Abstract
The Late Cretaceous of Western North America (Laramidia) supported a diverse dinosaur fauna, with duckbilled dinosaurs (Hadrosauridae) being among the most speciose and abundant members of this assemblage. Historically, collecting and preservational biases have meant that dinosaurs from Mexico and the American Southwest are poorly known compared to those of the northern Great Plains. However, evidence increasingly suggests that distinct species and clades inhabited southern Laramidia. Here, a new kritosaurin hadrosaurid, represented by the anterior part of a skull, is reported from the late Campanian of the Cerro del Pueblo Formation, ~72.5 Ma, in Coahuila, Mexico. The Cerro del Pueblo Formation kritosaur was originally considered to represent the same species as a saurolophine from the Olmos Formation of Sabinas, but the Sabinas hadrosaur is now considered a distinct taxon. More recently, the Cerro del Pueblo Formation kritosaur has been referred to Kritosaurus navajovius. We show it represents a new species related to Gryposaurus. The new species is distinguished by its large size, the shape of the premaxillary nasal process, the strongly downturned dentary, and massive denticles on the premaxilla’s palatal surface, supporting recognition of a new taxon, Coahuilasaurus lipani. The dinosaur assemblage of the Cerro del Pueblo Formation shows higher diversity than the contemporaneous fauna of the Horseshoe Canyon Formation in Alberta. Furthermore, Kritosaurini, Lambeosaurini, and Parasaurolophini all persist into the latest Campanian in southern Laramidia after disappearing from northern Laramidia. These patterns suggest declining herbivore diversity seen at high latitudes may be a local, rather than global phenomenon, perhaps driven by cooling at high latitudes in the Late Campanian and Maastrichtian.
Keywords:
Dinosauria; Hadrosauridae; Saurolophinae; Kritosaurini; Cretaceous; Campanian; Laramidia; biogeography 1. Introduction
The Campanian-Maastrichtian of western North America, or Laramidia, hosted a highly diverse dinosaur fauna [1]. Extensive fieldwork and study have made these faunas some of the best-known in the world and so important for understanding patterns of dinosaur evolution and extinction. Historically, collecting and studying have focused on northern Laramidia, particularly Alberta, Montana, Wyoming, and the Dakotas. Here, Cretaceous beds are extensively exposed, highly fossiliferous, and relatively easy to access, giving us perhaps the most detailed picture of dinosaurian ecosystems known from any time or place in the world.
However, fieldwork in the American Southwest and Mexico has slowly improved our understanding of these faunas and has shown that distinct species inhabited southern Laramidia [2,3,4,5,6,7,8,9,10]. Furthermore, at least some dinosaur clades were endemic to the south [4]. Community structures may have differed between the south and north as well, as suggested by the existence of giant titanosaurs [4,7], lambeosaurins [9], and kritosaurs [10] in the south, along with distinct paleofloras [11,12,13,14,15]. These differences also raise the possibility of regionally distinct patterns of evolution. Speciation and extinction rates may have differed between northern and southern Laramidia, as observed for Cenozoic ecosystems [16,17,18], and diversity trends could differ as well.
The southernmost dinosaur fauna known from the Late Cretaceous of Laramidia is that of the Cerro del Pueblo Formation in Coahuila State, Mexico (Figure 1). The Cerro del Pueblo Formation is dated to the end of the Campanian, approximately ~72.5 Ma [19,20,21], on the basis of ammonite biostratigraphy and magnetostratigraphy [19,20,21]. Strontium isotope dating of marine fossils suggests an age of 73 Ma ± 1 Ma [22], consistent with this assignment. The Cerro del Pueblo Formation is, therefore, contemporary with the lowermost Drumheller Member [23] of the Horseshoe Canyon Formation of Alberta [23,24].
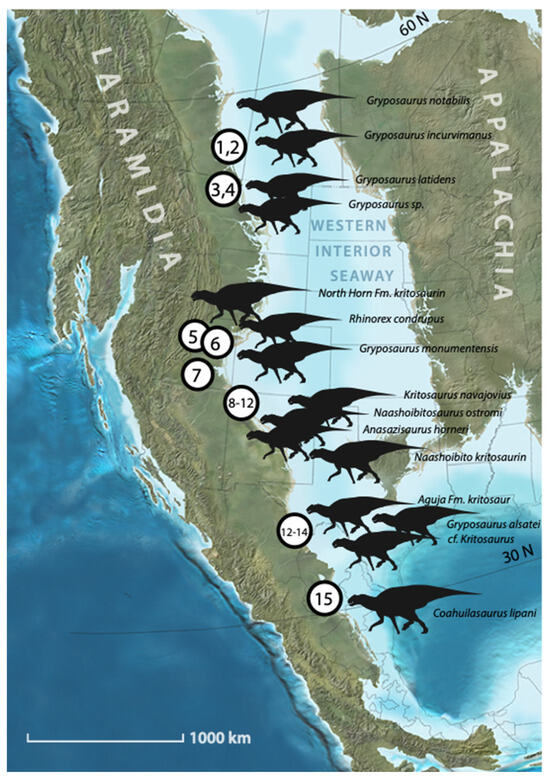
Figure 1.
Map showing localities of kritosaurins from the Late Cretaceous of western North America (Laramidia). 1,2, Gryposaurus notabilis (incl. G. incurvimanus) Dinosaur Park Formation, Alberta; 3, Gryposaurus latidens, Two Medicine Formation, Montana; 4, Gryposaurus sp., Bearpaw Shale, Montana; 5, Kritosaurini indet., North Horn Formation, Utah; 6, Rhinorex condrupus, Neslen Formation, Utah; 7, Gryposaurus monumentensis, Kaiparowits Formation, Utah; 8, Anazisaurus horneri, Farmington Formation of the Kirtland Formation of New Mexico; 9, Naashoibitosaurus ostromi, De-Na-Zin Member of the Kirtland Formation of New Mexico; 10, Kritosaurus navajovius, De-Na-Zin Member of the Kirtland Formation of New Mexico; 11, Naashoibito kritosaurin, Naashobito Member of the Kirtland Formation of New Mexico; 12, Gryposaurus (?) alsatei, upper Javelina Formation, Texas; 13, cf. Kritosaurus., UTEP P.37.7, basal Javelina Formation of Texas; Aguja Formation, Texas; 14, Aguja Fm. Kritosaur, AMNH 3079; 15, Coahuilasaurus lipani, Cerro del Pueblo Formation, Coahuila. Map by Ron Blakey.
A diverse assemblage of dinosaurs occurs in the Cerro del Pueblo Formation (Table 1), including theropods and ornithischians. Theropods include tyrannosaurids, the deinocheirid Paraxenisaurus normalensis [25], ornithomimids [26], caenagnathids [27], dromaeosaurids [28], and troodontids [28,29]. Ornithischians include ankylosaurs, ceratopsids, thescelosaurids, and hadrosaurids. Ankylosaurs include unnamed nodosaurids [30] and ankylosaurids [31]. At least two ceratopsids occur, an unnamed centrosaurine and an unnamed chasmosaurine [5]. Neornithischians include thescelosaurids [32] and both lambeosaurine and saurolophine hadrosaurids. Three lambeosaurines have been named from the Cerro del Pueblo: the lambeosaurin Velafrons coahuilensis [6], the parasaurolophin Tlatolophus galorum [33], and Latirhinus uitstlani [34,35]. Saurolophine hadrosaurids include indeterminate saurolophines [36] and IGM 6685 [19], a partial rostrum collected from Presa San Antonio, near Saltillo (Figure 2). The Presa San Antonio specimen is referable to Kritosaurini, and has previously been referred to Kritosaurus [19].

Table 1.
Dinosaurian fauna of the latest Campanian Cerro del Pueblo Formation, Coahuila, Mexico.
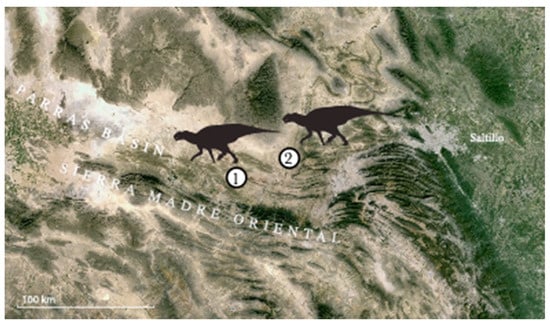
Figure 2.
Map showing (1) the Presa de San Antonio locality that produced IGM 6685, holotype of Coahuilasaurus lipani, and (2) referred specimen, BENC 1/1-007-2, BENC 1/1-007-1, BENC 1/1-007-3 fragmentary skull.
The Presa San Antonio hadrosaur, IGM 6685, is represented by the anterior part of a skull and jaws. It has a complicated taxonomic history. IGM 6685 was initially described in a paper along with a second hadrosaur specimen, PASAC 1 [19]. PASAC 1 is a large saurolophine collected near Sabinas in northern Coahuila [19]. PASAC 1 comes from the lower part (Zone 1) of the Olmos Formation and is represented by a very large and relatively complete skeleton. The specimen includes the maxilla, postorbital, braincase, quadrate, pterygoid, dentary, most of the vertebrae, scapulocoracoid, pelvis, and left hindlimb. IGM 6685 and PASAC 1 were originally interpreted as representing a single hadrosaurid species, identified as Kritosaurus sp. [10,19].
However, the overlapping parts of the dentary of IGM 6685 and PASAC 1 are morphologically distinct, one being short and robust and the other slender and elongated, showing that they belong to different species. Phylogenetic analysis further show that the specimens are not closely related [10,38]. The Presa de San Antonio specimen, IGM 6685, has been confirmed as a kritosaurin [10,38]. The affinities of PASAC 1 are unclear. Prieto-Marquez and Wagner (2013) recovered PASAC 1 in Saurolophini [39]; Prieto-Marquez (2014) placed it outside of a clade formed by Kritosaurini and Saurolophini; a similar result was recovered in 2023 by Prieto-Marquez and Wagner [40]. Ramirez-Velasco (2022) recovered PASAC 1 as part of Edmontosaurini [38]. The affinities of this animal remain uncertain, but it does not appear referable to Kritosaurini.
Following its initial description and referral to Kritosaurus sp., IGM 6685 was later referred to Kritosaurus navajovius by Prieto-Márquez [10]. However, recent study of the specimen suggests that although it represents a kritosaurin, it is distinct from Kritosaurus. Although the limited overlap between the two complicates comparisons, IGM 6685 differs markedly from Kritosaurus in the anatomy of the dentary and predentary. Instead, its anatomy suggests affinities with the kritosaurin Gryposaurus, especially G. monumentensis.
Furthermore, the Presa San Antonio hadrosaur is significantly younger than Kritosaurus. The Presa San Antonio specimen dates to the latest Campanian (Figure 3), ~72.5 Ma [19]; however, the holotype of K. navajovius comes from the De-Na-Zin member of the Kirtland Formation [10], dating to around 1 Ma earlier [24,41]. Given the rapid evolution and extinction seen in Cretaceous dinosaurs [42,43], with few species lasting more than a million years, it seems unlikely that IGM 6685 is referable to the significantly older K. navajovius. In fact, no dinosaurs from the Cerro del Pueblo Formation are referable to taxa known from the Kirtland Formation, even at the generic level (Table 1; Figure 1), suggesting substantial faunal turnover, endemism, or both.
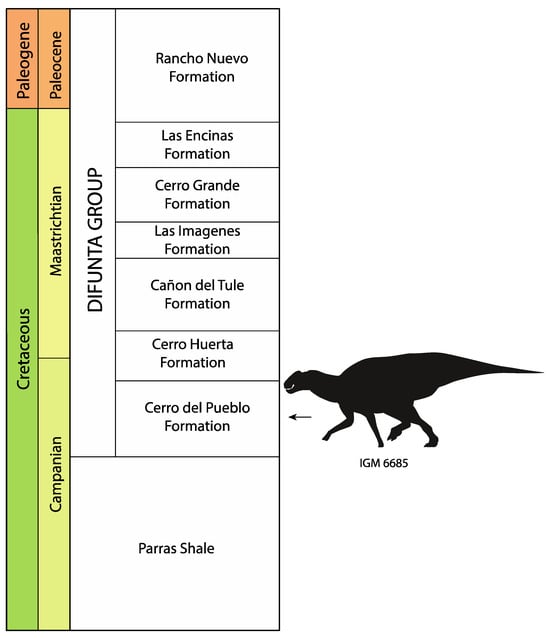
Figure 3.
Stratigraphic column showing stratigraphic position of IGM 6685, holotype of Coahuilasaurus lipani. Stratigraphy after [44].
Here, we redescribe IGM 6685 as a distinct taxon of kritosaurin. A second kritosaur, from Cerro del Pueblo Formation, outcrops at El Palmar, is tentatively referred to this species.
2. Materials and Methods
2.1. Materials Studied
Specimens described here include IGM 6685, the anterior rostrum of a hadrosaur skull from Presa de San Antonio, and a fragmentary skull comprising BENC 1/1-007-2, left maxilla, BENC 1/1-007-1, BENC 1/1-007-3 dentary fragments, from El Palmar, Municipality of General Cepeda. Both specimens come from the Cerro Del Pueblo Formation of the Difunta Group (Figure 3 and Figure 4), with IGM 6685 coming from near the base of the exposed outcrop.
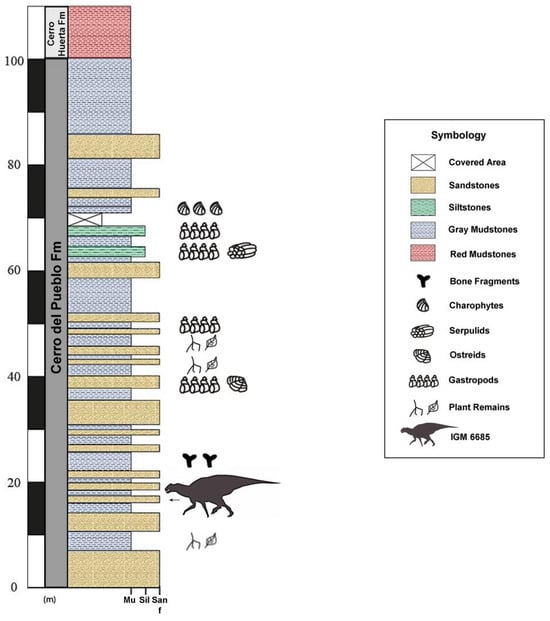
Figure 4.
Stratigraphy of the Cerro Del Pueblo Formation at the Presa de San Antonio locality showing the stratigraphic position of IGM 6685, holotype of Coahuilasaurus lipani.
2.2. Phylogenetic Analysis
Phylogenetic analysis was conducted using a modified version of a recently published data matrix of Ramírez-Velasco [38], constructed using Mesquite [45]. We revised nine codings of C. lipani (IGM 6685) and coded 29 additional characters. We added five new morphological characters (Ch. 335–340) to the matrix. A single biogeographic character was included for biogeographic reconstruction but not included in the final analysis. Four taxa (Malefica, Huallasaurus, Kelumapusaurus, “Gryposaurus” alsatei) were added. Secernosaurus was coded separately taxa from Huallasaurus. Sahaliyania was excluded as it is considered synonymous with Amurosaurus. We maintained 272 characters as ordered as in the original matrix. The data matrix consists of 90 taxa and 341 characters.
Two separate analyses were run, one using equal-weights parsimony and one using implied weighting with k = 2. Analyses were run in PAUP * 4.10 b10 [46] using a heuristic search. Due to missing data and conflicts in the data, equal-weights parsimony produced an extremely large number of trees; the analysis was therefore run until 25,000 trees were produced and a strict consensus was estimated. Implied weights produced a much smaller number of trees, from which the strict consensus was estimated. Both results are shown in the SI. The second analysis is better-resolved and agrees better with stratigraphy and so was used to reconstruct biogeography. Biogeography was reconstructed using parsimony using the “reconstruct” command in PAUP.
3. Results
3.1. Systematic Paleontology
Dinosauria Owen 1842 [47]
Ornithopoda Marsh 1881 [48]
Hadrosauridae Cope 1869 [49]
Saurolophinae Brown 1914 [50,51] (sensu Prieto-Márquez, 2010)
Kritosaurini Brett-Surman 1989 (sensu Prieto-Márquez, 2014)
Coahuilasaurus lipani gen. et sp. nov.
Etymology. Coahuila in reference to Coahuila State + Greek sauros, ‘lizard’; lipani references the Lipani Apache tribe of Coahuila.
Holotype. IGM 6685 (Figure 5), premaxillae, fragments of maxillae, right(?) vomer and pterygoid fragment, predentary, and anterior ends of the dentaries (Figure 6, Figure 7, Figure 8, Figure 9, Figure 10, Figure 11, Figure 12 and Figure 13).
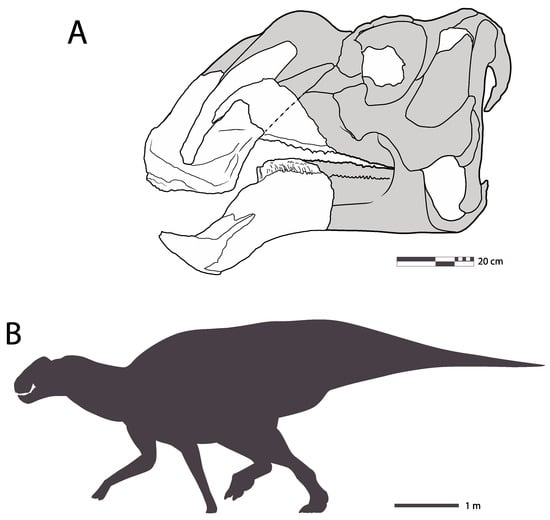
Figure 5.
IGM 6685, holotype Coahuilasaurus lipani. (A), rostrum with posterior skull reconstructed in left lateral view; (B), scaled silhouette. The animal is estimated to have been around 8 m in length.
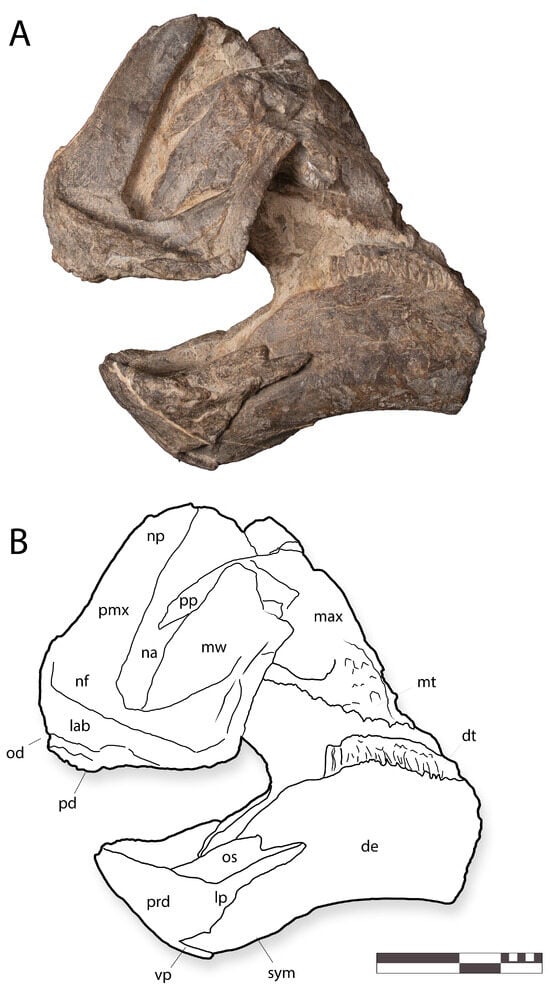
Figure 6.
IGM 6685, holotype of Coahuilasaurus lipani, in (A), left lateral view; (B), line drawings in left lateral view. Abbreviations: de, dentary; dt, dentary teeth; lab, labrum; lp, lateral process; mt, maxillary teeth; mw, maxillary wing; na, naris; nf, narial fossa; np, nasal process; od, oral denticles; os, occlusal shelf; pmx, premaxilla; pd, palatal denticles; pp, palatal process; sym, symphysis; vp, ventral process. Scale = 20 cm.
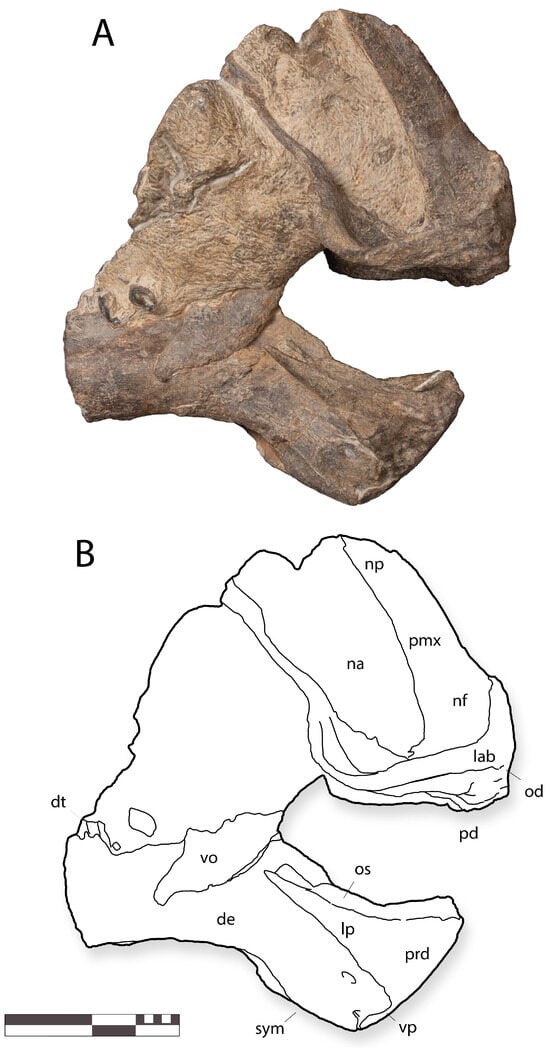
Figure 7.
IGM 6685, holotype of Coahuilasaurus lipani, in (A), right lateral view, (B), line drawings in right lateral view. Abbreviations: de, dentary; lab, labrum; lp, lateral process; na, naris; nf, narial fossa; np, nasal process; od, oral denticles; os, occlusal shelf; pmx, premaxilla; pd, palatal denticles; pp, premaxillary process; sym, symphysis; vo, vomer; vp, ventral process. Scale = 20 cm.
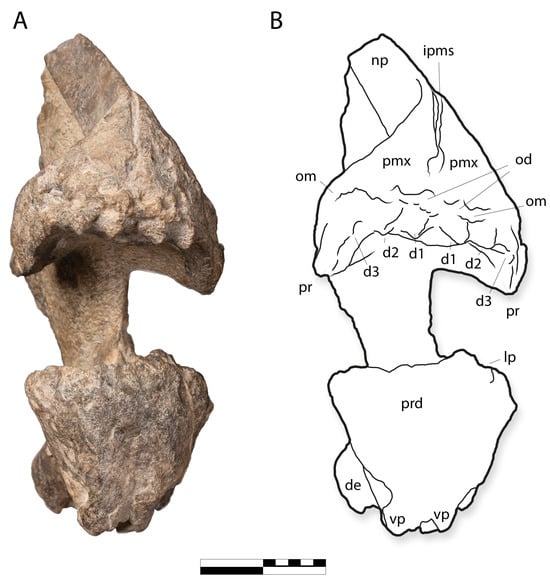
Figure 8.
IGM 6685, holotype of Coahuilasaurus lipani, in (A), anterior view, (B), line drawing in anterior view. d1–d3, palatal denticles 1–3; de, dentary; ipms, interpremaxillary suture; lp, lateral process; od, oral denticles; om, oral margin; np, nasal process; pmx, premaxilla; pr, palatal ridge; prd, predentary; vp, ventral process of predentary. Scale = 20 cm.
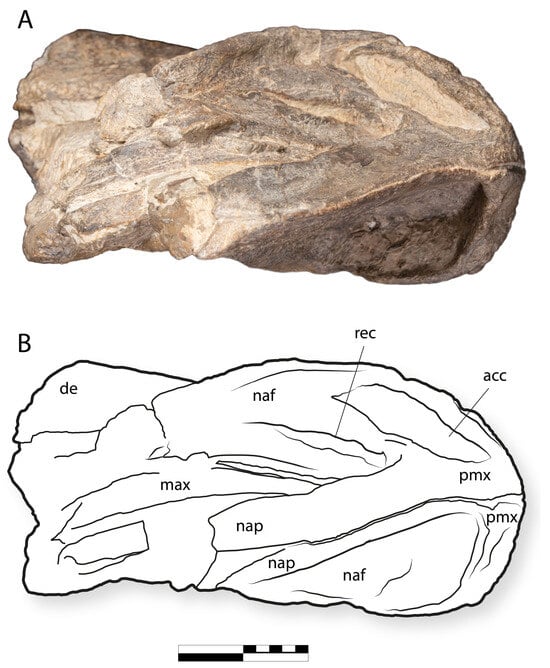
Figure 9.
IGM 6685, holotype of Coahuilasaurus lipani, (A) dorsal view, (B) line drawing in dorsal view. Abbreviations: acc, accessory fossa; de, dentary, max, maxilla; naf, narial fossa; nap, nasal process; pmx, premaxilla, rec, recess. Scale = 20 cm.
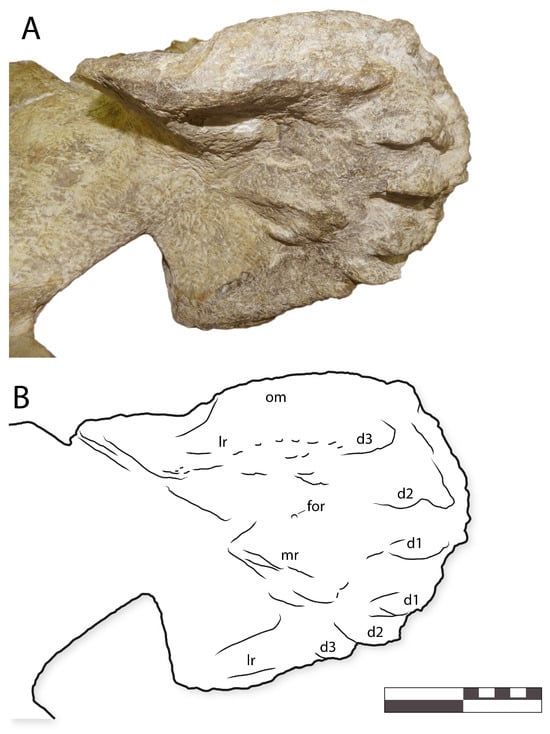
Figure 10.
IGM 6685, holotype of Coahuilasaurus lipani, (A) premaxillae in ventral view, (B) line drawing in ventral view. Abbreviations: d1–d3, palatal denticles 1–3; for, foramen; lr, lateral ridge; mr, median ridge, om, oral margin.
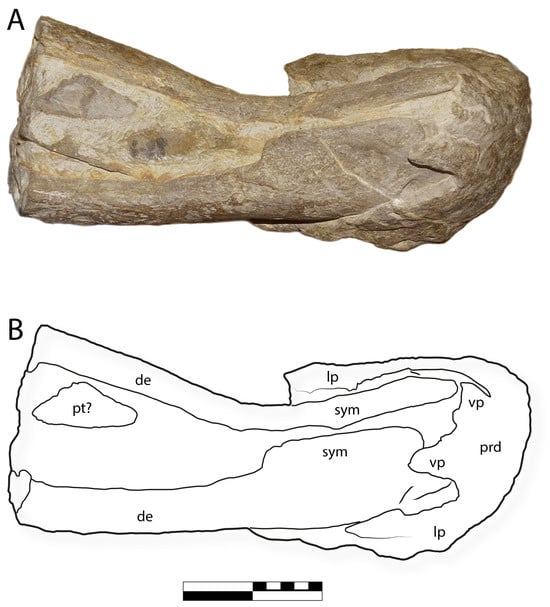
Figure 11.
IGM 6685, holotype of Coahuilasaurus lipani, premaxilla in (A), palatal view, (B), line drawing in palatal view. Abbreviations: de, dentary; lp, lateral process; prd, predentary; pt?, pterygoid; sym, symphysis; vp, ventral process. Scale = 20 cm.
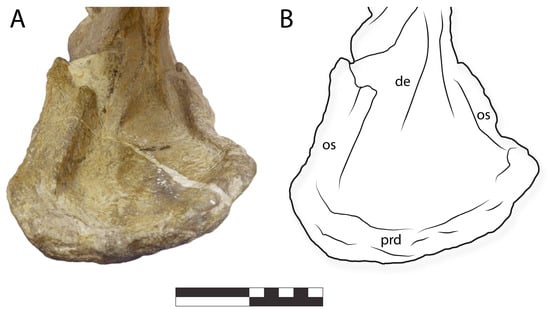
Figure 12.
IGM 6685, holotype of Coahuilasaurus lipani, (A) lower jaw in anterior view, (B), line drawing. Abbreviations: de, dentary; prd, predentary; os, occlusal shelf. Scale = 10 cm.
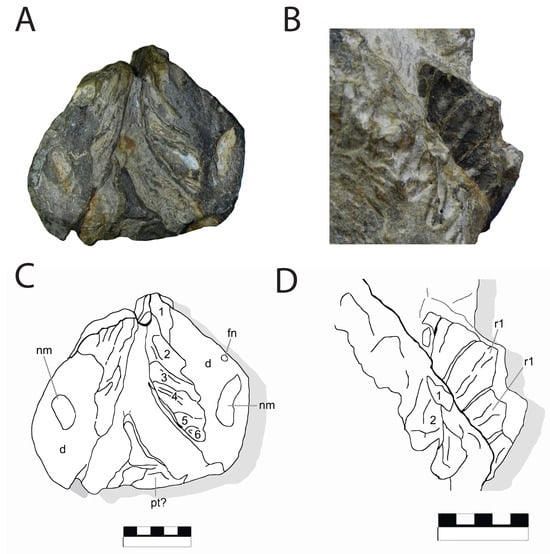
Figure 13.
Holotype of Coahuilasaurus lipani, IGM 6685, (A,C), dentaries in posterior view showing dentition. (B,D), dentition in occlusal view. Abbreviations: fn, nutrient foramina, nm, Meckelian canal, pt?, pterygoid, r, replacement teeth. Scale = 5 cm.
Referred specimens. BENC 1/1-007-2, left maxilla (Figure 14), BENC 1/1-007-1, BENC 1/1-007-3 dentary fragments (Figure 15 and Figure 16). BENC 1/1-007-2 represents a kritosaurin and shows the steeply rising ascending process of the maxilla seen in the Coahuilasaurus lipani type and the related Gryposaurus monumentensis, as well as a large number of replacement teeth in the dental battery. It is tentatively referred to as C. lipani.
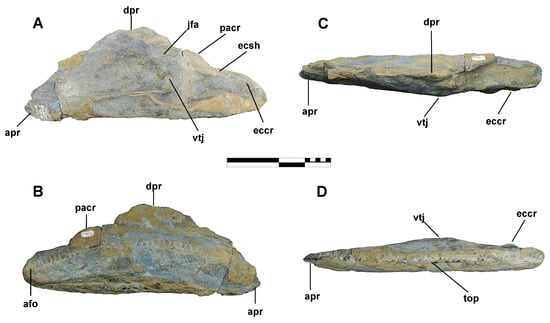
Figure 14.
Maxilla of Coahuilasaurus lipani, referred specimen BENC 1/1-007: Left maxilla BENC 1/1-007-2 in lateral (A), medial (B), dorsal (C), and ventral (D) views. Abbreviations: afo, alveolar foramina; apr, anteroventral process; dpr, dorsal process; eccr, ectopterygoid crest; ecsh, ectopterygoid shelf; jfa, jugal facet; pacr, palatine crest; top, tooth occlusal plane; vtj, ventral tubercle of the jugal. Scale = 20 cm.

Figure 15.
Left and right dentaries of cf. Coahuilasaurus lipani, specimen BENC 1/1-007: Left dentary BENC 1/1-007-1 in lateral (A) and medial (B) views. Right dentary BENC 1/1-007-3 in lateral (C) and medial (D) views. Abbreviations: al, alveoli; deb, dental battery; der, dental ramus; mkg, Meckelian groove. Scale = 20 cm.

Figure 16.
Dentary teeth of cf. Coahuilasaurus lipani BENC 1/1-007. (A), dental battery in medial view. (B), closeup showing tooth crowns. Scale = 5 cm.
Diagnosis. Large kritosaurin with an estimated length of ~8 m (Figure 5). Distinguished by the following unique character combination (* = autapomorphies). The anterior margin of premaxillae is strongly convex in lateral view; the anterodorsal margin of the narial process is subtly sinusoidal in lateral view, being concave anteriorly then weakly convex posteriorly *. The narial aperture is narrow and slot-like and strongly extended anteroventrally *. The premaxillary labrum is deep anteriorly, becoming shallower posteriorly. The posteroventral corner of the premaxillary beak is strongly angular, with the anteroventral margin and posteroventral margin forming approximately a right angle in lateral view *; the posteroventral margin of the premaxilla is strongly concave in lateral view. The premaxilla is short and broad in dorsal view. The oral margin of the beak bears a series of 5–6 small denticles. The palatal surface of the premaxilla forms a horseshoe-shaped ridge projecting far below the oral margin *; the ridge bears three pairs of massive, diamond-shaped denticles*. The nasal–premaxillary contact is posteriorly positioned, probably near the end of the nasal aperture. Very large narial flange of premaxilla *. The predentary oral margin is strongly concave in the lateral view and has a steeply inclined rostroventral margin, and the tip is hooked up strongly in lateral view (approached in Gryposaurus monumentensis). There is a narrow U-shaped notch between predentary lateral processes and ventral processes (shared with Gryposaurus monumentensis); the ventral processes are long and slender. The edentulous end of the dentary is strongly downturned, with a strongly convex anterodorsal margin, and a long ventral contact between the dentaries.
Locality and Horizon. IGM 6685 comes from Presa San Antonio (Figure 2) in the Parras Basin [19], near the SPA-88-9 quarry (Latirhinus type locality); 25°27′33″ N and 101°42′18″ W (Serrano-Brañas, 2006; pers. com. Hernández-Rivera). The outcrops here are part of the Cerro del Pueblo Formation (Figure 4). Invertebrate fossils suggest a mid-late Campanian age for the Cerro del Pueblo Formation [21], while magnetostratigraphy suggests the beds correlate with magnetochron 32n.3r—32n.2n, suggesting an age of ≥72.5 Ma [21].
Three specimens, found in association, BENC 1/1-007-2, a left maxilla (Figure 14), and BENC 1/1-007-1 and BENC 1/1-007-3, dentary fragments (Figure 15 and Figure 16), come from El Palmar, located in the Municipality of General Cepeda, at 25°34′18″ N and 101°20′09″ W. This site lies in the middle of the Cerro del Pueblo Formation.
Description. IGM 6685 (Figure 5, Figure 6, Figure 7, Figure 8, Figure 9, Figure 10, Figure 11, Figure 12 and Figure 13) comprises the premaxillae, left and right maxillae, vomer, predentary, and the tips of the left and right dentaries, with associated maxillary and dentary tooth batteries. The bones are articulated and preserved approximately in life position in a sandstone matrix with shells of gastropods and bivalves. Some deformation has occurred such that the bones are not entirely symmetrical between the left and right sides.
Premaxillae. The left and right premaxillae (Figure 6, Figure 7, Figure 8, Figure 9, Figure 10 and Figure 12) are well-preserved, with only the tips of the nasal processes and maxillary processes missing. The premaxillae would have contacted the nasals dorsally and the maxillae posteriorly (Figure 5).
As in other hadrosaurids, the premaxillae form an edentulous beak. The beak is transversely broad (Figure 9 and Figure 10), with the palatal surface being about as wide as long anteroposteriorly. The anterior margin of the premaxilla is broadly U-shaped in dorsal view (Figure 9).
In anterior view (Figure 8), the oral margin of the premaxilla is slightly convex. This corresponds to the dorsal margin of the predentary, which is concave in anterior view in contrast to the convex margin in Gryposaurus spp. [52,53]. The beak’s oral margin is damaged, but thickenings along the anterior surface suggest a series of 5–6 small denticles (Figure 8 and Figure 10). The enlarged oral denticles seen in G. alsatei [54] and the austrokritosaur Huallasaurus [55] are absent.
In lateral view (Figure 6 and Figure 7), the premaxilla’s anterior margin is vertically oriented and slightly convex, similar to G. monumentensis [52]. The premaxillary labrum, seen in lateral view, is deep anteriorly and shallower posteriorly, as in Gryposaurus [52,53] and “G”. alsatei [54]. Posteriorly, the labrum becomes very narrow as it extends toward the ventrolateral edge of the beak. This section of the labrum extends posterolaterally toward the ventrolateral apex of the beak, with a thin “u”-section and then a thick, lenticular section that later disappears, as seen in Gryposaurus notabilis [53] and G. monumentensis [52]. This lenticular zone forms a crest that delimits the lateral concavity.
The ventral margin of the beak is relatively straight in lateral view, then turns up posteriorly (Figure 6 and Figure 7), as in other Gryposaurus [52,53]. The premaxilla’s oral margin meets the maxillary process at a right angle to form a blunt apex, in contrast to the more rounded margin in Gryposaurus spp. [52,53] and other saurolophines such as Edmontosaurus [56,57]. However, a similar condition occurs in “Gryposaurus” alsatei and Brachylophosaurus canadensis [58].
In the ventral view, the palatal surface bears a broad, horseshoe-shaped ridge (Figure 10). The ridge bears a narrow but blunt triturating edge. This ridge lies slightly below the level of the beak’s oral margin laterally, as in “G”. alsatei [54], but anteriorly, the ridge projects far below the premaxilla’s oral margin so that the ridge is exposed in lateral view. This morphology is unique to IGM 6685 among kritosaurins.
Anteriorly, the palatal ridge bears six large denticles, three per side (Figure 8 and Figure 10). The palatal ridge in Gryposaurus latidens has at least four and perhaps five pairs of denticles [54]; “Gryposaurus” alsatei bears at least seven and perhaps as many as nine denticles per side [59]; the condition in other kritosaurins has not been described. The palatal denticles are very large and robust, unlike the small palatal denticles in “G”. alsatei [54] and G. latidens [59], and project ventrally. Similar to the condition in “G”. alsatei and G. latidens, palatal denticles anteroposteriorly elongate; they are triangular in anterior view and more crescentic in the lateral view.
The strong ventral projection of the premaxillae and large toothlike ventral structures are unique among hadrosaurids. Remarkably, they recall the modified, downwardly projecting palate of oviraptorosaurian theropods [60,61]. The conical denticles of the palatal surface would have occluded with the main body of the predentary anteriorly. Posterolateral to this, the palatal ridge would have occluded with the shelflike lateral processes of the predentary. This feature is presumably associated with a specialized diet and/or mechanism of food processing and presumably was used to crop some kind of tough or fibrous vegetation, such as palms.
The large size of the palatal denticles and the reduced oral denticles have led to misinterpretations of the anatomy of the animal, with the denticles of the palatal surface being mistaken for the denticles of the oral margin of the premaxilla [54,62]. The large conical denticles of Coahuilasaurus do not correspond to the large oral denticles of “G”. alsatei or other hadrosaurids, however. In “Gryposaurus” alsatei it is the oral denticles that are enlarged; in C. lipani, oral denticles are reduced, and the palatal denticles are instead hypertrophied.
In the center of the palatal surface, posterior to the denticles, lies a second, low, crescentic ridge, and behind this, a pair of oval premaxillary foramina (Figure 10).
The premaxilla’s nasal process is elongated (Figure 6 and Figure 7), as in other hadrosaurids [63], but more robust, as in Gryposaurus monumentensis [52] and G. notabilis [53], and unlike the more gracile narial process in G. latidens [59]. In cross-section, the narial process has a distinctly triangular shape, as in other kritosaurins such as G. notabilis. The lateral surface is flat, similar to G. notabilis, but unlike the concave condition in G. monumentensis [52]. The dorsal margin of the narial process has a relatively straight shape in lateral view, being weakly concave anteriorly and weakly convex posteriorly such that there is a slightly sinuous shape to the dorsal margin. This shape is unique among kritosaurins. No articulation for the nasals is preserved, suggesting that the nasals had a limited contribution to the nasal aperture.
The maxillary process is broad, thin, and concave where it forms the posterior narial fossa (Figure 6). The dorsal margin is sinuous in shape due to the presence of the anterior margin of the narial process (=dorsolateral flange of Prieto-Márquez, 2008), which projects laterodorsally. Together, the nasal process and the maxillary process delimit the naris. The naris is narrow compared to other kritosaurins [52,53]. This condition is approached in Gryposaurus TMP 1991.081.0001 [53].
A deep narial fossa surrounds the nasal aperture (Figure 6 and Figure 7). The narial fossa is broadly expanded anteriorly, dorsally, and ventrally to cover almost the entire lateroventral surface of the premaxilla, similar to Gryposaurus spp. [52,53], Edmontosaurus [56,57], Saurolophus [64], and Brachylophosaurus [58]. Ventrally, the narial fossa is divided into distinct anterior and posterior fossae, as in other saurolophines, with a strong rim and a lateral concavity separating them.
Maxillae. The anterior end of the left maxilla is preserved (Figure 6); a fragment of the right maxilla is buried in the matrix. The maxillary body is robust and, based on the position of the palatal process, appears to have been taller than in other saurolophines, a condition typical of kritosaurins [52,53], which is particularly well developed in Gryposaurus monumentensis.
The maxilla’s anterior section has two processes: the palatal process and the anterior process. The long and subrectangular palatal process is exposed through the nasal aperture, as in G. monumentensis [52]. The anterior process is short, with a strong ventral sloping. The left maxilla contacts the premaxilla at an angle of almost 90 degrees, but the maxilla’s occlusal margin forms an angle of around 45 degrees with the premaxilla in G. monumentensis, suggesting that the maxilla has been displaced from its natural position.
Dentaries. The dentary of G. lipani is extremely deep, more so than in Kritosaurus navajovius [10] or Gryposaurus spp. [52,53]. The dentaries have a strongly downturned symphysis as in ?Gryposaurus alsatei [54], Gryposaurus monumentensis [52], G. notabilis [53], Kritosaurus navajovius, and Austrokritosauria such as Secernosaurus koerneri [62]. In young G. notabilis [53] and G. latidens [59], the dentary symphysis is more weakly downturned, as is the symphysis in Saurolophus [65,66] and edmontosaurins [56,57].
The dentary’s edentulous anterodorsal margin (Figure 6 and Figure 7) is relatively short and dorsally convex, as in Gryposaurus monumentensis [52] and in contrast to Kritosaurus [10], where it is straight. The dentary’s lateral surface is flat dorsally and bears a ridge ventrally. The dentary has a convex lateral surface in “G”. alsatei [54] and Austrokritosauria [55]. The dentary tip bears a narrow, triangular process inserting into a V-shaped slot formed by the main body of the predentary and the predentary ventral process. This narrow, V-shaped contact is shared with G. monumentensis [52]. The anterior process is much broader in G. notabilis [67] and K. navajovius [10]. In the lateral view, a neurovascular canal lies near the ventral process of the predentary, and scattered neurovascular foramina lie below the dental battery.
Predentary. As in other hadrosaurids [63], the dentaries articulate with a large predentary bone anteriorly (Figure 6, Figure 7, Figure 8, Figure 11 and Figure 12), which, in life, bore a keratinous rhamphotheca. The predentary is triangular in lateral view. The predentary’s anterior margin rises up steeply relative to the long axis of the dentary, making almost a right angle relative to the long axis of the dentary symphysis. This steeply projecting symphysis is shared with G. monumentensis [52] but contrasts with the shallower angle formed by the predentary in other Gryposaurus species [53] and K. navajovius [10], where the tip projects more anteriorly.
The predentary’s oral margin is concave in anterior view (Figure 6), corresponding to the convex oral margin of the premaxillae. Laterally, the oral margin becomes deeply concave where the predentary would have contacted the caudolateral corners of the premaxillae.
The predentary lateral processes are short and broad, becoming wider posteriorly. They have a concave occlusal surface. Their lateral margin is sharp, their dorsal surface bears a trough, and medially, the trough is delimited by an elongate, convex, bladelike surface. The lateral processes are smaller than in G. monumentensis, covering less than half the dentary in lateral view, a condition shared with K. navajovius [10].
Ventrally, the predentary bears a short, slender ventral process (Figure 8 and Figure 11). This process has a short central lamina, with the ventral margin being concave, and a shallow concave external surface. Laterally, the central lamina bears a pair of finger-like structures with rounded tips, as in other kritosaurins [10].
Vomer. The anterior end of the right vomer is preserved and has been displaced to lie along the right dentary (Figure 11). The bone is very thin, flat, and triangular in shape, with a curved tip. The vomer’s posterior margin bears an excavation, as in Gryposaurus monumentensis [52] and Rhinorex condrupus [68].
Pterygoid. A bone that appears to represent the pterygoid is exposed posteroventrally between the dentaries (Figure 11). The dorsal part of the pterygoid is visible in posterior view; the ventral section is perpendicular to it in ventral view. Due to poor preservation, no other morphology of the pterygoid can be described; it cannot be identified as either the left or right pterygoid.
Dentition. The right maxilla preserves 20 alveoli (Figure 6). This suggests a high tooth count, as in Gryposaurus monumentensis, which has ~50 maxillary teeth [52]. The teeth form a stacked series of tooth families in each alveolar slot, with two functional teeth in each tooth family contributing to the grinding surface of the dental battery, as in other hadrosaurids [69].
The dentary preserves 15 alveoli and likely had around 40 teeth, as in G. monumentensis [52]. There are five or six stacked teeth in each tooth family (Figure 13) versus four or five in G. monumentensis [52]. In the middle of the toothrow, there are three functional teeth per tooth family, as in other kritosaurins [70]. Dentary teeth have a straight primary ridge and no accessory ridges, as in Gryposaurus spp. [70] and Rhinorex [68]. Denticles are highly reduced, as in other kritosaurins [10], with the exception of “G.” alsatei [54].
Referred Specimens. BENC 1/1-007 is represented by an almost complete left maxilla (BENC 1/1-007-2) (Figure 14) and partial left (BENC 1/1-007-1) and right dentaries (BENC 1/1-007-3) (Figure 15 and Figure 16).
Maxilla. As preserved, the maxilla BENC 1/1-007-2 is 470 mm long and 170 mm tall. In lateral view, the bone is massive and has a triangular outline (Figure 14A,B); dorsally, it has a straight appearance (Figure 14C). The bone’s anterior region is partially broken, and only part of the anteroventral process is preserved (Figure 14A,B). The premaxillary shelf of this process is flattened and not prominent. The crowns and roots of five teeth can be seen in this area (Figure 14A).
The middle of the maxilla has a tall and oval dorsal process, which is 100 mm long and 50 mm wide (Figure 14A). Below the dorsal process lies a prominent jugal facet. In addition, there are three large maxillary foramina in the middle region, each about 20 mm in diameter. The ventral tubercle of the jugal facet is moderately well-developed. The posterior region is dorsoventrally narrow and bears the contact surfaces for the palatine, pterygoid, and ectopterygoid bones. The palatine crest is long and broad, measuring 100 mm in length, although it is partially eroded and remains somewhat covered with matrix (Figure 14A).
The ectopterygoid crest is large and well-developed, measuring 150 mm long and 42 mm thick; this crest extends to the posterior end of the maxilla, where it curves downwards (Figure 14A). The ectopterygoid shelf is 45.5 mm wide, possesses a shallow concavity, and has a slight elevation that joins the palatal crest (Figure 14A,C). The lateroventral margin of the maxilla is nearly straight (Figure 14A,D). An arched line of 37 dental foramina can be seen on the medial side of the maxilla (Figure 14B). Ventrally, the maxilla has 42 tooth positions forming a straight tooth row (Figure 14D). The occlusal surface has a maximum of two functional teeth.
Dentary. Specimens BENC 1/1-007-1 and BENC 1/1-007-3 are fragmented and correspond to the middle of the dentaries. Both bones are robust and massive. The left dentary, BENC 1/1-007-1, is better preserved, measuring 350 mm long and 210 mm tall (Figure 15A,B); the right dentary, BENC 1/1-007-3, is 340 mm long and 170 mm tall and remains mostly covered by the rock matrix (Figure 15C,D). In medial view, both elements are deep below the dental battery, and the Meckelian groove can be observed (Figure 15B,D). Laterally, BENC 1/1-007-1 has a flat mandibular ramus and five foramina, whereas BENC 1/1-007-3 shows a slightly convex mandibular ramus (Figure 15A,C).
The dental battery is exposed and well-preserved in BENC 1/1-007-1 (Figure 15A,B and Figure 16); in BENC 1/1-007-3, it is partially fragmented and covered by matrix (Figure 15D). In both dentaries, there would be over 30 tooth positions. There are at least five teeth per alveolus, possibly more, but the ventral part of the toothrow is covered by matrix. At least two, and typically three, teeth are functional and contribute to the grinding surface in each alveolus. Alveoli measure 0.6 cm wide and 7.4 cm long on average.
Tooth crowns are diamond-shaped. Crowns average 4.6 cm tall and 1.3 cm wide. The height/width ratio of the crown is between 3.1 and 3.8, depending on the tooth position within the dental battery; the crown height/width ratio is 2.9 for Kritosaurus navajovius [10]. Crowns have a low, straight primary ridge and lack secondary ridges. Denticles are highly reduced, as in the holotype.
3.2. Phylogeny
Phylogenetic analysis using implied weighting (Figure 17) produced 171 most-parsimonious trees (tree length = 1448, Consistency index = 0.3363, Retention index = 0.7599, Rescaled consistency index (RC) = 0.2556). Phylogenetic analyses using equal weighting produce a large number of trees (tree length = 1429, Consistency index = 0.3408, Retention index = 0.7647, Rescaled consistency index = 0.2606).
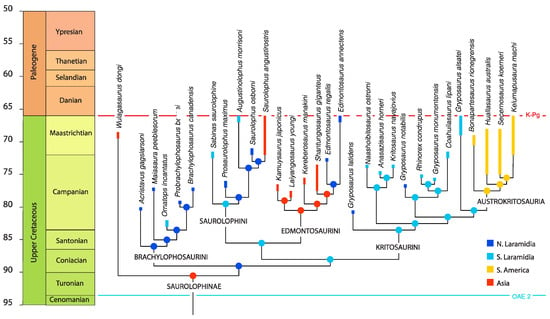
Figure 17.
Evolutionary tree showing the relationships of Coahuilasaurus lipani within Kritosaurini. Strict consensus of 171 most-parsimonious trees.
Both analyses recover three major subclades. The first comprises Coahuilasaurus lipani, Gryposaurus utahensis, Rhinorex condrupus, and Gryposaurus notabilis. The second consists of Naashoibitosaurus ostromi, Anasazisaurus horneri, and Kritosaurus navajovius. The third comprises the South American kritosaurins, the Austrokritosauria, as well as “Gryposaurus” alsatei. Implied weighting recovers Austrokritosauria as part of Kritosaurini, as found previously; the placement of Austrokritosauria is unresolved using equal weighting, and they do not emerge as part of Kritosaurini in all trees. In all analyses, under either implied weighting or equal weights parsimony, Coahuilasaurus is found to be sister to a lineage comprising Gryposaurus monumentensis and Rhinorex condrupus, with Gryposaurus notabilis lying just outside.
The choice of weighting scheme in phylogeny is debated. Although some have argued that implied weighting can recover incorrect trees [71], other analyses suggest it tends to increase accuracy over equal-weights parsimony [72], and in particular trees tend to converge towards the topology produced by implied weighting with subsequent revisions and new data [73]. In the two analyses done here, the implied weighting topology tends to push older taxa further down the tree and younger taxa further up, improving stratigraphic fit. More fossils and data are required to better resolve kritosaur phylogeny.
4. Discussion
4.1. Systematics and Diversity
IGM 6685 has previously been referred to Kritosaurus [10], but phylogenetic analysis shows that it is instead related to Gryposaurus monumentensis. IGM 6685 shares derived features with various kritosaurin species but, in particular, it resembles Gryposaurus monumentensis. The caudoventral margin of the premaxilla, between the ventral margin of the beak and maxilla, is concave in both G. lipani and G. monumentensis. The premaxilla is short anteroposteriorly in both taxa versus the more longirostrine G. notabilis. The short, strongly downturned symphyseal region of the dentary closely resembles Gryposaurus monumentensis [52], G. notabilis [53], and Kritosaurus navajovius [10]. G. latidens [59] has a longer, more weakly downturned symphyseal region; that of G. alsatei is strongly downcurved but elongated [54]. The strongly upturned tip of the predentary is shared exclusively with G. monumentensis among kritosaurins for which the predentary is known. Similarly, the narrow V-shaped dentary–predentary contact is uniquely shared with G. monumentensis. Unfortunately, Rhinorex condrupus lacks much of the premaxillae, the dentaries and the predentary, making comparisons difficult. However, Rhinorex uniquely resembles G. monumentensis in having a premaxilla that is strongly convex on its anterodorsal margin and which hangs downwards giving the snout a ‘droopy’ appearance. This character implies that G. monumentensis and Rhinorex may form a clade, as recovered here.
IGM 6685 is not closely related to Kritosaurus navajovius. Instead, Kritosaurus navajovius, Kritosaurus (Anasazisaurus) horneri, and Naashoibitosaurus ostromi form a distinct clade within Kritosaurini. They are united by the structure of the nasal ornament. As in other kritosaurins, the nasals are dorsoventrally expanded and united by a broad suture to form a crest on the snout; the dorsal margin of this crest bears a pair of low, blunt ridges. In Kritosaurus navajovius, Kritosaurus (Anasazisaurus) horneri, and, to a lesser degree, in Naashoibitosaurus ostromi, this crest is posteriorly extended, and the apex forms in a sort of tubercle or hornlet (Figure 18), which is located far behind the naris. The crest is more posteriorly extended in Kritosaurus navajovius and Kritosaurus (Anasazisaurus) horneri than in Naashoibitosaurus, although the Naashoibitosaurus holotype appears to be a juvenile, and so it seems possible the crest was better-developed in the adult. The nasal crest has a more primitive morphology in Gryposaurus notabilis, with a simple convex dorsal margin, and the apex is located just behind the external naris. The apex of the nasal crest is more anterior in Rhinorex condrupus. In G. latidens, the nasal ornament is still more primitive, being very low with limited expansion behind the naris. The more primitive structure of the nasal ornament suggests that G. latidens lies further down the tree, near the base of the kritosaurin radiation (Figure 17). A feature associated with the development of the nasal crest is the narial fossa. It is extensive and covers the entire crest in Kritosaurus navajovius and Kritosaurus (Anasazisaurus) horneri. It is less posteriorly extended in Naashoibitosaurus and Gryposaurus notabilis, but the fossa extends dorsally above the naris to the top of the crest. In G. latidens, the narial fossa shows limited development dorsal to the naris, a primitive condition. The condition is unclear in G. monumentensis and Rhinorex.
Despite the similarities of Coahuilasaurus to various species of Gryposaurus, the remarkably specialized morphology of the rostrum, with a unique downward projection of the palate, the hypertrophied palatal denticles, and the modified shape of the naris, justify recognition of a distinct genus. The morphological differences between Coahuilasaurus and other kritosaurins are considerable and at least as large as, for example, the differences in rostrum morphology between Triceratops and Torosaurus [74]. Furthermore, it is unclear that species previously assigned to Gryposaurus do, in fact, form a clade. Instead, the genus as traditionally conceived may be a wastebasket for different kritosaurs. It seems likely that a number of species, including ?Gryposaurus alsatei and G. latidens may require separate genera.
Kritosaurins are one of the more diverse lineages of hadrosaur in North America, being represented by a plethora of species from the latest Cretaceous of the Western Interior (Figure 18).

Figure 18.
A panoply of kritosaurs is known from the late Cretaceous of the Western Interior. Coahuilasaurus lipani IGM 6685, Gryposaurus monumentensis RAM 6786 [52], Gryposaurus notabilis ROM 873 [53]; Gryposaurus latidens MOR 478 [59], Rhinorex condrupus BYU 13528 [68], “Gryposaurus” alsatei TMM 46033-1 [54], Kritosaurus navajovius AMNH 5799 [62], Anasazisaurus horneri BYU 12950 [75].
Figure 18.
A panoply of kritosaurs is known from the late Cretaceous of the Western Interior. Coahuilasaurus lipani IGM 6685, Gryposaurus monumentensis RAM 6786 [52], Gryposaurus notabilis ROM 873 [53]; Gryposaurus latidens MOR 478 [59], Rhinorex condrupus BYU 13528 [68], “Gryposaurus” alsatei TMM 46033-1 [54], Kritosaurus navajovius AMNH 5799 [62], Anasazisaurus horneri BYU 12950 [75].

The striking variation in jaw morphology suggests specialization for distinct ecological niches and foods. The variation in cranial crest morphology in Kritosaurini also suggests that sexual selection drove the evolution of kritosaurins, as in the crested lambeosaurines and ceratopsids [76]. Although the bony crests of kritosaurins are not as elaborate, kritosaurins may have had elaborate soft-tissue structures. The related saurolophine Edmontosaurus, which lacks bony ornament, had a soft-tissue crest [77]; furthermore, the elaboration of the nasal crest in kritosaurs is associated with the elaboration of the narial fossae, suggesting some kind of soft tissue structure. Cranial ornament in birds is typically associated with heavily cornified (e.g., Casuarius, Fratercula, Numida, Bucerotidae) or fleshy structures (e.g., Cygnus olor) [78]. In birds, these display structures are sometimes boldly colored [78] or often are associated with bright colors and fleshy structures elsewhere on the face (e.g., Casuarius, Numida). The function of the kritosaurin crest is unclear. It was conspicuous and so presumably functioned for display; in modern birds, cranial ornaments are occasionally used in intraspecific combat as well, for example, in hornbills [79], and the relatively robust crest could conceivably have performed a similar function. The elaborate nasal chamber could also have been involved in vocalization, as proposed for lambeosaurs [80,81].
4.2. The Sabinas Hadrosaur
Kirkland et al. [19] identified the Cerro del Pueblo kritosaurin, IGM 6685, as belonging to the same taxon as PASAC 1, a large hadrosaurid skeleton from Sabinas, but this referral was later rejected.
First, the two hadrosaurs come from different basins and different formations. The Presa San Antonio specimen, IGM 6685, comes from the Cerro del Pueblo Formation in the Parras Basin. The Sabinas skeleton, PASAC 1, comes from the Olmos Formation in the Sabinas Basin, ~300 km away [19]. The formations are similar in age, but radiometric dating of the beds has not been undertaken, so the precise age of either formation remains unclear. Although there is good biostratigraphic data for the Cerro del Pueblo Formation, the particular age of the Olmos Formation is not well-constrained. Given the distance between the two localities and, more importantly, the lack of precise ages for either bed, it remains unclear whether the two animals overlapped in time. Even assuming they did, hadrosaurs were diverse, and it cannot be assumed that two saurolophines from the same horizon represent the same species.
Second (and more importantly), the overlapping bones of the two animals do not match. The dentary of the Sabinas hadrosaur has a long, slender, and weakly downturned symphysis in lateral view [19], similar to Edmontosaurus [57] and Saurolophus [65]. IGM 6685 has a short, deep, and strongly downturned symphysis, as in Gryposaurus [52,53] and Kritosaurus [10].
Third, phylogenetic analysis suggests the hadrosaurs come from different parts of the hadrosaurid tree. IGM 6685 represents a kritosaurin [10,19,38], whereas the Sabinas hadrosaur is variously recovered as an edmontosaurin [38] or a saurolophin [39]. The Cerro del Pueblo hadrosaurid, IGM 6685, therefore, is distinct from the Sabinas hadrosaurid.
The phylogenetic position of the “Sabinasaur” remains unclear. Under implied weighting, the Sabinas hadrosaur is nested in the Saurolophini, contrary to recent analyses that recovered it within Edmontosaurini [38] but consistent with the results of other studies [39]. However, in the equal-weights analysis, its phylogenetic position is unresolved with respect to Saurolophini and Edmontosaurini. Further study is needed.
4.3. Biogeography
Coahuilasaurus lipani differs from kritosaurins found further north in Laramidia [53,59,67] but may be related to G. monumentensis [52], suggesting it is part of a distinct, southern kritosaurin lineage. Other endemic taxa found in Coahuila include the lambeosaurs Velafrons coahuilensis [6], Latirhinus uitstlani [37], and Tlatolophus galorum [33], chasmosaurine Coahuilaceratops magnacuerna [5], and the deinocheirid Paraxenisaurus normalensis [25]. The distinctive nature of the Mexican dinosaur fauna, and southern dinosaurs in general [2,3,4,8,82,83,84], underscores the high endemism seen in the Late Cretaceous of Laramidia.
Furthermore, the persistence of kritosaurins in the south during the latest Campanian fits with a broader pattern where large herbivore lineages disappear from the northern Great Plains while persisting in the south. During the early Campanian, the kritosaur Gryposaurus latidens occurs in the Two Medicine Formation in Montana [59], and in the mid-Campanian, G. notabilis occurs in the Dinosaur Park Formation [53]. Kritosaurs subsequently disappeared from the northern Great Plains. They are not found in the latest Campanian or early Maastrichtian of the Horseshoe Canyon Formation of Alberta [23] or in the late Maastrichtian-aged Scollard, Frenchman, Hell Creek, or Lance formations of the northern Great Plains [1,85]. Meanwhile, C. lipani persists in Mexico in the latest Campanian, and “G.” alsatei in the late Maastrichtian of the Javelina Formation of Texas [54]. A fragmentary skull, AMNH 5797, shows kritosaurs persisted into the latest Maastrichtian in the Naashoibito Member of the Kirtland Formation in New Mexico [62], and a dentary from the latest Maastrichtian of the North Horn Formation (BYU 9432) shows their persistence in Utah.
Similarly, Parasaurolophini, represented by Parasaurolophus walkeri [86] in the Dinosaur Park Formation, disappeared from Canada in the latest Campanian [23]. Meanwhile, Tlatolophus shows that parasaurolophins survived until at least the end of the Campanian in Mexico [33]. Furthermore, Lambeosaurins persisted into the early Maastrichtian in Canada, represented by Hypacrosaurus altispinus [23,87], before vanishing in the late Maastrichtian from Canada [1,23] and the northern Great Plains. However, lambeosaurins survived into the latest Maastrichtian in New Mexico [88]. Finally, Deinocheiridae is present in the late Campanian of Alberta [89] but disappears afterward [23] while persisting into the latest Campanian in Mexico [25].
Herbivorous dinosaur taxa that became extinct in the northern Great Plains survived later or even up to the K-Pg boundary in the Southwest and Mexico. This contributes to the relatively high diversity seen in the Cerro Del Pueblo Formation ornithischians [5,6,19,33,37] compared to the contemporaneous fauna in the lower Horseshoe Canyon Formation [23]. Another striking pattern is that several major lineages, including both Triceratopsini [3,90] and Tyrannosaurini [90,91], appear to originate in the south and later spread north [8]. This “Out-of-the-Tropics” pattern has also been noted for marine invertebrates [92] so it would be unsurprising if dinosaurs behaved in a similar way. The southern parts of Laramidia may have been both a refugium for clades that became extinct in the north and a source of evolutionary novelty.
Evolution may proceed differently in different parts of the continent, with extinction rates, origination rates, and dispersal patterns differing regionally, especially with respect to latitude. It is also possible that cooling in the Campanian-Maastrichtian [93,94] was more severe in the Northern Great Plains, given that climate change tends to be more severe at higher latitudes. And it may be that the relatively low dinosaur diversity seen in the Northern Great Plains [1,85] was a localized phenomenon caused by local changes in climate, precipitation, and vegetation and not indicative of broader regional or global trends. However, the Hell Creek and other Northern Great Plains faunas are extremely well-studied [1,85], whereas other faunas—especially in southern North America—are less well-known [95,96]. This may distort our picture of dinosaur diversity. An apparent decline in dinosaur diversity in the Maastrichtian [97,98] may be, at least in part, an artifact of limited geographic sampling. Therefore, the distinct dinosaur faunas seen in the latest Campanian of Mexico (Figure 19) raise questions about the extent to which diversity patterns seen in the northern Great Plains [1,85] can be taken as representing global or even regional patterns.

Figure 19.
Cerro del Pueblo Formation dinosaur fauna.
5. Conclusions
A partial skull from the latest Campanian of the Cerro del Pueblo Formation of Coahuila, Mexico, represents a new species of kritosaurin, Coahuilasaurus lipani (Figure 20). Along with specimens from the Naashoibito Member of the Kirtland Formation in New Mexico, C. lipani documents the persistence of kritosaurins in the south after local extinction in northern Laramidia, a pattern also seen in Parasaurolophini and Lambeosaurini.

Figure 20.
Life reconstruction of Coahuilasaurus lipani, by C. Díaz Frías, 2023.
Southern faunas not only have distinct species, but different patterns of diversity change, compared to northern Laramidian faunas. Differences in species composition and community structure in different parts of Laramidia mean that further work on the dinosaurs of Mexico and the American Southwest is needed to understand the evolution of dinosaur diversity in western North America. Southern dinosaur faunas may have been characterized by both higher origination rates and lower extinction rates than northern faunas.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/d16090531/s1, Supplementary Data S1.
Author Contributions
Conceptualization, N.R.L. and J.K.; investigation, N.R.L., A.A.R.V., J.K., C.I.S.-B. and A.E.B.T.; writing—original draft preparation, N.R.L., A.A.R.V., J.K., C.I.S.-B. and A.E.B.T.; writing—review and editing, N.R.L., A.A.R.V., J.K., C.I.S.-B. and A.E.B.T.; visualization, N.R.L., A.A.R.V., C.I.S.-B. and A.E.B.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
All data is available in the manuscript or the online Supplementary Material.
Acknowledgments
Thanks to Jaime Urrutia-Fucugauchi for his gracious hospitality in Mexico City. We would like to thank the support and facilities granted during the review and preparation of material to V. Romero Mayen, J. Alvarado-Ortega, R. Hernández-Rivera, M. Montellano-Ballesteros, and G. Álvarez (IGM). Thanks to C. Díaz Frías for the life reconstruction of the new species. AARV thanks Consejo Nacional de Humanidades, Ciencia y Tecnología (CONAHCYT), and INAH for their support.
Conflicts of Interest
The authors declare no conflict of interest.
Institutional Abbreviations
AMNH, American Museum of Natural History, New York, New York, USA; BENC, Benemérita Escuela Normal de Coahuila, Coahuila, Mexico; BYU, Brigham Young University, Provo, Utah, USA; IGM, Institute of Geology, Universidad Autonoma de Mexico, Mexico City, Mexico; MOR, Museum of the Rockies, Bozeman, Montana, USA; PASAC, Paleontological Association of Sabinas, Coahuila, Sabinas, Mexico; RAM, Raymond Alf Museum, Claremont, California USA; ROM, Royal Ontario Museum, Toronto, Ontario, Canada; TMM, Texas Memorial Museum, Austin, Texas, USA; TMP, Royal Tyrrell Museum of Paleontology, Drumheller, Alberta, Canada.
References
- Weishampel, D.B.; Barrett, P.M.; Coria, R.A.; Le Loeuff, J.; Xu, X.; Zhao, X.; Sahni, A.; Gomani, E.; Noto, C.R. Dinosaur Distribution. In The Dinosauria, 2nd ed.; Weishampel, D.B., Dodson, P., Osmolska, H., Eds.; University of California Press: Berkeley, CA, USA, 2004; pp. 517–606. [Google Scholar]
- Sampson, S.D.; Loewen, M.A.; Farke, A.A.; Roberts, E.M.; Forster, C.A.; Smith, J.A.; Titus, A.L. New horned dinosaurs from Utah provide evidence for intracontinental dinosaur endemism. PLoS ONE 2010, 5, e12292. [Google Scholar] [CrossRef] [PubMed]
- Longrich, N.R. Titanoceratops ouranos, a giant horned dinosaur from the Late Campanian of New Mexico. Cretac. Res. 2011, 32, 264–276. [Google Scholar] [CrossRef]
- Lehman, T.M. Late Cretaceous Dinosaur Provinciality. In Mesozoic Vertebrate Life; Tanke, D.H., Carpenter, K., Farlow, J.A., Eds.; Life of the Past; Indiana University Press: Bloomington, IN, USA, 2001; pp. 310–328. [Google Scholar]
- Loewen, M.A.; Sampson, S.D.; Lund, E.K.; Farke, A.A.; Aguillón-Martínez, M.C.; de Leon, C.A.; Rodríguez-de la Rosa, R.A.; Getty, M.A.; Eberth, D.A. Horned dinosaurs (Ornithischia: Ceratopsidae) from the Upper Cretaceous (Campanian) Cerro del Pueblo Formation, Coahila, Mexico. In New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium; Ryan, M.J., Chinnery, B.J., Eberth, D.A., Eds.; Indiana University Press: Bloomington, IN, USA, 2010; pp. 99–116. [Google Scholar]
- Gates, T.A.; Sampson, S.D.; Delgado de Jesús, C.R.; Zanno, L.E.; Eberth, D.; Hernandez-Rivera, R.; Aguillón Martínez, M.C.; Kirkland, J.I. Velafrons coahuilensis, a new lambeosaurine hadrosaurid (Dinosauria: Ornithopoda) from the Late Campanian Cerro Del Pueblo Formation, Coahila, Mexico. J. Vertebr. Paleontol. 2007, 27, 917–930. [Google Scholar] [CrossRef]
- Lehman, T.M.; Coulson, A.B. A juvenile specimen of the sauropod dinosaur Alamosaurus sanjuanensis from the Upper Cretaceous of Big Bend National Park. J. Paleontol. 2002, 76, 156–172. [Google Scholar] [CrossRef]
- Dalman, S.G.; Loewen, M.A.; Pyron, R.A.; Jasinski, S.E.; Malinzak, D.E.; Lucas, S.G.; Fiorillo, A.R.; Currie, P.J.; Longrich, N.R. A giant tyrannosaur from the Campanian–Maastrichtian of southern North America and the evolution of tyrannosaurid gigantism. Sci. Rep. 2024, 14, 22124. [Google Scholar] [CrossRef]
- Prieto-Márquez, A.; Chiappe, L.M.; Joshi, S.H. The lambeosaurine dinosaur Magnapaulia laticaudus from the Late Cretaceous of Baja California, Northwestern Mexico. PLoS ONE 2012, 7, e38207. [Google Scholar] [CrossRef] [PubMed]
- Prieto-Márquez, A. Skeletal morphology of Kritosaurus navajovius (Dinosauria: Hadrosauridae) from the Late Cretaceous of the North American south-west, with an evaluation of the phylogenetic systematics and biogeography of Kritosaurini. J. Syst. Palaeontol. 2014, 12, 133–175. [Google Scholar] [CrossRef]
- Knowlton, F.H. Flora of the Fruitland and Kirtland Formations; US Government Printing Office: Washington, DC, USA, 1916.
- Wing, S.L.; Boucher, L.D. Ecological aspects of the Cretaceous flowering plant radiation. Annu. Rev. Earth Planet. Sci. 1998, 26, 379–421. [Google Scholar] [CrossRef]
- Estrada-Ruiz, E.; Martínez-Cabrera, H.I.; Callejas-Moreno, J.; Upchurch Jr, G.R. Cretaceous tropical rainforests from northern Mexico and its relationship with south-central North American floras. Polibotanica 2013, 36, 41–61. [Google Scholar]
- Wick, S.L.; Lehman, T.M. A new ceratopsian dinosaur from the Javelina Formation (Maastrichtian) of West Texas and implications for chasmosaurine phylogeny. Naturwissenschaften 2013. [Google Scholar] [CrossRef]
- Miller, I.M.; Johnson, K.R.; Kline, D.E.; Nichols, D.J.; Barclay, R.S. A Late Campanian Flora from the Kaiparowits. In At the Top of the Grand Staircase: The Late Cretaceous of Southern Utah; Titus, A., Loewen, M.A., Eds.; Indiana University Press: Indianapolis, IN, USA, 2013; pp. 107–131. [Google Scholar]
- Cardillo, M. Latitude and rates of diversification in birds and butterflies. Proc. R. Soc. London. Ser. B: Biol. Sci. 1999, 266, 1221–1225. [Google Scholar] [CrossRef]
- Mittelbach, G.G.; Schemske, D.W.; Cornell, H.V.; Allen, A.P.; Brown, J.M.; Bush, M.B.; Harrison, S.P.; Hurlbert, A.H.; Knowlton, N.; Lessios, H.A. Evolution and the latitudinal diversity gradient: Speciation, extinction and biogeography. Ecol. Lett. 2007, 10, 315–331. [Google Scholar] [CrossRef]
- Roy, K.; Jablonski, D.; Valentine, J.M.; Rosenberg, G. Marine latitudinal diversity gradients: Tests of causal hypotheses. Proc. Natl. Acad. Sci. USA 1998, 95, 3699–3702. [Google Scholar] [CrossRef]
- Kirkland, J.I.; Hernández-Rivera, R.; Gates, T.; Paul, G.S.; Nesbitt, S.; Serrano-Brañas, C.I.; Garcia-de la Garza, J.P. Large hadrosaurine dinosaurs from the latest Campanian of Coahuila, Mexico. New Mex. Mus. Nat. Hist. Sci. Bull. 2006, 35, 299–315. [Google Scholar]
- Serrano-Brañas, C.I.; Espinosa-Chávez, B.; Maccracken, S.A.; Torres-Rodríguez, E. The Cerro del Pueblo Formation, Unlocking the Environmental Data of an Extraordinary Ancient Ecosystem from Mexico. In Past Environments of Mexico: Unveiling the Past Environments of a Megadiverse Country Through its Fossil Record; Springer: Berlin/Heidelberg, Germany, 2024; pp. 405–426. [Google Scholar]
- de Jesús, C.R.D.; Eberth, D.A.; de la Rosa, R.A.R.; Lerbekmo, J.F.; Brinkman, D.B.; Sampson, S.D. Cerro del Pueblo Fm (Difunta Group, Upper Cretaceous), Parras Basin, southern Coahuila, Mexico: Reference sections, age, and correlation. Rev. Mex. De Cienc. Geol. 2004, 21, 335–352. [Google Scholar]
- Vogt, M.; Stinnesbeck, W.; Zell, P.; Kober, B.; Kontny, J.; Herzer, N.; Frey, E.; Rivera-Sylva, H.E.; Gutierrez, J.M.P.; Amezcua, N. Age and depositional environment of the “dinosaur graveyard” at Las Águilas, southern Coahuila, NE Mexico. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 441, 758–769. [Google Scholar] [CrossRef]
- Eberth, D.A.; Kamo, S.L. High-precision U–PB CA–ID–TIMS dating and chronostratigraphy of the dinosaur-rich Horseshoe Canyon Formation (Upper Cretaceous, Campanian–Maastrichtian), Red Deer River Valley, Alberta, Canada. Can. J. Earth Sci. 2020, 57, 1220–1237. [Google Scholar] [CrossRef]
- Ramezani, J.; Beveridge, T.L.; Rogers, R.R.; Eberth, D.A.; Roberts, E.M. Calibrating the zenith of dinosaur diversity in the Campanian of the Western Interior Basin by CA-ID-TIMS U–Pb geochronology. Sci. Rep. 2022, 12, 16026. [Google Scholar] [CrossRef]
- Serrano-Brañas, C.I.; Espinosa-Chávez, B.; Maccracken, S.A.; Gutiérrez-Blando, C.; de León-Dávila, C.; Ventura, J.F. Paraxenisaurus normalensis, a large deinocheirid ornithomimosaur from the Cerro del Pueblo Formation (Upper Cretaceous), Coahuila, Mexico. J. South Am. Earth Sci. 2020, 101, 102610. [Google Scholar] [CrossRef]
- Martinez, M.C.A. Fossil Vertebrates from the Cerro del Pueblo Formation, Coahuila, Mexico, and the Distribution of Late Campanian (Cretaceous) Terrestrial Vertebrate Faunas. Master’s Thesis, Southern Methodist University, Dallas, TX, USA, 2010. [Google Scholar]
- Serrano-Brañas, C.I.; Espinosa-Chávez, B.; Maccracken, S.A.; Guevara, D.B.; Torres-Rodríguez, E. First record of caenagnathid dinosaurs (Theropoda, Oviraptorosauria) from the Cerro del Pueblo Formation (Campanian, Upper Cretaceous), Coahuila, Mexico. J. South Am. Earth Sci. 2022, 119, 104046. [Google Scholar] [CrossRef]
- Ramírez-Velasco, A.; Hernández-Rivera, R. Diversity of late cretaceous dinosaurs from Mexico. Boletín Geológico Y Min. 2015, 126, 63–108. [Google Scholar]
- Aguillón-Martínez, M.C.; Rivera-Sylva, H.E. A Troodontid (Theropoda: Troodontidae) Neurocranium from the Cerro del Pueblo Formation (Late Campanian) of Coahuila, Mexico. Paleontol. Mex. 2023, 12, 99–105. [Google Scholar]
- Rivera-Sylva, H.E.; Carpenter, K.; Aranda-Manteca, F.J. Late cretaceous nodosaurids (Ankylosauria: Ornithischia) from Mexico. Rev. Mex. De Cienc. Geol. 2011, 28, 371–378. [Google Scholar]
- Rivera-Sylva, H.E.; Espinosa-Chávez, B. Ankylosaurid (dinosauria: Thyreophora) osteoderms from the upper Cretaceous Cerro del Pueblo Formation of Coahuila, Mexico. Carnets De Geol. 2006, 02, 1–5. [Google Scholar] [CrossRef][Green Version]
- Rivera-Sylva, H.E.; Frey, E.; Stinnesbeck, W.; Amezcua, N.; Flores Huerta, D. First occurrence of Parksosauridae in Mexico, from the Cerro del Pueblo Formation (Late Cretaceous; late Campanian) at Las Águilas, Coahuila. Boletín De La Soc. Geol. Mex. 2018, 70, 779–785. [Google Scholar] [CrossRef]
- Velasco, A.A.R.; Aguilar, F.J.; Hernández-Rivera, R.; Maussán, J.L.G.; Rodríguez, M.L.; Alvarado-Ortega, J. Tlatolophus galorum, gen. et sp. nov., a parasaurolophini dinosaur from the upper Campanian of the Cerro del Pueblo Formation, Coahuila, northern Mexico. Cretac. Res. 2021, 126, 104884. [Google Scholar] [CrossRef]
- Ramírez-Velasco, Á.A.; Espinosa-Arrubarrena, L.; Alvarado-Ortega, J. Review of the taxonomic affinities of Latirhinus uitstlani, an emblematic Mexican hadrosaurid. J. South Am. Earth Sci. 2021, 110, 103391. [Google Scholar] [CrossRef]
- Serrano-Brañas, C.I.; Prieto-Márquez, A. Taphonomic attributes of the holotype of the lambeosaurine dinosaur Latirhinus uitstlani from the late Campanian of Mexico: Implications for its phylogenetic systematics. J. South Am. Earth Sci. 2022, 114, 103689. [Google Scholar] [CrossRef]
- Ramírez-Velasco, A.A.; Hernández-Rivera, R.; Servin-Pichardo, R.; Evans, D.; Eberth, D. The hadrosaurian record from Mexico. In Hadrosaurs; Eberth, D.A., Evans, D.C., Eds.; Indiana University Press: Bloomington, IN, USA, 2014; Volume Bloomington and Indianapolis, pp. 340–360. [Google Scholar]
- Prieto-Márquez, A.; Serrano Brañas, C.I. Latirhinus uitstlani, a ‘broad-nosed’saurolophine hadrosaurid (Dinosauria, Ornithopoda) from the late Campanian (Cretaceous) of northern Mexico. Hist. Biol. 2012, 24, 607–619. [Google Scholar] [CrossRef]
- Ramírez-Velasco, A.A. Phylogenetic and biogeography analysis of Mexican hadrosauroids. Cretac. Res. 2022, 138, 105267. [Google Scholar] [CrossRef]
- Prieto-Márquez, A.; Wagner, J.R. A new species of saurolophine hadrosaurid dinosaur from the Late Cretaceous of the Pacific coast of North America. Acta Palaeontol. Pol. 2013, 58, 255–268. [Google Scholar]
- Prieto-Márquez, A.; Wagner, J.R. A new ‘duck-billed’dinosaur (Ornithischia: Hadrosauridae) from the upper Campanian of Texas points to a greater diversity of early hadrosaurid offshoots. Cretac. Res. 2023, 143, 105416. [Google Scholar] [CrossRef]
- Fowler, D.W. Revised geochronology, correlation, and dinosaur stratigraphic ranges of the Santonian-Maastrichtian (Late Cretaceous) formations of the Western Interior of North America. PLoS ONE 2017, 12, e0188426. [Google Scholar] [CrossRef]
- Mallon, J.C.; Evans, D.C.; Ryan, M.J.; Anderson, J.S. Megaherbivorous dinosaur turnover in the Dinosaur Park Formation (upper Campanian) of Alberta, Canada. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 350–352, 124–138. [Google Scholar] [CrossRef]
- Scannella, J.B.; Fowler, D.W.; Goodwin, M.B.; Horner, J.R. Evolutionary trends in Triceratops from the Hell Creek Formation, Montana. Proc. Natl. Acad. Sci. USA 2014, 111, 10245–10250. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Brañas, C.I.; Espinosa-Chávez, B.; Maccracken, S.A. Teredolites trace fossils in log-grounds from the Cerro del Pueblo Formation (Upper Cretaceous) of the state of Coahuila, Mexico. J. South Am. Earth Sci. 2019, 95, 102316. [Google Scholar] [CrossRef]
- Maddison, W.; Maddison, D. Mesquite: A Modular System for Evolutionary Analysis, v. 3.70; Mesquite Software, Inc.: Austin, TX, USA, 2024.
- Swofford, D.L. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods); 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Owen, R. Report on British fossil reptiles. Part II. Rep. Br. Assoc. Adv. Sci. 1842, 11, 60–204. [Google Scholar]
- Marsh, O.C. Classification of the Dinosauria. Am. J. Sci. 1881, 23, 81–86. [Google Scholar] [CrossRef]
- Cope, E.D. On the reptilian orders Pythonomorpha and Streptosauria. Proc. Boston Soc. Nat. Hist. 1869, 12, 250–267. [Google Scholar]
- Brett-Surman, M.K. A Revision of the Hadrosauridae (Reptilia: Ornithischia) and Their Evolution during the Campanian and Maastrichtian; The George Washington University: Washington, DC, USA, 1989. [Google Scholar]
- Brown, B. Corythosaurus casuarius, a new crested dinosaur from the Belly River Cretaceous; with Provisional classification of the family Trachodontidae. Bull. AMNH 1914, 33, 35. [Google Scholar]
- Gates, T.A.; Sampson, S. A new species of Gryposaurus (Dinosauria: Hadrosauridae) from the late Campanian Kaiparowits Formation, southern Utah, USA. Zool. J. Linn. Soc. 2007, 151, 351–376. [Google Scholar] [CrossRef]
- Lowi-Merri, T.M.; Evans, D.C. Cranial variation in Gryposaurus and biostratigraphy of hadrosaurines (Ornithischia: Hadrosauridae) from the Dinosaur Park Formation of Alberta, Canada. Can. J. Earth Sci. 2020, 57, 765–779. [Google Scholar] [CrossRef]
- Lehman, T.M.; Wick, S.L.; Wagner, J.R. Hadrosaurian dinosaurs from the Maastrichtian Javelina Formation, Big Bend National Park, Texas. J. Paleontol. 2016, 90, 333–356. [Google Scholar] [CrossRef]
- Rozadilla, S.; Brissón-Egli, F.; Agnolín, F.L.; Aranciaga-Rolando, A.M.; Novas, F.E. A new hadrosaurid (Dinosauria: Ornithischia) from the Late Cretaceous of northern Patagonia and the radiation of South American hadrosaurids. J. Syst. Palaeontol. 2021, 19, 1207–1235. [Google Scholar] [CrossRef]
- Xing, H.; Mallon, J.C.; Currie, M.L. Supplementary cranial description of the types of Edmontosaurus regalis (Ornithischia: Hadrosauridae), with comments on the phylogenetics and biogeography of Hadrosaurinae. PLoS ONE 2017, 12, e0175253. [Google Scholar] [CrossRef] [PubMed]
- Campione, N.E. Cranial variation in Edmontosaurus (Hadrosauridae) from the Late Cretaceous of North America. N. Am. Paleontol. Conv. 2009, NAPC 2009, 95. [Google Scholar]
- Prieto-Marquez, A. New information on the cranium of Brachylophosaurus canadensis (Dinosauria, Hadrosauridae), with a revision of its phylogenetic position. J. Vertebr. Paleontol. 2005, 25, 144–156. [Google Scholar] [CrossRef]
- Prieto-Márquez, A. The skull and appendicular skeleton of Gryposaurus latidens, a saurolophine hadrosaurid (Dinosauria: Ornithopoda) from the early Campanian (Cretaceous) of Montana, USA. Can. J. Earth Sci. 2012, 49, 510–532. [Google Scholar] [CrossRef]
- Clark, J.M.; Norell, M.A.; Rowe, T. Cranial anatomy of Citipati osmolskae (Theropoda, Oviraptorosauria) and a reinterpretation of the holotype of Oviraptor philoceratops. Am. Mus. Novit. 2002, 3364, 1–24. [Google Scholar] [CrossRef]
- Osmólska, H.; Currie, P.J.; Barsbold, R. Oviraptorosauria. In The Dinosauria; Weishampel, D.B., Dodson, P., Osmolska, H., Eds.; University of California Press: Berkeley, CA, USA, 2004; pp. 165–183. [Google Scholar]
- Prieto-Márquez, A. A re-evaluation of Secernosaurus koerneri and Kritosaurus australis (Dinosauria, Hadrosauridae) from the Late Cretaceous of Argentina. J. Vertebr. Paleontol. 2010, 30, 813–837. [Google Scholar] [CrossRef]
- Horner, J.R.; Weishampel, D.B.; Forster, C.A. Hadrosauridae. In The Dinosauria; Weishampel, D.B., Dodson, P., Osmolska, H., Eds.; University of California Press: Berkeley, CA, USA, 2004; pp. 438–463. [Google Scholar]
- Brown, B. A crested dinosaur from the Edmonton Cretaceous. Bull. Am. Mus. Nat. Hist. 1912, 31, 131. [Google Scholar]
- Brown, B. The skeleton of Saurolophus, a crested duck-billed dinosaur from the Edmonton Cretaceous. Bull. AMNH 1913, 32, 19. [Google Scholar]
- Maryanska, T.; Osmólska, H. Cranial anatomy of Saurolophus angustirostris with comments on the Asian Hadrosauridae (Dinosauria). Palaeontol. Pol. 1981, 42, 5–24. [Google Scholar]
- Lambe, L. On Gryposaurus notabilis, a new genus and species of trachodont dinosaur from the Belly River Formation of Alberta, with a description of the skull of Chasmosaurus belli. Ott. Nat. 1914, 27, 145–155. [Google Scholar]
- Gates, T.A.; Scheetz, R. A new saurolophine hadrosaurid (Dinosauria: Ornithopoda) from the Campanian of Utah, North America. J. Syst. Palaeontol. 2015, 13, 711–725. [Google Scholar] [CrossRef]
- Erickson, G.M.; Krick, B.A.; Hamilton, M.; Bourne, G.R.; Norell, M.A.; Lilleodden, E.; Sawyer, W.G. Complex dental structure and wear biomechanics in hadrosaurid dinosaurs. Science 2012, 338, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Prieto-Marquez, A. Global phylogeny of Hadrosauridae (Dinosauria: Ornithopoda) using parsimony and Bayesian methods. Zool. J. Linn. Soc. 2010, 159, 435–502. [Google Scholar] [CrossRef]
- Congreve, C.R.; Lamsdell, J.C. Implied weighting and its utility in palaeontological datasets: A study using modelled phylogenetic matrices. Palaeontology 2016, 59, 447–462. [Google Scholar] [CrossRef]
- Goloboff, P.A. Self-Weighted Optimization: Tree Searches and Character State Reconstructions under Implied Transformation Costs. Cladistics 1997, 13, 225–245. [Google Scholar]
- Ezcurra, M.D. Exploring the effects of weighting against homoplasy in genealogies of palaeontological phylogenetic matrices. Cladistics 2024, 40, 242–281. [Google Scholar] [CrossRef]
- Longrich, N.R.; Field, D. Torosaurus is not Triceratops: Ontogeny in Chasmosaurine Ceratopsids as a Case Study in Dinosaur Taxonomy. PLoS ONE 2012, 7, e32623. [Google Scholar] [CrossRef]
- Lucas, S.G.; Spielmann, J.A.; Sullivan, R.M.; Hunt, A.P.; Gates, T.A. Anasazisaurus, a hadrosaurian dinosaur from the Upper Cretaceous of New Mexico. New Mex. Mus. Nat. Hist. Sci. Bull. 2006, 35, 293–298. [Google Scholar]
- Knell, R.J.; Naish, D.; Tomkins, J.L.; Hone, D.W.E. Sexual selection in prehistoric animals: Detection and implications. Trends Ecol. Evol. 2013, 28, 38–47. [Google Scholar] [CrossRef]
- Bell, P.R.; Fanti, F.; Currie, P.J.; Arbour, V.M. A mummified duck-billed dinosaur with a soft-tissue cock’s comb. Curr. Biol. 2014, 24, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Mayr, G. A survey of casques, frontal humps, and other extravagant bony cranial protuberances in birds. Zoomorphology 2018, 137, 457–472. [Google Scholar] [CrossRef]
- Kinnaird, M.F.; Hadiprakarsa, Y.-Y.; Thiensongrusamee, P. Aerial jousting by helmeted hornbills Rhinoplax vigil: Observations from Indonesia and Thailand. Ibis 2003, 145, 506–508. [Google Scholar] [CrossRef]
- Weishampel, D.B. Acoustic analyses of potential vocalization in lambeosaurine dinosaurs (Reptilia: Ornithischia). Paleobiology 1981, 7, 252–261. [Google Scholar] [CrossRef]
- Evans, D.C.; Ridgely, R.; Witmer, L.M. Endocranial anatomy of lambeosaurine hadrosaurids (Dinosauria: Ornithischia): A sensorineural perspective on cranial crest function. Anat. Rec. Adv. Integr. Anat. Evol. Biol. Adv. Integr. Anat. Evol. Biol. 2009, 292, 1315–1337. [Google Scholar] [CrossRef]
- Lehman, T.M. Late Campanian dinosaur biogeography in the Western Interior of North America. In Dinofest International, Proceedings of a Symposium Sponsored by Arizona State University, Special Publication, Academy of Natural Sciences; Wolberg, D.A., Stump, E., Eds.; Academy of Natural Sciences: Philidelphia, PA, USA, 1997; pp. 223–240. [Google Scholar]
- Gates, T.; Sampson, S.; Zanno, L.; Roberts, E.; Eaton, J.; Nydam, R.; Hutchison, J.; Smith, J.; Loewen, M.; Getty, M. Biogeography of terrestrial and freshwater vertebrates from the Late Cretaceous (Campanian) Western Interior of North America. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 291, 371–387. [Google Scholar] [CrossRef]
- Sampson, S.D.; Lund, E.K.; Loewen, M.A.; Farke, A.A.; Clayton, K.E. A remarkable short-snouted horned dinosaur from the Late Cretaceous (late Campanian) of southern Laramidia. Proc. R. Soc. B 2013, 280. [Google Scholar] [CrossRef]
- Russell, D.; Manabe, M. Synopsis of the Hell Creek (uppermost Cretaceous) dinosaur assemblage. GSA Spec. Pap. 2002, 361, 169–176. [Google Scholar]
- Parks, W.A. Parasaurolophus walkeri: A new genus and species of crested trachodont dinosaur. Univ. Tor. Stud. Geol. Ser. 1922, 13, 5–32. [Google Scholar]
- Brown, B. A new trachodont dinosaur, Hypacrosaurus, from the Edmonton Cretaceous of Alberta. Bull. Am. Mus. Nat. Hist. 1913, 32, 395–406. [Google Scholar]
- Sullivan, R.M.; Jasinski, S.E.; Guenther, M.F.; Lucas, S.G. The first lambeosaurin (Dinosauria, Hadrosauridae, Lambeosaurinae) from the Upper Cretaceous Ojo Alamo Formation (Naashoibito Member), San Juan Basin, New Mexico. New Mex. Mus. Nat. Hist. Sci. Bull. 2011, 53, 405–417. [Google Scholar]
- Longrich, N.R. A new, large ornithomimid from the Dinosaur Park Formation of Alberta, Canada: Implications for the study of dissociated dinosaur remains. Palaeontology 2008, 51, 983–997. [Google Scholar] [CrossRef]
- Lawson, D.A. Tyrannosaurus and Torosaurus, Maestrichtian dinosaurs from Trans-Pecos, Texas. J. Paleontol. 1976, 50, 158–164. [Google Scholar]
- Loewen, M.A.; Irmis, R.B.; Sertich, J.J.; Currie, P.J.; Sampson, S.D. Tyrant dinosaur evolution tracks the rise and fall of Late Cretaceous oceans. PLoS ONE 2013, 8, e79420. [Google Scholar] [CrossRef]
- Jablonski, D.; Roy, K.; Valentine, J.W. Out of the tropics: Evolutionary dynamics of the latitudinal diversity gradient. Science 2006, 314, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Price, G.D.; Twitchett, R.J.; Wheeley, J.R.; Buono, G. Isotopic evidence for long term warmth in the Mesozoic. Sci. Rep. 2013, 3, 1438. [Google Scholar] [CrossRef] [PubMed]
- Grossman, E.L.; Joachimski, M.M. Ocean temperatures through the Phanerozoic reassessed. Sci. Rep. 2022, 12, 8938. [Google Scholar] [CrossRef]
- Gilmore, C.W. Reptilian fauna of the North Horn Formation of central Utah. United States Geol. Surv. Prof. Pap. 1946, 210-C, 29–53. [Google Scholar]
- Jasinski, S.E.; Sullivan, R.M.; Lucas, S.G. Taxonomic composition of the Alamo Wash local fauna from the Upper Cretaceous Ojo Alamo Formation (Naashoibito Member), San Juan Basin, New Mexico. New Mex. Mus. Nat. Hist. Sci. Bull. 2011, 53, 216–265. [Google Scholar]
- Condamine, F.L.; Guinot, G.; Benton, M.J.; Currie, P.J. Dinosaur biodiversity declined well before the asteroid impact, influenced by ecological and environmental pressures. Nat. Commun. 2021, 12, 3833. [Google Scholar] [CrossRef]
- Sakamoto, M.; Benton, M.J.; Venditti, C. Dinosaurs in decline tens of millions of years before their final extinction. Proc. Natl. Acad. Sci. USA 2016, 113, 5036–5040. [Google Scholar] [CrossRef] [PubMed]



















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).