
Arachnid Assemblage Composition Diverge between South- and North-Facing Slopes in a Levantine Microgeographic Site †

 , , and
, , and
Abstract

1. Introduction
2. Materials and Methods
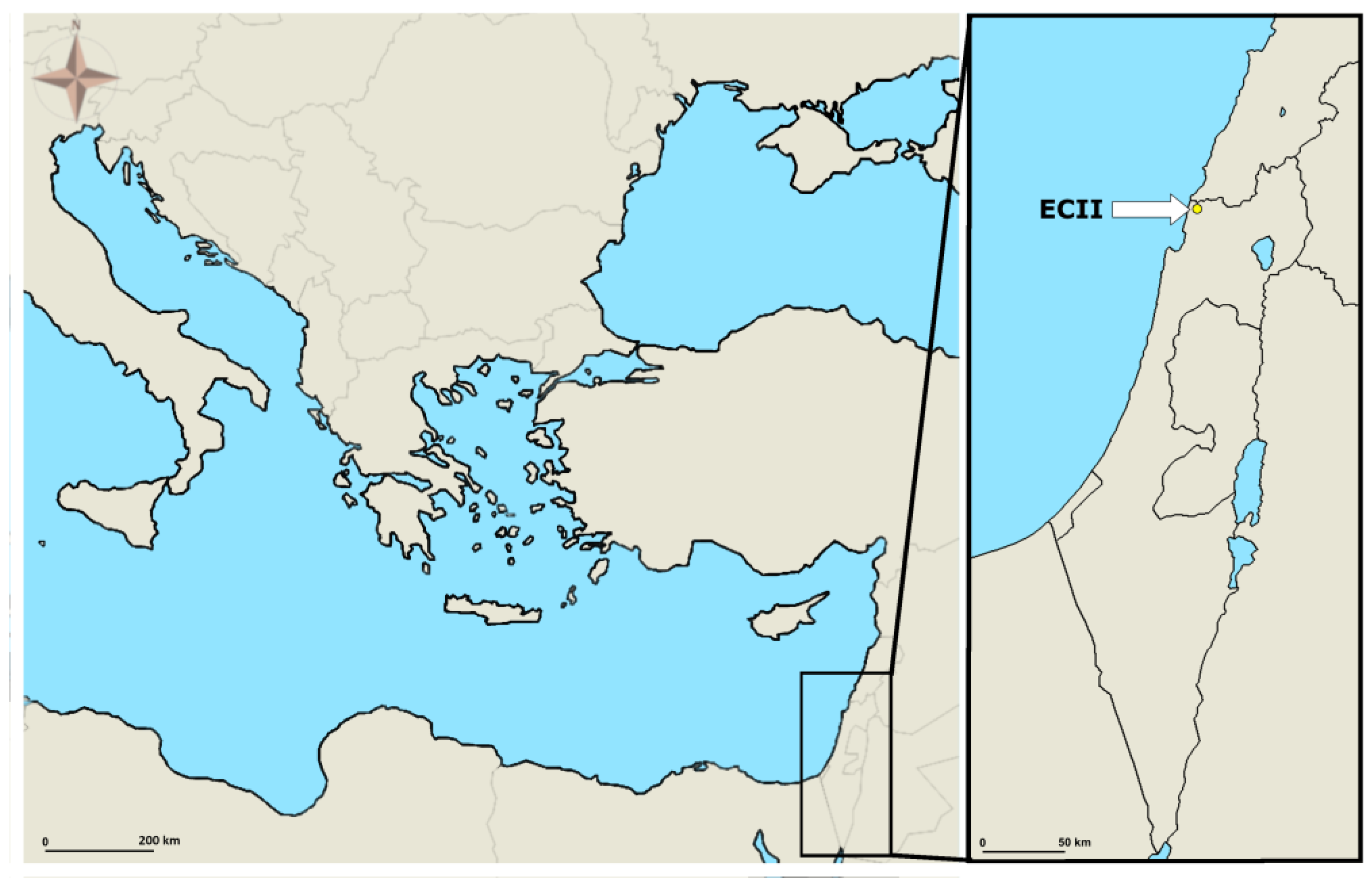
2.1. The Study Area
2.2. Arachnid Sampling, Sorting, and Identification
2.3. Statistical Analysis
2.4. Species Descriptions
2.5. DNA Extraction, COI Sequence Acquisition and Analysis
3. Results
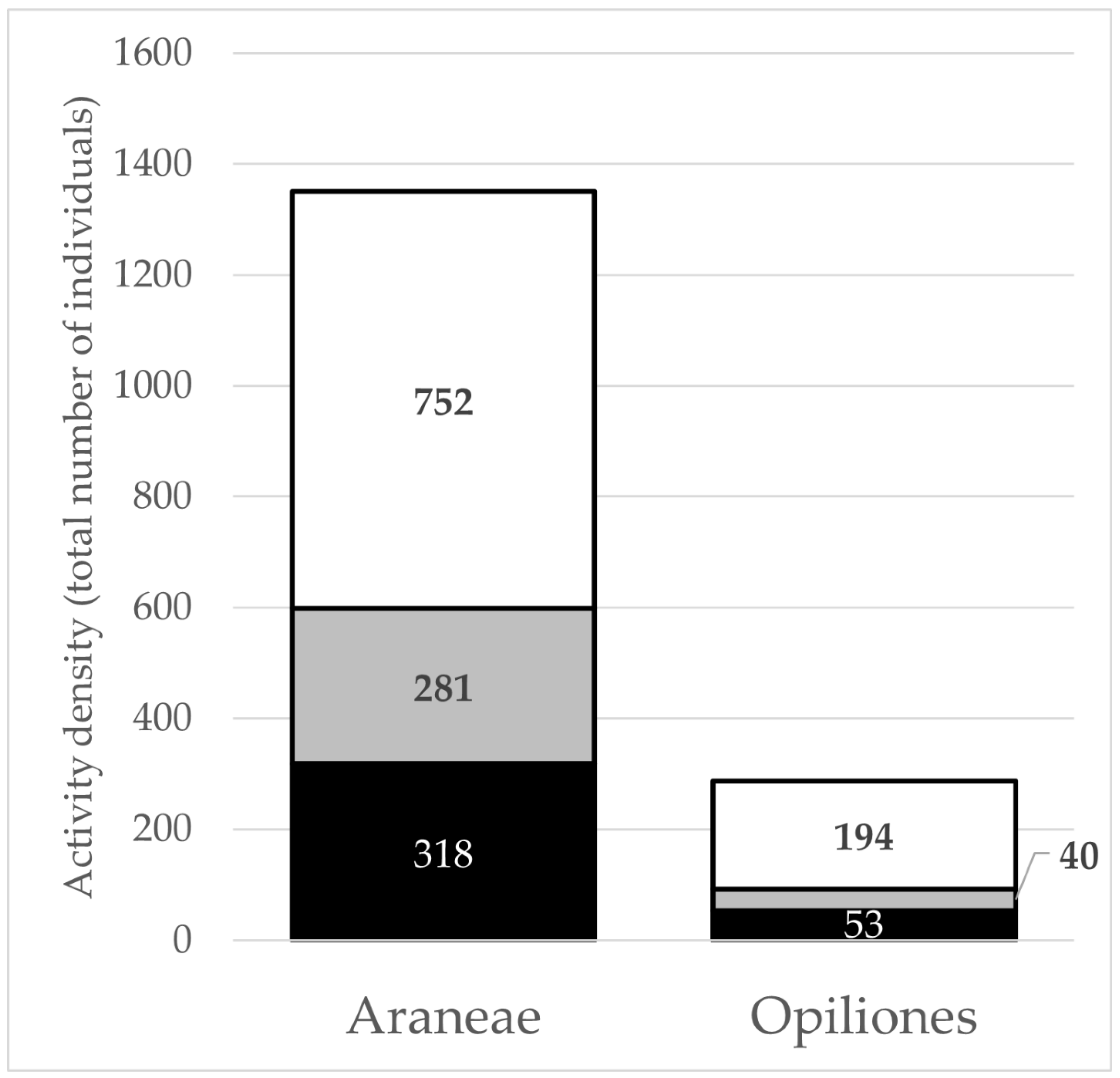
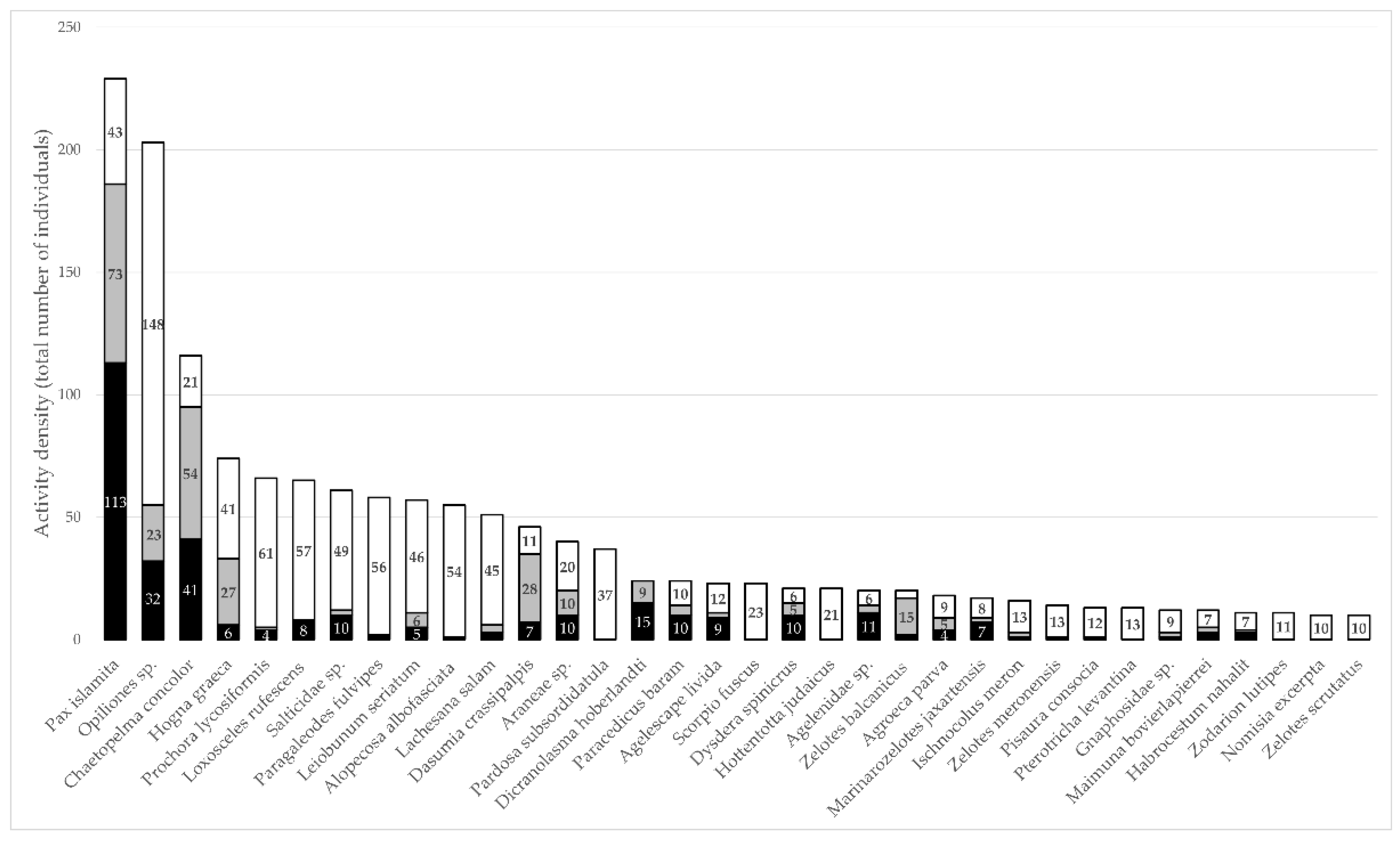
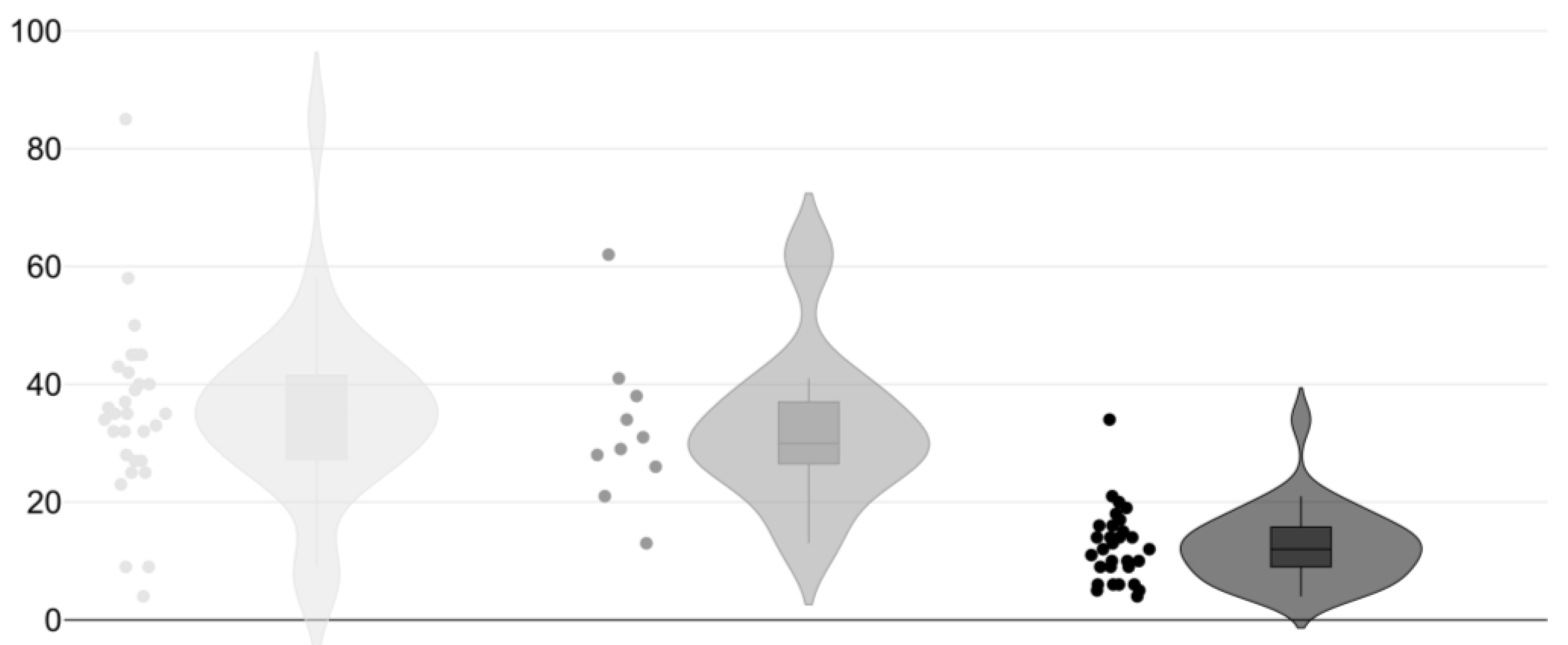
3.1. Arachnid Abundance, Richness, Distribution, and Biogeography
3.2. Seasonality
3.3. Habitat and Microhabitat Use
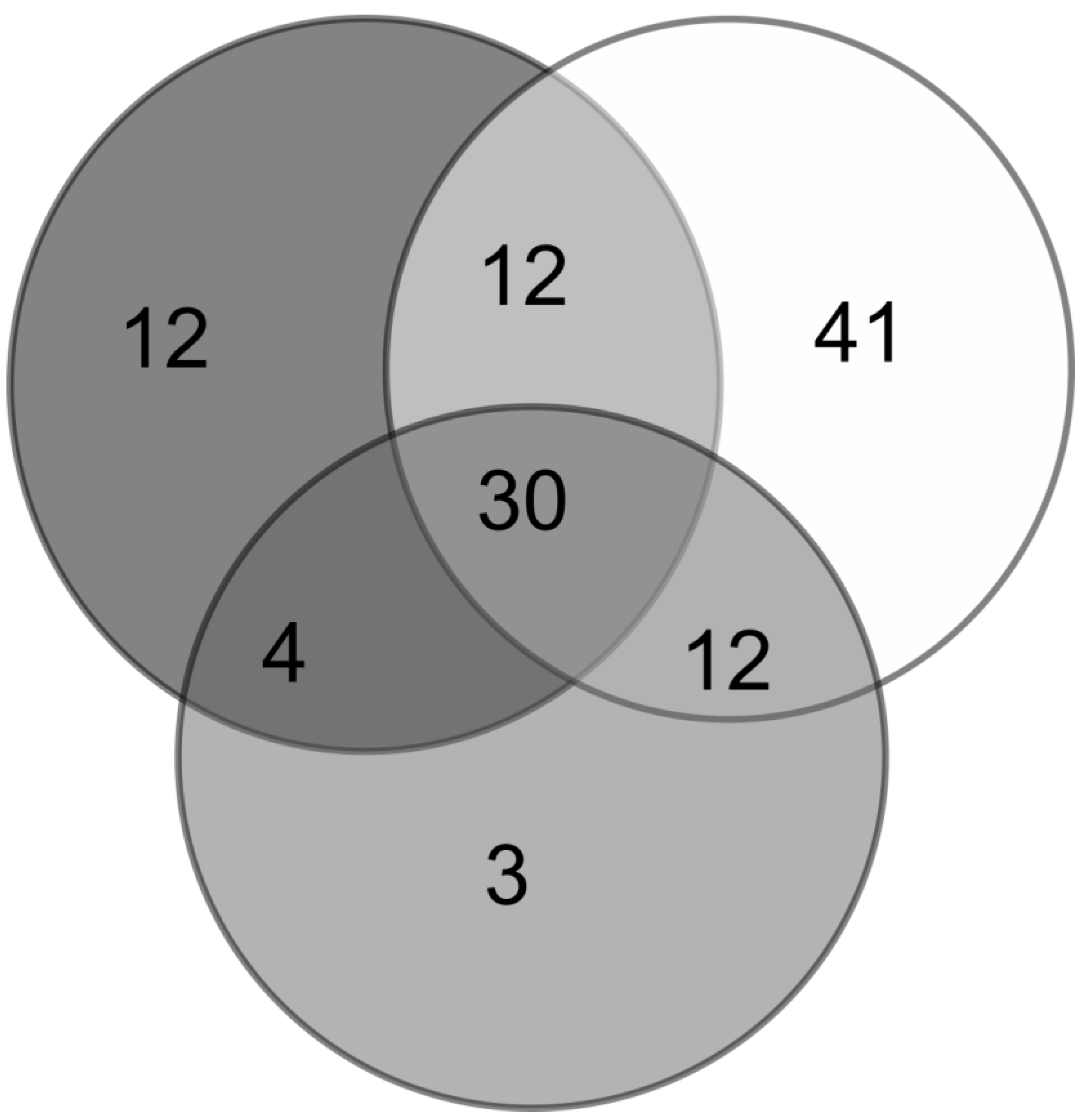
3.4. Arachnid Assemblage Composition
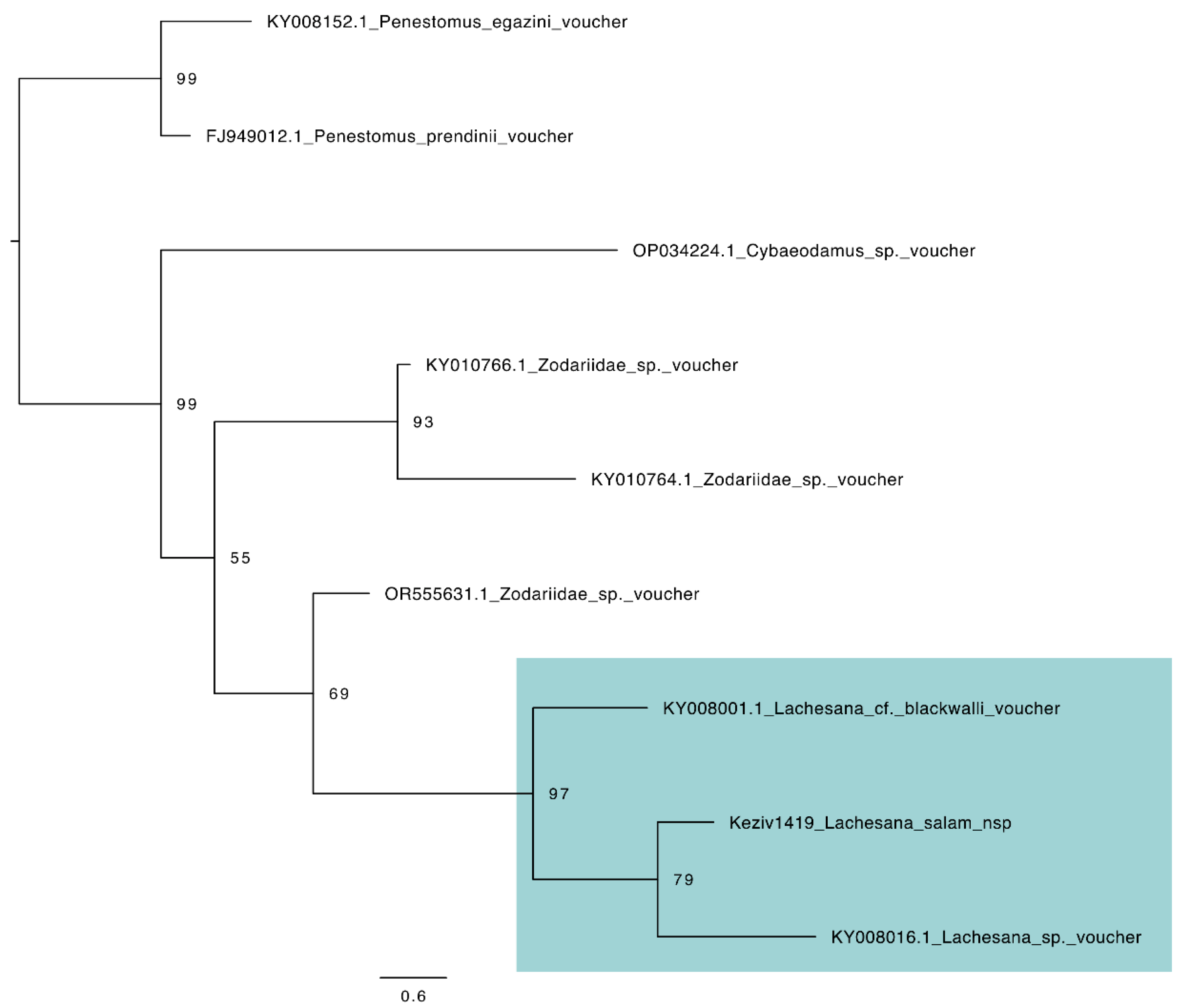
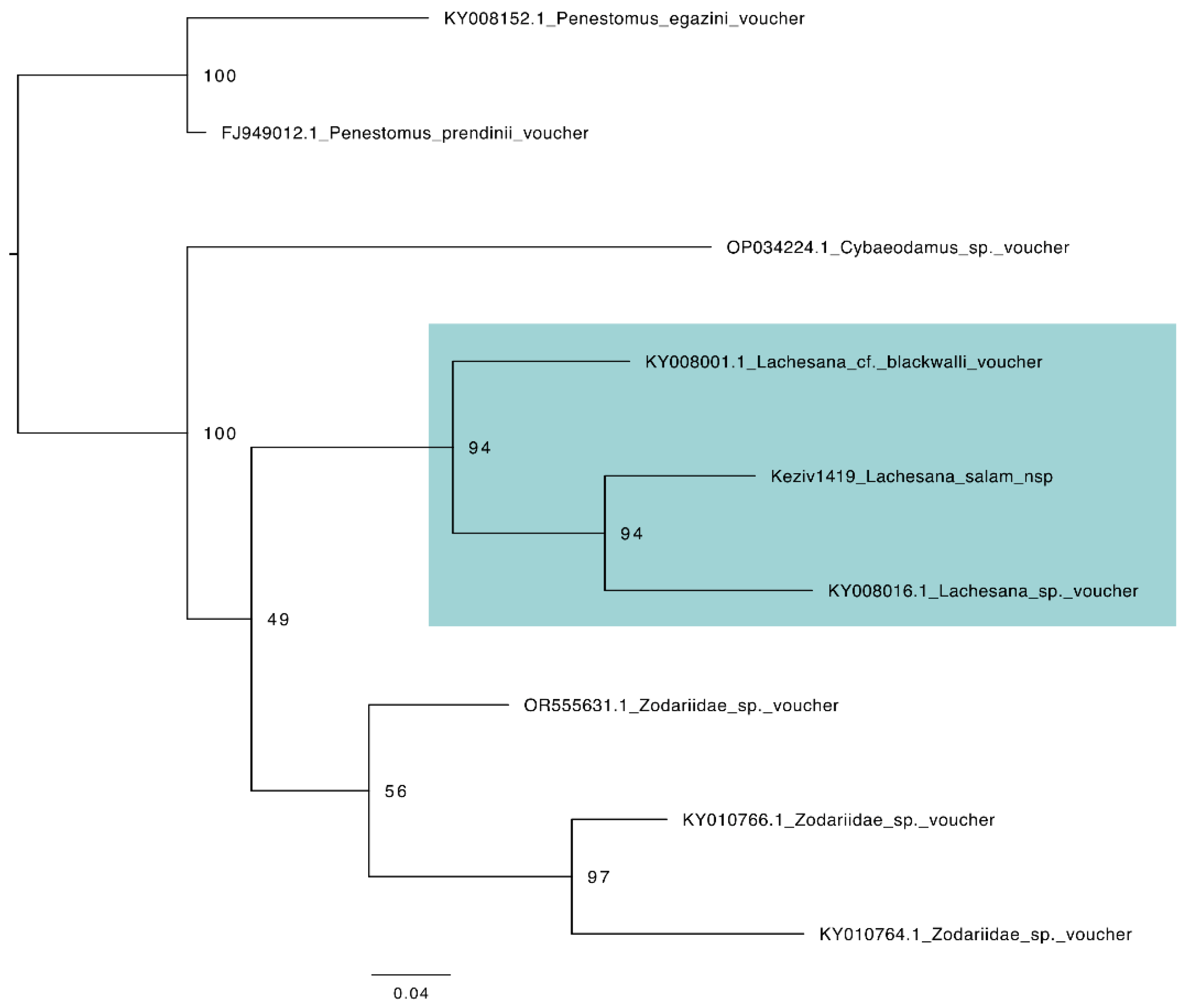
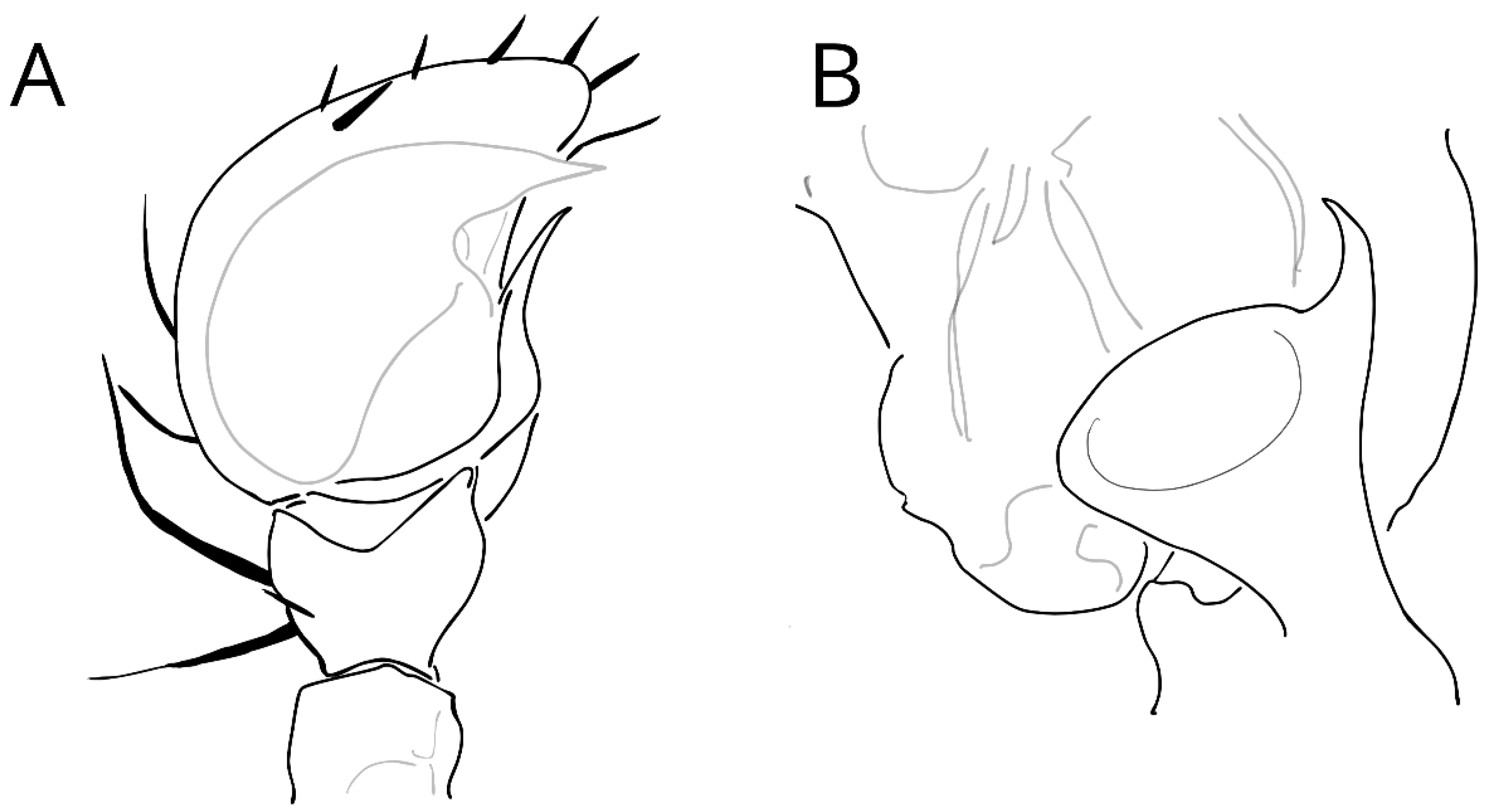
3.5. Species Description
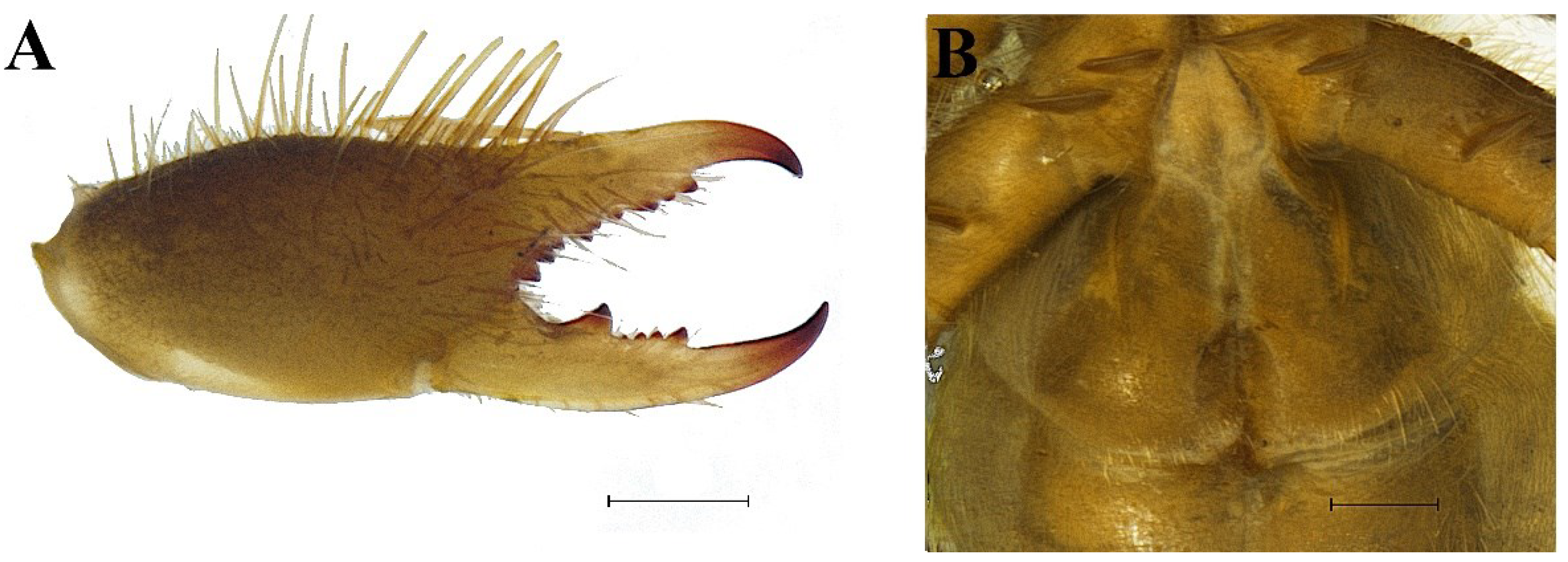
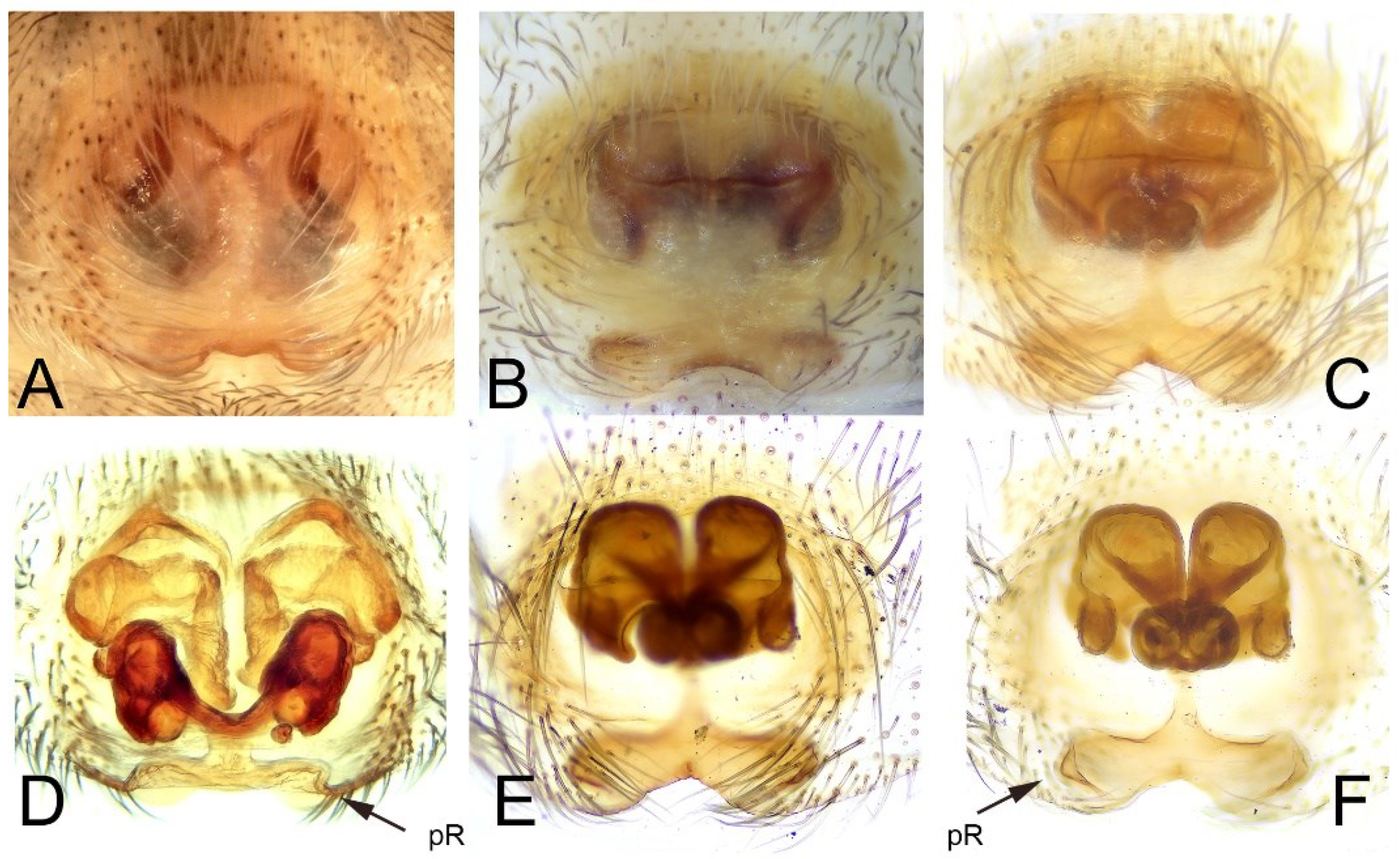
3.5.1. Taxonomy and Description
Family Salticidae Blackwall, 1841.Genus Habrocestum Simon, 1876
Family Zodariidae Simon, 1864
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specimen Code | Species | Sex | Year Collected | Locality | GenBank Accession |
|---|---|---|---|---|---|
| Keziv 1419 | Lachesanasalam | Male | 2020 | Nahal Keziv, Galilee, Israel | PQ300647 |
| >seq96__20967_G01_7591858.ab1 NNNNNNGNTNGNNNCCTGATATAGCTTTTCCTCGAATGAATAATTTAAGATTTTGAT- TATTGCCTCCTTCTTTGTTGTTGTTATTCATTTCTTCTATAGTAGAAATAGGAGTTGGAGCAGGATGAACAATCTA TCCTCCATTAGCTTCTTTAATAGGTCATTCTGGTGAATCTGTCGATTTTGC- TATTTTTTCGCTTCATTTAGCAGGTGCTTCTTCTATTTTAGGCTCTGTAAATTTTA TTTCAACTATTATTAATATACGTTCTTATGGAATAAGAATGGAAAAAGTTCCTTTGTTT- GTTTGGTCTGTCTTAGTAACAACTGTATTGTTATTATTGTCTTTACCTGTATTAGCAGGTGCTATTACTATATTATTA ACAGATCGAAATTTTAATACTTCATTTTTTGATCCGGCTGGAGGAGGAGATCCAATTTT- GTTTCAACATCTATTTTNCNNNNNTTGAAAACCCTGAAN | |||||
References
- Willis, K.J.; Whittaker, R.J. Species Diversity: Scale Matters. Science 2002, 295, 1245–1248. [Google Scholar] [CrossRef]
- Trivellone, V.; Bougeard, S.; Giavi, S.; Krebs, P.; Balseiro, D.; Dray, S.; Moretti, M. Factors Shaping Community Assemblages and Species Co-Occurrence of Different Trophic Levels. Ecol. Evol. 2017, 7, 4745–4754. [Google Scholar] [CrossRef]
- Pulliam, H.R. On the Relationship between Niche and Distribution. Ecol. Lett. 2000, 3, 349–361. [Google Scholar] [CrossRef]
- Zobel, M. The Relative of Species Pools in Determining Plant Species Richness: An Alternative Explanation of Species Coexistence? Trends Ecol. Evol. 1997, 12, 266–269. [Google Scholar] [CrossRef]
- Zobel, M.; Otto, R.; Laanisto, L.; Naranjo-Cigala, A.; Pärtel, M.; Fernández-Palacios, J.M. The Formation of Species Pools: Historical Habitat Abundance Affects Current Local Diversity: Species Pool Formation and Current Diversity. Glob. Ecol. Biogeogr. 2011, 20, 251–259. [Google Scholar] [CrossRef]
- Martín-Devasa, R.; Jiménez-Valverde, A.; Leprieur, F.; Baselga, A.; Gómez-Rodríguez, C. Dispersal Limitation Shapes Distance-decay Patterns of European Spiders at the Continental Scale. Glob. Ecol. Biogeogr. 2024, 33, e13810. [Google Scholar] [CrossRef]
- Jiménez-Valverde, A.; Lobo, J.M. Determinants of Local Spider (Araneidae and Thomisidae) Species Richness on a Regional Scale: Climate and Altitude vs. Habitat Structure. Ecol. Entomol. 2007, 32, 113–122. [Google Scholar] [CrossRef]
- Shreve, F. Conditions Indirectly Affecting Vertical Distribution on Desert Mountains. Ecology 1922, 3, 269. [Google Scholar] [CrossRef]
- Nevo, E.; Travleev, A.P.; Belova, N.A.; Tsatskin, A.; Pavlíček, T.; Kulik, A.F.; Tsvetkova, N.N.; Yemshanov, D.C. Edaphic Interslope and Valley Bottom Differences at “Evolution Canyon”, Lower Nahal Oren, Mount Carmel, Israel. Catena 1998, 33, 241–254. [Google Scholar] [CrossRef]
- Pavlícek, T.; Sharon, D.; Kravchenko, V.; Saaroni, H.; Nevoa, E. Microclimatic Interslope Differences Underlying Biodiversity Contrasts in “Evolution Canyon”, Mt. Carmel, Israel. Isr. J. Earth Sci. 2003, 52, 1–9. [Google Scholar] [CrossRef]
- Nevo, E. Asian, African and European Biota Meet at ‘Evolution Canyon’ Israel: Local Tests of Global Biodiversity and Genetic Diversity Patterns. Proc. R. Soc. Lond. B 1995, 262, 149–155. [Google Scholar] [CrossRef]
- Nevo, E. Evolution in Action across Phylogeny Caused by Microclimatic Stresses at “Evolution Canyon”. Theor. Popul. Biol. 1997, 52, 231–243. [Google Scholar] [CrossRef]
- Nevo, E. Evolution of Genome–Phenome Diversity under Environmental Stress. Proc. Natl. Acad. Sci. USA 2001, 98, 6233–6240. [Google Scholar] [CrossRef]
- Finkel, M.; Fragman, O.; Nevo, E. Biodiversity and Interslope Divergence of Vascular Plants Caused by Sharp Microclimatic Differences at “Evolution Canyon II”, Lower Nahal Keziv, Upper Galilee, Israel. Isr. J. Plant Sci. 2001, 49, 285–296. [Google Scholar] [CrossRef]
- Finkel, M.; Chikatunov, V.; Nevo, E. Coleoptera of Evolution Canyon II: Lower Nahal Keziv, Western Upper Galilee, Israel; Series Faunistica; Pensoft: Sofia, Bulgaria; Moscow, Russia, 2002. [Google Scholar]
- Finkel, M.; Ofer, J.; Beharav, A.; Nevo, E. Species Interslope Divergence of Ants Caused by Sharp Microclimatic Stresses at ‘Evolution Canyon’ II, Lower Nahal Keziv, Western Upper Galilee, Israel. Isr. J. Entomol. 2015, 44–45, 63–73. [Google Scholar]
- Grishkan, I.; Nevo, E.; Wasser, S.P.; Beharav, A. Adaptive Spatiotemporal Distribution of Soil Microfungi in ‘Evolution Canyon’ II, Lower Nahal Keziv, Western Upper Galilee, Israel: Soil Fungi Distribution in ‘Evolution Canyon’ II, Israel. Biol. J. Linn. Soc. 2003, 78, 527–539. [Google Scholar] [CrossRef]
- Nevo, E. “Evolution Canyon,” a Potential Microscale Monitor of Global Warming across Life. Proc. Natl. Acad. Sci. USA 2012, 109, 2960–2965. [Google Scholar] [CrossRef] [PubMed]
- Por, F.D. An Outline of the Zoogeography of the Levant1. Zool. Scr. 1975, 4, 5–20. [Google Scholar] [CrossRef]
- Yom-Tov, Y.; Tchernov, E. The Zoogeography of Israel: The Distribution and Abundance at a Zoogeographical Crossroad; Monographiae Biologicae; Springer: Dordrecht, The Netherlands, 1988. [Google Scholar]
- Greenstone, M.H. Determinants of Web Spider Species Diversity: Vegetation Structural Diversity vs. Prey Availability. Oecologia 1984, 62, 299–304. [Google Scholar] [CrossRef]
- Halaj, J.; Ross, D.; Moldenke, A. Habitat Structure and Prey Availabilty as Predictors of the Abundance Community Organization of Spiders in Western Oregon Forest Canopies. J. Arachnol. 1998, 26, 203–220. [Google Scholar]
- Košulič, O.; Michalko, R.; Hula, V. Impact of Canopy Openness on Spider Communities: Implications for Conservation Management of Formerly Coppiced Oak Forests. PLoS ONE 2016, 11, e0148585. [Google Scholar] [CrossRef]
- Siemann, E.; Tilman, D.; Haarstad, J.; Ritchie, M. Experimental Tests of the Dependence of Arthropod Diversity on Plant Diversity. Am. Nat. 1998, 152, 738–750. [Google Scholar] [CrossRef]
- Spears, L.R.; MacMahon, J.A. An Experimental Study of Spiders in a Shrub-Steppe Ecosystem: The Effects of Prey Availability and Shrub Architecture. J. Arachnol. 2012, 40, 218–227. [Google Scholar] [CrossRef]
- Cardoso, P.; Silva, I.; De Oliveira, N.G.; Serrano, A.R.M. Seasonality of Spiders (Araneae) in Mediterranean Ecosystems and Its Implications in the Optimum Sampling Period. Ecol. Entomol. 2007, 32, 516–526. [Google Scholar] [CrossRef]
- Chatzaki, M.; Trichas, A.; Markakis, G.; Mylonas, M. Seasonal Activity of the Ground Spider Fauna in a Mediterranean Ecosystem (Mt Youchtas, Crete, Greece). In Proceedings of the 17th European Colloquium of Arachnology, Edinburgh, UK, 14–18 July 1997; British Arachnological Society, Burnham Beeches: Buckinghamshire, UK, 1998. [Google Scholar]
- Ganem, Z.; Ferrante, M.; Lubin, Y.; Armiach Steinpress, I.; Gish, M.; Sharon, R.; Harari, A.R.; Keasar, T.; Gavish-Regev, E. Effects of Natural Habitat and Season on Cursorial Spider Assemblages in Mediterranean Vineyards. Insects 2023, 14, 782. [Google Scholar] [CrossRef]
- Gavish-Regev, E.; Lubin, Y.; Coll, M. Migration Patterns and Functional Groups of Spiders in a Desert Agroecosystem. Ecol. Entomol. 2008, 33, 202–212. [Google Scholar] [CrossRef]
- Malumbres-Olarte, J.; Crespo, L.C.; Domènech, M.; Cardoso, P.; Moya-Laraño, J.; Ribera, C.; Arnedo, M.A. How Iberian Are We? Mediterranean Climate Determines Structure and Endemicity of Spider Communities in Iberian Oak Forests. Biodivers. Conserv. 2020, 29, 3973–3996. [Google Scholar] [CrossRef]
- Pitta, E.; Zografou, K.; Poursanidis, D.; Chatzaki, M. Effects of Climate on Spider Beta Diversity across Different Mediterranean Habitat Types. Biodivers. Conserv. 2019, 28, 3971–3988. [Google Scholar] [CrossRef]
- Zografou, K.; Adamidis, G.C.; Komnenov, M.; Kati, V.; Sotirakopoulos, P.; Pitta, E.; Chatzaki, M. Diversity of Spiders and Orthopterans Respond to Intra-Seasonal and Spatial Environmental Changes. J. Insect Conserv. 2017, 21, 531–543. [Google Scholar] [CrossRef]
- Pozsgai, G.; Lhoumeau, S.; Amorim, I.R.; Boieiro, M.; Cardoso, P.; Costa, R.; Ferreira, M.T.; Leite, A.; Malumbres-Olarte, J.; Oyarzabal, G.; et al. The BALA Project: A Pioneering Monitoring of Azorean Forest Invertebrates over Two Decades (1999–2022). Sci. Data 2024, 11, 368. [Google Scholar] [CrossRef]
- Armiach, I.; Bernstein, I.; Tang, Y.; Dayan, T.; Gavish-Regev, E. Activity-Density Data Reveal Community Structure of Lycosidae at a Mediterranean Shrubland. Aramit 2016, 52, 16–24. [Google Scholar] [CrossRef]
- Lubin, Y.; Ferrante, M.; Musli, I.; Lövei, G.L. Diversity of Ground-Active Spiders in Negev Desert Habitats, Israel. J. Arid. Environ. 2020, 183, 104252. [Google Scholar] [CrossRef]
- Lubin, Y.; Angel, N.; Assaf, N. Ground Spider Communities in Experimentally Disturbed Mediterranean Woodland Habitats. Aramit 2011, 40, 85–93. [Google Scholar] [CrossRef]
- Whitehouse, M.E.A.; Shochat, E.; Shachak, M.; Lubin, Y. The Influence of Scale and Patchiness on Spider Diversity in a Semi-Arid Environment. Ecography 2002, 25, 395–404. [Google Scholar] [CrossRef]
- Sneh, A.; Bartov, Y.; Weissbrod, T.; Rosensaft, M. Geological Map of Israel 1998. Available online: https://www.gov.il/he/pages/israel-map-1-200k (accessed on 20 August 2024).
- Israel Meteorological Service Website. Available online: https://ims.gov.il/he/data_gov (accessed on 20 August 2024).
- Sharon, D. Areal Pattern of Rainfall in a Small Watershed as Affected by Wind and Other Meteorological Conditions. In Results of Research on Experimental Basins; International Association of Hydrology Sciences Publication, 1970; pp. 3–11. Available online: https://eurekamag.com/research/018/258/018258119.php (accessed on 20 August 2024).
- Sharon, D. The Distribution of Hydrologically Effective Rainfall Incident on Sloping Ground. J. Hydrol. 1980, 46, 165–188. [Google Scholar] [CrossRef]
- Sharon, D.; Adar, E.; Lieberman, G. Observations on the Differential Hydrological and/or Erosional Response of Opposite-Lying Slopes, as Related to Incident Rainfall. Isr. J. Earth Sci. 1983, 32, 71–74. [Google Scholar]
- Sharon, D.; Arazi, A. The Distribution of Wind-Driven Rainfall in a Small Valley: An Empirical Basis for Numerical Model Verification. J. Hydrol. 1997, 201, 21–48. [Google Scholar] [CrossRef]
- Cottle, H.J. Vegetation on North and South Slopes of Mountains in Southwestern Texas. Ecology 1932, 13, 121–134. [Google Scholar] [CrossRef]
- Stephenson, S.L. Exposure-Induced Differences in the Vegetation, Soils, and Microclimate of North- and South-Facing Slopes in Southwestern Virginia. Va. J. Sci. 1982, 33, 36–50. [Google Scholar]
- Palamarev, E. Paleobotanical Evidences of the Tertiary History and Origin of the Mediterranean Sclerophyll Dendroflora. Pl Syst Evol. 1989, 162, 93–107. [Google Scholar] [CrossRef]
- Levy, G. Crab-Spiders of Six Genera from Israel (Araneae: Thomisidae). Isr. J. Zool. 1973, 22, 107–141. [Google Scholar] [CrossRef]
- Levy, G. Araneae: Thomisidae; Fauna Palaestina Arachnida; The Israel Academy of Sciences and Humanities: Jerusalem, Israel, 1985; ISBN 978-965-208-058-5. [Google Scholar]
- Levy, G. The Family of Huntsman Spiders in Israel with Annotations on Species of the Middle East (Araneae: Sparassidae). J. Zool. 1989, 217, 127–176. [Google Scholar] [CrossRef]
- Levy, G. Spiders of the Genus Lachesana and a New Storenoid Genus from Israel (Araneae: Zodariidae). Zool. J. Linn. Soc. 1990, 98, 327–362. [Google Scholar] [CrossRef]
- Levy, G. The Spider Genera Palaestina, Trygetus, Zodarion and Ranops (Araneae, Zodariidae) in Israel with Annotations on Species of the Middle East. Isr. J. Zool. 1992, 38, 67–110. [Google Scholar]
- Levy, G. Revision of the Spider Subfamily Gnaphosinae in Israel (Araneae: Gnaphosidae). J. Nat. Hist. 1995, 29, 919–981. [Google Scholar] [CrossRef]
- Levy, G. The Agelenid Funnel-Weaver Family and the Spider Genus Cedicus in Israel (Araneae, Agelenidae and Cybaeidae). Zool. Scr. 1996, 25, 85–122. [Google Scholar] [CrossRef]
- Levy, G. Araneae Theridiidae; The Israel Academy of Sciences and Humanities: Jerusalem, Israel, 1998; ISBN 978-965-208-013-4. [Google Scholar]
- Levy, G. New Thomisid and Philodromid Spiders (Araneae) from Southern Israel. Bull. Br. Arachnol. Soc. 1999, 11, 185–190. [Google Scholar]
- Levy, G. Spiders of the Genera Anagraphis and Talanites (Araneae, Gnaphosidae) from Israel. Isr. J. Zool. 1999, 45, 215–223. [Google Scholar] [CrossRef]
- Levy, G. Spiders of Six Uncommon Drassodine Genera (Araneae: Gnaphosidae) from Israel. Isr. J. Zool. 1999, 45, 427–452. [Google Scholar] [CrossRef]
- Levy, G. Spiders of the Families Anyphaenidae, Hahniidae, Ctenidae, Zoridae, and Hersiliidae (Araneae) from Israel. Isr. J. Zool. 2003, 49, 1–31. [Google Scholar] [CrossRef]
- Levy, G. Spiders of the Genera Drassodes and Haplodrassus (Araneae, Gnaphosidae) from Israel. Isr. J. Zool. 2004, 50, 1–37. [Google Scholar] [CrossRef]
- Levy, G. Calommata (Atypidae) and New Spider Species (Araneae) from Israel. Zootaxa 2007, 1551, 1–30. [Google Scholar] [CrossRef]
- Levy, G. New Ground-Spider Genera and Species with Annexed Checklist of the Gnaphosidae (Araneae) of Israel. Zootaxa 2009, 2066, 1–49. [Google Scholar] [CrossRef]
- Magalhaes, I.L.F.; Aharon, S.; Ganem, Z.; Gavish-Regev, E. A New Semi-Cryptic Filistata from Caves in the Levant with Comments on the Limits of Filistata Insidiatrix (Forsskål, 1775) (Arachnida: Araneae: Filistatidae). EJT 2022, 831, 149–174. [Google Scholar] [CrossRef]
- Nentwig, W.; Blick, T.; Bosmans, R.; Gloor, D.; Hänggi, A.; Kropf, C. Spiders of Europe. Version 08.2023. Available online: https://araneae.nmbe.ch/ (accessed on 11 August 2023).
- Roberts, M. Collins Field Guide to the Spiders of Britain and Northern Europe; Harper Collins: London, UK, 1996. [Google Scholar]
- Tanasevitch, A. On Linyphiid Spiders (Araneae) from the Eastern and Central Mediterranean Kept at the Museum d’histoire Naturelle, Geneva. Rev. Suisse Zool. 2011, 118, 49–91. [Google Scholar] [CrossRef]
- Levy, G.; Amitai, P. Scorpiones; Fauna Palaestina; Israel Academy of Sciences and Humanities: Jerusalem, Israel, 1980; ISBN 978-965-208-016-5. [Google Scholar]
- Yahalomi, O.; Zvik, Y.; Tsoar, A.; Gavish-Regev, E. The Scorpions of Israel Field Guide, 1st ed.; Israel Nature Protection Authority: Jerusalem, Israel, 2018. [Google Scholar]
- Levy, G.; Shulov, A. The Solifuga of Israel. Isr. J. Zool. 1964, 13, 102–120. [Google Scholar] [CrossRef]
- Warburg, S.; Aharon, S.; Armiach Steinpress, I.; Sharma, P.P.; Harms, D.; Gavish-Regev, E. Pseudoscorpions of Israel: Annotated Checklist and Key, With New Records of Two Families (Arachnida: Pseudoscorpiones). Taxonomy 2023, 3, 466–495. [Google Scholar] [CrossRef]
- ter Braak, C.; Smilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination (Version 5.10); Microcomputer Power: Ithaca, NY, USA, 2018. [Google Scholar]
- ter Braak, C.; Smilauer, P. Canoco—Software for Ordination 2020.
- Lepš, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using CANOCO, 1st ed.; Cambridge University Press: Cambridge, UK, 2003; ISBN 978-0-521-81409-6. [Google Scholar]
- Jesussek, M.; Volk-Jesussek, H. Statisty. Available online: https://statisty.app/ (accessed on 20 August 2024).
- Magalhaes, I.L.F. Spreadsheets to Expedite Taxonomic Publications by Automatic Generation of Morphological Descriptions and Specimen Lists. Zootaxa 2019, 4624, 147–150. [Google Scholar] [CrossRef]
- Logunov, D.V. Taxonomic-faunistic notes on the jumping spiders of the Mediterranean (Aranei: Salticidae). Arthropoda Sel. 2015, 24, 33–85. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- World Spider Catalog Version 25. Available online: http://wsc.nmbe.ch (accessed on 25 June 2024).
- Natural History Museum Bern World Solifugae Catalog. Available online: http://wac.nmbe.ch (accessed on 25 June 2024).
- Cardoso, P.; Pekár, S.; Jocqué, R.; Coddington, J.A. Global Patterns of Guild Composition and Functional Diversity of Spiders. PLoS ONE 2011, 6, e21710. [Google Scholar] [CrossRef] [PubMed]
- Prószyński, J. Atlas Rysunków Diagnostycznych Mniej Znanych Salticidae 2; Zeszyty Naukowe Wyższej Szkoly Rolniczo-Pedagogicznej: Siedlcach, Poland, 1987. [Google Scholar]
- Prószyński, J. Salticidae (Araneae) of the Levant. Ann. Zool. Warszawa 2003, 53, 1–180. [Google Scholar]
- Simon, E. Monographie Des Espèces Européennes de La Famille Des Attides (Attidae Sundewall.—Saltigradae Latreille). Ann. Société Entomol. Fr. 1868, 4, 11–72. [Google Scholar]
- Metzner, H. Jumping Spiders (Arachnida: Araneae: Salticidae) of the World. Available online: https://www.jumping-spiders.com (accessed on 29 June 2024).
- Metzner, H. Die Springspinnen (Araneae, Salticidae) Griechenlands. Andrias 1999, 14, 1–279. [Google Scholar]
- Lecigne, S. A New Species of Sintula (Linyphiidae), Redescription of Brigittea Iinocens (Dictynidae) and Eight Spider Species Newly Recorded for Turkey (Araneae). Arachnol. Mitteilungen Arachnol. Lett. 2021, 62, 11–34. [Google Scholar] [CrossRef]
- Prószyński, J. On Mostly New Species of Salticidae (Aranei) from Levant. Arthropoda Sel. 1999, 8, 231–262. [Google Scholar]
- Oger, P. Les Araignées de Belgique et de France. Available online: https://arachno.piwigo.com/ (accessed on 20 August 2024).
- Bosmans, R.; Gavalas, I. The Spiders (Araneae) of the Tiny Greek Island Iraklia (Kiklades), with the Descriptions of 5 New Harpactea Species (Araneae: Dysderidae), 3 Species New to Europe and 8 New to Greece. Parnass. Arch. 2023, 11 (Suppl. 1), 1–91. [Google Scholar]
- Thaler, K.; Knoflach, B. A New Lachesana Species from Greece and Additional Records from the Near East (Arachnida, Araneae, Zodariidae). Bull. Br. Arachnol. Soc. 2004, 13, 41–46. [Google Scholar]
- Simon, E. Histoire Naturelle des Araignées; Roret: Paris, France, 1892. [Google Scholar]
- Pickard-Cambridge, O. General List of the Spiders of Palestine and Syria, with Descriptions of Numerous New Species, and Characters of Two New Genera. Proc. Zool. Soc. Lond. 1872, 40, 183–354. [Google Scholar] [CrossRef]
- Zamani, A.; Marusik, Y.M. Revision of the Spider Family Zodariidae (Arachnida, Araneae) in Iran and Turkmenistan, with Seventeen New Species. ZooKeys 2021, 1035, 145–193. [Google Scholar] [CrossRef] [PubMed]
- Barriga, J.C.; Lassaletta, L.; Moreno, A.G. Ground-Living Spider Assemblages from Mediterranean Habitats under Different Management Conditions. J. Arachnol. 2010, 38, 258–269. [Google Scholar] [CrossRef]
- Lin, S.; Vasseur, L.; You, M.-S. Seasonal Variability in Spider Assemblages in Traditional and Transgenic Rice Fields. Environ. Entomol. 2016, 45, 537–546. [Google Scholar] [CrossRef]
- Zapata, W.; Vergara-Moreno, D.; Carrillo-Pallares, M.; Segovia-Paccini, A.; Navas, S.G.R.; Malumbres-Olarte, J. Seasonal Diversity of Spider Assemblages (Araneae: Arachnida) in the “Guillermo Piñeres” Botanical Garden, Turbaco–Colombia. Neotrop. Biodivers. 2023, 9, 17–28. [Google Scholar] [CrossRef]
- Gillespie, R.G.; Bennett, G.M.; De Meester, L.; Feder, J.L.; Fleischer, R.C.; Harmon, L.J.; Hendry, A.P.; Knope, M.L.; Mallet, J.; Martin, C.; et al. Comparing Adaptive Radiations Across Space, Time, and Taxa. J. Hered. 2020, 111, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Rolán-Alvarez, E. Sympatric Speciation as a By-Product of Ecological Adaptation in the Galician Littorina Saxatilis Hybrid Zone. J. Molluscan Stud. 2007, 73, 1–10. [Google Scholar] [CrossRef]
- Muma, M.H. The Life Cycle of Eremobates durangonus (Arachnida: Solpugida). Fla. Entomol. 1966, 49, 233–242. [Google Scholar] [CrossRef]
- Junqua, C. Recherches Biologiques et Histophysiologiques Sur Un Solifuge Saharien Othoes Saharae Panouse. In Me’moires du Muse´um National d’Histoire Naturelle, Se´ries A; Muséum national d’Histoire naturelle: Paris, France, 1966; pp. 1–124. [Google Scholar]
- Punzo, F. Natural History and Life Cycle of the Solifuge Eremobates marathoni Muma & Brookhart (Solifugae, Eremobatidae). Bull. Br. Arachnol. Soc. 1998, 11, 111–118. [Google Scholar]
- Jones, R.R.; Batista-Perales, D.L.; Garcia, E.L. Home on the Range: A Pilot Study on Solifuge (Solifugae: Eremobatidae) Site Fidelity at Rocky Mountain Arsenal National Wildlife Refuge. J. Arachnol. 2022, 50, 47–50. [Google Scholar] [CrossRef]
- Hrušková-Martišová, M.; Pekár, S.; Bilde, T. Coercive Copulation in Two Sexually Cannibalistic Camel-spider Species (Arachnida: Solifugae). J. Zool. 2010, 282, 91–99. [Google Scholar] [CrossRef]






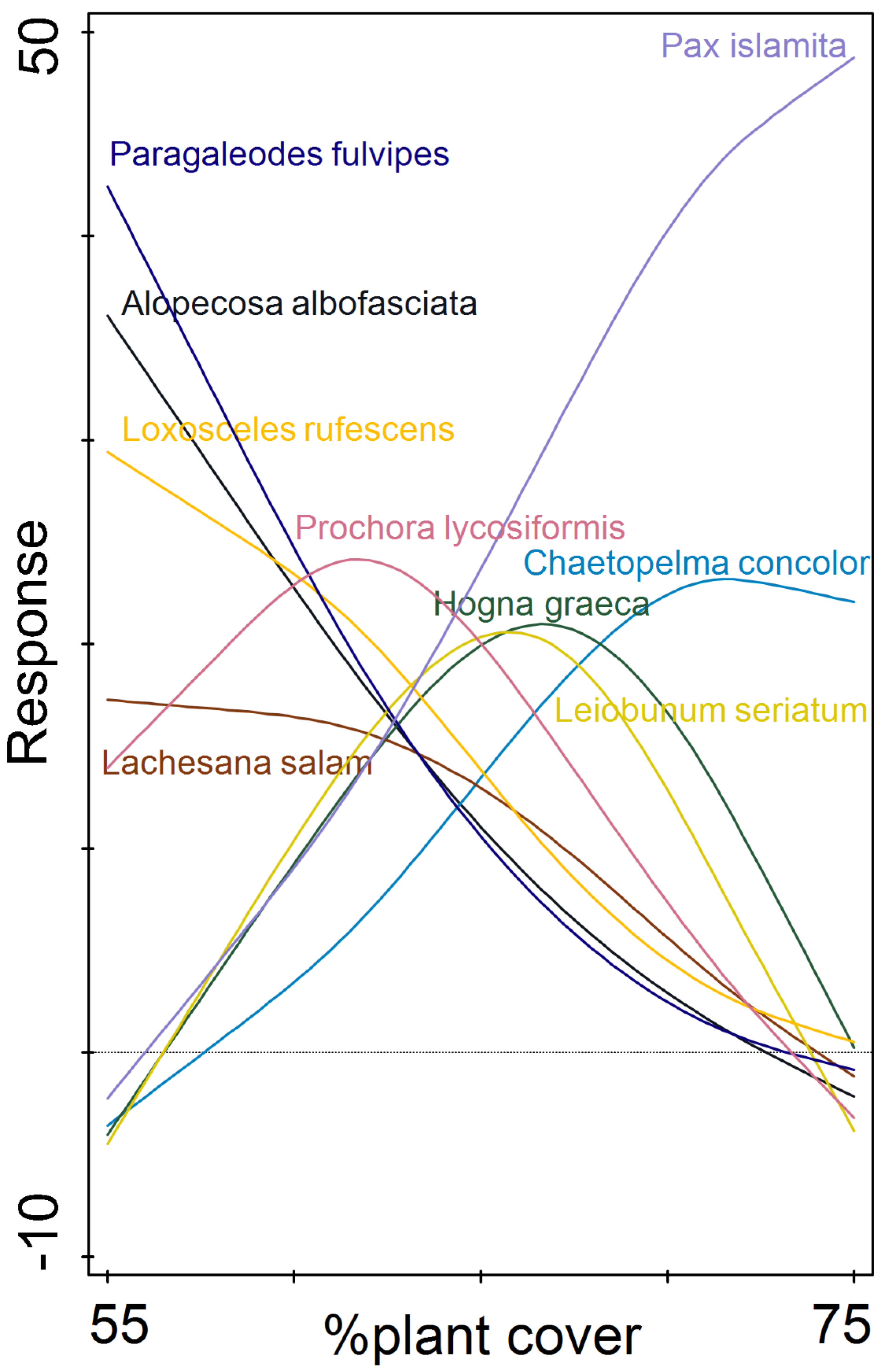
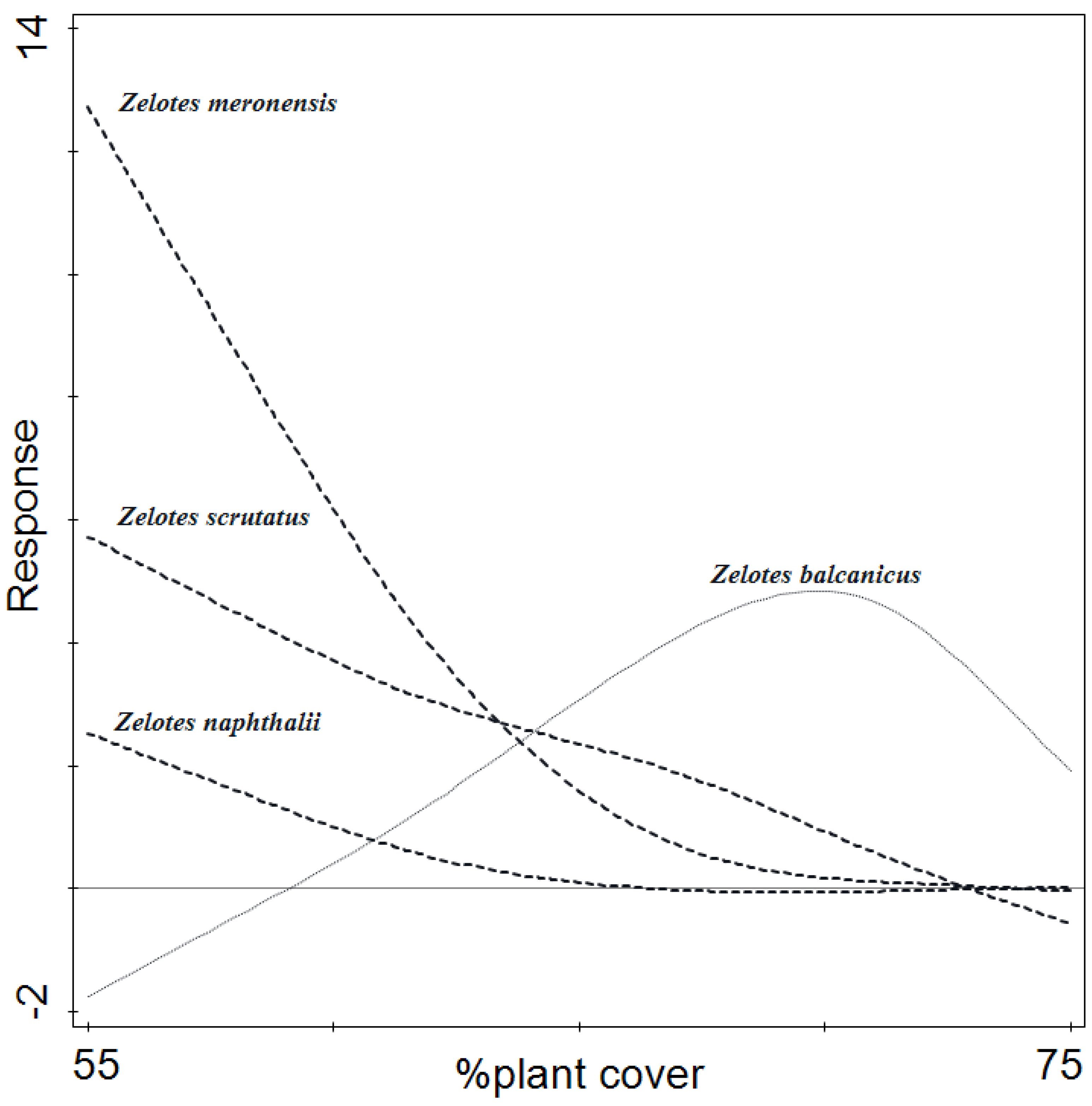

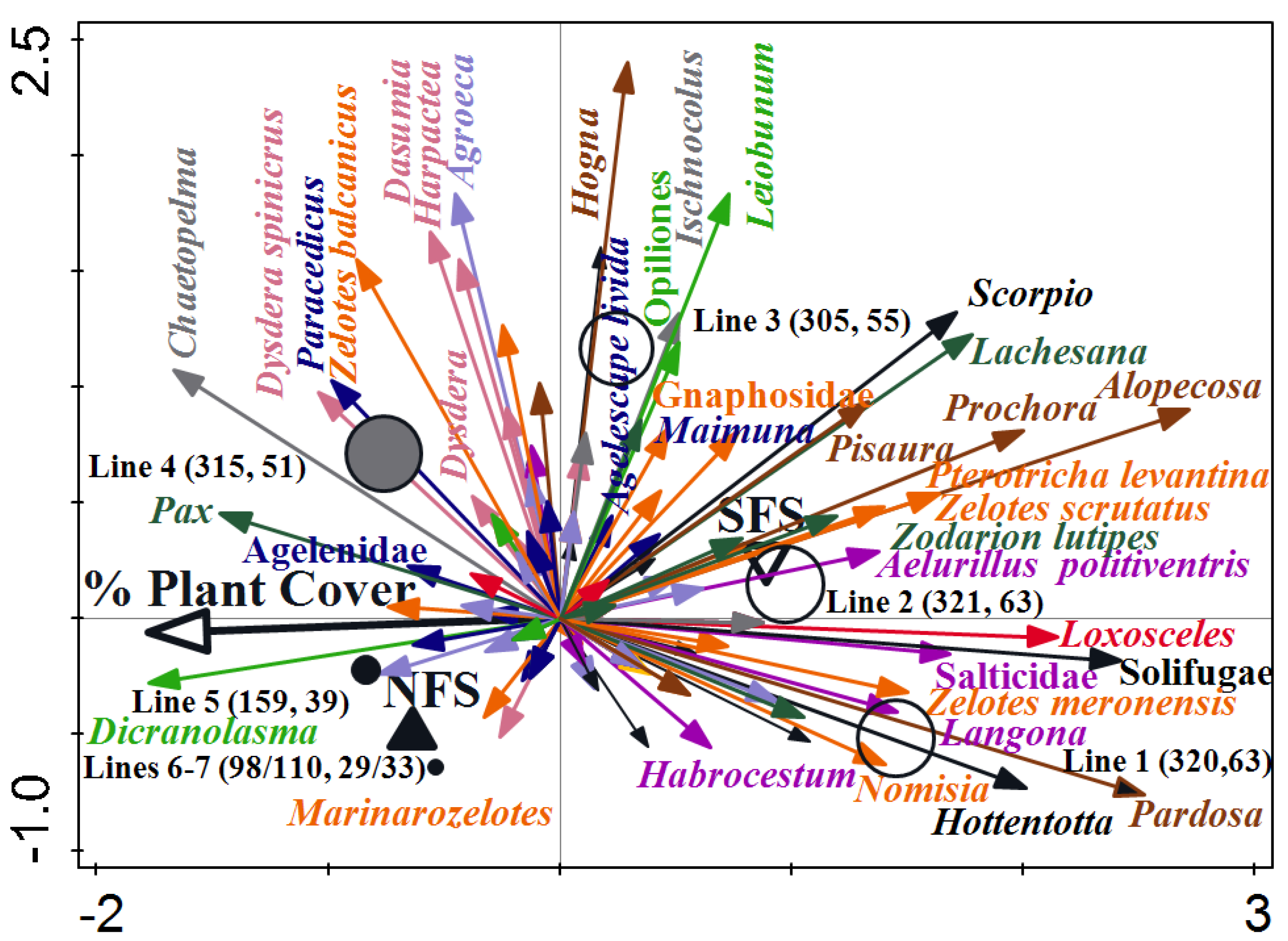



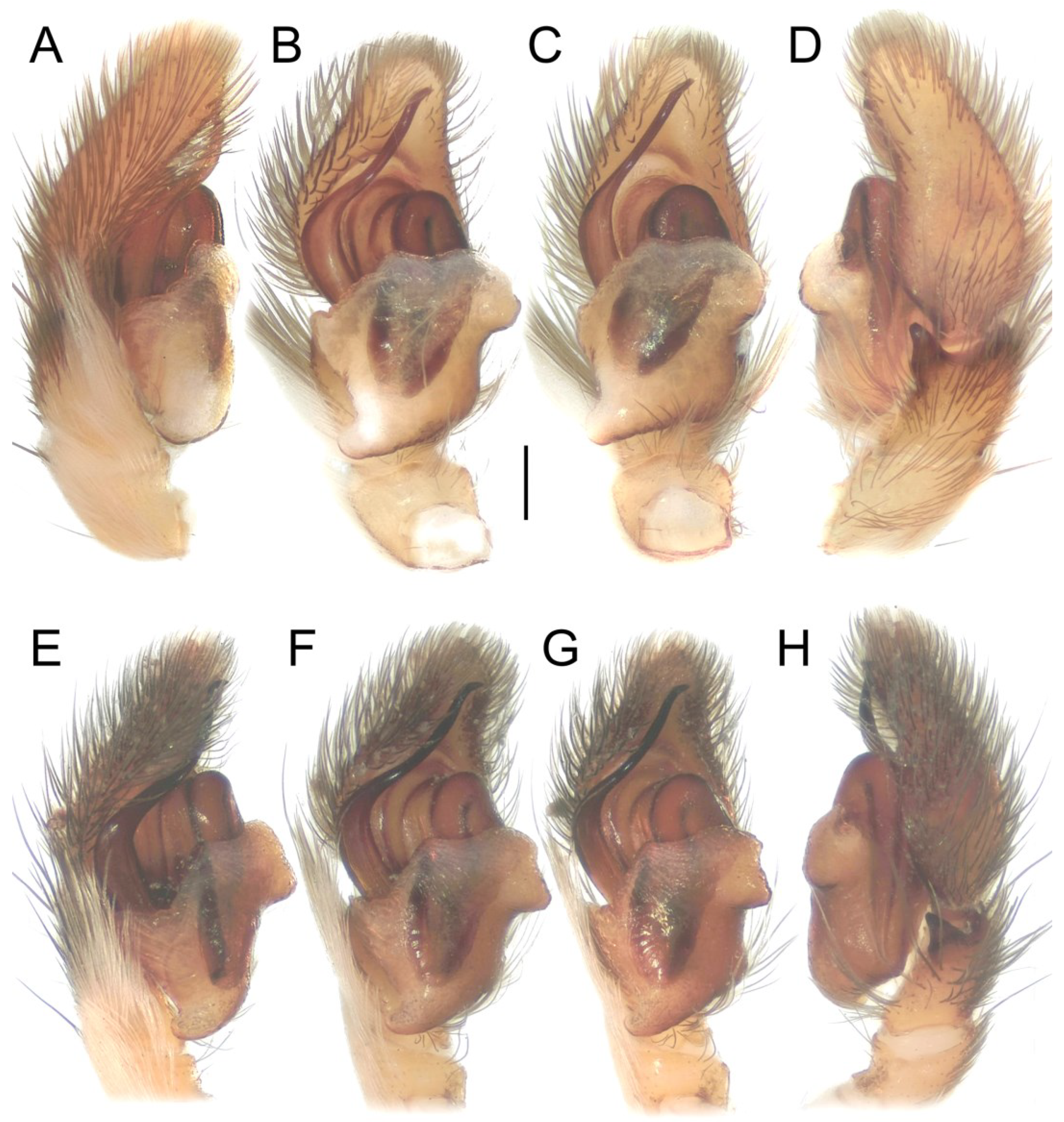
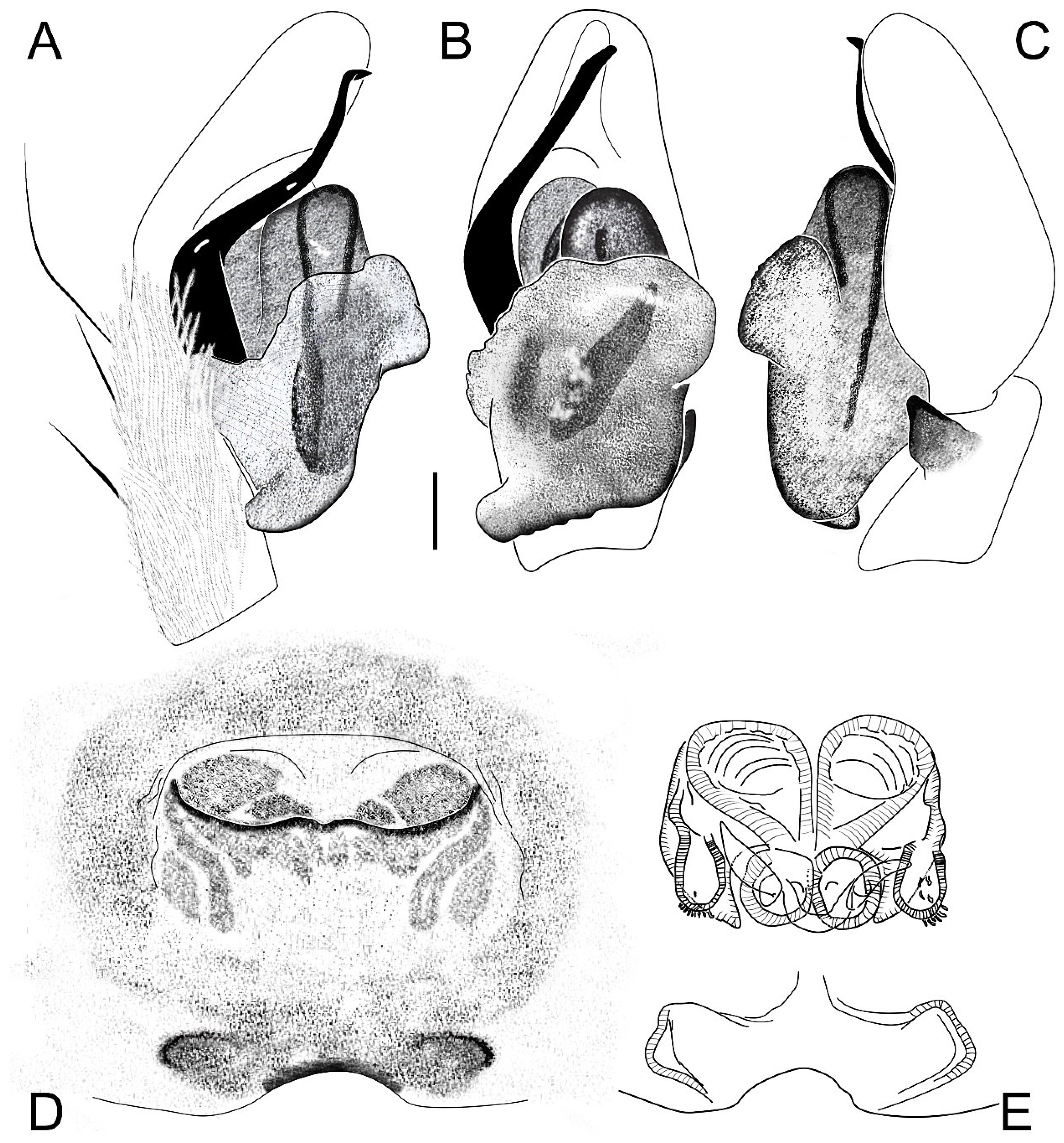


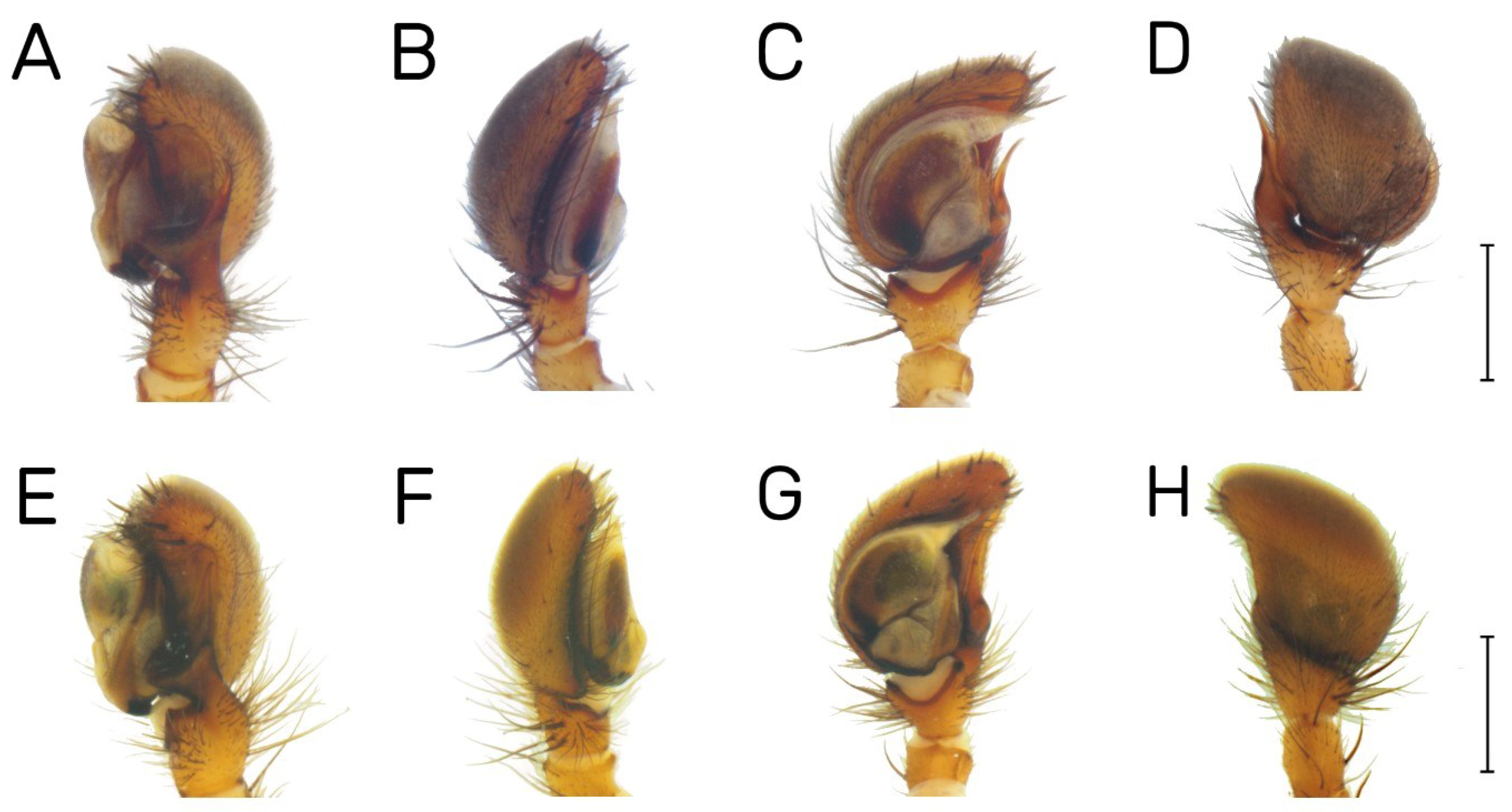
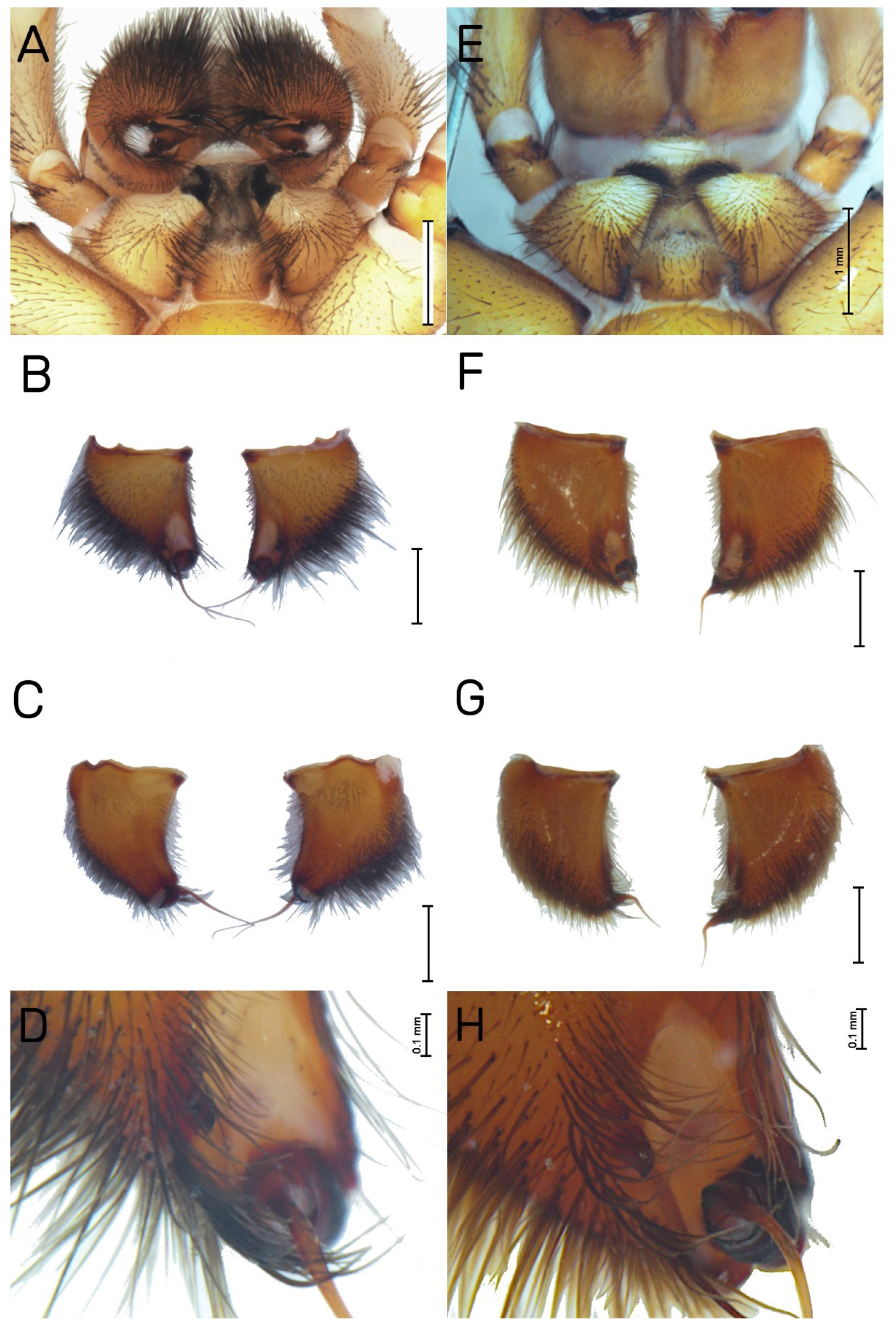
| Habitat | Number of Traps | Number of Individuals | |||
|---|---|---|---|---|---|
| Minimum | Maximum | Median | Mean | ||
| SFS | 30 | 4 | 85 | 35 | 35 |
| VB | 10 | 13 | 62 | 30 | 32.3 |
| NFS | 30 | 4 | 34 | 12 | 12.5 |
| SFS: Under tree canopy | 7 | 4 | 50 | 36 | 34 |
| SFS: Under shrub | 9 | 9 | 45 | 32 | 31.67 |
| SFS: Grassland in open patches | 14 | 9 | 85 | 35 | 37.64 |
| Holo/Para | AME | ALE | PME | PLE | AME-AME | AME-PME | PME-PME |
|---|---|---|---|---|---|---|---|
| HOLO | 0.18 | 0.17 | 0.18 | 0.13 | 0.09 | 0.16 | 0.16 |
| PARA | 0.17 | 0.16 | 18 | 0.14 | 0.08 | 0.11 | 0.13 |
| leg | Femur | Patella | Tibia | Metatarsus | Tarsus | Total |
|---|---|---|---|---|---|---|
| I | 4d, 2-2 (r-l) la | 1 (r) la | 1 (r) la, 10ve | 8ve | 11ve | 39 |
| II | 4d, 4-4 (r-l) la | 1 (r) la | 2 (r) la, 8ve | 1 (r) la, 9ve | 9ve | 42 |
| III | 4d, 3-2 (r-l) la | 1d, 2-1 (r-l) la | 3d, 3-2 (r-l) la, 6ve | 1d, 4-3 (r-l) la, 13ve | 2 (r) la, 10ve | 60 |
| IV | 4d, 3-2(r-l) la | 1d, 2-1 (r-l) la | 3d, 3-2 (r-l) la, 10ve | 1d, 4-3 (r-l) la, 13ve | 11ve | 63 |
| leg | Femur | Patella | Tibia | Metatarsus | Tarsus | Total |
|---|---|---|---|---|---|---|
| I | 4.32/4.06 | 1.79/1.78 | 3.3/3.04 | 3.29/3.16 | 2.3/2.09 | 15.00/14.13 |
| II | 4.04/3.62 | 1.81/1.74 | 2.77/2.69 | 3.00/3.23 | 2.10/2.04 | 13.72/13.32 |
| III | 3.87/3.61 | 1.78/1.64 | 2.15/2.18 | 4.17/3.83 | 2.44/2.12 | 14.41/13.38 |
| IV | 4.52/4.38 | 1.83/1.89 | 2.88/2.98 | 5.56/5.47 | 2.7/2.58 | 17.49/17.30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Finkel, M.; Ben-Asher, A.; Shmula, G.; Armiach Steinpress, I.; Ganem, Z.; Hammouri, R.; Garcia, E.; Szűts, T.; Gavish-Regev, E. Arachnid Assemblage Composition Diverge between South- and North-Facing Slopes in a Levantine Microgeographic Site. Diversity 2024, 16, 540. https://doi.org/10.3390/d16090540
Finkel M, Ben-Asher A, Shmula G, Armiach Steinpress I, Ganem Z, Hammouri R, Garcia E, Szűts T, Gavish-Regev E. Arachnid Assemblage Composition Diverge between South- and North-Facing Slopes in a Levantine Microgeographic Site. Diversity. 2024; 16(9):540. https://doi.org/10.3390/d16090540
Chicago/Turabian StyleFinkel, Meir, Amit Ben-Asher, Gur Shmula, Igor Armiach Steinpress, Zeana Ganem, Rami Hammouri, Erika Garcia, Tamás Szűts, and Efrat Gavish-Regev. 2024. "Arachnid Assemblage Composition Diverge between South- and North-Facing Slopes in a Levantine Microgeographic Site" Diversity 16, no. 9: 540. https://doi.org/10.3390/d16090540
APA StyleFinkel, M., Ben-Asher, A., Shmula, G., Armiach Steinpress, I., Ganem, Z., Hammouri, R., Garcia, E., Szűts, T., & Gavish-Regev, E. (2024). Arachnid Assemblage Composition Diverge between South- and North-Facing Slopes in a Levantine Microgeographic Site. Diversity, 16(9), 540. https://doi.org/10.3390/d16090540