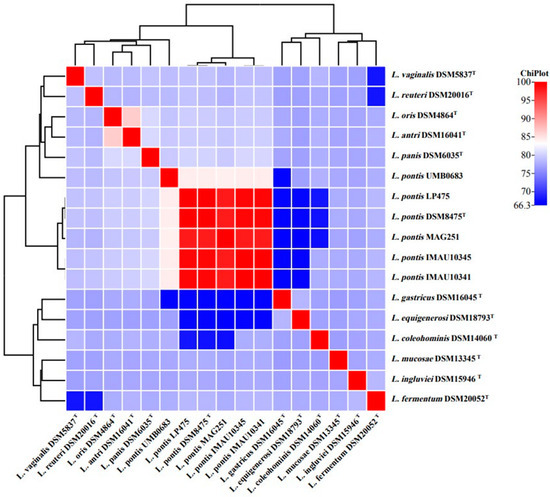
Figure 1.
ANI analysis of L. pontis and closely related strains.
3.3. Core Genome and Pangenome Analysis
In order to compare the genetic differences between different strains, we used the Prokka (v1.1.1) and Roary (v3.13.0) software, with greater than 90% amino acid identity as the standard; the numbers of core genome and pangenome genes were counted [32]. The pangenome sets of the five strains of L. pontis contained 2166 genes, of which 1281 were core genome genes. There were eight and seven unique genes derived from fermented milk strains IMAU10341 and IMAU10345, respectively, which were all pseudogenes; 37 and 52 unique genes in sourdough-derived strains DSM8475T and LP475, respectively; and 294 specific genes in the animal-derived strain MAG.251 (Figure 2).
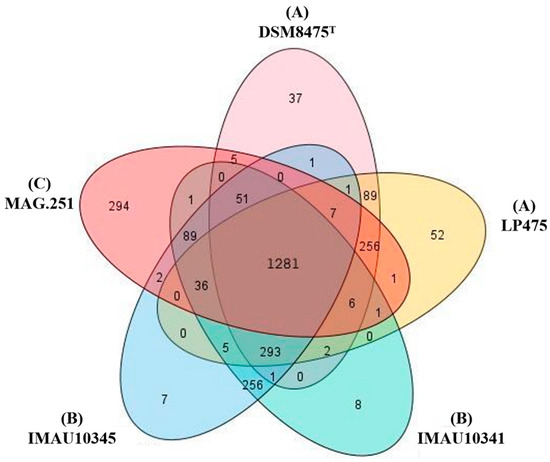
Figure 2.
Venn diagram of core genomes of five L. pontis strains divided into three classes. (A) Sourdough, (B) fermented milk, (C) broiler chicken gastrointestinal tract.
3.4. Phylogenetic Tree
A phylogenetic tree directly shows the genetic distances and evolutionary relationships between individuals in the same population. Building a phylogenetic tree is a necessary prerequisite for the study of population structure and species evolution [33]. To further understand the population structure of L. pontis, we used neighbor joining to construct a phylogenetic tree, with a bootstrap value of 1000 [34]. This was based on comparisons between the nucleic acid sequences of the L. pontis strains and closely related species as outgroups: L. oris DSM4864T, L. antri DSM16041T, L. panis DSM6035T, L. reuteri DSM20016T, and L. vaginalis DSM5837T. The five strains of L. pontis were different from each other due to the development of genetic diversity during evolution; they were divided into three main branches (Figure 3). Isolates from sourdough (L. pontis LP475 and L. pontis DSM8475T) were grouped together in one branch; strains from fermented milk (L. pontis IMAU10341 and L. pontis IMAU10345) were included in a second branch; and the branch containing the strain isolated from the broiler chicken gastrointestinal tract (L. pontis MAG.251) was closer to the root of the outgroup.
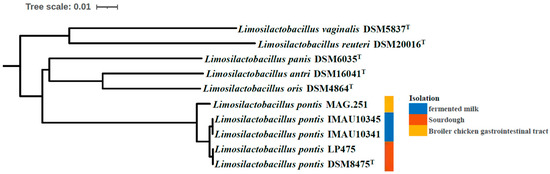
Figure 3.
Phylogenetic tree constructed based on 279 core genes of five L. pontis strains and a further five closely related strains. The colors signify the origins of the strains. Blue = fermented milk, red = sourdough, yellow = broiler chicken gastrointestinal tract.
3.5. Functional Gene Analysis
The functional genes of the five L. pontis strains were predicted and annotated by RAST. A total of 23 functional categories were annotated (Figure 4). The L. pontis strains evaluated had more genes related to the metabolism of nucleosides and nucleotides, proteins, amino acids and derivatives, and carbohydrates than any other type of gene. Strains L. pontis IMAU10341 and L. pontis IMAU10345, from fermented milk, had significantly more genes involved in the metabolism of carbohydrates, amino acids and derivatives, fatty acids, lipids, and isoprenoids than the other strains; genes involved in cell wall and capsule development, toxicity, disease, and defense were also more abundant in these two strains than the other isolates. The number of genes related to protein metabolism was highest in the sourdough isolate, L. pontis LP475. Genes related to cofactors, vitamins, prosthetic groups, and pigments were most numerous in the broiler chicken gastrointestinal tract strain, L. pontis MAG.251.
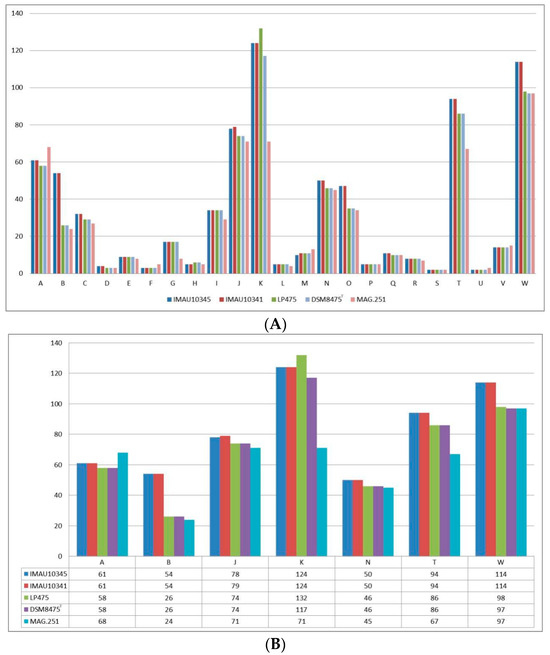
Figure 4.
Annotation of L. pontis’s functional genes based on RAST database. (A) All RAST comment results and (B) some of the comment results. A: cofactors, vitamins, prosthetic groups, and pigments; B: cell wall and capsule; C: virulence, disease, and defense; D: potassium metabolism; E: miscellaneous; F: phages, prophages, transposable elements, and plasmids; G: membrane transport; H: iron acquisition and metabolism; I: RNA metabolism; J: nucleosides and nucleotides; K: protein metabolism; L: cell division and cell cycle; M: regulation and cell signaling; N: DNA metabolism; O: fatty acids, lipids, and isoprenoids; P: dormancy and sporulation; Q: respiration; R: stress response; S: metabolism of aromatic compounds; T: amino acids and derivatives; U: sulfur metabolism; V: phosphorus metabolism; W: carbohydrates.
3.6. Carbohydrate-Active Enzyme Analysis
To further study the differences in the genome of L. pontis and explore the reasons for the diversity in L. pontis, Prokka annotation and RAST annotation were performed for each strain. It was found that the proportion of carbohydrate-utilization-related functions in each genome was significantly different, and the number of related functional genes in the strains was quite different. Therefore, the abundance of the carbohydrate-related active enzymes amongst the strains was compared. CAZy is an enzyme database resource, based on the similarity of the amino acid sequences in the protein domains, which can synthesize or decompose various complex carbohydrates and sugar complexes [35]. The main function of the GH family is to hydrolyze or rearrange glycosidic bonds, including 152 gene families; the GT family is mainly responsible for the formation of glycosidic bonds, including 105 gene families; the function of the PL family is to break glycosidic bonds (not by hydrolysis), including 28 gene families; the CE family can hydrolyze carbohydrate esterases, including 16 gene families; the AA family is an oxidoreductase, which works in synergy with carbohydrate-active enzymes and includes 14 gene families; the CBM family is attached to carbohydrates, including 83 gene families [36].
The CAZy annotation showing the distribution and abundance of CE, GH, and GT family genes in L. pontis was visualized as a heat map and clustered using the hierarchical clustering method. This showed that L. pontis was divided into three groups, representing the three different isolation sources. The predictions showed that the five strains contained 11 GH, eight GT, and one CE (Figure 5). There were differences in the composition of carbohydrate-active enzymes in the L. pontis strains isolated from different sources. L. pontis contained eight GT families, among which GT2 and GT4 were the most widely distributed, and GT2 was most abundant in strains from fermented milk. The GT32 family and GT101 family were only found in L. pontis MAG.251. L. pontis contained 11 GT families, of which GT36 and GT73 were the most widely distributed. GH53 was only found in sourdough strains. The functional genes varied in relation to the source and habitats, which may have resulted in evolution in different directions under the selection pressure of the living environment. GT2 and GT4 (glycosyltransferases, GTs), involved in the synthesis of cellulose and sucrose, were detected in L. pontis and were regarded as the largest family of carbohydrate enzymes, especially in strains isolated from fermented milk.
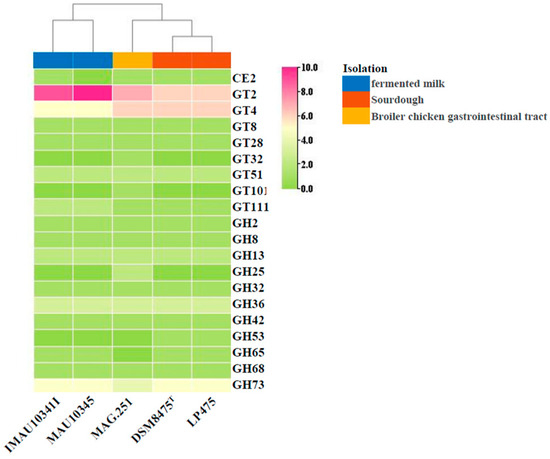
Figure 5.
Heatmap of CAZyme families of L. pontis from different strains. Blue = fermented milk, red = sourdough, yellow = broiler chicken gastrointestinal tract.
3.7. Bacteriocin
In recent years, the use of software combined with databases has enabled us to quickly identify operons that can encode bacteriocins in the bacterial genome. Most studies use BAGEL to identify potential bacteriocin operons [37]. Bacteriocins can be roughly divided into three classes; Class I bacteriocins encompass all peptides that undergo enzymatic modification during biosynthesis (heat-stable); Class II bacteriocins are unmodified bacteriocins (heat-stable); and Class III bacteriocins are large-molecular-weight and heat-labile antimicrobial proteins, usually composed of different domains [38].
We also used BAGEL to predict the bacteriocin operons in the five strains of L. pontis. The bacteriocin gene cluster prediction found that both L. pontis MAU10341 and L. pontis IMAU10345 from fermented milk contained the same core peptide, which was enterolysin A (Figure 6). Bacteriocin was not detected in the other three strains. Enterolysin A belongs to the Class III bacteriocins, is a novel antimicrobial protein, can degrade cell walls, and is mainly purified from an Enterococcus faecalis culture. Enterolysin A inhibits the growth of selected Enterococci [39]. The function of stationary-phase bacteriocin production might also be to kill bacteria in the same environment and thereby reduce the number of bacteria that compete for limiting nutrients [39]. Fermented milk is rich in nutrients and contains many types and a number of strains, so the strains need to produce bacteriocins for niche competition.
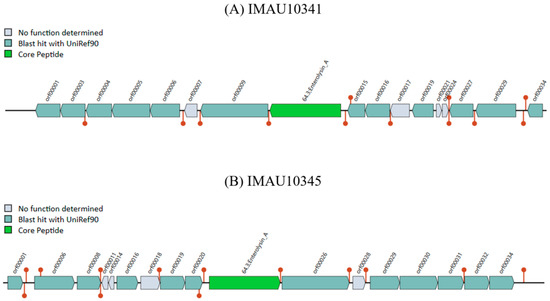
Figure 6.
Bacteriocin gene clusters of IMAU10341 and IMAU10345. (A) IMAU10341 has one core peptide; (B) IMAU10345 has one core peptide.
4. Discussion
In this study, we undertook a comparative genomics analysis of the whole-genome sequences of five strains of L. pontis from different sources. It was found that L. pontis had a smaller genome and higher G + C content than those in the same genus. The length of the genome sequences of most Lactobacillus species is between 1.80 and 2.90 Mb, and the genome size of L. pontis was in the normal range. The G + C content of most Lactobacillus genomes is below 50%, and the G + C content of Ligilactobacillus salivarius UCC118 is at least 33% [39]. The G + C content of L. pontis in Lactobacillus was higher. The high G + C content of bacterial genomes results in a faster growth rate and a more stable DNA structure [40]. Not only is the genome density relatively large, but also the ability to resist high temperatures and alkaline environments is enhanced [41]. Compared with nomadic species and free-living species, the G + C content of the host-adapted Lactobacillus species was decreased, but the G + C content of Lactobacillus pontis was higher than that of other Lactobacillus species [42]. Based on the core gene phylogenetic tree, the fermented milk, sourdough, and animal isolates were placed in different branches, and the branch containing animal isolates was closer to the common ancestor. The conclusions from the ANI analysis were consistent with those of the phylogenetic tree analysis. The ANI values of L. pontis IMAU10341 and L. pontis IMAU10345 were high and different from the ANI value of the type strain. Pairs of strains isolated from similar samples were closely related, but not identical. Strains from fermented milk, sourdough, and the broiler chicken gastrointestinal tract naturally clustered into several categories. In addition, there were some differences in the ANI values between strains from different sources and the type strain, and the ANI values of L. pontis strains from different hosts indicated that the species had sufficient genetic diversity to adapt to its different habitats.
In this study, the number of genes specific to a strain was similar when the strains were from the same source, but differed greatly when the strains were from different sources. The unique genes of L. pontis were under positive selection to adapt to different environments, and this study also demonstrated that food and intestinal habitats exerted different selective pressures related to the growth rate and metabolism [43]. The unique genes of L. pontis IMAU10341 and L. pontis IMAU10345 were pseudogenes. Some original genes of bacteria are degraded or inactivated if they are no longer needed. The formation of pseudogenes is one way in which genes decline or approach deletion from the genome [44]. Therefore, it is speculated that fermented milk, which is rich in readily available nutrition, represents a highly suitable environment for L. pontis strains and leads to the phenomenon of genome degradation in strains from fermented milk compared with strains from other sources.
Shuo Wang et al. [34] analyzed the phylogenetic relationships of L. ruminis, and a phylogenetic tree was created based on 91 orthologue genes that constituted the core genome. They found that strains from porcine, bovine, and human feces were divided into three clades; from the phylogenetic tree, it was clear that strains from the same sample type were clustered, closely related, and had the potential for niche adaptation. In our study, L. pontis MAG.251 was closest to the root, indicating that its genome was closer to its ancestors than the other strains evaluated. In different environments, the evolutionary process for species adaptation is different. The adaptation of bacterial genomes to specific environments will accumulate genetic variations related to the environmental characteristics. The genomes of the other four strains all changed due to environmental changes. Strains from different sources clustered together, indicating that the phylogenetic relationship reflected potential niche adaptations, which was consistent with the ANI results. The adaptive evolution of microorganisms to specific niches is also reflected in their functional genes, and specific niches will shape specific functional genes. In order to reduce the energy consumption of reproduction, strains living in specific niches will lose redundant functional genes related to the environment and simultaneously evolve specific genes adapted to these niches [45].
The functional gene annotation results showed that the L. pontis strains from fermented milk (L. pontis IMAU10341 and L. pontis IMAU10345) were rich in carbohydrate-related genes and genes involved in amino acids and derivatives. Genes related to the metabolism of fatty acids, lipids, and isoprenoids were significantly more abundant in fermented milk strains than in other strains, while genes for protein metabolism were more abundant in the sourdough strain L. pontis LP475 than in other strains. Based on RAST annotations, carbohydrates are the main energy source for the growth and development of organisms. Studies have shown that LAB can specifically adapt to the environment by obtaining carbohydrates related to the environment or losing non-essential carbohydrate genes. The differences among the five L. pontis strains in terms of their carbohydrate genes may be related to their different isolation sources; strains from the same isolation source were more similar to each other. Strains from fermented milk had more cell wall and capsule genes than other strains. This may be due to the rich environment and the greater number of microorganisms, which increase the thickness of the cell wall in order to resist external antibacterial substances. LAB require exogenous amino acids and peptides, which are provided by the hydrolysis of casein in fermented milk. Many LAB are deficient in a variety of amino acids [46]. In addition to enabling microbial growth, polypeptides, amino acids, and their derivatives also contribute to the distinctive texture and flavor of fermented milk, so the strains from fermented milk contained more genes associated with the metabolism of amino acids and their derivatives. These results also show that the functional genes of L. pontis in particular living environments function differently in their hosts.
In addition, all L. pontis strains contained genes for the production of extracellular polysaccharides, which can protect against external stresses. Because L. pontis contained more genes associated with carbohydrate metabolism, we also undertook a carbohydrate enzyme analysis. There were differences in the composition of carbohydrate-active enzymes. L. pontis contained eight GT families, among which GT2 and GT4 were the most widely distributed. Studies have shown that GT2 and GT4 are rich in glycosyltransferases in naturally fermented milk. In fermented milk, these substances, especially polysaccharide compounds, are related to the final viscosity and water-holding capacity of fermented milk products. The L. pontis strains were rich in GT2 and GT4, indicating that they had stronger sugar synthesis abilities. In addition, the GT32 family, which participates in the conversion of mannose, glucose, galactose, and others, and the GT101 family, which represents β-glucosyltransferase, were found only in L. pontis MAG.251, which may be related to its animal origin. GH53 was only found in sourdough isolates, which is related to its ability to degrade hemicellulose [13]. Carbohydrates are the main constituents of the cells and the main energy source and play a regulatory role in life activities [47]. Thus, the strains of L. pontis from different sources had different carbohydrate-active enzymes in order to adapt to their different habitats.
Bacteriocins are bacteriostatic substances produced by LAB in order to compete for ecological niches and enabling them to compete under real-life conditions [33]. Among the five strains of L. pontis, only those from fermented milk had predicted bacteriocins, which may be due to the rich nutritional content of fermented milk, leading to the diversity and abundance of other bacteria; bacteriocin is required for survival under niche competition.
The results of this study provide a reference for an understanding of the evolution of L. pontis. The functional genes of strains from different sources varied in order to adapt to various environments. L. pontis from fermented milk contained numerous carbohydrate-related genes and more genes for cell wall, capsule, and amino acid metabolism and its derivatives than strains from other sources. Protein metabolism and fatty acid, lipid, and isoprene metabolism genes were significantly more abundant in sourdough strains than other strains. In addition, all L. pontis strains contained genes for the production of extracellular polysaccharides, which protect from outside attacks. The two L. pontis strains from fermented milk exhibited the bacteriocin operon, which increases their application potential.
In order to adapt to different habitats, strains from different sources possess different phylogenetic relationships, functional genomic characteristics, carbohydrate enzymes, and bacteriocins. At present, there are few published genomes for L. pontis, which limits the in-depth study of the species, and it may be necessary to further explore the appropriate culture conditions and increase the number of strains from various sources and samples for research.
5. Conclusions
In this study, the genome sequences of five L. pontis strains provided a basis for an analysis of this species. L. pontis was characterized by high G + C content and potential niche adaptations. Moreover, the strains of L. pontis from different sources had a range of genetic diversity, which was closely related to their living environments; this lays a foundation for an exploration of the genetic basis and molecular evolution rules of L. pontis’ adaptation to different habitats. However, due to the limited number of available strain sequences currently, more strain sequences need to be obtained for further comparative genomics studies to verify this. As more L. pontis strains are sequenced in the future, more L. pontis genomes from different environments can be compared, which will provide a strong indication of the factors affecting their genetic diversity and habitat adaptation.
Author Contributions
Conceptualization, X.L. and Q.L.; methodology, X.L., Q.L. and W.L. (Weicheng Li); formal analysis, W.L. (Weicheng Li), Y.L. and L.Z.; writing—review and editing, X.L.; resources, W.L. (Wenjun Liu); data curation, W.L. (Wenjun Liu). All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by grants from the National Natural Science Foundation of China (U23A20260), the Science and Technology Development Fund Project for Central Guiding Local development (2022ZY0209) and the Inner Mongolia Natural Science Foundation Outstanding Youth Fund project (2023JQ07), both awarded to W.L. (Wenjun Liu).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Vogel, R.F.; BöCKER, G.; Stolz, P.; Ehrmann, M.; Fanta, D.; Ludwig, W.; Pot, B.; Kersters, K.; Schleifer, K.H.; Hammes, W.P. Identification of Lactobacilli from sourdough and description of Lactobacillus pontis sp. nov. Int. J. Syst. Evol. Microbiol. 1994, 44, 223–229. [Google Scholar] [CrossRef][Green Version]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.; Harris, H.M.; Mattarelli, P.; O’toole, P.W.; Pot, B.; Vandamme, P.; Walter, J. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Maya-Barrios, A.; Lira-Hernandez, K.; Jiménez-Escobar, I.; Hernández, L.; Ortiz-Hernandez, A.; Jiménez-Gutiérrez, C.; López-Velázquez, G.; Gutiérrez-Castrellón, P. Limosilactobacillus reuteri ATCC PTA 5289 and DSM 17938 as adjuvants to improve evolution of pharyngitis/tonsillitis in children: Randomised controlled trial. Benef. Microbes 2021, 12, 137–145. [Google Scholar] [CrossRef]
- Bao, Y.; Zhang, Y.; Zhang, Y.; Liu, Y.; Wang, S.; Dong, X.; Wang, Y.; Zhang, H. Screening of potential probiotic properties of Lactobacillus fermentum isolated from traditional dairy products. Food Control 2010, 21, 695–701. [Google Scholar] [CrossRef]
- Shazadi, K.; Arshad, N. Evaluation of inhibitory and probiotic properties of lactic acid bacteria isolated from vaginal microflora. Folia Microbiol. 2022, 67, 427–445. [Google Scholar] [CrossRef]
- Saviano, A.; Brigida, M.; Migneco, A.; Gunawardena, G.; Zanza, C.; Candelli, M.; Franceschi, F.; Ojetti, V. Lactobacillus Reuteri DSM 17938 (Limosilactobacillus reuteri) in diarrhea and constipation: Two sides of the same coin? Medicina 2021, 57, 643. [Google Scholar] [CrossRef] [PubMed]
- Talarico, T.; Casas, I.; Chung, T.C.; Dobrogosz, W. Production and isolation of reuterin, a growth inhibitor produced by Lactobacillus reuteri. Antimicrob. Agents Chemother. 1988, 32, 1854–1858. [Google Scholar] [CrossRef]
- Arqués, J.L.; Fernández, J.; Gaya, P.; Nuñez, M.; Rodríguez, E.; Medina, M. Antimicrobial activity of reuterin in combination with nisin against food-borne pathogens. Int. J. Food Microbiol. 2004, 95, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Dobrogosz, W.J.S.E. 5413960 Antibiotic reuterin: Dobrogosz Walter J.; Lindgren Sven E Raleigh, NC, United States Assigned to Biogaia AB. Biotechnol. Adv. 1996, 14, 288. [Google Scholar] [CrossRef]
- Feyereisen, M.; Mahony, J.; Kelleher, P.; Roberts, R.J.; O’Sullivan, T.; Geertman, J.-M.A.; van Sinderen, D. Comparative genome analysis of the Lactobacillus brevis species. BMC Genom. 2019, 20, 416. [Google Scholar] [CrossRef]
- Francis, F.; Kim, J.; Ramaraj, T.; Farmer, A.; Rush, M.C.; Ham, J.H. Comparative genomic analysis of two Burkholderia glumae strains from different geographic origins reveals a high degree of plasticity in genome structure associated with genomic islands. Mol. Genet. Genom. 2013, 288, 195–203. [Google Scholar] [CrossRef]
- Oh, P.L.; Benson, A.K.; Peterson, D.A.; Patil, P.B.; Moriyama, E.N.; Roos, S.; Walter, J. Diversification of the gut symbiont Lactobacillus reuteri as a result of host-driven evolution. ISME J. 2010, 4, 377–387. [Google Scholar] [CrossRef]
- Jiang, J.Y.; Li, K.; Xiao, Y.; Zhong, A.; Tang, J.J.; Duan, Y.F.; Li, Z.J. Limosilactobacillus reuteri Regulating Intestinal Function: A Review. Fermentation 2022, 9, 19. [Google Scholar] [CrossRef]
- Tyagi, A.; Yeon, S.J.; Chelliah, R.; Oh, D.H. Draft genome sequence of Limosilactobacillus reuteri, isolated from human breast milk. Microbiol. Resour. Announc. 2023, 12, 112822. [Google Scholar] [CrossRef]
- Liu, W.; Bao, Q.; Jirimutu; Qing, M.; Siriguleng; Chen, X.; Sun, T.; Li, M.; Zhang, J.; Yu, J. Isolation and identification of lactic acid bacteria from Tarag in Eastern Inner Mongolia of China by 16S rRNA sequences and DGGE analysis. Microbiol. Res. 2012, 167, 110–115. [Google Scholar] [CrossRef]
- Yu, J.; Zhao, J.; Song, Y.; Zhang, J.; Yu, Z.; Zhang, H.; Sun, Z. Comparative genomics of the herbivore gut symbiont Lactobacillus reuteri reveals genetic diversity and lifestyle adaptation. Front. Microbiol. 2018, 9, 1151. [Google Scholar] [CrossRef]
- Tanaka, T.; Sakai, R.; Kobayashi, R.; Hatakeyama, K.; Matsunaga, T. Contributions of phosphate to DNA adsorption/desorption behaviors on aminosilane-modified magnetic nanoparticles. Langmuir 2009, 25, 2956–2961. [Google Scholar] [CrossRef]
- Modi, A.; Vai, S.; Caramelli, D.; Lari, M. The Illumina Sequencing Protocol and the NovaSeq 6000 System. Methods Mol. Biol. 2021, 2242, 15–42. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y. SOAPdenovo2: An empirically improved memory-efficient short-read de novo assembler. GigaScience 2012, 1, 2047-217X-1-18. [Google Scholar] [CrossRef]
- Xu, M.; Guo, L.; Gu, S.; Wang, O.; Zhang, R.; Peters, B.A.; Fan, G.; Liu, X.; Xu, X.; Deng, L. TGS-GapCloser: A fast and accurate gap closer for large genomes with low coverage of error-prone long reads. GigaScience 2020, 9, 94. [Google Scholar] [CrossRef]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA-DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef]
- Chen, J.; Yang, X.; Chen, J.; Cen, Z.; Guo, C.; Jin, T.; Cui, Y. SISP: A Fast Species Identification System for Prokaryotes Based on Total Nucleotide Identity of Whole Genome Sequences. Infect. Dis. Transl. Med. 2015, 1, 30–55. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Duranti, S.; Milani, C.; Lugli, G.A.; Mancabelli, L.; Turroni, F.; Ferrario, C.; Mangifesta, M.; Viappiani, A.; Sánchez, B.; Margolles, A. Evaluation of genetic diversity among strains of the human gut commensal Bifidobacterium adolescentis. Sci. Rep. 2016, 6, 23971. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Cantarel, B.L.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The Carbohydrate-Active EnZymes database (CAZy): An expert resource for glycogenomics. Nucleic Acids Res. 2009, 37, 233–238. [Google Scholar] [CrossRef]
- van Heel, A.J.; de Jong, A.; Song, C.; Viel, J.H.; Kok, J.; Kuipers, O.P. BAGEL4: A user-friendly web server to thoroughly mine RiPPs and bacteriocins. Nucleic Acids Res. 2018, 46, 278–281. [Google Scholar] [CrossRef]
- Johnson, B.; Shneiderman, B. Tree-Maps: A Space-Filling Approach to the Visualization of Hierarchical Information Structures; IEEE: San Diego, CA, USA, 1991; pp. 284–291. [Google Scholar] [CrossRef]
- Wang, S.; Yang, B.; Ross, R.P.; Stanton, C.; Zhao, J.; Zhang, H.; Chen, W. Comparative genomics analysis of Lactobacillus ruminis from different niches. Genes 2020, 11, 70. [Google Scholar] [CrossRef]
- Balakirev, E.S.; Krupnova, T.N.; Ayala, F.J. DNA variation in the phenotypically-diverse brown alga Saccharina japonica. BMC Plant Biol. 2012, 108, 1471–2229. [Google Scholar] [CrossRef]
- Accetto, T.; Avguštin, G. Polysaccharide utilization locus and CAZYme genome repertoires reveal diverse ecological adaptation of Prevotella species. Syst. Appl. Microbiol. 2015, 38, 453–461. [Google Scholar] [CrossRef]
- Cao, H.; Ekstrom, A.; Yin, Y. Plant Carbohydrate Active Enzyme (CAZyme) Repertoires: A Comparative Study; Springer International Publishing: New York, NY, USA, 2015; pp. 115–134. [Google Scholar] [CrossRef]
- Vilma, K.; Mantas, S.; Kristina, B.; Naujokaitytė, G.; Mulkytė, K.; Malakauskas, M.; Maruška, A. Current state of purification, isolation and analysis of bacteriocins produced by lactic acid bacteria. Appl. Microbiol. Biotechnol. 2017, 101, 1323–1335. [Google Scholar] [CrossRef]
- Alvarez-Sieiro, P.; Montalban-Lopez, M.; Mu, D.; Kuipers, O. Bacteriocins of lactic acid bacteria: Extending the family. Appl. Microbiol. Biotechnol. 2016, 100, 2939–2951. [Google Scholar] [CrossRef]
- Sinéad, C.; Li, Y.; Riedel, C.U.; O’Toole, P.W.; Hill, C.; Gahan, C.G.M. From the Cover: Bacteriocin production as a mechanism for the antiinfective activity of Lactobacillus salivarius UCC118. Proc. Natl. Acad. Sci. USA 2007, 104, 761. [Google Scholar] [CrossRef]
- Bobay, L.-M.; Ochman, H. Impact of recombination on the base composition of bacteria and archaea. Mol. Biol. Evol. 2017, 34, 2627–2636. [Google Scholar] [CrossRef]
- Musto, H.; Naya, H.; Zavala, A.; Romero, H.; Alvarez-Valín, F.; Bernardi, G. Correlations between genomic GC levels and optimal growth temperatures in prokaryotes. FEBS Lett. 2004, 573, 73–77. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Mining genome traits that determine the different gut colonization potential of Lactobacillus and Bifidobacterium species. Microb. Genom. 2021, 7, 000581. [Google Scholar] [CrossRef]
- Maldonado-Gómez, M.X.; Martínez, I.; Bottacini, F.; O’Callaghan, A.; Ventura, M.; van Sinderen, D.; Hillmann, B.; Vangay, P.; Knights, D.; Hutkins, R.W. Stable engraftment of Bifidobacterium longum AH1206 in the human gut depends on individualized features of the resident microbiome. Cell Host Microbe 2016, 20, 515–526. [Google Scholar] [CrossRef]
- Lerat, E.; Ochman, H. Recognizing the pseudogenes in bacterial genomes. Nucleic Acids Res. 2005, 33, 3125–3132. [Google Scholar] [CrossRef]
- Jung, J.; Kim, K.; Yoo, D.; Lee, C.; Kang, J.; Cho, K.; Kang, D.-K.; Kwak, W.; Yoon, S.H.; Sohn, H. Comparative genomic analysis of Lactobacillus plantarum GB-LP4 and identification of evolutionarily divergent genes in high-osmolarity environment. Genes Genom. 2018, 40, 217–223. [Google Scholar] [CrossRef]
- Walter, J.; Britton, R.A.; Roos, S. Host-microbial symbiosis in the vertebrate gastrointestinal tract and the Lactobacillus reuteri paradigm. Proc. Natl. Acad. Sci. USA 2011, 108, 4645–4652. [Google Scholar] [CrossRef]
- El Kafsi, H.; Binesse, J.; Loux, V.; Buratti, J.; Boudebbouze, S.; Dervyn, R.; Kennedy, S.; Galleron, N.; Quinquis, B.; Batto, J.M. Lactobacillus delbrueckii ssp. lactis and ssp. bulgaricus: A chronicle of evolution in action. BMC Genom. 2014, 15, 407. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).