
Biodiversity and Possible Bio-Indicators of Mediterranean Temporary Ponds in Southern Apulia, Italy



Abstract
:1. Introduction
2. Material and Methods
2.1. Study Area
2.2. Field Surveys
2.3. Floristic Data
2.4. Habitat MTP 3170* Data
2.5. Aquatic Faunal Data
2.6. Data Analysis
3. Results
3.1. Abiotic Characteristics of the Water
3.2. Flora
3.3. Aquatic Fauna
3.4. Habitat MTP 3170* Conservation Status
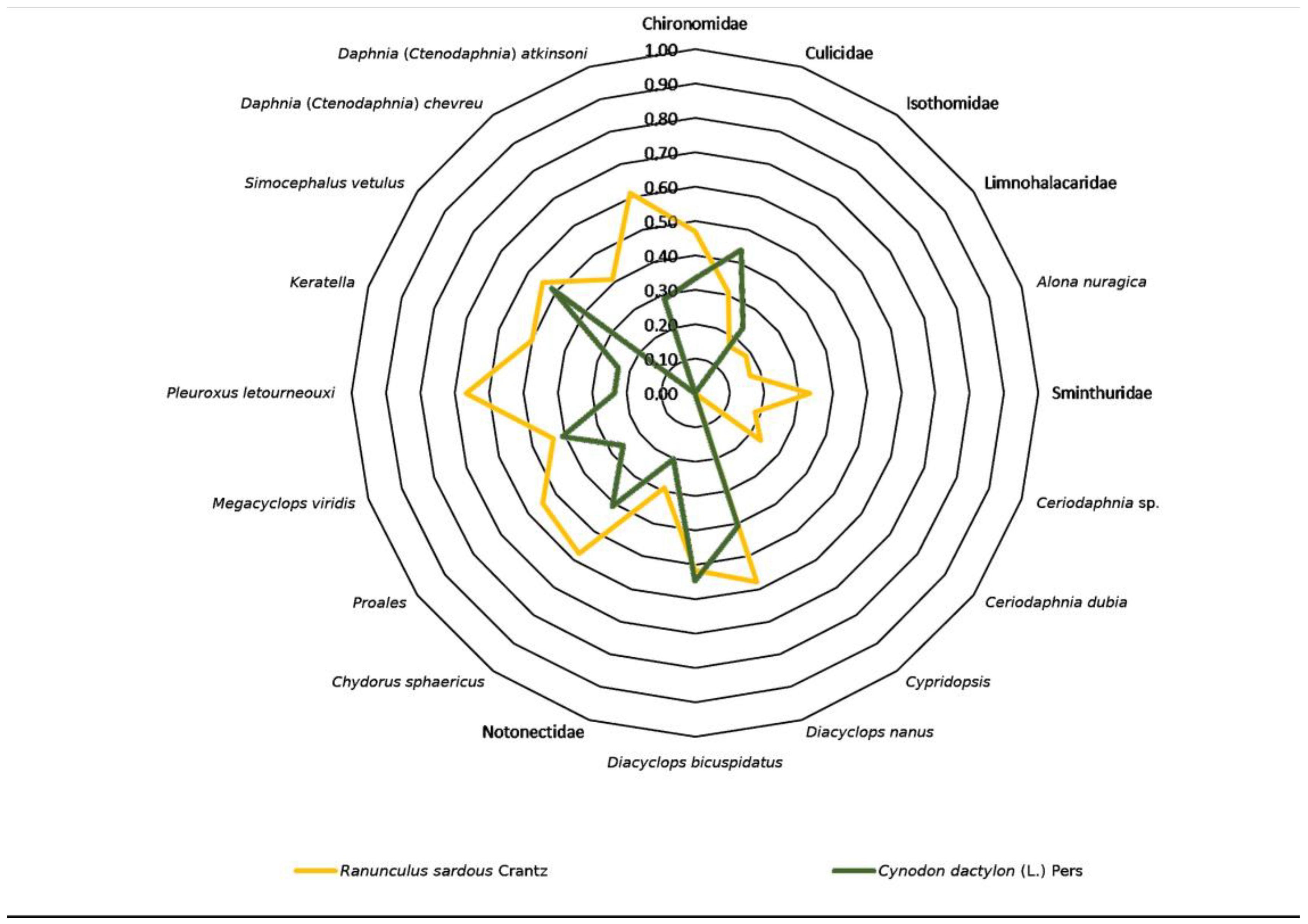
3.5. Flora–Fauna Relationships
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Oertli, B.; Céréghino, R.; Hull, A.; Miracle, M.R. Pond conservation: From science to practice. Hydrobiologia 2009, 634, 157–165. [Google Scholar] [CrossRef]
- Oertli, B.; Joye, D.A.; Castella, E.; Juge, R.; Cambin, D.; Lachavanne, J.B. Does size matter? The relationship between pond area and biodiversity. Biol. Conserv. 2002, 104, 59–70. [Google Scholar] [CrossRef]
- Williams, P.; Whitfield, M.; Biggs, J.; Bray, S.; Fox, G.; Nicolet, P.; Sear, D. Comparative biodiversity of rivers streams, ditches and ponds in an agricultural landscape in Southern England. Biol. Conserv. 2004, 115, 329–341. [Google Scholar] [CrossRef]
- Grillas, P.; Gauthier, P.; Yavercovski, N.; Perennou, C. Mediterranean Temporary Pools. Volume 1–Enjeux de Conservation, Fonctionnement et Gestion; Station biologique de la Tour du Valat: Arles, France, 2004; 121p, ISBN 2-9103-6849-1. [Google Scholar]
- Williams, D.D. The Biology of Temporary Waters; Oxford University Press: Oxford, UK, 2006; 352p. [Google Scholar]
- Alfonso, G.; Beccarisi, L.; Pieri, V.; Frassanito, A.; Belmonte, G. Using crustaceans to identify different pond types. A case study from the Alta Murgia National Park, Apulia (South-eastern Italy). Hydrobiologia 2016, 782, 53–69. [Google Scholar] [CrossRef]
- Bagella, S.; Gascón, S.; Filigheddu, R.; Cogoni, A.; Boix, D. Mediterranean Temporary Ponds: New challenges from a neglected habitat. Hydrobiologia 2016, 782, 1–10. [Google Scholar] [CrossRef]
- Oertli, B.; Joye, D.A.; Indermuehle, N.R.; Juge & Lacahvanne, J.B. lst European pond workshop “Conservation and Monitoring of Pond Biodiversity”. Arch. Des. Sci. 2004, 57, 69–71. [Google Scholar]
- Céreghino, R.; Biggs, J.; Oertli, B.; Declerck, S. The ecology of European ponds: Defining the characteristics of a neglected freshwater habitat. Hydrobiologia 2008, 597, 1–6. [Google Scholar] [CrossRef]
- EPCN The Pond Manifesto. 2008. Available online: http://camp,s.hesge.ch/epcn/projects.asp (accessed on 5 July 2024).
- Miracle, M.R.; Oertli, B.; Céréghino, R.; Hull, A. Preface: Conservation of European ponds-current knowledge and future needs. Limnetica 2010, 29, 1–8. [Google Scholar] [CrossRef]
- Boix, D.; Biggs, J.; Céréghino, R.; Hull, A.P.; Kalettka, T.; Oertli, B. Pond research and management in Europe: “Small is Beautiful”. Hydrobiologia 2012, 689, 1–9. [Google Scholar] [CrossRef]
- Brysiewicz, A.; Czerniejewski, P.; Bonisławska, M. Effect of Diverse Abiotic Conditions on the Structure and Biodiversity of Ichthyofauna in Small, Natural Water Bodies Located on Agricultural Lands. Water 2020, 12, 2674. [Google Scholar] [CrossRef]
- Sługocki, L.; Brysiewicz, A. Divergence of zooplankton assemblages in unstable fishless and stable fish ponds. Hydrobiologia 2024, 851, 3881–3896. [Google Scholar] [CrossRef]
- Mustafa, A.; Mudian, P.; Tarunamulia, T.; Kamariah, K.; Ratnawati, E.; Athirah, A.; Asaf, R.; Tahe, S.; Makmur, M.; Taukhid, I.; et al. Water quality changes in the coastal area of intensive whiteleg shrimp brackish water pond aquaculture. J. Water Land Dev. 2024, 61, 130–142. [Google Scholar]
- European Commission. Interpretation Manual of European Union Habitats; EUR 28; European Environment, Nature and Biodiversity; European Commission: Brussels, Belgium, 2013. [Google Scholar]
- Ruggiero, A.; Céréghino, R.; Figuerola, J.; Marty, P.; Angélibert, S. Farm ponds make a contribution to the biodiversity of aquatic insects in a French agricultural landscape. Comptes Rendus Biol. 2008, 331, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Bagella, S.; Gascón, S.; Caria, M.C.; Sala, J.; Boix, D. Cross-taxon congruence in Mediterranean temporary wetlands: Vascular plants, crustaceans, and coleopterans. Community Ecol. 2011, 12, 40–50. [Google Scholar] [CrossRef]
- Nicolet, P.; Biggs, J.; Fox, G.; Hodson, M.J.; Reynolds, C.; Whitfield, M.; Williams, P. The wetland plant and macroinvertebrate assemblages of temporary ponds in England and Wales. Biol. Conserv. 2004, 120, 265–282. [Google Scholar] [CrossRef]
- Angeler, D.G.; Viedma, O.; Cirujano, S.; Alvarez-Cobelas, M.; Sánchez-Carrillo, S. Microinvertebrate and plant beta diversity in dry soils of a semiarid agricultural wetland complex. Mar. Freshw. Res. 2008, 59, 418–428. [Google Scholar] [CrossRef]
- Landi, M.; Piazzini, S.; Nucci, A.; Saveri, C.; Angiolini, C. Can macrophytes be a surrogate for amphibians and physico-chemical features in pond classifications? Aquat. Bot. 2012, 101, 1–7. [Google Scholar] [CrossRef]
- Rouisii, M.; Boix, D.; Muller, S.D.; Gascón, S.; Ruhí, A.; Sala, J.; Bouattour, A.; Ben Haj Jilani, I.; Ghrabi-Gammar, Z.; Ben Saad-Limam, S.; et al. Spatio-temporal variability of faunal and floral assemblages in Mediterranean temporary wetlands. Comptes Rendus Biol. 2014, 337, 695–708. [Google Scholar] [CrossRef]
- Ruhí, A.; Chappuis, E.; Escoriza, D.; Jover, M.; Sala, J.; Boix, D.; Gascón, S.; Gacia, E. Environmental filtering determines community patterns in temporary wetlands: A multi-taxon approach. Hydrobiologia 2014, 723, 25–39. [Google Scholar] [CrossRef]
- Rundle, S.D.; Foggo, A.; Choiseul, V.; Bilton, D.T. Are distribution patterns linked to dispersal mechanism? An investigation using pond invertebrate assemblages. Freshw. Biol. 2002, 47, 1571–1581. [Google Scholar] [CrossRef]
- Eitam, A.; Blaustein, L.; Van Damme, K.; Dumont, H.J.; Martens, K. Crustacean species richness in temporary pools: Relationships with habitat traits. Hydrobiologia 2004, 525, 125–130. [Google Scholar] [CrossRef]
- Della Bella, V.; Bazzanti, M.; Chiarotti, F. Macroinvertebrate diversity and conservation status of Mediterranean ponds in Italy: Water permanence and mesohabitat influence. Aquat. Conserv. 2005, 15, 583–600. [Google Scholar] [CrossRef]
- Serrano, L.; Fahd, K. Zooplankton communities across a hydroperiod gradient of temporary ponds in the Donana National Park (SW Spain). Wetlands 2005, 25, 101–111. [Google Scholar] [CrossRef]
- Tavernini, S.; Mura, G.; Rossetti, G. Factors influencing the seasonal phenology and composition of zooplankton communities in mountain temporary pools. Int. Rev. Hydrobiol. 2005, 90, 358–375. [Google Scholar] [CrossRef]
- Waterkeyn, A.; Grillas, P.; Vanschoenwinkel, B.; Brendonck, L. Invertebrate community patterns in Mediterranean temporary wetlands along hydroperiod and salinity gradients. Freshw. Biol. 2008, 53, 1808–1822. [Google Scholar] [CrossRef]
- Florencio, M.; Serrano, L.; Gómez-Rodríguez, C.; Millán, A.; Díaz-Paniagua, C. Inter- and intra-annual variations of macroinvertebrate assemblages are related to the hydroperiod in Mediterranean temporary ponds. Hydrobiologia 2009, 634, 167–183. [Google Scholar] [CrossRef]
- Gilbert, J.D.; De Vicente, I.; Ortega, F.; Jiménez-Melero, R.; Parra, G.; Guerrero, F. A comprehensive evaluation of the crustacean assemblages in southern Iberian Mediterranean wetlands. J. Limnol. 2015, 74, 169–181. [Google Scholar] [CrossRef]
- Seminara, M.; Vagaggini, D.; Stoch, F. Long-term monitoring of astatic water bodies: Microcrustaceans as indicators of hydroperiod length in ponds and pools. Rend. Lincei 2015, 26, 345–352. [Google Scholar] [CrossRef]
- Boix, D.; Kneitel, J.; Robson, B.J.; Duchet, C.; Zùñiga, L.; Day, J.; Gascòn, S.; Sala, J.; Quintana, X.D.; Blaustein, L. Invertebrates of Freshwater Temporary Ponds in Mediterranean Climates Invertebrates in Freshwater Wetlands; Springer: New York, NY, USA, 2016; pp. 141–189. [Google Scholar]
- Diaz-Paniagua, C.; Fernandez-Zamudio, R.; Florencio, M.; Garcia Murillo, P.; Gomez-Rodriguez, C.; Portheault, A.; Serrano, L.; Siljestrom, P. Temporay ponds from Donana National Park: A system of natural habitats for the preservation of aquatic flora and fauna. Limnetica 2010, 29, 41–58. [Google Scholar]
- Semlitsh, R.D. Amphibian Conservation; Smithsonian Books: Washington, DC, USA; London, UK, 2003; 336p. [Google Scholar]
- Bisquert-Ribes, M.; Martínez, A.; Castillo-Escrivà, A.; Gálvez, Á.; Iepure, S.; De Manuel, B.; Marinho, M.; Martins, F.; Olmo, C.; Mesquita-Joanes, F.; et al. An evaluation of crustacean and rotifer diversity and composition in a temporary pond metacommunity: Comparing results from field samples and a hatching experiment. Freshw. Biol. 2023, 68, 378–390. [Google Scholar] [CrossRef]
- Alfonso, G.; Belmonte, G.; Ernandes, P.; Zuccarello, V. Stagni Temporanei Mediterranei in Puglia. Biodiversità ed Aspetti di un Habitat Poco Conosciuto; Grifo: Lecce, Italy, 2011; 43p. [Google Scholar]
- Alfonso, G.; Frassanito, A.G.; Modesti, F.; Belmonte, G. Crostacei di Stagni e Pozze del Parco Nazionale Dell’alta Murgia; Parco nazionale dell’Alta Murgia: Bari, Italy, 2017; 64p. [Google Scholar]
- Caldara, M.; Fatiguso, R.; Garganese, A.; Pennetta, L. Bibliografia Geologica Della Puglia; SAFRA: Bari, Italy, 1990. [Google Scholar]
- Blasi, C.; Michetti, L. The Climate of Italy; Blasi, C., Boitani, L., La Posta, S., Manes, F., Marchetti, M., Eds.; Biodiversity in Italy; Palombi Editori: Rome, Italy, 2007; pp. 57–66. [Google Scholar]
- Ernandes, P.; Gigante, D.; Beccarisi, L.; Marchiori, S.; Venanzoni, R.; Zuccarello, V. Isoëto-Nanojuncetea in Puglia (S-Italy): First phytosociological survey. Plant Sociol. 2017, 54, 23–36. [Google Scholar]
- Tomaselli, V.; Beccarisi, L.; Brullo, S.; Cambria, S.; Forte, L.; Minissale, P.; Veronico, G. Phytosociological research on temporary ponds in Apulia (southern Italy). Mediterr. Bot. 2020, 41, 15–41. [Google Scholar] [CrossRef]
- Pignatti, S. Flora d’Italia, 2nd ed.; Edagricole: Bologna, Italy, 2017; Volume 1, 1064p. [Google Scholar]
- Pignatti, S. Flora d’Italia, 2nd ed.; Edagricole: Bologna, Italy, 2017; Volume 2, 1178p. [Google Scholar]
- Pignatti, S. Flora d’Italia, 2nd ed.; Edagricole: Bologna, Italy, 2018; Volume 3, 1287p. [Google Scholar]
- Pignatti, S. Flora d’Italia, 2nd ed.; Edagricole: Bologna, Italy, 2019; Volume 4, 1054p. [Google Scholar]
- Tutin, T.G.; Burges, N.A.; Chater, A.O.; Edmondson, J.R.; Heywood, V.H.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. (Eds.) Flora Europaea, 2nd ed.; Volume 1: Psilotaceae to Platanaceae; Cambridge University Press: Cambridge, UK, 1993; 630p. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. (Eds.) Flora Europaea; Volume 2: Rosaceae to Umbelliferae; Cambridge University Press: Cambridge, UK, 1968; 500p. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. (Eds.) Flora Europaea; Volume 3: Diapensiaceae to Myoporaceae, 1. paperback print; Cambridge University Press: Cambridge, UK, 1972; 416p. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. (Eds.) Flora Europaea; Volume 4: Plantaginaceae to Compositae (and Rubiaceae); Cambridge University Press: Cambridge, UK, 1976; 552p. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. (Eds.) Flora Europaea; Volume 5: Alismataceae to Orchidaceae (Monicotyledones); Cambridge University Press: Cambridge, UK, 1980; 506p. [Google Scholar]
- Landucci, F.; Acosta, A.T.R.; Agrillo, E.; Attorre, F.; Biondi, E.; Cambria, V.E.; Chiarucci, A.; Del Vico, E.; De Sanctis, M.; Facioni, L.; et al. VegItaly: The Italian collaborative project for a national vegetation database. Plant Biosyst. 2012, 146, 756–763. [Google Scholar] [CrossRef]
- Angelini, P.; Casella, L.; Grignetti, A.; Genovesi, P. (Eds.) Manuali Per il Monitoraggio di Specie e Habitat di Interesse Comunitario (Direttiva 92/43/CEE) in Italia: Habitat; ISPRA: Lombardy, Italy, 2016; 294p. [Google Scholar]
- Biondi, E.; Blasi, C.; Burrascano, S.; Casavecchia, S.; Copiz, R.; Del Vico, E.; Galdenzi, D.; Gigante, D.; Lasen, C.; Spampinato, G.; et al. Manuale Italiano di Interpretazione Degli Habitat Della Direttiva 92/43/CEE. 2009. Available online: http://vnr.unipg.it/habitat (accessed on 5 July 2024).
- Biondi, E.; Blasi, C. Prodromo Alla Vegetazione d’Italia. 2015. Available online: http://www.prodromo-vegetazione-italia.org (accessed on 5 July 2024).
- Galasso, G.; Conti, F.; Peruzzi, L.; Alessandrini, A.; Ardenghi, N.M.G.; Bacchetta, G.; Banfi, E.; Barberis, G.; Bernardo, L.; Bouvet, D.; et al. A second update to the checklist of the vascular flora alien to Italy. Plant Biosyst. 2024, 158, 297–340. [Google Scholar] [CrossRef]
- Galasso, G.; Conti, F.; Peruzzi, L.; Ardenghi, N.M.G.; Banfi, E.; Celesti-Grapow, L.; Albano, A.; Alessandrini, A.; Bacchetta, G.; Ballelli, S.; et al. An updated checklist of the vascular flora alien to Italy. Plant Biosyst. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Conti, F.; Manzi, A.; Pedrotti, F. Liste Rosse Regionali Delle Piante d’Italia; Società Botanica Italiana e WWF Italia: Roma, Italy, 1997. [Google Scholar]
- Scoppola, A.; Spampinato, G. (Eds.) Atlante delle specie a rischio di estinzione (CD-ROM). In Stato Delle Conoscenze Sulla Flora Vascolare d’Italia; Palombi Editori: Roma, Italy, 2005. [Google Scholar]
- Zito, P.; Sajeva, M.; Rocco, M. Le specie vegetali italiane presenti nella normativa CITES dell’Unione Europea. Inf. Bot. Ital. 2008, 40, 43–69. [Google Scholar]
- Bilz, M.; Kell, S.P.; Maxted, N.; Lansdown, R.V. European Red List of Vascular Plants; Publications Office of the European Union: Luxembourg, 2011; 145p. [Google Scholar]
- Rossi, G.; Montagnani, C.; Gargano, D.; Peruzzi, L.; Abeli, T.; Ravera, S.; Cogoni, A.; Fenu, G.; Magrini, S.; Gennai, M.; et al. (Eds.) Lista Rossa della Flora Italiana. 1. Policy Species e Altre Specie Minacciate; Comitato Italiano IUCN e Ministero dell’Ambiente e della Tutela del Territorio e del Mare: Roma, Italy, 2013. [Google Scholar]
- European Environment Agency. Reference Portal for Reporting Under Article 17 of the Habitats Directive. Eionet: Central Data Repository. 2023. Available online: https://cdr.eionet.europa.eu/help/habitats_art17 (accessed on 5 July 2024).
- Robinson, A.H.; Morrison, J.L.; Muehrcke, P.C.; Kimerling, A.J.; Guptill, S.C. Elements of Cartography, 6th ed.; John Wiley & Sons: Hoboken, NJ, USA, 1995. [Google Scholar]
- Helm, A.; Zobel, M.; Moles, A.T.; Szava-Kovats, R.; Pärtel, M. Characteristic and derived diversity: Implementing the species pool concept to quantify conservation condition of habitats. Divers. Distrib. 2015, 21, 711–721. [Google Scholar] [CrossRef]
- Bijlsma, R.J.; Agrillo, E.; Attorre, F.; Boitani, L.; Brunner, A.; Evans, P.; Foppen, R.; Gubbay, S.; Janssen, J.A.M.; van Kleunen, A.; et al. Defining and Applying the Concept of Favourable Reference Values for Species and Habitats Under the EU Birds and Habitats Directives; Technical report; Wageningen Environmental Research: Wageningen, The Netherlands, 2019; Available online: https://library.wur.nl/WebQuery/wurpubs/fulltext/469035 (accessed on 5 July 2024).
- Dussart, B. Les Copépodes Des Eaux Continentales d’Europe Occidentale. I. Calanoïdes et Harpacticoïdes; N. Boubée et Cie: Paris, France, 1967. [Google Scholar]
- Kiefer, F. Das Zooplankton der Binnengewässer. Freilebende Copepoda; Die Binnengewässer. 2. E.; Schweizerbart’sche Verlagbuchhandlung: Stuttgart, Germany, 1978; 343p. [Google Scholar]
- Petkovski, T.K. Calanoida (Crustacea-Copepoda); Prirodonaučen Muzej na Makedonija: Skopje, Yugoslavia, 1983; 182p. [Google Scholar]
- Stella, E. Copepoda: Calanoida; Calderini: Bologna, Italy, 1984; 101p. [Google Scholar]
- Einsle, U. Crustacea Copepoda. Calanoida and Cyclopoida; Süsswasserfauna Mitteleuropas, 8/4-1; Gustav Fisher Verlag: Jena, Germany, 1993. [Google Scholar]
- Margaritora, F.G. Cladocera; Calderini: Bologna, Italy, 1985. [Google Scholar]
- Alonso, M. Crustacea, Branchiopoda; Museo Nacional de Ciencias Naturales: Madrid, Spain, 1996; 486p. [Google Scholar]
- Dussart, B. Les Copépodes des Eaux Continentales d’Europe Occidentale. II. Cyclopoïdes; N. Boubée et Cie: Paris, France, 1969. [Google Scholar]
- Meisch, C. Freshwater Ostracoda of Western and Central Europe; Spektrum Akademischer Verlag: Heidelberg, Germany, 2000; 522p. [Google Scholar]
- Cottarelli, V.; Mura, G. Anostraci, Notostraci, Concostraci Crustacea: Anostraca, Notostraca, Conchostraca; Consiglio Nazionale Delle Ricerche: Roma, Italy, 1983; Volume 18. [Google Scholar]
- Argano, R. Isopodi (Crustacea: Isopoda); Consiglio Nazionale Delle Ricerche: Roma, Italy, 1979; Volume 5. [Google Scholar]
- De Caceres, M.; Oliva, F.; Font, X.; Vives, S. Ginkgo, a program for non-standard multivariate fuzzy analysis. Adv. Fuzzy Sets Syst. 2007, 2, 41–56. [Google Scholar]
- Zadeh, L.A. Fuzzy sets. Inf. Control 1965, 8, 338–353. [Google Scholar] [CrossRef]
- Bezdek, J.C. Pattern Recognition With Fuzzy Objective Function Algorithms; Kluwer Academic Publishers: Norwell, MA, USA, 1981. [Google Scholar]
- Feoli, E.; Zuccarello, V. Syntaxonomy: A source of useful fuzzy sets for environmental analysis? Coenoses 1988, 3, 141–147. [Google Scholar]
- Alfonso, G.; Belmonte, G. Calanoida (Crustacea Copepoda) from the inland waters of Apulia (south−eastern Italy). J. Limnol. 2011, 70, 57–68. [Google Scholar] [CrossRef]
- Brown, K.S. Vanishing pools taking species with them. Science 1998, 281, 626. [Google Scholar] [CrossRef]
- Gascón, S.; Machado, M.; Sala, J.; Cancela Da Fonseca, L.; Cristo, M.; Boix, D. Spatial characteristics and species niche attributes modulate the response by aquatic passive dispersers to habitat degradation. Mar. Freshwater Res. 2012, 63, 232–245. [Google Scholar] [CrossRef]
- Bagella, S.; Gascón, S.; Caria, M.C.; Sala, J.; Mariani, M.A.; Boix, D. Identifying key environmental factors related to plant and crustacean assemblages in Mediterranean temporary ponds. Biodivers. Conserv. 2010, 19, 1749–1768. [Google Scholar] [CrossRef]
- Bagella, S. Which perspectives for Mediterranean temporary ponds in the European Union in the third millennium? Biodivers. Conserv. 2023, 32, 839–849. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Acronym | Site Name | Apulian Code ([37]) | Latitude (WGS84) | Longitude (WGS84) | Altitude (m a.s.l.) | Morphological Type | Brief Description | Pond Area (m2) |
|---|---|---|---|---|---|---|---|---|
| BEL | Masseria Bellimento | PU058 | 17°55′21.2″ | 40°11′46.7″ | 0 | Dolines | System of two dolines 15 m apart. | 1720 |
| DTO | groove beside Padule Rotondo (Bosco don Tommaso) | PU233 | 18°16′49.6″ | 39°58′39.8″ | 113 | Dolines | A doline crossed centrally by an artificial canal and surrounded by cultivated fields. | 3103 |
| FEL | Felline | PU156 | 18°7′16.3″ | 39°55′52.9″ | 44 | Cupular pools | System of many small limestone cupular pools. | 212 |
| FER | Laccu Feretru | PU059 | 18°11′7.1″ | 40°13′0.6″ | 66 | Dolines | A doline surrounded by cultivated fields and a stone quarry. | 899 |
| IAC | Iacorizzo | PU234 | 17°49′4.0″ | 40°23′11.7″ | 63 | Dolines | A doline with a long to permanent hydroperiod, surrounded by cultivated fields also subject to flooding | 2368 |
| LEU | Leuca: vaschette su calcare | PU252, PU253, PU254 | 18°21′10.3″ | 39°48′19.4″ | 50 | Cupular pools | A spread system of limestone cupular pools, within a maximum distance of 1.3 km. | 480 |
| MAA | Madonna del Lago A | PU126 | 18°10′3.6″ | 40°16′41.5″ | 57 | Dolines | A pond with a relatively long flood phase. | 287 |
| MAN | Bosco Mangiato | PU042 | 17°14′43.0″ | 40°44′26.6″ | 427 | Dolines | A pond in a wooded context. | 345 |
| PAL | Masseria I Pali | PU124, PU125 | 18°12′49.5″ | 39°50′54.1″ | 46 | Cupular pools | Two limestone cupular pool systems, 70 m apart. | 270 |
| PMA | Padula Mancina | PU020 | 18°18′37.0″ | 39°59′8.6″ | 102 | Dolines | A late hydroperiod pond, surrounded by earlier-drying flooded meadows. | 8165 |
| POS | Posticeddu | PU227 | 17°50′26.9″ | 40°41′24.3″ | 9 | Waterlogged soils | A seasonally bogged coastal site. | 1076 |
| SEM | Masseria Semeraro | PU231 | 17°3′46.7″ | 40°36′45.8″ | 137 | Cupular pools | Small limestone cupular pools, in shrubby landscape. | 22 |
| SPE | Lo Specchione | PU235 | 17°53′4.5″ | 40°29′40.1″ | 72 | Dolines | A doline surrounded by cultivated fields. | 753 |
| SPL | Lago Splendore | PU218 | 16°22′57.5″ | 40°45′54.5″ | 457 | Dolines | A pond in a wooded context. | 2145 |
| SUR | Suriani 3 | PU100 | 18°14′16.7″ | 40°4′31.3″ | 110 | Waterlogged soils | Site is part of a wetland made up of marshes and artificial canals, surrounded by cultivated fields. | 71,043 |
| ZE1 | Paduli 1, strada Silva (Zello) | PU103 | 18°14′5.2″ | 40°3′38.5″ | 116 | Waterlogged soils | Area subject to flooding between cultivated fields. | 4367 |
| Site | Estimated Area (m2) | Areal Criterion | FCSi | Structural Criterion | High Importance Pressure Category | Fun-ctional Criterion | Habitat 3170 Conservation Status |
|---|---|---|---|---|---|---|---|
| BEL | 1299 | NA | −0.1 | − | PA04 | − | Bad |
| DTO | 310 | NA | −0.5 | − | PA05 | − | Bad |
| FEL | 23 | + | −0.1 | − | + | Favourable | |
| FER | 62 | + | −0.5 | − | PA05 | − | Inadequate |
| IAC | 7 | NA | −0.4 | − | PA04, PA17 | − | Bad |
| LEU | 480 | + | NA | NA | NA | Favourable | |
| MAA | 86 | + | −0.3 | - | + | Favourable | |
| MAN | 173 | NA | 0.0 | + | + | Favourable | |
| PAL | 216 | − | −0.3 | − | + | Inadequate | |
| PMA | 3280 | NA | −0.4 | − | PA05 | − | Bad |
| POS | 215 | + | −0.2 | − | + | Favourable | |
| SEM | 7 | NA | 0.4 | + | + | Favourable | |
| SPE | 377 | NA | 0.0 | − | PA17 | − | Bad |
| SPL | 1073 | NA | −0.6 | − | PA05 | − | Bad |
| SUR | 49,730 | NA | −0.4 | − | PA05 | − | Bad |
| ZE1 | 2620 | NA | 0.2 | + | PA05 | − | Favourable |
| Species | g-1 | g-2 | g-3 |
|---|---|---|---|
| Lythrum hyssopifolia L. | 0.85 | 0.42 | 0.30 |
| Mentha pulegium L. | 0.41 | 0.83 | 0.77 |
| Polypogon subspathaceus Req. | 0.35 | 0.71 | 0.19 |
| Juncus pygmaeus Rich. ex Thuill. | 0.08 | 0.65 | 0.13 |
| Lythrum borysthenicum (Schrank) Litv. | 0.04 | 0.56 | 0.10 |
| Ranunculus sardous Crantz | 0.05 | 0.50 | 0.55 |
| Juncus bufonius L. | 0.42 | 0.55 | 0.14 |
| Symphyotrichum squamatum (Spreng.) G.L. Nesom | 0.41 | 0.40 | 0.52 |
| Paspalum distichum L. | 0.09 | 0.50 | 0.28 |
| Lotus angustissimus L. | 0.05 | 0.50 | 0.35 |
| Poa infirma Kunth | 0.47 | 0.01 | 0.01 |
| Isolepis cernua (Vahl) Roem. et Schult. | 0.08 | 0.46 | 0.13 |
| Cynodon dactylon (L.) Pers. | 0.03 | 0.09 | 0.44 |
| Fauna\Flora | Mentha puleg. L. | Polypogon subspat. Req. | Lythrum hyssopif. L. | Ranunculus sardous Crantz | Juncus bufonius L. | Juncus pygm. Rich. ex Thuil | Lythrum boryst. (Schrank) | Isolepis cernua (Vahl) Roem. et Schult. | Symphyotrichum squam. (Spreng.) G.L. Nesom | Paspalum distic. L. | Lotus angust. L. | Cynodon dactylon (L.) Pers. | Poa infirma Kunth |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chironomidae | 0.70 | 0.55 | 0.71 | 0.47 | 0.59 | 0.52 | 0.43 | 0.43 | 0.55 | 0.52 | 0.65 | 0.33 | 0.41 |
| Culicidae | 0.46 | 0.29 | 0.80 | 0.31 | 0.46 | 0.17 | 0.19 | 0.19 | 0.43 | 0.51 | 0.34 | 0.44 | 0.53 |
| Isothomidae | 0.37 | 0.31 | 0.72 | 0.17 | 0.67 | 0.37 | 0.20 | 0.41 | 0.46 | 0.37 | 0.37 | 0.24 | 0.58 |
| Limnohalacaridae | 0.40 | 0.51 | 0.63 | 0.18 | 0.73 | 0.40 | 0.45 | 0.45 | 0.51 | 0.20 | 0.40 | 0.00 | 0.63 |
| Alona nuragica | 0.49 | 0.46 | 0.58 | 0.17 | 0.33 | 0.37 | 0.41 | 0.41 | 0.31 | 0.37 | 0.37 | 0.00 | 0.29 |
| Sminthuridae | 0.37 | 0.46 | 0.58 | 0.33 | 0.50 | 0.55 | 0.41 | 0.41 | 0.31 | 0.37 | 0.18 | 0.00 | 0.29 |
| Ceriodaphnia sp. | 0.40 | 0.51 | 0.32 | 0.18 | 0.18 | 0.20 | 0.22 | 0.45 | 0.34 | 0.00 | 0.20 | 0.00 | 0.32 |
| C. dubia | 0.35 | 0.22 | 0.41 | 0.24 | 0.24 | 0.26 | 0.29 | 0.00 | 0.22 | 0.26 | 0.26 | 0.00 | 0.82 |
| Cypridopsis | 0.21 | 0.00 | 0.50 | 0.00 | 0.29 | 0.00 | 0.00 | 0.00 | 0.27 | 0.00 | 0.00 | 0.00 | 1.00 |
| Diacyclops nanus | 0.64 | 0.53 | 0.63 | 0.58 | 0.58 | 0.47 | 0.53 | 0.35 | 0.67 | 0.47 | 0.63 | 0.41 | 0.25 |
| D. bicuspidatus | 0.76 | 0.36 | 0.45 | 0.52 | 0.13 | 0.28 | 0.32 | 0.32 | 0.36 | 0.57 | 0.57 | 0.55 | 0.22 |
| Notonectidae | 0.75 | 0.27 | 0.50 | 0.29 | 0.43 | 0.32 | 0.18 | 0.53 | 0.53 | 0.47 | 0.32 | 0.20 | 0.25 |
| Chydorus sphaericus | 0.85 | 0.40 | 0.63 | 0.58 | 0.43 | 0.47 | 0.53 | 0.18 | 0.53 | 0.79 | 0.63 | 0.41 | 0.25 |
| Proales sp. | 0.54 | 0.34 | 0.47 | 0.55 | 0.18 | 0.40 | 0.45 | 0.22 | 0.34 | 0.60 | 0.60 | 0.26 | 0.32 |
| Megacyclops viridis | 0.75 | 0.40 | 0.63 | 0.43 | 0.43 | 0.32 | 0.35 | 0.18 | 0.67 | 0.63 | 0.47 | 0.41 | 0.00 |
| Pleuroxus letourneouxi | 0.49 | 0.62 | 0.29 | 0.67 | 0.33 | 0.37 | 0.41 | 0.41 | 0.46 | 0.18 | 0.18 | 0.24 | 0.00 |
| Keratella sp. | 0.74 | 0.31 | 0.43 | 0.50 | 0.17 | 0.37 | 0.41 | 0.20 | 0.46 | 0.73 | 0.55 | 0.24 | 0.00 |
| Simocephalus vetulus | 0.67 | 0.17 | 0.47 | 0.55 | 0.18 | 0.20 | 0.22 | 0.00 | 0.51 | 0.60 | 0.40 | 0.52 | 0.00 |
| Daphnia (Ctenodaphnia) chevreuxi | 0.60 | 0.76 | 0.35 | 0.41 | 0.41 | 0.67 | 0.75 | 0.50 | 0.19 | 0.67 | 0.45 | 0.00 | 0.00 |
| Daphnia (Ctenodaphnia) atkinsoni | 0.60 | 0.57 | 0.18 | 0.61 | 0.20 | 0.45 | 0.50 | 0.50 | 0.38 | 0.45 | 0.67 | 0.29 | 0.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beccarisi, L.; Zuccarello, V.; Accogli, R.; Belmonte, G. Biodiversity and Possible Bio-Indicators of Mediterranean Temporary Ponds in Southern Apulia, Italy. Diversity 2024, 16, 559. https://doi.org/10.3390/d16090559
Beccarisi L, Zuccarello V, Accogli R, Belmonte G. Biodiversity and Possible Bio-Indicators of Mediterranean Temporary Ponds in Southern Apulia, Italy. Diversity. 2024; 16(9):559. https://doi.org/10.3390/d16090559
Chicago/Turabian StyleBeccarisi, Leonardo, Vincenzo Zuccarello, Rita Accogli, and Genuario Belmonte. 2024. "Biodiversity and Possible Bio-Indicators of Mediterranean Temporary Ponds in Southern Apulia, Italy" Diversity 16, no. 9: 559. https://doi.org/10.3390/d16090559