

Environmental Variables Outpace Biotic Interactions in Shaping a Phytoplankton Community

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Field Sampling
2.3. Samples and Data Analysis of Abiotic Environmental Variables
2.4. Samples and Data Analysis of Plankton Community
2.5. Statistical Analysis
3. Results
3.1. Regional Climate and Physical and Chemical Conditions
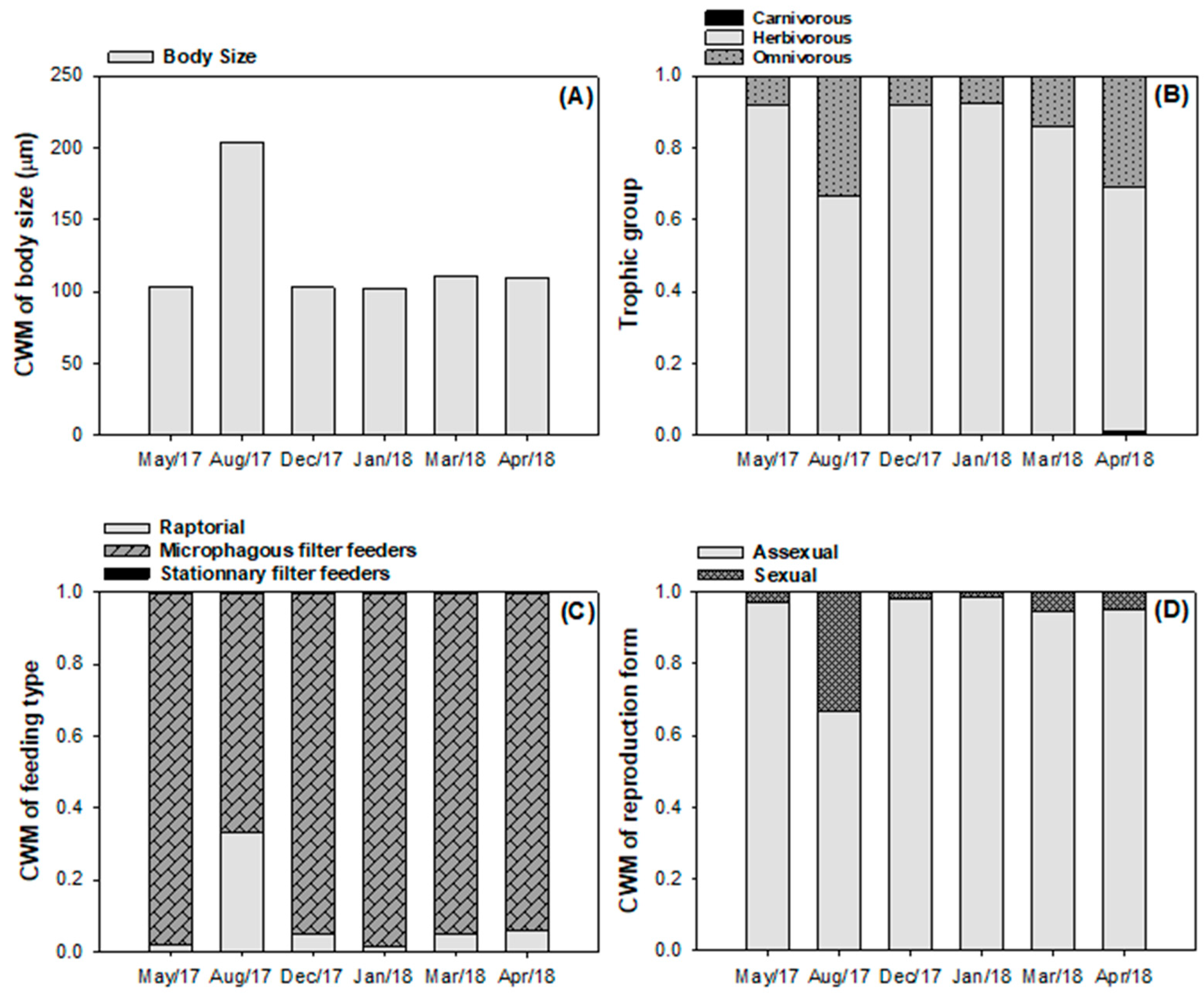
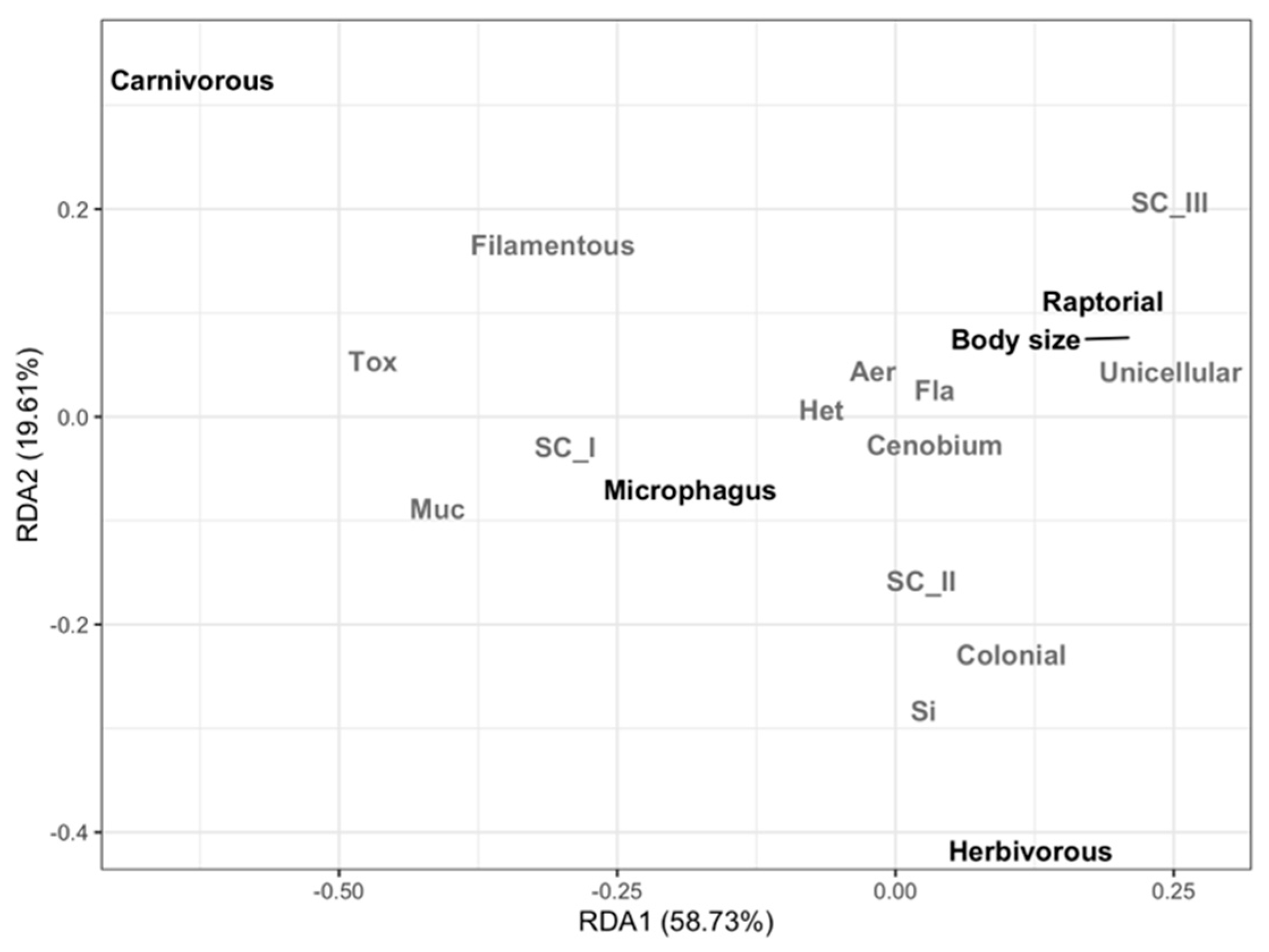
3.2. Taxonomic and Functional Diversity of the Phytoplankton Community
3.3. Taxonomic and Functional Diversity of Zooplankton Community
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reynolds, C.S. Ecology of Phytoplankton; Cambridge University Press: New York, NY, USA, 2006. [Google Scholar]
- Lürling, M. Grazing resistance in phytoplankton. Hidrobiologia 2021, 848, 237–249. [Google Scholar] [CrossRef]
- Costa, M.R.A.; Cardoso, M.M.L.; Selmeczy, G.B.; Padisák, J.; Becker, V. Phytoplankton functional responses induced by extreme hydrological events in a tropical reservoir. Hydrobiologia 2024, 851, 849–867. [Google Scholar] [CrossRef]
- Magalhães, L.; Rangel, L.M.; Rocha, A.M.; Cardoso, S.J.; Silva, L.H.S. Responses of morphology-based phytoplankton functional groups to spatial variation in two tropical reservoirs with long water-residence time. Inland. Waters 2020, 11, 29–43. [Google Scholar] [CrossRef]
- Carvalho, L. Top-down control of phytoplankton in a shallow hypertrophic lake: Little Mere (England). Hydrobiologia 1994, 275–276, 53–63. [Google Scholar] [CrossRef]
- Jeppesen, E.; Sondergaard, M.; Jensen, J.P.; Mortensen, E.; Sortkjaer, O. Fish-induced changes in zooplankton grazing on phytoplankton and bacterioplankton: A long-term study in shallow hypertrophic Lake Sebygaard. J. Plankton Res. 1996, 18, 1605–1625. [Google Scholar] [CrossRef]
- Rangel, L.M.; Soares, M.C.S.; Paiva, R.; Silva, L.H.S. Morphology-based functional groups as effective indicators of phytoplankton dynamics in a tropical cyanobacteria-dominated transitional river–reservoir system. Ecol. Indic. 2016, 64, 217–227. [Google Scholar] [CrossRef]
- Josué, I.I.P.; Cardoso, S.J.; Miranda, M.; Mucci, M.; Ali, K.G.; Roland, F.; Marinho, M.M. Cyanobacteria dominance drives zooplankton functional dispersion. Hydrobiologia 2019, 831, 149–161. [Google Scholar] [CrossRef]
- Heathcote, A.J.; Filstrup, C.T.; Kendall, D.; Downing, J.A. Biomass pyramids in lake plankton: Influence of cyanobacteria size and abundance. Inland. Waters 2016, 6, 250–257. [Google Scholar] [CrossRef]
- Krztoń, W.; Kosiba, J.; Pociecha, A.; Wilk-Woźniak, E. The effect of cyanobacterial blooms on bio- and functional diversity of zooplankton communities. Biodivers. Conserv. 2019, 28, 815–1835. [Google Scholar] [CrossRef]
- Krztoń, W.; Kosiba, J. Variations in zooplankton functional groups density in freshwater ecosystems exposed to cyanobacterial blooms. Sci. Total Environ. 2020, 730. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.R.A.; Attayde, J.L.; Becker, V. Effects of water level reduction on the dynamics of phytoplankton functional groups in tropical semi-arid shallow lakes. Hydrobiologia 2016, 778, 75–89. [Google Scholar] [CrossRef]
- Dalu, T.; Wasserman, R.J. Cyanobacteria dynamics in a small tropical reservoir: Understanding spatio-temporal variability and influence of environmental variables. Sci. Total Environ. 2018, 643, 835–841. [Google Scholar] [CrossRef] [PubMed]
- Mesquita, M.C.B.; Lürling, M.; Door, F.; Pinto, E.; Marinho, M.M. Combined effect of light and temperature on the production of saxitoxins in Cylindrospermopsis raciborskii strains. Toxins 2019, 11, 38. [Google Scholar] [CrossRef]
- Mesquita, M.C.B.; Prestes, A.C.C.; Gomes, A.M.A.; Marinho, M.M. Direct effects of temperature on growth of different tropical phytoplankton species. Microb. Ecol. 2020, 79, 1–11. [Google Scholar] [CrossRef]
- Barbosa, L.G.; Barbosa, F.A.R.; Bicudo, C.E.M. Is thermal stability a factor that influences environmental heterogeneity and phytoplankton distribution in tropical lakes? Acta Limnol. 2018, 30. [Google Scholar] [CrossRef]
- Longhi, M.L.; Beinser, B.E. Patterns in taxonomic and functional diversity of Lake Phytoplankton. Freshw. Biol. 2010, 55, 1349–1366. [Google Scholar] [CrossRef]
- Rangel, L.M.; Silva, L.H.S.; Rosa, P.; Roland, F.; Huszar, V.L.M. Phytoplankton biomass is mainly controlled by hydrology and phosphorus concentrations in tropical hydroelectric reservoirs. Hydrobiologia 2012, 693, 3–28. [Google Scholar] [CrossRef]
- Reynolds, C.S. Vegetation Processes in the Pelagic: A Model for Ecosystem Theory; Ecology Institute: Oldendorf, Germany, 1997. [Google Scholar]
- Scheffer, M. Ecology of Shallow Lakes; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998; 357p. [Google Scholar]
- Leitão, E.; Ger, K.A.; Panosso, R. Selective grazing by a tropical copepod (Notodiaptomus iheringi) facilitates Microcystis dominance. Front. Microbiol. 2018, 9, 301. [Google Scholar] [CrossRef]
- Leitão, E.; Panosso, R.; Molica, R.; Ger, K.A. Top-down regulation of filamentous cyanobacteria varies among a raptorial versus current feeding copepod across multiple prey generations. Freshw. Biol. 2020, 66, 142–156. [Google Scholar] [CrossRef]
- Severiano, J.S.; Almeida-Melo, V.L.S.; Bittencourt-Oliveira, M.C.; Chia, M.A.; Moura, A.N. Effects of increased zooplankton biomass on phytoplankton and cyanotoxins: A tropical mesocosm study. Harmful Algae 2018, 71, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Faithfull, C.; Goetze, E. Copepod nauplii use phosphorus from bacteria, creating a short circuit in the microbial loop. Ecol. Lett. 2019, 22, 1462–1471. [Google Scholar] [CrossRef]
- Jagadeesan, L.; Jyothibabu, R.; Arunpandi, N.; Anjusha, A.; Parthasarathi, S.; Pandiyarajan, R.S. Feeding preference and daily ration of 12 dominant copepods on mono and mixed diets of phytoplankton, rotifers, and detritus in a tropical coastal water. Environ. Monit. Assess. 2017, 189, 503. [Google Scholar] [CrossRef]
- Colina, M.; Calliari, D.; Carballo, C.; Kruk, C. A trait-based approach to summarize zooplankton– phytoplankton interactions in freshwaters. Hydrobiologia 2016, 767, 221–233. [Google Scholar] [CrossRef]
- Sommer, U.; Sommer, F.; Santer, B.; Jamieson, C.; Boersma, M.; Becker, C.; Hansen, T. Complementary impact of copepods and cladocerans on phytoplankton. Ecol. Lett. 2001, 4, 545–550. [Google Scholar] [CrossRef]
- Sommer, U.; Soomer, F. Cladocerans versus copepods: The cause of contrasting top–down controls on freshwater and marine phytoplankton. Oecologia 2006, 147, 183–194. [Google Scholar] [CrossRef]
- Rangel, L.M.; Silva, L.H.S.; Faassen, E.J.; Lürling, M.; Ger, K.A. Copepod prey selection and grazing efficiency mediated by chemical and morphological defensive traits of cyanobacteria. Toxins 2020, 12, 465. [Google Scholar] [CrossRef]
- Amorim, C.H.; Valença, C.R.; Moura-Falcão, R.H.; Moura, A.N. Seasonal variations of morpho-functional phytoplankton groups influence the top-down control of a cladoceran in a tropical hypereutrophic lake. Aquat. Ecol. 2019, 53, 453–464. [Google Scholar] [CrossRef]
- Severiano, J.S.; Amaral, C.B.; Diniz, A.S.; Moira, A.N. Species-specific response of phytoplankton to zooplankton grazing in tropical eutrophic reservoirs. Acta Limnol. 2021, 33. [Google Scholar] [CrossRef]
- Kiørboe, T. How zooplankton feed: Mechanisms, traits and trade-offs. Biol. Rev. 2011, 86, 311–339. [Google Scholar] [CrossRef]
- Kruk, C.; Huszar, V.L.M.; Peeters, E.T.H.; Bonilla, S.; Costa, L.; Lürling, M.; Reynolds, C.S.; Scheffer, M. A morphological classification capturing functional variation in phytoplankton. Freshw. Biol. 2010, 55, 614–627. [Google Scholar] [CrossRef]
- Litchman, E.; Ohman, M.D.; Kiørboe, T. Trait-based approaches to zooplankton communities. J. Plankton Res. 2013, 35, 473–484. [Google Scholar] [CrossRef]
- Violle, C.; Navas, M.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Carscadden, K.; Mirotchnick, N. Beyond species: Functional diversity and the maintenance of ecological processes and services. J. App Ecol. 2011, 48, 1079–1087. [Google Scholar] [CrossRef]
- Barnett, A.J.; Finlay, K.; Beisner, B.E. Functional diversity of crustacean zooplankton communities: Towards a trait-based classification. Freshw. Biol. 2007, 52, 796–813. [Google Scholar] [CrossRef]
- Graco-Roza, C.; Soinine, J.; Corrêa, G.; Pacheco, F.S.; Miranda, M.; Domingos, P.; Marinho, M.M. Functional rather than taxonomic diversity reveals changes in the phytoplankton community of a large dammed river. Ecol. Indic. 2021, 121, 107048. [Google Scholar] [CrossRef]
- Inepac (Instituto Estadual do Patrimônio Cultural). Inventário de identificação dos reservatórios da CEDAE. Levantado por M.G. Ferraz, M.G. Mendonça e Rui Veloso (1998) e Iracema Franco (2006). Secretaria de Estado de Cultura, RJ. Available online: https://www.ipatrimonio.org/wp-content/uploads/2019/08/ipatrimonio-Rio-de-Janeiro-Reservatorio-Livramento-Fonte-Inepac-compactado.pdf (accessed on 12 April 2023).
- Menezes, M. New species of pigmented flagellates from southeastern Brazil. Arch. Protistenkd. 1996, 147, 101–105. [Google Scholar] [CrossRef]
- Menezes, M.; Bicudo, C.E.M. Flagellate green algae from four water bodies in the state of Rio de Janeiro, Southeast Brazil. Hoehnea 2008, 35, 435–468. [Google Scholar] [CrossRef]
- Pereira, U.J. Regulação do Fitoplâncton, Dinâmica Trófica Pelágica e Abordagem Experimental Aplicada ao Biocontrole de Cylindrospermopsis raciborskii em Reservatório Eutrófico Tropical (Reservatório do Camorim, Parque Estadual da Pedra Branca, RJ). Ph.D. Thesis, Federal University of Rio de Janeiro, Rio de Janeiro, Brazil, 2018. [Google Scholar]
- Cole, G.A. Textbook of Limnology; Waveland Press Inc.: Long Grove, IL, USA, 1994. [Google Scholar]
- Padisák, J.; Barbosa, F.; Koschel, R.; Krienitz, L. Deep layer cyanoprokaryota maxima in temperate and tropical lakes. Arch. Hydrobiol. Spec. Issues Advanc. Limnol. 2003, 58, 175–199. [Google Scholar]
- Sas, H. Lake Restoration by Reduction of Nutrient Loading: Expectations, Experiences, Extrapolations; Academia Verlag Richarz: St. Augustin, FL, USA, 1989. [Google Scholar]
- Uhelinger, V. Étude statistique des méthodes de dénobrement planctonique. Arch. Sci. 1964, 17, 121–123. [Google Scholar]
- Utermöhl, H. Zur Vervollkommnung der quantitativen Phytoplankton-Methodik: Mit 1 Tabelle und 15 abbildungen im Text und auf 1 Tafel. Int. Ver. Theor. Angew. Limnol. Mitteilungen 1958, 9, 1–38. [Google Scholar] [CrossRef]
- Lund, J.W.G.; Klipling, C.; Le Cren, E.D. The inverted microscope method of estimating algal numbers and the statistical basis of estimating by counting. Hydrobiologia 1958, 11, 143–170. [Google Scholar] [CrossRef]
- Hoek, C.V.D.; Mann, D.G.; Jahns, H.M. Algae: An Introduction to Phycology; Cambridge University Press: New York, NY, USA, 1995. [Google Scholar]
- Round, F.E.; Crawford, R.M.; Mann, D.G. The Diatoms. Biology and Morphology of Genera; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota I. Teil Chroococcales; Ettl, H., Gärtner, G., Heynig, H., Mollenhauer, D., Eds.; Süsswasserflora von Mitteleuropa; Gustav Fischer Verlag: Jena, Germany, 1999. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota 2. Teil Oscillatoriales. Büdel, B., Krienitz, L., Gärtner, G., Schagerl, M., Eds.; Süsswasserflora von Mitteleuropa; Gustav Fischer Verlag: Jena, Germany, 2005. [Google Scholar]
- Hillebrand, H.; Dürseken, D.; Kirschiel, D.; Pollingher, U.; Zohary, T. Biovolume calculation for pelagic and benthic microalgae. J. Phycol. 1999, 35, 403–424. [Google Scholar] [CrossRef]
- Carpenter, E.J.; Subramaniam, A.; Capone, D.G. Biomass and primary productivity of the cyanobacterium Trichodesmium spp. in the tropical N Atlantic Ocean. Deep-Sea Res. 2004, 51, 173–203. [Google Scholar] [CrossRef]
- Menden-Deuer, S.; Lessard, E.J. Carbon to volume relationships for dinnoflagellates, diatoms, and other protist plankton. L&O 2000, 4, 569–579. [Google Scholar]
- Montagnes, D.J.S.; Franklim, D.J. Effect of temperature on diatom volume, growth rate, and carbon and nitrogen content: Reconsidering some paradigms. L&O 2001, 46, 2008–2018. [Google Scholar]
- Verity, P.G.; Robertson, C.; Tronzo, C.R.; Andrews, M.G.; Nelson, J.R.; Sieracki, M.E. Relationships between cell volume and the carbon and nitrogen content of marine photosynthetic nanoplankton. L&O 1992, 37, 1434–1446. [Google Scholar]
- Prepas, E.E. Some statistical methods for the design of experiments and analysis of samples. In A Manual on Methods for the Assessment of Secondary Productivity in Fresh Waters, 2nd ed.; Downing, J.A., Rigler, F.H., Eds.; Blackwell Scientific Publications: Oxford, UK, 1984; pp. 266–335. [Google Scholar]
- Ruttner-Kolisko, A. Suggestions for biomass calculation of plankton rotifers. Arch. Hydrobiol. 1977, 8, 71–76. [Google Scholar]
- Mccauley, E. The estimation of the abundance and biomass of zooplankton in samples. In A Manual on Methods for the Assessment of Secondary Productivity in Fresh Waters, 2nd ed.; Dowing, J.A., Rigler, F.H., Eds.; Blackwell Scientific Publications: Oxford, UK, 1984; pp. 228–264. [Google Scholar]
- Bottrell, H.H.; Duncan, A.; Gliwicz, Z.M.; Grygierek, E.; Herzig, A.; Hillbrichtilkowska, A.; Kurasawa, H.; Larsson, P.; Weglenska, T. Are view of some problems in zooplankton production studies. Nor. J. Zool. 1976, 24, 419–456. [Google Scholar]
- Pace, M.L.; Orcutt, J.D. The relative importance of protozoans, rotifers, and crustaceans in a freshwater zooplankton community. L&O 1981, 26, 822–830. [Google Scholar]
- Idris, B.A.G.; Fernando, C.H. Two new species of cladoceran crustaceans of the genera Macrothrix baird and Alona baird from Malaysia. Hydrobiologia 1981, 76, 8–85. [Google Scholar] [CrossRef]
- Masundire, H.M. Mean individual dry weight and length-weight regressions of some zooplankton of Lake Kariba. Hydrobiologia 1994, 272, 231–238. [Google Scholar] [CrossRef]
- Matsumura-Tundisi, T.; Rietzler, A.C.; Tundisi, J.G. Biomass (dry weight and carbon content) of plankton crustacea from Broa reservoir (Sao Carlos, S.P.-Brazil) and its fluctuation across one year. Hydrobiologia 1989, 179, 229–236. [Google Scholar] [CrossRef]
- Panarelli, E.A.; Casanova, S.M.C.; Henry, R. Secondary production and biomass of Cladocera in marginal lakes after the recovery of their hydrologic connectivity in a river–reservoir transition zone. L&O Res. Manag. 2010, 15, 319–334. [Google Scholar]
- Culver, D.A.; Boucherle, M.M.; Bean, D.J.; Fletcher, J.W. Biomass of freshwater crustacean zooplankton from length-weight regressions. Can. J. Fish. Aquat. Sci. 1985, 42, 1380–1390. [Google Scholar] [CrossRef]
- Burns, C.W. Relation between filtering rate, temperature and body size in four species of Daphnia. L&O 1969, 14, 693–700. [Google Scholar]
- Edmondson, W.T.; Winberg, G.G. A Manual on Methods for the Assessment of Secondary Productivity in Fresh Waters; IBP Handbook No. 17; Blackwell: Oxford, UK, 1971; p. 358. [Google Scholar]
- Hessen, D.O. Factors determining the nutritive status and production of zooplankton in a humic lake. J. Plankton Res. 1989, 11, 649–664. [Google Scholar] [CrossRef]
- Persson, G.; Ekbohm, G. Estimation of dry weight in zooplankton populations: Methods applied to crustacean populations from lakes in the Kuokkel Area, Northern Sweden. Arch. Hydrobiol. 1980, 59, 225–246. [Google Scholar]
- Rosen, R.H. Length-dry weight relationships of some freshwater zooplankton. J. Freshw. Ecol. 1981, 1, 225–229. [Google Scholar] [CrossRef]
- Latja, R.; Salomen, K. Carbon analysis for determination of individual biomasses of planktonic animals. Verh. Internat Verein Limnol. 1978, 20, 2556–2560. [Google Scholar] [CrossRef]
- Lokko, K.; Virro, T.; Kotta, J. Seasonal variability in the structure and functional diversity of psammic rotifer communities: Role of environmental parameters. Hydrobiologia 2017, 796, 287–307. [Google Scholar] [CrossRef]
- Obertegger, U.; Smith, H.A.; Flaim, G.; Wallace, R.L. Using the guild ratio to characterize pelagic rotifer communities. Hydrobiologia 2011, 662, 157–162. [Google Scholar] [CrossRef]
- Dolédec, S.; Chessel, D.; Ter Braak, C.J.F.; Champely, S. Matching species traits to environmental variables: A new three-table ordination method. Environ. Ecol. Stat. 1996, 3, 143–166. [Google Scholar] [CrossRef]
- Dray, S.; Choler, P.; Dolédec, S.; Peres-Neto, P.R.; Thuiller, W.; Pavoine, S.; Ter Braak, C.J.F. Combining the fourth-corner and the RLQ methods for assessing trait responses to environmental variation. Ecology 2014, 95, 14–21. [Google Scholar] [CrossRef]
- Legendre, P.; Galzin, R.; Harmelin-Vivien, M.L. Relating behavior to habitat: Solutions to the fourth-corner problem. Ecology 1997, 78, 547–562. [Google Scholar] [CrossRef]
- Dray, S.; Dufour, A.B. The ade4 package: Implementing the duality diagram for ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef]
- Ricotta, C.; Moretti, M. CWM and Rao’s quadratic diversity: A unified framework for functional ecology. Oecologia 2011, 167, 181–188. [Google Scholar] [CrossRef]
- Pla, L.; Casanoves, F.; Di Rienzo, J. Quantifying Functional Biodiversity; Springer: Dordrecht, The Netherlands, 2011. [Google Scholar]
- Laliberté, E.; Legendre, P. A distance-based frameworkfor measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B. FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology. R Package Version 1. 2014, 0–12. Available online: https://citeseerx.ist.psu.edu/document?repid=rep1&type=pdf&doi=c0123be0f0737359fa9303aece03fd500d2499b1 (accessed on 25 May 2024).
- Kleyer, M.; Dray, S.; Bello, F.; Leps, J.; Pakeman, R.J.; Strauss, B.; Thuiller, W.; Lavorel, S. Assessing species and community functional responses to environmental gradients: Which multivariate methods? J. Veg. Sci. 2012, 23, 805–821. [Google Scholar] [CrossRef]
- Rangel, L.M.; Silva, L.H.S.; Arcifa, M.S.; Perticarrari, A. Driving forces of the diel distribution of phytoplankton functional groups in a shallow tropical lake (Lake Monte Alegre, Southeast Brazil). Braz. J. Biol. 2009, 69, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Garnier, E.; Lavorel, S.; Ansquer, P.; Castro, H.; Cruz, P.; Dolezal, J.; Eriksson, O.; Fortunel, C.; Grigulis, K. Assessing the effects of land-use change on plant traits, communities and ecosystem functioning in grasslands: A standardized methodology and lessons from an application to 11 European sites. Ann. Bot. 2007, 99, 967–985. [Google Scholar] [CrossRef] [PubMed]
- Bergström, A.N.; Jansson, M.; Drakare, S.; Blomqvist, P. Occurrence of mixotrophic flagellates in relation to bacterioplankton production, light regime and availability of inorganic nutrients in unproductive lakes with differing humic contents. Freshw. Biol. 2003, 48, 868–877. [Google Scholar] [CrossRef]
- Ahlgren, G.; Lundstedt, L.; Brett, M.; Forsberg, C. Lipid composition and food quality of some freshwater phytoplankton for cladoceran zooplankters. J. Plankton Res. 1990, 12, 809–818. [Google Scholar] [CrossRef]
- Boersma, M. The nutritional quality of P-limited algae for Daphnia. L&O 2000, 45, 1157–1161. [Google Scholar]
- Gasol, J.M.; Duarte, C.M. Comparative analyses in aquatic microbial ecology: How far do they go? FEMS Microbiol. Ecol. 2000, 31, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Litchman, E.; Klausmeier, C.A. Trait-based community ecology of phytoplankton. Annu. Rev. Ecol. Evol. Syst. 2007, 39, 615–639. [Google Scholar] [CrossRef]
- Lopes, M.R.M.; Bicudo, C.E.M.; Ferragut, M.C. Short term spatial and temporal variation of phytoplankton in a shallow tropical oligotrophic reservoir, southeast Brazil. Hydrobiologia 2005, 542, 235–247. [Google Scholar] [CrossRef]
- Marinho, M.M.; Azevedo, S.M.F.O. Influence of N/P ratio on competitive abilities for nitrogen and phosphorus by Microcystis aeruginosa and Aulacoseira distans. Aquat. Ecol. 2007, 41, 525–533. [Google Scholar] [CrossRef]
- Boersma, M.; Mathew, K.A.; Niehoff, B.; Schoo, K.L.; Franco-Santos, R.M.; Meunier, C.L. Temperature driven changes in the diet preference of omnivorous copepods: No more meat when it’s hot? Ecol. Lett. 2016, 19, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Gusha, M.N.C.; Dalu, T.; Wasserman, R.J.; McQuaid, C.D. Zooplankton grazing pressure is insufficient for primary producer control under elevated warming and nutrient levels. Sci. Total Environ. 2019, 651, 410–418. [Google Scholar] [CrossRef] [PubMed]
- Angilletta, M.J.; Dunham, A.E. The temperature-size rule in ectotherms: Simple evolutionary explanations may not be general. Am. Nat. 2003, 162, 332–342. [Google Scholar] [CrossRef]
- Havens, K.E.; Pinto-Coelho, R.M.; Beklioğlu, M.; Christoffersen, K.S.; Jeppsen, R.; Lauridsen, T.L.; Mazumde, A.; Méthot, G.; Alloul, B.P.; Tavşanoğlu, U.N.; et al. Temperature effects on body size of freshwater crustacean zooplankton from Greenland to the tropics. Hydrobiologia 2015, 743, 27–35. [Google Scholar] [CrossRef]
- Burns, C.W. The relationship between body size of filter-feeding Cladocera and the maximum size of particle ingested. L&O 1968, 13, 675–678. [Google Scholar]
- Lenz, J. Introduction. In Zooplankton Methodology Manual; Harris, R.P., Wiebe, P.H., Lenz, J., Skjoidal, H.R., Huntley, M., Eds.; Academic Press: San Diego, CA, USA, 2000. [Google Scholar]
- Arndt, H. Rotifers as predators on components of the microbial web (bacteria, heterotrophic flagellates, ciliates)—A review. Hydrobiologia 1993, 255, 231–246. [Google Scholar] [CrossRef]
- Ger, K.A.; Leitao, E.; Panosso, R. Potential mechanisms for the tropical copepod Notodiaptomus to tolerate Microcystis toxicity. J. Plankton Res. 2016, 38, 843–854. [Google Scholar] [CrossRef]
- Titocci, J.; Bom, M.; Fink, P. Morpho-functional traits reveal differences in size fractionated phytoplankton communities but do not significantly affect zooplankton grazing. Microorganisms 2022, 10, 182. [Google Scholar] [CrossRef]
- Hansen, B. The size ratio between planktonic predators and their prey. L&O 1994, 39, 395–403. [Google Scholar]
- Fuchs, H.; Franks, P. Plankton community Properties determined by nutrients and size-selective feeding. Mar. Ecol. Prog. Ser. 2010, 413, 1–15. [Google Scholar] [CrossRef]
- Eskinazi Sant’Anna, E.M.; Maia-Barbosa, P.M.; Barbosa, F.A.R. On the natural diet of Daphnia laevis in the eutrophic Pampulha Reservoir (Belo Horizonte, Minas Gerais). Braz. J. Biol. 2002, 62, 445–452. [Google Scholar] [CrossRef]
- Rothhaupt, K.O. Changes of the functional responses of the rotifers Brachionus rubens and Brachionus calyciflorus with particle sizes. L&O 1990, 35, 24–32. [Google Scholar]
- Miracle, M.R.; Vicente, E.; Sarma, S.S.S.; Nandino, S. Planktonic rotifer feeding in hypertrophic conditions. Int. Rev. Hydrobiol. 2014, 99, 141–150. [Google Scholar] [CrossRef]
- Soares, M.C.S.; Lürling, M.; Huszar, V.L.M. Responses of the rotifer Brachionus calyciflorus to two tropical toxic cyanobacteria (Cylindrospermopsis raciborskii and Microcystis aeruginosa) in pure and mixed diets with green algae. J. Plankton Res. 2010, 32, 999–1008. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (MBFG) | Descript | Representative Taxa | Toxicity | Grazing Susceptibility |
|---|---|---|---|---|
 | Small organisms with high surface/volume | Chlorella minutissima; Monoraphidium minutum | No | High |
 | Small-flagellated organisms with siliceous exoskeletal structures | Chromulina gyrans; Dinobryon cylindricum | No | Low |
 | Large filaments with aerotopes | Dolichospermum sp.; Raphidiopsis raciborskii | Yes | Low |
 | Organisms of medium size lacking specialized traits | Scenedesmus acutus; Chlorella sp. | No | High |
 | Organisms of medium size lacking specialized traits | Scenedesmus acutus; Chlorella sp. | No | High |
 | Non-flagellated organisms with siliceous exoskeletons | Thalassiosira weissflogi; Cyclotella sp. | No | Medium |
 | Large mucilaginous colonies | Microcystis aeruginosa | Yes | Low |
| Months | Maximum Depth (m) | Euphotic Zone (m) | RWCS |
|---|---|---|---|
| May 2017 | 2.50 | 2.50 | 33.95 |
| August 2017 | 2.70 | 2.70 | 21.93 |
| December 2017 | 2.10 | 2.10 | 63.62 |
| January 2018 | 2.90 | 2.90 | 145.59 |
| March 2018 | 2.70 | 2.70 | 100.70 |
| April 2018 | 2.30 | 2.16 | 38.89 |
| Surface | Bottom | |||||||
|---|---|---|---|---|---|---|---|---|
| Min | Max | Med | CV | Min | Max | Med | CV | |
| SRP | 3.5 | 6.2 | 4.8 | 0.19 | 3.6 | 5.3 | 4.8 | 0.12 |
| DIN | 46.7 | 170.5 | 103.9 | 0.40 | 89.3 | 236.2 | 165.7 | 0.28 |
| SRSi | 39.6 | 836.7 | 271.3 | 0.84 | 8.6 | 1368.2 | 153.8 | 1.38 |
| DIN:SRP | 4.4 | 36.9 | 18 | 0.54 | 10.7 | 53.9 | 27.9 | 0.48 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mesquita, M.C.B.; Graco-Roza, C.; de Magalhães, L.; Ger, K.A.; Marinho, M.M. Environmental Variables Outpace Biotic Interactions in Shaping a Phytoplankton Community. Diversity 2024, 16, 438. https://doi.org/10.3390/d16080438
Mesquita MCB, Graco-Roza C, de Magalhães L, Ger KA, Marinho MM. Environmental Variables Outpace Biotic Interactions in Shaping a Phytoplankton Community. Diversity. 2024; 16(8):438. https://doi.org/10.3390/d16080438
Chicago/Turabian StyleMesquita, Marcella C. B., Caio Graco-Roza, Leonardo de Magalhães, Kemal Ali Ger, and Marcelo Manzi Marinho. 2024. "Environmental Variables Outpace Biotic Interactions in Shaping a Phytoplankton Community" Diversity 16, no. 8: 438. https://doi.org/10.3390/d16080438
APA StyleMesquita, M. C. B., Graco-Roza, C., de Magalhães, L., Ger, K. A., & Marinho, M. M. (2024). Environmental Variables Outpace Biotic Interactions in Shaping a Phytoplankton Community. Diversity, 16(8), 438. https://doi.org/10.3390/d16080438