
Functional Diversity and Ecosystem Services of Birds in Productive Landscapes of the Colombian Amazon


Abstract

1. Introduction
2. Materials and Methods
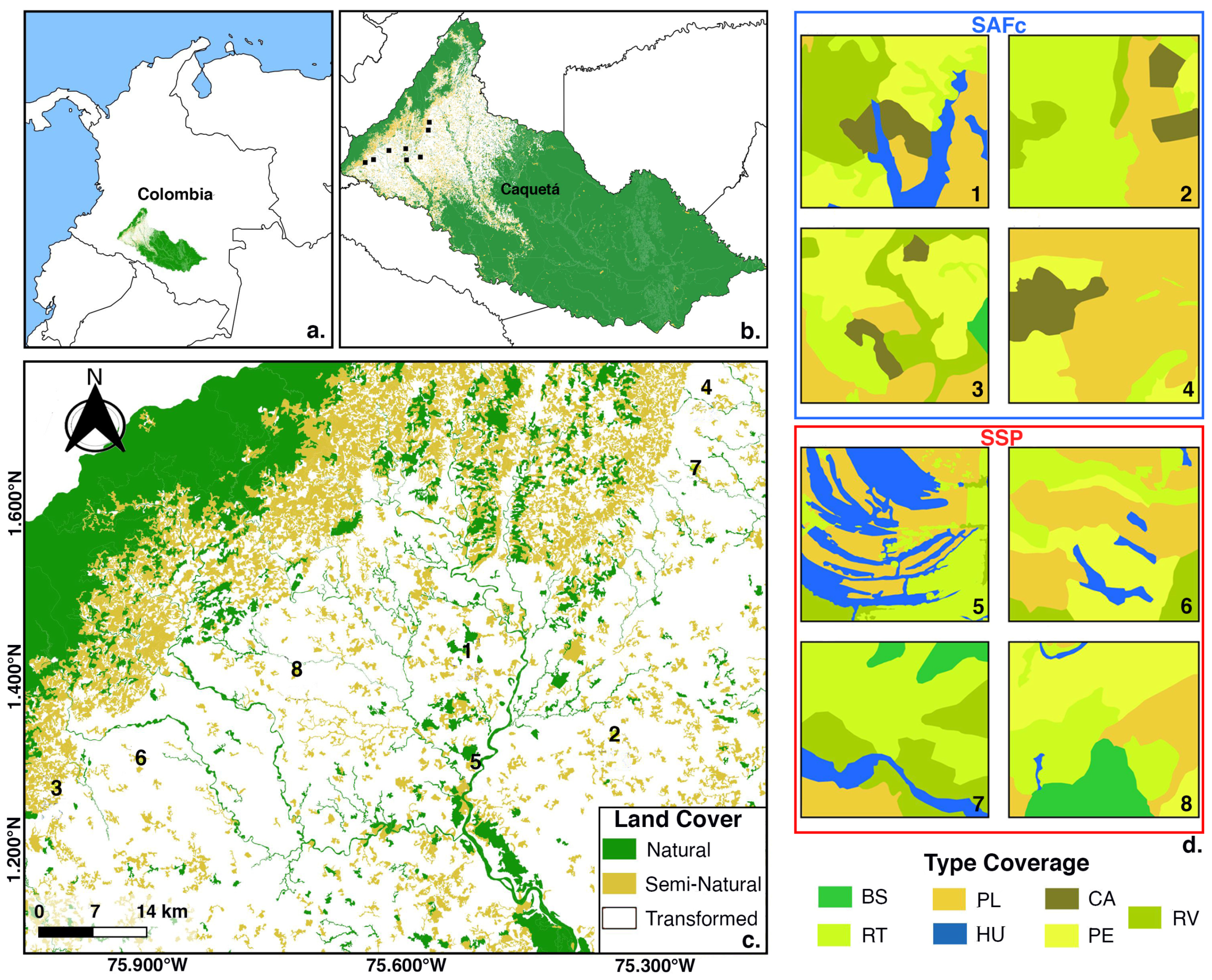
2.1. Study Area
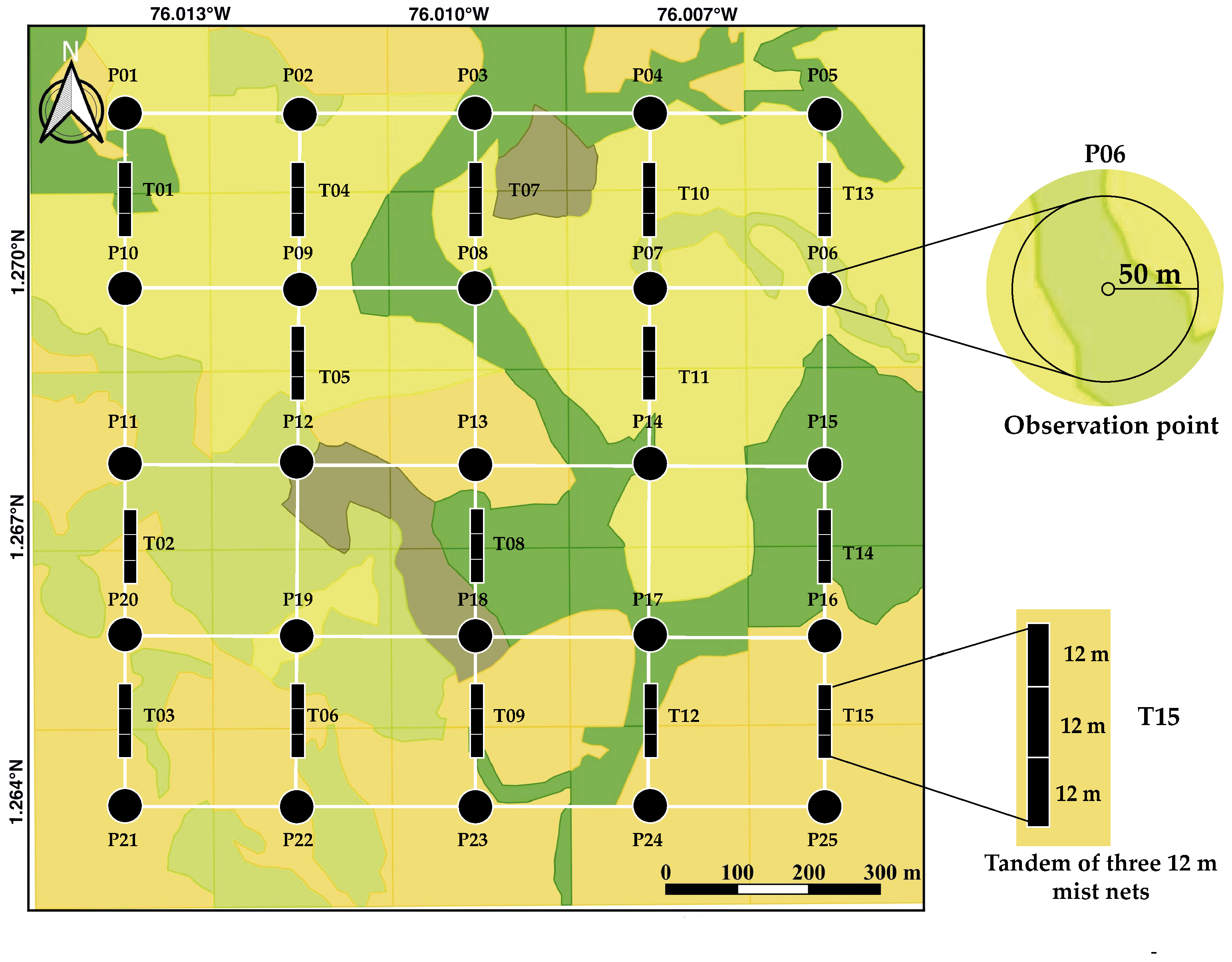
2.2. Bird Census and Functional Traits
2.3. Statistical Analysis
3. Results
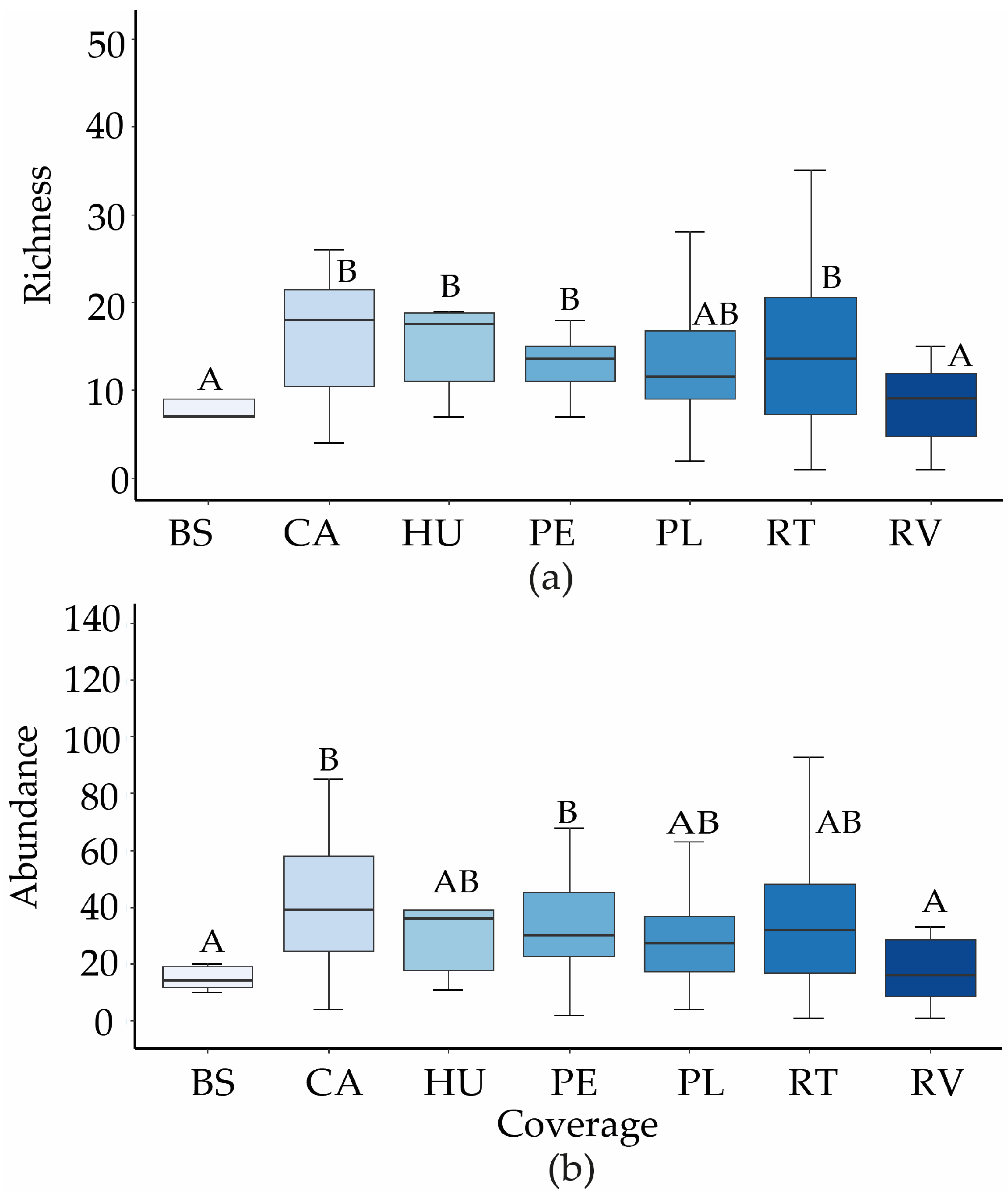
3.1. Functional Diversity of the Bird Community by Vegetation Cover Types and Systems
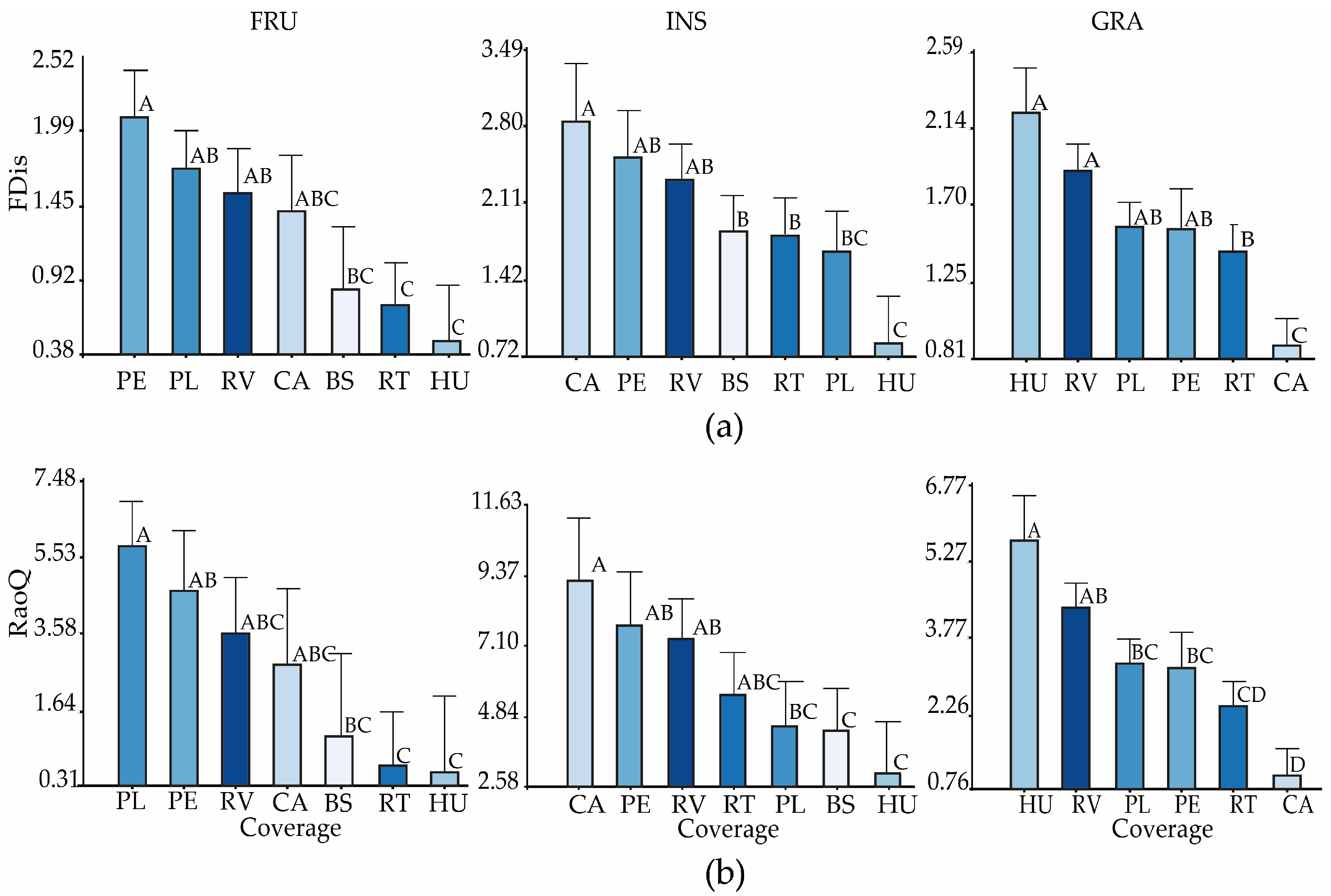
3.2. Functional Diversity of the Bird Community by Guilds, Vegetation Cover Types, and Systems
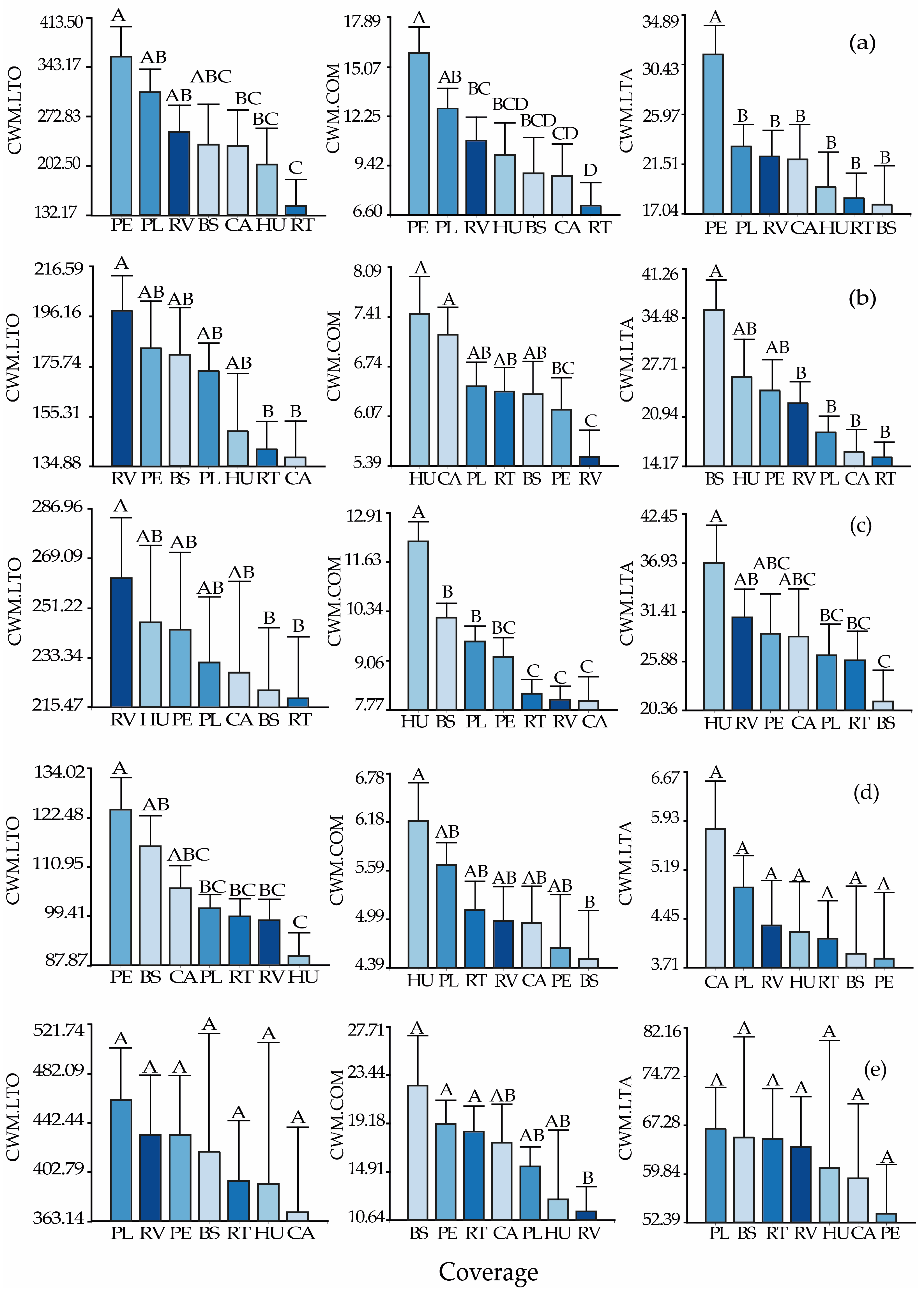
3.3. Functional Composition of the Bird Community by Vegetation Cover Types and Systems
3.4. Functional Composition of the Bird Community by Guilds, Vegetation Cover Types, and Systems
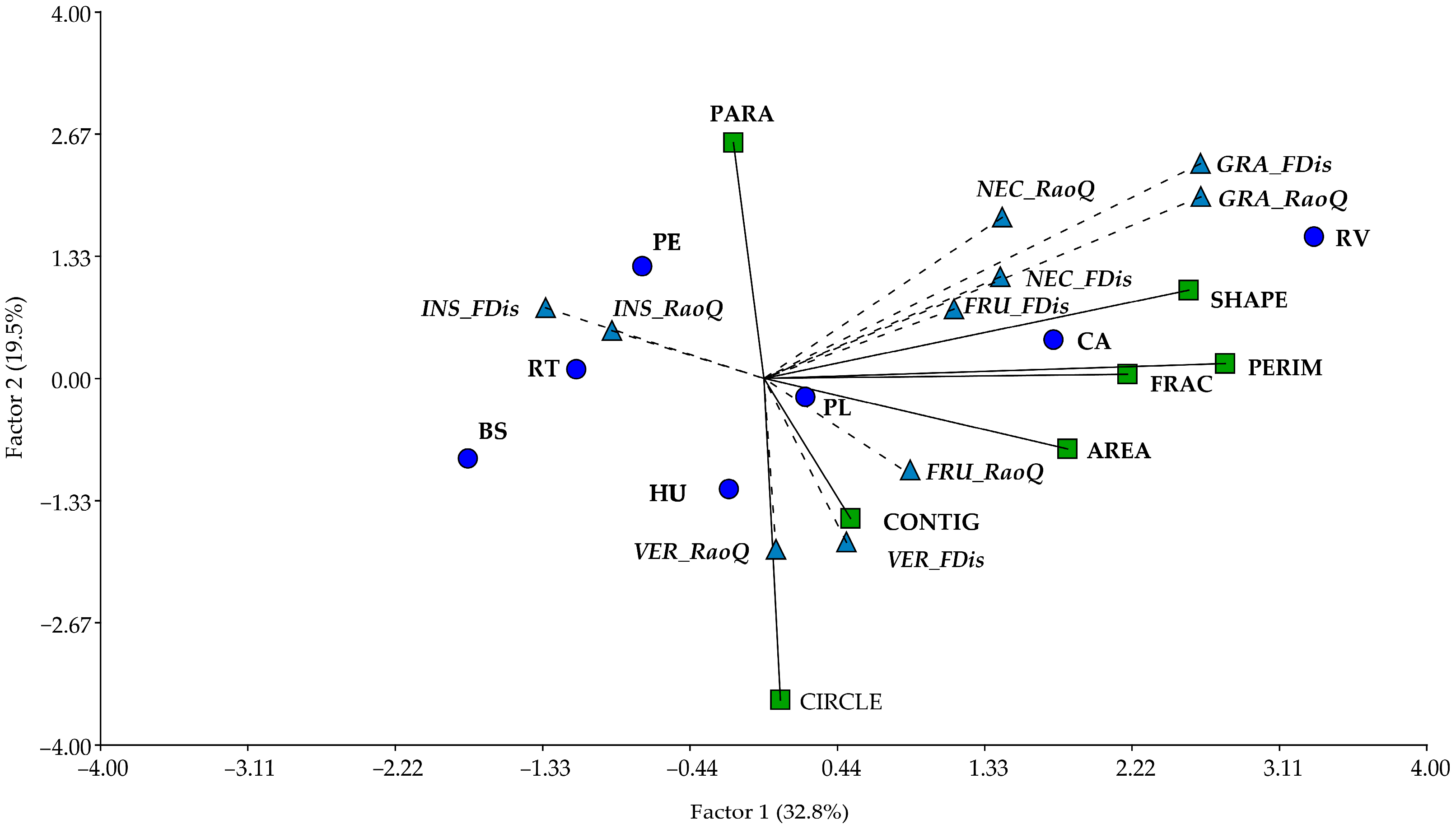
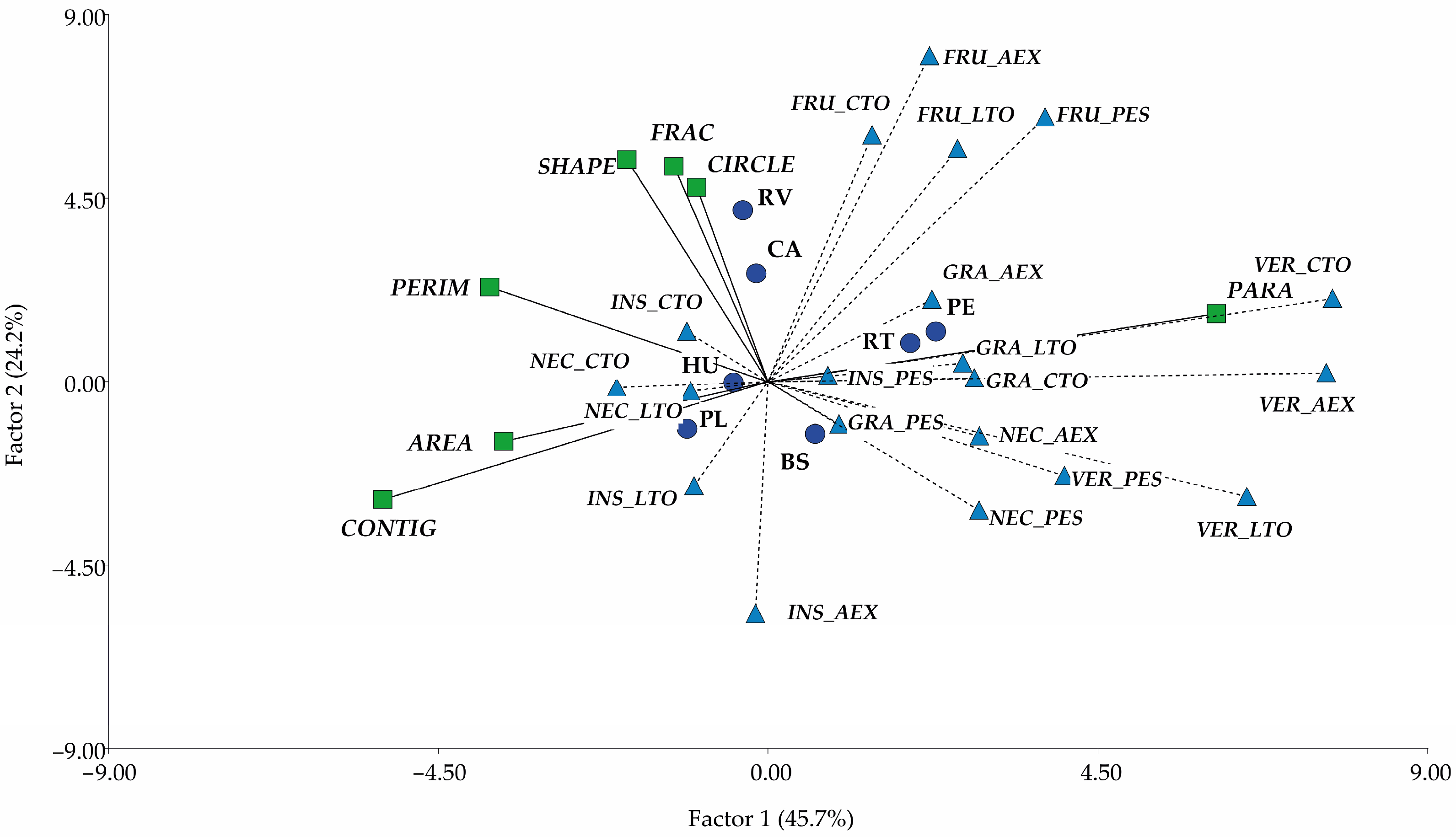
3.5. Relationship Between Functional Diversity and Composition and Landscape Structure
4. Discussion
4.1. Functional Diversity and Composition of Bird Communities by Vegetation Cover and System
4.2. Functional Diversity of Bird Communities by Guilds, Vegetation Cover Types, and Systems
4.3. Relationship Between Landscape Structure and Functional Diversity and Composition
4.4. Implications for Ecosystem Service Provision
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| FRU | Frugivores |
| INS | Insectivores |
| GRA | Granivores |
| NEC | Nectarivores |
| VER | Vertebrate Consumers |
| BS | Secondary Forests |
| CA | Cacao Agroforestry Crops |
| HU | Wetlands |
| PE | Weedy Pastures |
| PL | Clean Pastures |
| RT | Early Successional Shrubs |
| RV | Late Successional Shrubs |
| FRAC | Mean Fractal Index |
| CONTIG | Contiguity Index |
| SHAPE | Shape Index |
| AREA | Total Area |
| CIRCLE | Circularity Index |
| PERIM | Perimeter–Area Ratio |
| PARA | Mean Perimeter–Area Ratio |
| LTO | Total Body Length |
| LTA | Tarsus Length |
| AEX | Extended Wing |
| COM | Commissure |
| ALT | Beak Height |
| CTO | Total Culmen Length |
| PES | Weight |
References
- Armenteras, D.; Murcia, U.; González, T.M.; Barón, O.J.; Arias, J.E. Scenarios of land use and land cover change for NW Amazonia: Impact on forest intactness. Glob. Ecol. Conserv. 2019, 17, e00567. [Google Scholar] [CrossRef]
- Hänggli, A.; Levy, S.A.; Armenteras, D.; Bovolo, C.I.; Brandão, J.; Rueda, X.; Garrett, R.D. A systematic comparison of deforestation drivers and policy effectiveness across the Amazon biome. Environ. Res. Lett. 2023, 18, 073001. [Google Scholar] [CrossRef]
- Velásquez Valencia, A.; Bonilla Gómez, M.A. Influence of the configuration and heterogeneity of the agroforestry and silvopastoral mosaics on the bird community, Andean Amazon of Colombia. Rev. Biol. Trop. 2019, 67, 306–320. [Google Scholar] [CrossRef]
- Fontúrbel, F.E.; Betancurt-Grisales, J.F.; Vargas-Daza, A.M.; Castaño-Villa, G.J. Effects of habitat degradation on bird functional diversity: A field test in the Valdivian rainforest. For. Ecol. Manag. 2022, 522, 120466. [Google Scholar] [CrossRef]
- Koo, H.; Kleemann, J.; Cuenca, P.; Noh, J.K.; Fürst, C. Implications of landscape changes for ecosystem services and biodiversity: A national assessment in Ecuador. Ecosyst. Serv. 2024, 69, 101652. [Google Scholar] [CrossRef]
- Gebremichael, G.; Hundera, K.; De Decker, L.; Aerts, R.; Lens, L.; Atickem, A. Bird Community Composition and Functional Guilds Response to Vegetation Structure in Southwest Ethiopia. Forests 2022, 13, 2068. [Google Scholar] [CrossRef]
- Giraldo, L.P.; Chará, J.; Velásquez-Valencia, A.; Ordoñez-García, M. Vegetation structure and bird diversity in silvopastoral systems of the Amazonian piedmont of Colombia. Agrofor. Syst. 2024, 98, 2253–2266. [Google Scholar] [CrossRef]
- Habel, J.C.; Mulwa, M.; Wagner, L.; Schmitt, T.; Teucher, M.; Ulrich, W. Diverging responses to natural and anthropogenic habitats in aftrotropical birds and butterflies. Biodivers. Conserv. 2024, 33, 971–985. [Google Scholar] [CrossRef]
- Zamora-Marín, J.M.; Zamora-López, A.; Oliva-Paterna, F.J.; Torralva, M.; Sánchez-Montoya, M.M.; Calvo, J.F. From small waterbodies to large multi-service providers: Assessing their ecological multifunctionality for terrestrial birds in Mediterranean agroecosystems. Agric. Ecosyst. Environ. 2024, 359, 108760. [Google Scholar] [CrossRef]
- Murillo-Sandoval, P.J.; Clerici, N.; Correa-Ayram, C. Rapid loss in landscape connectivity after the peace agreement in the Andes-Amazon region. Glob. Ecol. Conserv. 2022, 38, e02205. [Google Scholar] [CrossRef]
- Mena, J.L.; Yagui, H.; Tejeda, V.; Cabrera, J.; Pacheco-Esquivel, J.; Rivero, J.; Pastor, P. Abundance of jaguars and occupancy of medium- and large-sized vertebrates in a transboundary conservation landscape in the northwestern Amazon. Glob. Ecol. Conserv. 2020, 23, e01079. [Google Scholar] [CrossRef]
- Negret Torres, P.J. Development, Armed Conflict and Conservation: Improving the Effectiveness of Conservation Decisions in Conflict Hotspots Using Colombia as a Case Study. Ph.D. Thesis, The University of Queensland, Brisbane, Australia, 2020. Available online: https://espace.library.uq.edu.au/view/UQ:4cfbe2d (accessed on 25 October 2024).
- Johnson, B.A.; Kumar, P.; Okano, N.; Dasgupta, R.; Shivakoti, B.R. Nature-based solutions for climate change adaptation: A systematic review of systematic reviews. Nat.-Based Solut. 2022, 2, 100042. [Google Scholar] [CrossRef]
- Baptiste, B.; Pinedo-Vasquez, M.; Gutierrez-Velez, V.H.; Andrade, G.I.; Vieira, P.; Estupiñán-Suárez, L.M.; Londoño, M.C.; Laurance, W.; Lee, T.M. Greening peace in Colombia. Nat. Ecol. Evol. 2017, 1, 0102. [Google Scholar] [CrossRef]
- Castro, P.; Azul, A.M.; Leal Filho, W.; Azeiteiro, U.M. (Eds.) Climate Change-Resilient Agriculture and Agroforestry: Ecosystem Services and Sustainability; Climate Change Management; Springer International Publishing: Cham, Switzerland, 2019; Available online: http://link.springer.com/10.1007/978-3-319-75004-0 (accessed on 14 August 2024).
- Clerici, N.; Armenteras, D.; Kareiva, P.; Botero, R.; Ramírez-Delgado, J.P.; Forero-Medina, G.; Ochoa, J.; Pedraza, C.; Schneider, L.; Lora, C.; et al. Deforestation in Colombian protected areas increased during post-conflict periods. Sci. Rep. 2020, 10, 4971. [Google Scholar] [CrossRef] [PubMed]
- Nair, P.K.R. Agroforestry Systems and Environmental Quality: Introduction. J. Environ. Qual. 2011, 40, 784–790. [Google Scholar] [CrossRef]
- Murgueitio Restrepo, E.; Chara Orozco, J.D.; Barahona Rosales, R.; Cuartas Cardona, C.A.; Naranjo Ramirez, J.F. Intensive Silvopastoral Systems (SSPi), a tool for mitigating and adapting to climate change. Trop. Subtrop. Agroecosystems 2014, 17, 501–507. Available online: https://www.revista.ccba.uady.mx/ojs/index.php/TSA/article/view/1558 (accessed on 14 August 2024).
- Davis, S.; Guerreiro Milheiras, S.; Olivier, P.L.; Barnes, L.; Shirima, D.; Kioko, E.; Sallu, S.M.; Ishengoma, E.; Marshall, A.R.; Pfeifer, M. Cropland can support high bird diversity in heterogeneous rural tropical landscapes. Bird Conserv. Int. 2024, 34, e13. [Google Scholar] [CrossRef]
- Frank, C.; Hertzog, L.; Klimek, S.; Schwieder, M.; Tetteh, G.O.; Böhner, H.G.S.; Röder, N.; Levers, C.; Katzenberger, J.; Kreft, H. Woody semi-natural habitats modulate the effects of field size and functional crop diversity on farmland birds. J. Appl. Ecol. 2024, 61, 987–999. [Google Scholar] [CrossRef]
- Alvarado, F.; Escobar, F.; Williams, D.R.; Arroyo-Rodríguez, V.; Escobar-Hernández, F. The role of livestock intensification and landscape structure in maintaining tropical biodiversity. Nichols E, editor. J. Appl. Ecol. 2018, 55, 185–194. [Google Scholar] [CrossRef]
- Álvarez, F.; Casanoves, F.; Suárez, J.C.; Rusch, G.M.; Ngo Bieng, M.A. An assessment of silvopastoral systems condition and their capacity to generate ecosystem services in the Colombian Amazon. Ecosyst. People 2023, 19, 2213784. [Google Scholar] [CrossRef]
- Williams, B.A.; Grantham, H.S.; Watson, J.E.M.; Alvarez, S.J.; Simmonds, J.S.; Rogéliz, C.A.; Da Silva, M.; Forero-Medina, G.; Etter, A.; Nogales, J.; et al. Minimising the loss of biodiversity and ecosystem services in an intact landscape under risk of rapid agricultural development. Environ. Res. Lett. 2020, 15, 014001. [Google Scholar] [CrossRef]
- Gantz, A.G.; Yáñez-Alvarado, M.; Zamorano, G.; Smith-Ramírez, C.; Valdivia, C.E.; Rau, J.R. Contrasting responses of taxonomic diversity and functional groups of birds to the agricultural landscape of southern Chile. Ornithol. Res. 2024, 32, 357–371. [Google Scholar] [CrossRef]
- Ocampo-Ariza, C.; Hanf-Dressler, T.; Maas, B.; Novoa-Cova, J.; Thomas, E.; Vansynghel, J.; Steffan-Dewenter, I.; Tscharntke, T. Regional differences of functional and taxonomic bird diversity in tropical agroforests of Peru. Conserv. Sci. Pract. 2024, 6, e13123. [Google Scholar] [CrossRef]
- Anderle, M.; Brambilla, M.; Hilpold, A.; Matabishi, J.G.; Paniccia, C.; Rocchini, D.; Rossin, J.; Tasser, E.; Torresani, M.; Tappeiner, U.; et al. Habitat heterogeneity promotes bird diversity in agricultural landscapes: Insights from remote sensing data. Basic Appl. Ecol. 2023, 70, 38–49. [Google Scholar] [CrossRef]
- Nogueira, O.M.A.; Palmeirim, A.F.; Peres, C.A.; Dos Santos-Filho, M. Synergistic effects of habitat configuration and land-use intensity shape the structure of bird assemblages in human-modified landscapes across three major neotropical biomes. Biodivers. Conserv. 2021, 30, 3793–3811. [Google Scholar] [CrossRef]
- Bain, G.C.; MacDonald, M.A.; Hamer, R.; Gardiner, R.; Johnson, C.N.; Jones, M.E. Changing bird communities of an agricultural landscape: Declines in arboreal foragers, increases in large species. R. Soc. Open Sci. 2020, 7, 200076. [Google Scholar] [CrossRef]
- Molina-Marin, D.A.; Rodas-Rua, J.C.; Lara, C.E.; Rivera-Páez, F.A.; Fontúrbel, F.E.; Castaño-Villa, G.J. Effects of Landscape Configuration on the Body Condition of Migratory and Resident Tropical Birds. Diversity 2022, 14, 432. [Google Scholar] [CrossRef]
- Peña, R.; Schleuning, M.; Miñarro, M.; García, D. Variable relationships between trait diversity and avian ecological functions in agroecosystems. Funct. Ecol. 2023, 37, 87–98. [Google Scholar] [CrossRef]
- Jacoboski, L.I.; Hartz, S.M. Using functional diversity and taxonomic diversity to assess effects of afforestation of grassland on bird communities. Perspect. Ecol. Conserv. 2020, 18, 103–108. [Google Scholar] [CrossRef]
- Lisón, F.; Matus-Olivares, C.; Troncoso, E.; Catalán, G.; Jiménez-Franco, M.V. Effect of forest landscapes composition and configuration on bird community and its functional traits in a hotspot of biodiversity of Chile. J. Nat. Conserv. 2022, 68, 126227. [Google Scholar] [CrossRef]
- Hewes, A.E.; Cuban, D.; Groom, D.J.E.; Sargent, A.J.; Beltrán, D.F.; Rico-Guevara, A. Variable evidence for convergence in morphology and function across avian nectarivores. J. Morphol. 2022, 283, 1483–1504. [Google Scholar] [CrossRef] [PubMed]
- Campo-Celada, M.; Jordano, P.; Benítez-López, A.; Gutiérrez-Expósito, C.; Rabadán-González, J.; Mendoza, I. Assessing short and long-term variations in diversity, timing and body condition of frugivorous birds. Oikos 2022, 2022, oik.08387. [Google Scholar] [CrossRef]
- Landázuri, O.S.; Tinoco, B.A.; Espinosa, C.I.; Jiménez-Franco, M.V.; Robledano, F. Effects of landscape composition and configuration on Andean birds are influenced by spatial scale. For. Ecol. Manag. 2024, 563, 121960. [Google Scholar] [CrossRef]
- Pizo, M.A.; Morales, J.M.; Ovaskainen, O.; Carlo, T.A. Frugivory Specialization in Birds and Fruit Chemistry Structure Mutualistic Networks across the Neotropics. Am. Nat. 2021, 197, 236–249. [Google Scholar] [CrossRef]
- Karp, D.S.; Chaplin-Kramer, R.; Meehan, T.D.; Martin, E.A.; DeClerck, F.; Grab, H.; Gratton, C.; Hunt, L.; Larsen, A.E.; Martínez-Salinas, A.; et al. Crop pests and predators exhibit inconsistent responses to surrounding landscape composition. Proc. Natl. Acad. Sci. USA 2018, 115, E7863–E7870. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Siefer, P.; Olmos-Moya, N.; Fontúrbel, F.E.; Lavandero, B.; Pozo, R.A.; Celis-Diez, J.L. Bird-mediated effects of pest control services on crop productivity: A global synthesis. J. Pest. Sci. 2022, 95, 567–576. [Google Scholar] [CrossRef]
- Bitani, N.; Cordier, C.P.; Ehlers Smith, D.A.; Ehlers Smith, Y.C.; Downs, C.T. Responses of bird functional communities to anthropogenic disturbances in the naturally fragmented Southern Mistbelt Forests in the Midlands of KwaZulu-Natal, South Africa. For. Ecol. Manag. 2024, 562, 121977. [Google Scholar] [CrossRef]
- Şekercioğlu, Ç.H.; Sutherland, W.J.; Buechley, E.R.; Li, B.V.; Ocampo-Peñuela, N.; Mahamued, B.A. Avian biodiversity collapse in the Anthropocene: Drivers and consequences. Front. Ecol. Evol. 2023, 11, 1202621. [Google Scholar] [CrossRef]
- Xu, Y.; Wu, Z.; Liu, X.; Han, Q.; Liu, P.; Wang, D.; Wang, J. Patch shape, connectivity and functional groups moderate the responses of bird diversity to agricultural diversification—A meta-analysis. Glob. Ecol. Conserv. 2024, 55, e03204. [Google Scholar] [CrossRef]
- Tilman, D. Functional Diversity. Encycl. Biodivers. 2001, 3, 109–120. [Google Scholar]
- Martínez-Salinas, A.; DeClerck, F.; Vierling, K.; Vierling, L.; Legal, L.; Vílchez-Mendoza, S.; Avelino, J. Bird functional diversity supports pest control services in a Costa Rican coffee farm. Agric. Ecosyst. Environ. 2016, 235, 277–288. [Google Scholar] [CrossRef]
- Figueroa-Alvarez, J.A.; Ortega-Álvarez, R.; Manson, R.H.; Sosa, V.J.; Vázquez-Reyes, L.D.; Medina Mena, I.; Bautista Bautista, L. Insectivorous birds and potential pest control services: An occupancy study of functional groups in a coffee landscape in Oaxaca, Mexico. Perspect. Ecol. Conserv. 2024, 22, 331–341. [Google Scholar] [CrossRef]
- Alvarez-Alvarez, E.A.; Almazán-Núñez, R.C.; Corcuera, P.; González-García, F.; Brito-Millán, M.; Alvarado-Castro, V.M. Land use cover changes the bird distribution and functional groups at the local and landscape level in a Mexican shaded-coffee agroforestry system. Agric. Ecosyst. Environ. 2022, 330, 107882. [Google Scholar] [CrossRef]
- Lessi, B.; Geneletti, D.; Cortinovis, C.; Dias, M.; Reis, M. Bird richness and Ecosystems Services across an urban to natural gradient in south-eastern Brazil: Implications for landscape planning and future scenarios. One Ecosyst. 2024, 9, e114955. [Google Scholar] [CrossRef]
- Alvarado Sandino, C.O.; Barnes, A.P.; Sepúlveda, I.; Garratt, M.P.D.; Thompson, J.; Escobar-Tello, M.P. Examining factors for the adoption of silvopastoral agroforestry in the Colombian Amazon. Sci. Rep. 2023, 13, 12252. [Google Scholar] [CrossRef] [PubMed]
- Acevedo-Charry, O.; Peña-Alzate, F.Á.; Beckers, J.; Cabezas, M.; Coral-Jaramillo, B.; Janni, O.; Ocampo, D.; Peñuela-Gomez, S.M.; Rocha-López, D.; Socolar, J.B.; et al. Avifauna del interfluvio de la cuenca media Caquetá Putumayo (Japurá-Içá), al sur de la Amazonia colombiana y su respuesta a la huella humana. Rev. Acad. Colomb. Cienc. Exactas Físicas Nat. 2021, 45, 229–249. [Google Scholar] [CrossRef]
- Decaëns, T.; Martins, M.B.; Feijoo, A.; Oszwald, J.; Dolédec, S.; Mathieu, J.; de Sartre, X.A.; Bonilla, D.; Brown, G.; Cuellar Criollo, Y.A.; et al. Biodiversity loss along a gradient of deforestation in Amazonian agricultural landscapes. Conserv. Biol. 2018, 32, 1380–1391. [Google Scholar] [CrossRef] [PubMed]
- Pulecio-Suarez, I.L.; Velázquez-Valencia, A.; Cruz-Trujillo, E.J. Riqueza y Abundancia de la comunidad de aves en tres sistemas de producción en el Piedemonte Amázonico. Rev. Inv. Agroemp. 2015, 1, 11–17. [Google Scholar]
- Agudelo-Hz, W.J.; Castillo-Barrera, N.C.; Uriel, M.G. Scenarios of land use and land cover change in the Colombian Amazon to evaluate alternative post-conflict pathways. Sci. Rep. 2023, 13, 2152. [Google Scholar] [CrossRef]
- Kim, J.H.; Park, S.; Kim, S.H.; Kang, K.; Waldman, B.; Lee, M.H.; Yu, M.; Yang, H.; Yong Chung, H.; Lee, E.J. Structural implications of traditional agricultural landscapes on the functional diversity of birds near the Korean Demilitarized Zone. Ecol. Evol. 2020, 10, 12973–12982. [Google Scholar] [CrossRef]
- Santos, J.S.D.; Dodonov, P.; Oshima, J.E.F.; Martello, F.; Santos De Jesus, A.; Ferreira, M.; Silva-Neto, C.M.; Ribeiro, M.C.; Garcia Collevatti, R. Landscape ecology in the Anthropocene: An overview for integrating agroecosystems and biodiversity conservation. Perspect. Ecol. Conserv. 2021, 19, 21–32. [Google Scholar] [CrossRef]
- IDEAM. Leyenda Nacional de Coberturas de la Tierra. Metodología Corine Land Cover Adaptada Para Colombia. Escala 1:100.000. Primera Edición. Vol. 1; Instituto de Hidrología, Meteorología y Estudios Ambientales: Bogotá, Colombia, 2010; 73p.
- McGarigal, K.; Cushman, S.; Ene, E. FRAGSTATS v4: Spatial Pattern Analysis Program for Categorical Maps. 2023. Available online: https://www.fragstats.org (accessed on 19 September 2024).
- Pliscoff, P.; Simonetti, J.A.; Grez, A.A.; Vergara, P.M.; Barahona-Segovia, R.M. Defining corridors for movement of multiple species in a forest-plantation landscape. Glob. Ecol. Conserv. 2020, 23, e01108. [Google Scholar] [CrossRef]
- Coddington, C.P.J.; Cooper, W.J.; Mokross, K.; Luther, D.A. Forest structure predicts species richness and functional diversity in Amazonian mixed-species bird flocks. Biotropica 2023, 55, 467–479. [Google Scholar] [CrossRef]
- Tobias, J.A.; Sheard, C.; Pigot, A.L.; Devenish, A.J.M.; Yang, J.; Sayol, F.; Neate-Clegg, M.H.; Alioravainen, N.; Weeks, T.L.; Barber, R.A.; et al. AVONET: Morphological, ecological and geographical data for all birds. Ecol. Lett. 2022, 25, 581–597. [Google Scholar] [CrossRef] [PubMed]
- Remsen, J.; Areta, J.; Bonaccorso, E.; Claramunt, S.; Lane, D.; Robbins, M.; Stiles, F.G.; Zimmer, K.J. A Classification of the Bird Species of South America. South American Classification Committee; Museum of Natural Science, Louisiana State University: Baton Rouge, LA, USA, 2025; Available online: https://www.museum.lsu.edu/~Remsen/SACCBaseline.htm (accessed on 29 August 2024).
- Echeverry-Galvis, M.Á.; Acevedo-Charry, O.; Avendaño, J.E.; Gómez, C.; Stiles, F.G.; Estela, F.A.; Cuervo, A.M. Official List of Birds of Colombia 2022: Additions, Taxonomic Changes, and Status Updates. Ornitol. Colomb. 2022, 22, 25–51. [Google Scholar] [CrossRef]
- Ayerbe-Quiñones, F. Illustrated Guide to Colombian Birdlife, 3rd ed.; WCS Colombia: Cali, Colombia, 2023; 459p. [Google Scholar]
- McMullan, M. Field Guide to the Birds of Colombia, 2023 ed.; McMullan Birding & Publishers: Cali, Colombia, 2023; 528p. [Google Scholar]
- Jarrett, C.; Smith, T.B.; Claire, T.T.R.; Ferreira, D.F.; Tchoumbou, M.; Elikwo, M.N.F.; Wolfe, J.D.; Brzeski, K.; Welch, A.; Hanna, R. Bird communities in African cocoa agroforestry are diverse but lack specialized insectivores. J. Appl. Ecol. 2021, 58, 1237–1247. [Google Scholar] [CrossRef]
- Michel, N.L.; Whelan, C.J.; Verutes, G.M. Ecosystem services provided by Neotropical birds. Condor 2020, 122, duaa022. [Google Scholar] [CrossRef]
- Pejchar, L.; Clough, Y.; Ekroos, J.; Nicholas, K.A.; Olsson, O.; Ram, D.; Tschumi, M.; Smith, H.G. Net Effects of Birds in Agroecosystems. BioScience 2018, 68, 896–904. [Google Scholar] [CrossRef]
- Di Rienzo, J.; Macchiavelli, R.E.; Casanoves, F. Linear Mixed Models: Applications in InfoStat, 1st ed.; National University of Córdoba Argentina: Córdoba, Argentina, 2017; Available online: https://scholar.google.com.co/citations?view_op=view_citation&hl=es&user=yumGXjoAAAAJ&cstart=20&pagesize=80&citation_for_view=yumGXjoAAAAJ:ML0RJ9NH7IQC (accessed on 25 January 2025).
- Di Rienzo, J.; Balzarini, M.; Gonzalez, L.; Casanoves, F.; Tablada, M.; Robledo, C.W. InfoStat Software, Version 2020; National University of Córdoba: Córdoba, Argentina, 2020. Available online: https://www.infostat.com.ar (accessed on 30 January 2025).
- Bonfim, F.C.G.; Dodonov, P.; Cazetta, E. Landscape composition is the major driver of the taxonomic and functional diversity of tropical frugivorous birds. Landsc. Ecol. 2021, 36, 2535–2547. [Google Scholar] [CrossRef]
- Chapman, P.M.; Tobias, J.A.; Edwards, D.P.; Davies, R.G. Contrasting impacts of land-use change on phylogenetic and functional diversity of tropical forest birds. J. Appl. Ecol. 2018, 55, 1604–1614. [Google Scholar] [CrossRef]
- Conti, G.; Enrico, L.; Jaureguiberry, P.; Cuchietti, A.; Lipoma, M.L.; Cabrol, D. The role of functional diversity in the provision of multiple ecosystem services: An empirical analysis in the dry Chaco of Córdoba, central Argentina. Ecosistemas 2018, 27, 60–74. [Google Scholar]
- Villéger, S.; Mason, N.W.H.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Pla, L.; Casanoves, F.; Di Rienzo, J. Quantifying Functional Biodiversity; Springer Briefs in Environmental Science; Springer; Dordrecht, The Netherlands, 2012; Available online: https://link.springer.com/10.1007/978-94-007-2648-2 (accessed on 17 December 2024).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Viena, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 17 December 2024).
- Laliberté, E.; Legendre, P.; Shipley, B. FD-Package Version 1.0 to 12.3: Measuring Functional Diversity (FD) from Multiple Traits, and Other Tools for Functional Ecology; License GPL-2; 2014. Available online: https://www.scirp.org/reference/ReferencesPapers?ReferenceID=1820451 (accessed on 17 December 2024).
- Lepš, J.; De Bello, F. Differences in trait–environment relationships: Implications for community weighted means tests. J. Ecol. 2023, 111, 2328–2341. [Google Scholar] [CrossRef]
- Casanoves, F.; Pla, L.; Di Rienzo, J.A.; Díaz, S. FDiversity: A software package for the integrated analysis of functional diversity. Methods Ecol. Evol. 2011, 2, 233–237. [Google Scholar] [CrossRef]
- Marcolin, F.; Lakatos, T.; Gallé, R.; Batáry, P. Fragment connectivity shapes bird communities through functional trait filtering in two types of grasslands. Glob. Ecol. Conserv. 2021, 28, e01687. [Google Scholar] [CrossRef]
- Balzarini, M.; Gonzalez, L.; Tablada, M.; Casanoves, F.; Di Rienzo, J.; Robledo, C.W. InfoStat Statistical Software: User Manual, 1st ed.; Group InfoStat, FCA, National University of Córdoba: Córdoba, Argentina, 2008; 335p. [Google Scholar]
- Rosenfield, M.F.; Miedema Brown, L.; Anand, M. Increasing cover of natural areas at smaller scales can improve the provision of biodiversity and ecosystem services in agroecological mosaic landscapes. J. Environ. Manag. 2022, 303, 114248. [Google Scholar] [CrossRef]
- Velásquez-Trujillo, V.; Betancurt-Grisales, J.F.; Vargas-Daza, A.M.; Lara, C.E.; Rivera-Páez, F.A.; Fontúrbel, F.E.; Castaño-Villa, G.J. Bird Functional Diversity in Agroecosystems and Secondary Forests of the Tropical Andes. Diversity 2021, 13, 493. [Google Scholar] [CrossRef]
- Luck, G.W.; Carter, A.; Smallbone, L. Changes in Bird Functional Diversity across Multiple Land Uses: Interpretations of Functional Redundancy Depend on Functional Group Identity. PLoS ONE 2013, 8, e63671. [Google Scholar] [CrossRef] [PubMed]
- Carvajal Alfaro, V.; Oviedo, P.E. Composición de aves asociada a fincas de cacao orgánico en Región Huetar Norte, Costa Rica. Uniciencia 2024, 38, 138–146. [Google Scholar] [CrossRef]
- Cabral, J.P.; Faria, D.; Morante-Filho, J.C. Landscape composition is more important than local vegetation structure for understory birds in cocoa agroforestry systems. For. Ecol. Manag. 2021, 481, 118704. [Google Scholar] [CrossRef]
- Coelho, M.T.P.; Raniero, M.; Silva, M.I.; Hasui, É. The effects of landscape structure on functional groups of Atlantic forest birds. Wilson J. Ornithol. 2016, 128, 520–534. [Google Scholar] [CrossRef]
- Gutiérrez García, G.A.; Gutiérrez-Montes, I.; Hernández Núñez, H.E.; Suárez Salazar, J.C.; Casanoves, F. Relevance of local knowledge in decision-making and rural innovation: A methodological proposal for leveraging participation of Colombian cocoa producers. J. Rural. Stud. 2020, 75, 119–124. [Google Scholar] [CrossRef]
- Hernández-Núñez, H.E.; Gutiérrez-Montes, I.; Sánchez-Acosta, J.R.; Rodríguez-Suárez, L.; Gutiérrez-García, G.A.; Suárez-Salazar, J.C.; Casanoves, F. Agronomic conditions of cacao cultivation: Its relationship with the capitals endowment of Colombian rural households. Agrofor. Syst. 2020, 94, 2367–2380. [Google Scholar] [CrossRef]
- Bennett, R.E.; Sillett, T.S.; Rice, R.A.; Marra, P.P. Impact of cocoa agricultural intensification on bird diversity and community composition. Conserv. Biol. 2022, 36, e13779. [Google Scholar] [CrossRef] [PubMed]
- De Moraes, K.F.; Lima, M.G.M.; Gonçalves, G.S.R.; Cerqueira, P.V.; Santos, M.P.D. The future of endemic and threatened birds of the Amazon in the face of global climate change. Ecol. Evol. 2024, 14, e11097. [Google Scholar] [CrossRef]
- Karp, D.S.; Rominger, A.J.; Zook, J.; Ranganathan, J.; Ehrlich, P.R.; Daily, G.C. Intensive agriculture erodes β-diversity at large scales. Ecol. Lett. 2012, 15, 963–970. [Google Scholar] [CrossRef]
- Carucci, T.; Whitehouse-Tedd, K.; Yarnell, R.W.; Collins, A.; Fitzpatrick, F.; Botha, A.; Santangeli, A. Ecosystem services and disservices associated with vultures: A systematic review and evidence assessment. Ecosyst. Serv. 2022, 56, 101447. [Google Scholar] [CrossRef]
- Šekercioğlu, C.H. Bird functional diversity and ecosystem services in tropical forests, agroforests and agricultural areas. J. Ornithol. 2012, 153, 153–161. [Google Scholar] [CrossRef]
- Mayani-Parás, F.; Moreno, C.E.; Escalona-Segura, G.; Botello, F.; Munguía-Carrara, M.; Sánchez-Cordero, V. Classification and distribution of functional groups of birds and mammals in Mexico. PLoS ONE 2023, 18, e0287036. [Google Scholar] [CrossRef] [PubMed]
- Smith, O.M.; Kennedy, C.M.; Echeverri, A.; Karp, D.S.; Latimer, C.E.; Taylor, J.M.; Wilson-Rankin, E.E.; Owen, J.P.; Snyder, W.E. Complex landscapes stabilize farm bird communities and their expected ecosystem services. J. Appl. Ecol. 2022, 59, 927–941. [Google Scholar] [CrossRef]
- Matuoka, M.A.; Benchimol, M.; Almeida-Rocha, J.M.D.; Morante-Filho, J.C. Effects of anthropogenic disturbances on bird functional diversity: A global meta-analysis. Ecol. Indic. 2020, 116, 106471. [Google Scholar] [CrossRef]
- Adelino, J.R.P.; Calsavara, L.C.; Willrich, G.; Rosa, G.L.M.; Lima, M.R.; Dos Anjos, L. Ecosystem functions of birds as a tool to track restoration efficiency in Brazil. Ornithol. Res. 2020, 28, 38–50. [Google Scholar] [CrossRef]
- Gaston, K.J. Birds and ecosystem services. Curr. Biol. 2022, 32, R1163–R1166. [Google Scholar] [CrossRef]
- Karp, D.S.; Mendenhall, C.D.; Sandí, R.F.; Chaumont, N.; Ehrlich, P.R.; Hadly, E.A.; Daily, G.C. Forest bolsters bird abundance, pest control and coffee yield. Ecol. Lett. 2013, 16, 1339–1347. [Google Scholar] [CrossRef]
- Whelan, C.J.; Şekercioğlu, Ç.H.; Wenny, D.G. Why birds matter: From economic ornithology to ecosystem services. J. Ornithol. 2015, 156, 227–238. [Google Scholar] [CrossRef]
- Frenzel, M.; Everaars, J.; Schweiger, O. Bird communities in agricultural landscapes: What are the current drivers of temporal trends? Ecol. Indic. 2016, 65, 113–121. [Google Scholar] [CrossRef]
- Le Clech, S.; Van Bussel, L.G.J.; Lof, M.E.; De Knegt, B.; Szentirmai, I.; Andersen, E. Effects of linear landscape elements on multiple ecosystem services in contrasting agricultural landscapes. Ecosyst. Serv. 2024, 67, 101616. [Google Scholar] [CrossRef]
- Benedetti, Y.; Morelli, F.; Munafò, M.; Assennato, F.; Strollo, A.; Santolini, R. Spatial associations among avian diversity, regulating and provisioning ecosystem services in Italy. Ecol. Indic. 2020, 108, 105742. [Google Scholar] [CrossRef]
- Linden, V.M.G.; Grass, I.; Joubert, E.; Tscharntke, T.; Weier, S.M.; Taylor, P.J. Ecosystem services and disservices by birds, bats and monkeys change with macadamia landscape heterogeneity. J. Appl. Ecol. 2019, 56, 2069–2078. [Google Scholar] [CrossRef]
- Mortimer, R.; Saj, S.; David, C. Supporting and regulating ecosystem services in cacao agroforestry systems. Agrofor. Syst. 2018, 92, 1639–1657. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Description |
|---|---|
| Cocoa agroforestry crops (CA) | Plantations of Theobroma cacao (L.) intercropped with various tree species such as Hevea brasiliensis (Müll. Arg.) and Acacia mangium (Willd.). These systems typically occur on acidic, low-fertility soils—primarily ultisols or oxisols—with good drainage. They feature partial shade that moderates light intensity and promotes a humid microclimate. Water and thermal stress are low, and soil moisture retention is generally high. |
| Wetlands (HU) | Natural freshwater bodies located in flood-prone areas of alluvial terraces or between foothill valleys, featuring aquatic herbaceous vegetation such as Juncus L. These systems are characterized by hydromorphic soils with high water saturation and low oxygen availability, typically gleysols or fluvisols rich in organic matter. Light intensity is generally high, though it varies depending on the density of surrounding vegetation. |
| Early successional fallows (RT) | Shrub-herbaceous vegetation between 2–5 m in height, with dominant dbh < 10 cm, and species from genera such as Miconia (Ruiz and Pav.) and Spathiphyllum (H.W. Schott and S.L. Endlicher). Found on disturbed soils—typically ultisols or inceptisols—with low to moderate fertility and surface compaction. High light intensity favors the regeneration of heliophilous species. |
| Late successional fallows (RV) | Vegetation older than ten years, primarily arboreal, with heights of 8–12 m, dominant dbh > 10 cm, and an irregular canopy, with species such as Ochroma pyramidale (Cav. ex Lam. Urb.) and Piptocoma discolor (Kunth Pruski). Partially recovered soils, such as ultisols or inceptisols, with higher organic matter content. Low light intensity due to the canopy, resulting in a more shaded and humid understory. |
| Clean pastures (PL) | Areas dominated by Urochloa spp. with over 70% ground Cover. Disturbed soils lacking tree cover, typically compacted ultisols or oxisols with low fertility and organic matter content. These areas are exposed to direct solar radiation, resulting in high light intensity and elevated temperatures, which increase evapotranspiration and reduce soil moisture retention. |
| Secondary forest (BS) | Arboreal vegetation with pioneer species, height exceeding 12 m, dbh between 15–20 cm, a multi-layered canopy, and tree cover greater than 30%, with species such as Zygia longifolia (Willd. Britton and Rose) and Ormosia coccinea (Aubl.). Partially recovered soils, typically ultisols or inceptisols with high organic matter content and good water retention. Light intensity is low, with gradients of light that support both heliophilous and shade-tolerant species. |
| Weedy pastures (PE) | Areas with Urochloa spp. mixed with weeds, under 1.5 m in height, including species like Bellucia pentamera (Naudin). Degraded or compacted soils, such as ultisols with low fertility and poor nutrient retention. These areas experience high exposure to solar radiation, with intense light and elevated surface temperatures. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz-Cháux, J.T.; Velasquez-Valencia, A.; Martínez-Salinas, A.; Casanoves, F. Functional Diversity and Ecosystem Services of Birds in Productive Landscapes of the Colombian Amazon. Diversity 2025, 17, 305. https://doi.org/10.3390/d17050305
Díaz-Cháux JT, Velasquez-Valencia A, Martínez-Salinas A, Casanoves F. Functional Diversity and Ecosystem Services of Birds in Productive Landscapes of the Colombian Amazon. Diversity. 2025; 17(5):305. https://doi.org/10.3390/d17050305
Chicago/Turabian StyleDíaz-Cháux, Jenniffer Tatiana, Alexander Velasquez-Valencia, Alejandra Martínez-Salinas, and Fernando Casanoves. 2025. "Functional Diversity and Ecosystem Services of Birds in Productive Landscapes of the Colombian Amazon" Diversity 17, no. 5: 305. https://doi.org/10.3390/d17050305
APA StyleDíaz-Cháux, J. T., Velasquez-Valencia, A., Martínez-Salinas, A., & Casanoves, F. (2025). Functional Diversity and Ecosystem Services of Birds in Productive Landscapes of the Colombian Amazon. Diversity, 17(5), 305. https://doi.org/10.3390/d17050305