Invasion by Exotic Earthworms Alters Biodiversity and Communities of Litter- and Soil-dwelling Oribatid Mites

Abstract
: Exotic earthworms are drivers of biotic communities in invaded North American forest stands. Here we used ecologically important oribatid mite (Arachnida: Acari) communities, as model organisms to study the responses of litter- and soil-dwelling microarthropod communities to exotic earthworm invasion in a northern temperate forest. Litter- and soil-dwelling mites were sampled in 2008–2009 from forest areas: (1) with no earthworms; (2) those with epigeic and endogeic species, including Lumbricus rubellus Hoffmeister; and (3) those with epigeic, endogeic, and anecic earthworms including L. terrestris L. Species richness and diversity of litter- and soil-dwelling (0–2 cm soil depth) oribatid mites was 1–2 times higher in sites without earthworms than in sites with worms. Similarly, litter-dwelling oribatid mites were between 72 and 1,210 times more abundant in earthworm-free sites than in sites with worms. Among earthworm invaded sites, abundance of litter-dwelling oribatid mites in sites without the anecic L. terrestris was twice as high in May and 28 times higher in October, compared to sites with L. terrestris. Species richness, diversity, and abundance of oribatid mites were greater in litter-layers than in the soil-layers that showed a varied response to earthworm invasion. Species compositions of both litter- and soil-dwelling oribatid mite communities of forests with no earthworms were markedly different from those with earthworms. We conclude that exotic earthworm invasions are associated with significant declines of species diversity, numbers, and compositional shifts in litter- and soil-inhabiting communities. These faunal shifts may contribute to earthworm effects on soil processes and food web dynamics in historically earthworm-free, northern temperate forests.1. Introduction
During the last few centuries, northern temperate forests of North America have experienced many anthropogenic disturbances, such as habitat loss and fragmentation, changes in natural disturbances regimes, and the introduction of exotic species [1-4]. For example, exotic earthworms (Oligochaeta: Lumbricidae) native to Europe and Asia, have caused some of the most permanent changes in northern temperate forests [5-7]. Almost all earthworm species present in these once glaciated areas in North America are non-native, as native species were likely eradicated during the last glacial period [8]. Activities by exotic earthworms can lead to abiotic and biotic changes, such as a near absence of litter or organic layer, intermixing of soil layers, altered soil porosity, texture and chemical attributes, increased competition for microbial food resources within two to five years of introduction, and shifts in the composition and abundance of plants and animals [9-13]. Due to the widespread alteration of ecosystem structure and function caused by earthworms, these taxa are generally considered as “ecosystem engineers” in both the native and non-native ranges [6,7,10].
The ecological impacts of exotic earthworms vary depending upon the species. Earthworms are generally classified into three groups: epigeic, endogeic, and anecic [11,14-16]. Epigeic species such as Lumbricus rubellus Hoffmeister are litter-dwelling species that use shallow burrows, and feed on leaf litter and other coarse organic matter. Endogeic species, such as Aporrectodea caliginosa (Savigny) and Octalasion tyrtaeum (Savigny), dwell in deep burrows in the mineral soil where they ingest mineral soil to digest organic matter and microflora. Anecic species such as L. terrestris L. are typically large, and can form deep (1–2 m) vertical burrows; however, like epigeic species, anecic species feed on leaf litter and coarse organic matter at the surface [7]. Amongst the three guilds of earthworms, anecic species cause the greatest soil disturbance (rapid reduction of litter layer and intermixing of the litter and mineral layers), followed by endogeic (disturbance of mineral layer), and epigeic (rapid reduction of litter layer) species [7].
Litter- and soil-dwelling mites are among the most species-rich, widely distributed, and abundant animal taxa. Among mites, oribatid species are a diverse and functionally important group in soil and litter layers [17]. In some instances, over 100 species and 100,000 mites may be found per m2 in northern temperate forests [18]. Oribatid mites are sensitive to changes in physical and chemical habitat conditions, and have limited dispersal abilities [18]. Oribatid mite species are largely particle-feeding saprophagous and mycetophagous, and are important for litter break-down and decomposition on the forest floor [18]. These mites are also important vectors for distribution of fungal spores, some of which are beneficial mycorrhizal species. For example, 31 fungal taxa were associated with oribatid mites, some of which were not common soil species indicating that these fungal species may be transported to new colonization sites by mites and/or their predators [19]. Mites and other microarthropods are also important prey for larger invertebrates and some forest vertebrates such as woodland salamanders [20,21]. Overall, shifts in oribatid mite abundance or composition could have implications for forest nutrient and food web dynamics.
Earthworm activities structurally and functionally alter litter and soil layers, and hence, the habitat of soil microarthropods. There have been a number of studies that have assessed the interactions of earthworms with soil microarthropods such as mites (Arachnida: Acari) and collembolans (Insecta: Collembola) in their native habitats of Europe and Asia [22-28]. These studies reported that earthworm activity may enhance microarthropod diversity and abundance. In contrast, there have been relatively few studies on the interactions between exotic earthworms and soil microarthropods in their non-native habitats in North America [29-31]. These reports indicated that the effects of exotic earthworms are variable, as depending upon the native taxa and geographic location. For example, in a lodgepole pine (Pinus contorta Douglas ex Louden) forest in Alberta, there was an increased species richness and diversity, but decreased density of oribatid mites with high earthworm activity, depending upon the soil layer [29,30]. Octolasion tyrtaeum reduced both species richness and abundance of soil microarthropods in aspen (Populus tremuloides Michx.) forests of the Canadian Rockies [11]. At present, there is little information about how microarthropods may respond to exotic earthworms belonging to different guilds (epigeic, endogeic, and anecic) in northern temperate (Acer saccharum Marsh) forests.
We measured changes in microarthropod communities, specifically of oribatid mites, in habitats invaded by exotic earthworms in a northern hardwood forest. Our specific research objectives were to determine whether: (1) species richness and diversity, abundance, and composition of oribatid mites differed between habitats with and without invasive earthworms; and (2) species richness and diversity, abundance, and composition of oribatid mites differed between earthworm invaded habitats with or without anecic species.
2. Methods
2.1. Study Sites
We sampled soil mites from the 1,700 ha Arnot Teaching and Research Forest, Cornell University (N42°16″ W76°28″) located in the Allegheny Plateau in New York State. Temperatures range from a mean annual summer high of 22 °C to a mean winter low of −4 °C. Annual precipitation is around 1,000 mm, and there is significant snow-cover in winter [32]. The soils in the region are derived from bedrock till, consisting mostly of shales from the Upper Devonian Period [32,33]. Soils are primarily acidic Dystrochrepts and Fragiochrepts of the Lordstown-Volusia-Mardin Series [32,33]. Glaciers played an important role in the erosion of the landscape, and deposited rocks, boulders, sand and clay from ice and glacial melt water [34]. These deposits currently occur as moraines, outwash plains, and valley trains in the Arnot Forest [34]. The forest floor litter is well developed (∼4 cm thick), and overlays an acidic (pH 4.5–5.0) mineral horizon [32,35]. The dominant overstory tree species in the Arnot Forest includes sugar maple, red maple (A. rubrum L.), beech (Fagus grandifolia Ehrh.), white ash (Fraxinus americana L.), basswood (Tilia americana L.), and hemlock (Tsuga canadensis L.) [32].
2.2. Study Design
Study sites were three replicate areas in each of three types of habitats as follows: (1) those with no earthworms (hereafter referred to as “no earthworm site”); (2) those with exotic epigeic and endogeic earthworm species (hereafter referred to as “habitat without anecic earthworms”); and (3) those with epigeic, endogeic, and anecic earthworms including L. terrestris (hereafter referred to as “habitat with anecic earthworms”). Species documented during this study period in the earthworm habitats lacking anecic species were: L. rubellus, O. tyrtaeum, and A. caliginosa (Bohlen et al, unpublished data). We did detect six small juvenile L. terrestris, which are anecic, in one study plot; however, we did not find any adult L. terrestris in the plot during the year prior to this study period, during the study, or in the year subsequent to this study. We speculate that these juveniles dispersed into the plot from nearby habitats with L. terrestris. Species documented during this study period in the earthworm habitats with anecic species were: L. terrestris, L. rubellus, O. tyrtaeum, and A. caliginosa (Bohlen et al, unpublished data).
These sites were part of a larger study examining the impact of earthworm invasions on nutrient dynamics in forest soils, so the methods reflect the larger objectives of that study. Within each study site, eight 1 m × 1.25 m study plots were established. Each plot was surrounded by ∼10 cm tall plastic garden edging flush with the mineral soil, and adjacent plots were separated by >4 m. The edging prevented leaf litter from moving laterally in or out of the plot. A 1 × 0.25 m rough-cut, sugar maple board was placed in the center of each plot for sampling salamanders in a related study, creating two quadrants totaling 1 m2 of exposed forest floor within each plot. In October 2007, fresh litterfall was removed from each quadrant and a coarse-mesh nylon screen (hole size = 6 cm2) was positioned on the underlying forest floor and anchored at the corners. About 400 g (weighed to ±0.01 g) of air-dried sugar maple litter was added to each plot to roughly match leaf litterfall in the study area. A second coarse-mesh screen was positioned on the added litter and anchored to confine the litter and prevent subsequent addition of litterfall. In fall 2008, fresh sugar maple litter was collected in the stands and about 400 g was again added to each plot and secured in place with a third coarse mesh screen. Because standard quantities and composition of litter was added to plots, there was limited potential for any differences in litterfall composition or volume to contribute to differences in microarthropod differences among habitats.
Two plots in each site were destructively sampled on 21 May and 10 October 2008 for a total of six plots per habitat (n = 6) per sampling date. In each plot, all of the litter remaining between screens was placed inside a cotton pillow case and held in a cooler for transport to the lab. Next, the underlying forest floor horizons (Oe + Oa) were collected by excavating with hand spades to the top of the mineral soil. Forest floor horizons were also placed on a cotton pillowcase in a cooler. Finally, mineral soil was cored to 20 cm depth using 5 cm sharpened split-PVC corers. Soil samples from several cores (generally 6–8) were composited for each quadrant by depth. All samples were stored cold until processed within 48 h.
We used Tullgren funnels to extract invertebrates. Leaf litter, organic horizon, or mineral soil samples were placed on top of a fine mesh and suspended under a 60 Watt incandescent bulb for seven days until the sample was completely dry. Invertebrates were collected in 35% EtOH and stored for later identification. All arthropods were sorted to order, and soil mites were separated for identification and enumeration. We sub-sampled catches that had greater than 250 oribatid individuals. To subsample, we divided samples in EtOH into 2 mL samples, and then we stirred and poured each subsample into a 55 cm2 circular petri dish, with a 6 × 6 cm2 grid etched on the bottom. Mites were allowed to settle to the bottom to ensure random but even distribution around the grid. All mites in five randomly selected cells were collected for species identification.
All oribatid mites from these five subsamples were pooled and mounted onto slides for species identification. Oribatid mites were cleared in 85% lactic acid solution for 24–72 hours, depending upon the level of sclerotization of each species. Darker and more robust species were cleared for a longer period of time (∼72 hours). Cleared mites were mounted on cavity slides with 60% sodium lactate syrup solution, and edges of cover-slips sealed with a clear nail-paint. All oribatid mites were identified to species- and genera-level using available taxonomic keys and descriptions [18], and verified by an oribatid mite systematist. The synoptic collections of oribatid mites were deposited at the Georgia Museum of Natural History, University of Georgia, Athens, USA.
2.3. Statistical Analyses
Before analyses, we pooled leaf litter and organic horizon layers (hereafter referred to as litter) for each plot, and we standardized all abundance to m2 for all litter and soil samples. Venn diagrams were used to determine relationships of mite species richness (total number of species) between sampling times and depths, and habitats. We compared oribatid mite species diversity using Shannon-Weiner, Simpson's Diversity (reciprocal form), and Evenness indices [36]. For abundance analyses, data were log-transformed to meet assumptions of normality and homogeneity of variance. To provide resolution on which species accounted for differences in abundance among habitats, we used two-way multiple analysis of variance (MANOVA) to test the effects of habitat and season on the abundance of the three most abundant mite species. Roy's Greatest Root test was used to assess the significance of two factors and their interactions. Standardized canonical coefficients of canonical variates were used to evaluate the relationships between each species' abundance and the main factors [37]. These standardized canonical coefficients were obtained by multiplying each coefficient with standard deviation of the variable [37]. Similar signs (+ or −) refer to variables correlated in a similar way, whereas opposite signs refer to variables correlated in an opposite way. A transformed scale was used to show trends when there was a significant interaction between two factors. Due to differences in collection technique (whole litter extraction versus soil cores) and differences in mite abundance, we conducted separate analyses for litter and soil samples.
Next, we used nonmetric multidimensional scaling (NMS) to compare mite communities [38,39]. Data were first checked for skewness, kurtosis, and coefficients of variation (CVs). As CV was >100%, data were log-transformed. A preliminary NMS ordination was done using Sorenson distance measure on six axes using random coordinates, 15 iterations to evaluate stability, stability criterion of 0.0005, 250 iterations at 0.2 step lengths, step down in dimensionality, and 50 runs for real data. Final stress versus the number of dimensions was plotted to find the correct number of dimensions, and stability of the solution was checked using the plot of stress versus number of iterations. Final NMS ordination analysis was run using three dimensions, 1 run of real data, and 250 iterations. The final stress was 11.15 and 12.36 for litter- and soil-dwelling mites, respectively which is intermediate (10–20) for most community data sets, and the final stability was ∼0.00045 [39]. An ordination graph was created using the means (±SE) for ordination scores for each combination of sampling times, sampling depths, and habitats along the two hypothetical axes that had the highest R2 value.
3. Results
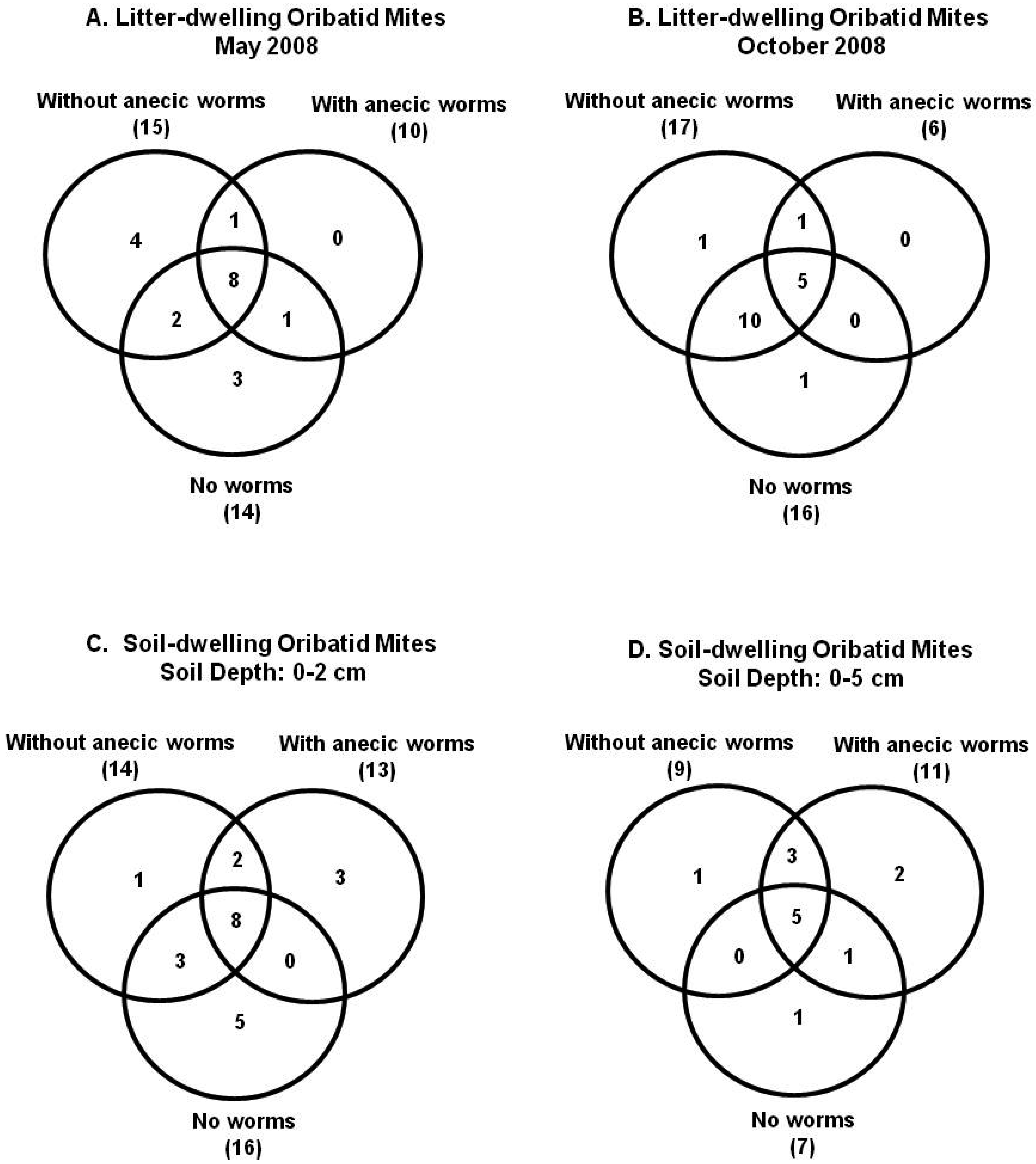
For litter-layer, a total number of 36,221 oribatid mite individuals belonging to 23 species were identified (Supplement A, Table 1). Three species accounted for 89% of all oribatid mite individuals in the study. Scheloribates sp.1 was most abundant (45% of total mite individuals) followed by Oribatula tibialis (Nicolet) (30%) and Parachipteria sp. 1 (14%) (Supplement A). Venn diagrams for May sampling for litter-dwelling mites indicated that similar numbers of unique species were present among the three habitats (Figure 1A). However, in October, more species were shared between no earthworm and earthworm invaded habitats without anecic species. We did not find any oribatid mite species unique to habitats with anecic earthworm (Figure 1B). Species richness, diversity, and evenness in earthworm-invaded habitats were half of those in the non-invaded habitats (Table 1). Oribatid mite species diversity was higher in October than in May, and diversity differences among habitats were greater in October than in May (Table 1).
For soil-layer, 1,457 oribatid mite individuals belonging to 23 species were recorded (Supplement B). Three species accounted for 77% of all individuals. Gemmazetes crosbyi (Berlese) was the most abundant species (38.8% of total mite individuals), followed by Scheloribates sp. 1 (24.4%), and Oppiella nova (Oudemans) (13.37%) (Supplement B). Venn diagrams for 0–2 cm soil layer indicated that the no earthworm habitats had the most unique species and the highest species richness (Figure 1C). About 35% of species were shared between all three habitats. In contrast, 0–5 cm soil depth had about an equal number of shared species with the highest species richness in habitats with anecic earthworm (Figure 1D). About 22% of species were shared between all the sites. Species richness and diversity was about 1–1.5 times higher in habitats without earthworms in 0–2 cm depth, and in habitats with anecic worms in 0–5 cm soil depth (Table 2). Evenness of oribatid soil communities was 1.2 times higher in habitats without anecic worms in 0–2 cm depth, and in no worm habitats in 0–5 cm soil depth (Table 2).
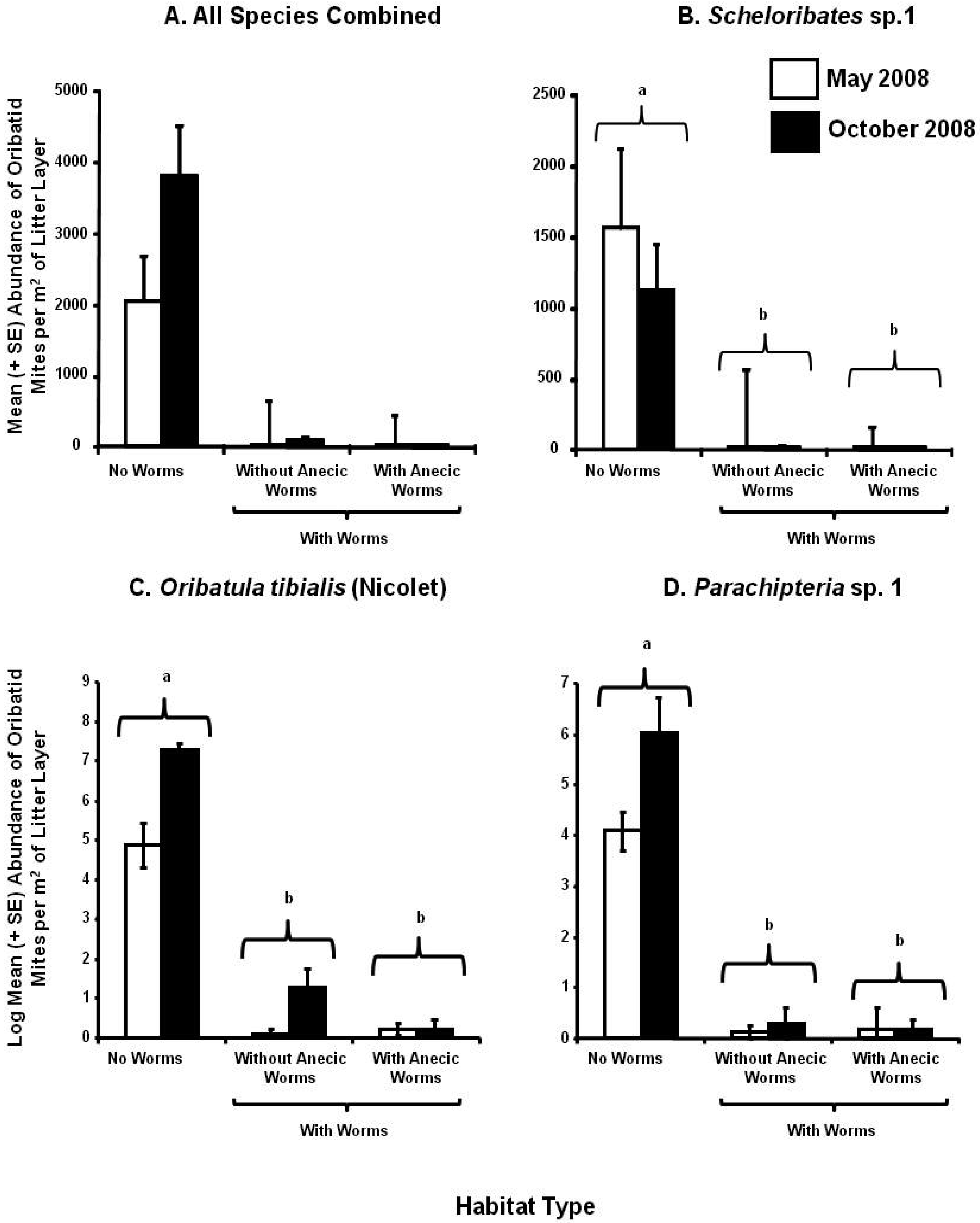
Roy's Greatest Root for these three most abundant species indicated that there was a significant interaction between sampling time and habitat for litter-dwelling oribatid mites (Table 3). Depending on the season, there were 72–1,210 times more oribatid mites per m2 in the no earthworm habitats than in habitats with earthworms (Figure 2A). There were 2–28 times more oribatid mites found in habitats without anecic earthworms than in habitats with anecic earthworms (Figure 2A). We found significantly greater numbers of Scheloribates sp.1 in no worm than in worm invaded habitats, with 92 and 576 times more individuals in habitats without and with anecic earthworms, respectively (Figure 2B; Supplement A). Both O. tibialis and Parachipteria sp. 1 were more abundant in no-worm than in worm habitats (8–26 and 24–28 times, respectively) (Figure 2C, D). About 1.5–2 times more individuals of these two species were caught in October than in May samples (Figure 2C, D). The abundance of Scheloribates sp.1 was negatively associated with the other two species, whereas O. tibialis and Parachipteria sp. 1 were positively associated with each other (Table 3).
Roy's Greatest Root indicated that habitat was the only significant factor explaining differences in the abundances of the three most abundant soil-dwelling mite species (Table 4); however, differences among habitats were relatively small compared to differences among habitats seen in the litter-layer mite abundances (Figure 3A). About 2.5–3 times more G. crosbyi were found in soil habitats without anecic earthworms as compared to the other two habitats (Figure 3B). There were no significant differences in the abundances of Scheloribates sp. 1 or O. nova between habitats (Figure 3C, D). However, five times more O. nova were found in 0–2 than in 0–5 cm soil depth (Figure 3D).
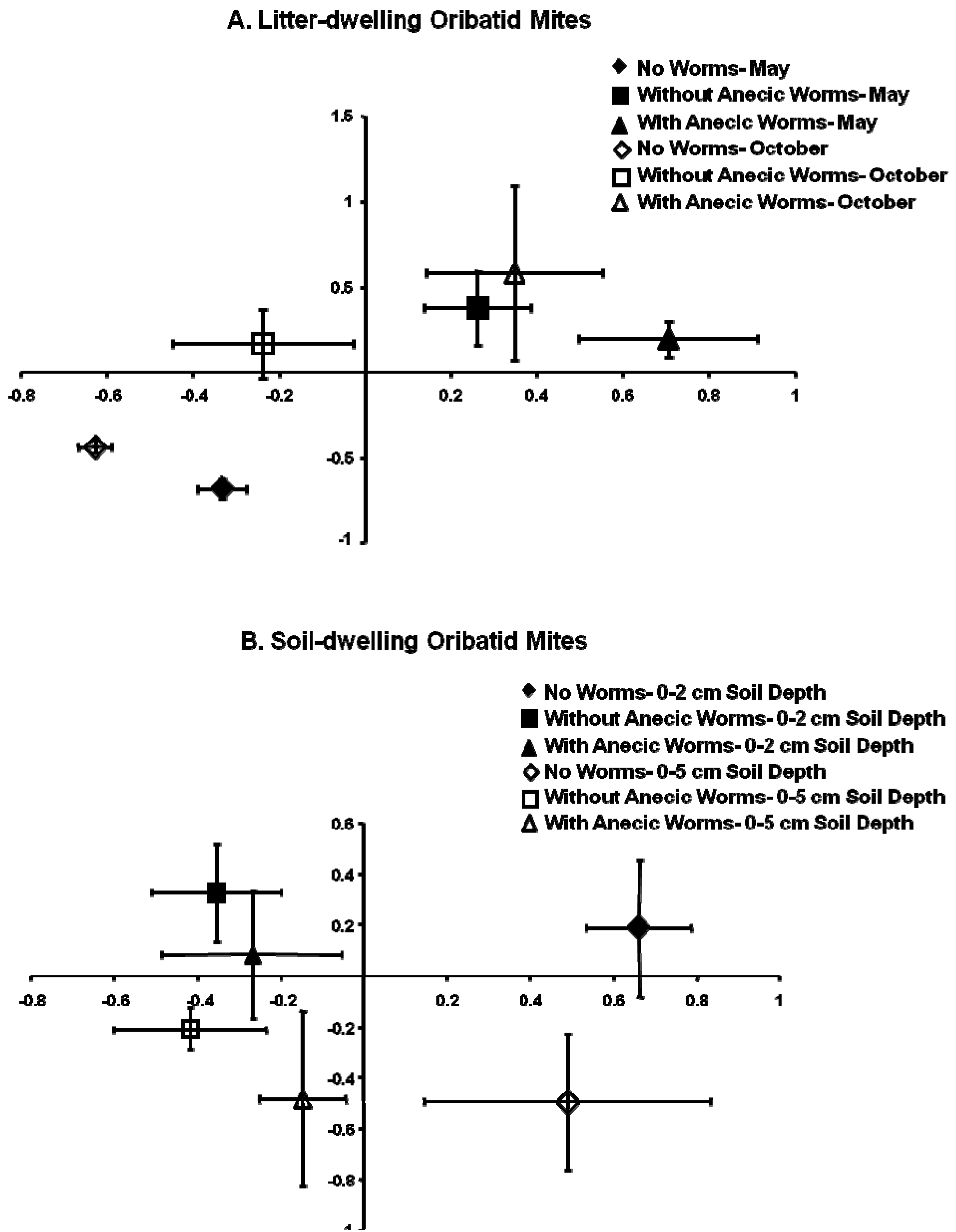
The ordination plot for litter-dwelling oribatid mites constructed using NMS with the plot scores showed that no earthworm habitats had the most distinct oribatid mite communities (R2 for axis 1 = 24%, R2 for axis 2= 36%, r = 0.426) (Figure 4A). Earthworm invaded habitats were arranged at the opposite end of the hypothetical gradient with no overlap along the y-axis. Habitats without anecic earthworms in October were similar to mite composition with habitats without earthworms along the x-axis, and there was considerable overlap in species composition between habitats with and without anecic earthworms (Figure 4A). Mite communities in no earthworm habitats differed between May and October, indicating species turnover with time; however, species composition of two sampling periods for both earthworm habitats were generally similar to each other, along one axis (Figure 4A).
The ordination graph for soil-dwelling oribatid mite communities created from mean NMS scores for the first two axes (R2 for axis 1 = 17%, R2 for axis 2 = 25%, r = −0.12) showed that species composition of oribatid mites for the no worm habitats at two depths were distinct as they were arranged on the opposite end of the hypothetical gradient (Figure 4B). In contrast, oribatid species composition of earthworm-invaded habitats at two soil depths was quite similar to each other, as there was a considerable overlap. There was some separation of the two soil depths along the y-axis (Figure 4B).
4. Discussion
Our results indicated that the invasion by earthworms changed the richness, diversity, abundance, and composition of oribatid mites in a northern temperate forest. The five major trends evident in this study were: (1) species richness and diversity of litter- and soil-mites (only 0–2 cm depth) were reduced by earthworm invasion; (2) abundance of litter-dwelling mites including the three most abundant species was reduced by several orders of magnitude by earthworm invasion; (3) total numbers of soil-dwelling oribatid mites was lower than that of litter-dwelling mites, and species varied in their response to earthworm invasion within the soil; (4) earthworm free habitats had a more distinct oribatid mite community than earthworm invaded habitats; and (5) between earthworm invaded habitats, those with anecic species had the least diverse communities and lowest number of mites; however, differences between worm habitats with and without anecic species were small compared to differences between habitats with and without earthworms.
Other studies have documented mixed results regarding interactions between earthworms and soil microarthropods. For example, in boreal lodgepole pine [30] and aspen [11] forests, epigeic exotic earthworms depressed the diversity and densities of oribatid mites. Some of these results were dependent upon the soil layer sampled, as there were high oribatid mite species richness and diversity in the L layer compared to the FH layer [30]. Addition of carbon and nitrogen to beech (Fagus sylvatica L.) forests in Germany reduced density of mites and Collembola, perhaps due to the enhanced activity and biomass of earthworms [40]. In a microcosm study, earthworms reduced the numbers of microarthropods such as Collembola and Acari [27]. In contrast, middens or defecated soil and litter at the mouths of earthworm burrows (L. terrestris) had greater numbers of Collembola and prostigmatid mites associated with them (results dependent upon habitat-type and season) within white ash dominated forests of Pennsylvania [41]. There was higher diversity of microarthropods, especially larger-sized Collembola in high earthworm density patches in the French West Indies [23]. Such trends suggest that the effects of native and exotic earthworms are highly varied, based on site at various scales, time since invasion and of sampling, and the study taxa.
The adverse effects of exotic earthworms were evident in the three most abundant litter-dwelling species: Scheloribates sp.1, O. tibialis, and Parachipteria sp. 1, where the response of the latter two species varied depending on time of sampling. These three species belong to oribatid families (Scheloribatidae, Oribatulidae, and Achipteriidae, respectively) that are species-rich and among the most common and widely distributed taxa in soil- and litter-layers [18]. Since most oribatid mites have long life-cycles and low reproductive potential [18], such changes to densities of these oribatid mites may be relatively long-term. At present, it is unclear whether these oribatid mite communities may be slow to recover from earthworm-caused disturbances. Such effects can create a cascade effect within ecosystems, as these microarthropod species are known to be important prey items for amphibian and insect species [12,18,20,21].
The activity of earthworms may have adversely affected oribatid mite communities through abiotic and biotic alterations at the soil-litter interface in these forest stands. Some of these major abiotic alterations may include creation of burrows with mucus lining, increased soil porosity through formation of macropores, changes in soil texture, chemistry, moisture, and temperature, and reduced habitat through shredding, and thinning of litter layer [42,43]. For example, depth of litter or organic layer is known to positively affect oribatid mite species [44]. Some of the major biotic factors may be due to increased predation by mesostigmatid mites and pseudoscorpions because of larger pores in the soil [45], ingestion and direct mortality of mites by earthworms, and loss/competition for fungi and other micro-organisms as a food source. Abundant oribatid species in our study such as O. tibialis are known to be a selective feeder on ectomycorrhizal fungi [46], and hence, competition and loss of their food source may have led to substantial reductions in their abundance. Earthworms indirectly led to increased activity of soil microorganisms due to mixing of organic into mineral layer [47]. Similarly, exotic earthworms may lower the fungi:bacteria ratio in the forest soils [48]. It is also possible that greater presence of bacteria may have indirectly or directly led to further loss of food sources for native oribatid mite species [48].
In terms of species richness and diversity (0–5 cm depth), and abundance, soil-dwelling oribatid mite species responded differently than litter-dwelling species, especially Scheloribates sp. 1 that was present in both the two soil layers. Although we caught 25 times fewer mites in the soil than in the litter layer, total abundance of soil-dwelling mites did not differ between earthworm and no earthworm sites indicating that other small-scale abiotic factors may be more important for these fauna. Additionally, more G. crosbyi were found in the soil of the earthworm invaded habitats (without anecic species) compared to no worm habitats, suggesting that their abundance in the soil was enhanced by earthworm activities [41]. It is possible that G. crosbyi is surviving well in the new micro-habitats created by earthworms, such as increased organic material in soil layer, can better escape predation by other arthropods such as centipedes and pseudoscorpions, and/or is attracted to and feeds on earthworm mucus and urine as documented for some Collembola species [24,26,28]. The abundance of one common species in the soil, O. nova, was not affected by the presence of earthworms. Oppiella nova is parthenogenetic, long-lived, and one of the most widely distributed mite species around the world [49]. It is known to be a numerically dominant species in a variety of ecosystems, and is generally considered to be disturbance-adapted [50-53]. Similar to our study, O. nova was not affected by soil compaction after forest harvesting [52], unresponsive to habitat simplification through litter alterations [50], and positively affected by younger stand ages [54] in European and North American forests. The effects observed in our study may be early in the stage of earthworm invasion, and there may be greater cumulative impacts over time. Continued sampling is needed to assess whether and how long it would take for soil mites to be affected by earthworms within these forest stands.
Species compositions of both litter- and soil-dwelling oribatid mites were affected by the presence of exotic earthworms. Mite communities in the no earthworm habitats were distinct from earthworm-dominated sites. In both the soil and litter samples, no-worm habitats were generally arranged on the opposite end of the hypothetical environmental gradient to that of the earthworm-invaded habitats. We speculate that habitat specialist species are perhaps being replaced by disturbance-adapted species such as O. nova within earthworm-infested sites. Such a turnover of oribatid mite communities may be permanent as earthworm densities increase and disperse within forest stands [55]. Thus, successional dynamics of litter- and soil-dwelling oribatid mite communities in earthworm-dominated stands will differ considerably from stands that are earthworm free with perhaps ensuing impacts on forest processes over time.
5. Conclusions
As “ecosystem engineers”, exotic invasive species of European and Asian earthworms have significantly impacted forest structure and function in North American forest soils [6,7,10]. Overall, our study provides evidence that exotic earthworms present in a northern hardwood forest stand have altered species richness and diversity, abundance, and composition of oribatid mites. The mechanisms for such changes to mites are unknown at the present time; however, other studies have clearly documented that earthworms affect abiotic and biotic components on forest floor [14,47]. Such impacts on microarthropod communities are likely to be long-term and have cascading ecological impacts on other inter-dependent taxa within forest stands [10,12]. Hence, to maintain forest biodiversity, effective quarantine, and eradication, containment measures may be needed to reduce the current population levels and prevent the spread of exotic earthworms in the non-invaded forest stands [55,56].





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Month (2008) | Treatment Type | Species Richness | Evenness | Shannon-Weiner's Diversity Index | Simpson's Diversity Index |
|---|---|---|---|---|---|
| May | Without worms | 6.83 ± 0.79 | 0.97 ± 0 | 1.83 ± 0.12 | 0.82 ± 0.02 |
| Without anecic worms | 5.0 ± 1.44 | 0.78 ± 0.16 | 1.28 ± 0.32 | 0.62 ± 0.13 | |
| With anecic worms | 3.83 ± 1.01 | 0.81 ± 0.16 | 1.08 ± 0.30 | 0.57 ± 0.13 | |
| October | Without worms | 11.0 ± 0.86 | 0.97 ± 0 | 2.31 ± 0.09 | 0.89 ± 0 |
| Without anecic worms | 7.5 ± 0.89 | 0.96 ± 0 | 1.90 ± 0.11 | 0.84 ± 0.02 | |
| With anecic worms | 2.0 ± 0.47 | 0.48 ± 0.23 | 0.53 ± 0.25 | 0.32 ± 0.15 |
| Sampling Depth (cm) | Treatment Type | Species Richness | Evenness | Shannon-Weiner's Diversity Index | Simpson's Diversity Index |
|---|---|---|---|---|---|
| 0–2 | Without worms | 6.33 ± 1.23 | 0.73 ± 0.04 | 1.30 ± 0.16 | 0.62 ± 0.06 |
| Without anecic worms | 6.0 ± 0.26 | 0.63 ± 0.08 | 1.13 ± 0.16 | 0.55 ± 0.07 | |
| With anecic worms | 4.67 ± 0.95 | 0.75 ± 0.03 | 1.06 ± 0.15 | 0.57 ± 0.05 | |
| 0–5 | Without worms | 3.33 ± 0.67 | 0.88 ± 0.12 | 0.99 ± 0.21 | 0.57 ± 0.09 |
| Without anecic worms | 5.0 ± 0.58 | 0.78 ± 0.05 | 1.26 ± 0.17 | 0.64 ± 0.07 | |
| With anecic worms | 6.0 ± 2.0 | 0.87 ± 0.05 | 1.36 ± 0.35 | 0.68 ± 0.10 |
| A. Roy's Greatest Root | |||||
|---|---|---|---|---|---|
| Source | Value | F-value | Num DF | Den DF | P-value |
| Habitat | 16.234 | 156.93 | 3 | 29 | <0.001 |
| Sampling Time | 1.324 | 12.36 | 3 | 28 | <0.001 |
| Habitat × Sampling Time | 0.619 | 5.99 | 3 | 29 | 0.003 |
| B. Standardized Canonical Coefficients | |||||
|---|---|---|---|---|---|
| Source | Scheloribates sp.1 | Oribatula tibialis | Parachipteria sp.1 | ||
| Habitat | 0.68 | 2.209 | 1.146 | ||
| Sampling Time | −2.48 | 4.0239 | 0.676 | ||
| Habitat × Sampling Time | −1.489 | 3.416 | 1.351 | ||
| A. Roy's Greatest Root | |||||
|---|---|---|---|---|---|
| Source | Value | F-value | Num DF | Den DF | P-value |
| Habitat | 1.235 | 8.23 | 3 | 20 | <0.001 |
| Sampling Time | 0.460 | 2.91 | 3 | 19 | 0.061 |
| Habitat × Sampling Time | 0.340 | 2.27 | 3 | 20 | 0.112 |
| B. Standardized Canonical Coefficients | |||
|---|---|---|---|
| Source | Gemmazetes crosbyi | Scheloribates sp. 1 | Oppiell a nova |
| Habitat | 1.387 | −0.230 | −0.519 |
| Sampling Time | 0.182 | 0.617 | 0.804 |
| Habitat × Sampling Time | −0.353 | 1.047 | 0.379 |
| Oribatid Mite Species | May 2008 | October 2008 | Total Numbers | ||||
|---|---|---|---|---|---|---|---|
| Without Worms | Without Anecic Worms | With Anecic Worms | Without Worms | Without Anecic Worms | With Anecic Worms | ||
| Carabodes niger (Banks, 1895) | 55 | 0 | 0 | 77 | 2 | 0 | 134 |
| Damaeus angustipes (Banks, 1905) | 33 | 3 | 1 | 363 | 116 | 0 | 516 |
| Damaeus sp. 1 | 0 | 0 | 0 | 44 | 1 | 0 | 45 |
| Eulohmannia ribagai (Berlese, 1910) | 0 | 1 | 0 | 0 | 0 | 0 | 1 |
| Galumna sp. 1 | 0 | 7 | 0 | 11 | 34 | 0 | 52 |
| Galumna sp. 2 | 11 | 2 | 5 | 0 | 0 | 0 | 18 |
| Gemmazetes crosbyi (Berlese, 1908) | 0 | 0 | 0 | 22 | 40 | 0 | 62 |
| Hypochthonius sp. 1 | 0 | 1 | 1 | 0 | 0 | 0 | 2 |
| Metabelba sp. 1 | 77 | 4 | 0 | 286 | 67 | 0 | 434 |
| Nothrus palustris (Koch, 1876) | 0 | 34 | 0 | 0 | 0 | 0 | 34 |
| Oppia sp. 1 | 0 | 2 | 0 | 0 | 0 | 0 | 2 |
| Oppiella nova (Oudemans, 1902) | 33 | 1 | 0 | 209 | 47 | 0 | 290 |
| Oribatella sp. 1 | 0 | 0 | 0 | 165 | 4 | 0 | 169 |
| Oribatula tibialis (Nicolet, 1855) | 1375 | 1 | 2 | 9262 | 33 | 3 | 10676 |
| Parachipteria sp. 1 | 495 | 1 | 2 | 4532 | 5 | 2 | 5037 |
| Phthiracarus sp. 1 | 33 | 17 | 7 | 132 | 44 | 6 | 239 |
| Phthiracarus sp. 2 | 11 | 17 | 7 | 0 | 6 | 6 | 47 |
| Platynothrus peltifer (Koch, 1839) | 22 | 0 | 0 | 176 | 1 | 0 | 199 |
| Rhysotritia ardua (Koch, 1841) | 33 | 22 | 19 | 242 | 6 | 1 | 323 |
| Scheloribates sp.1 | 9394 | 59 | 27 | 6809 | 118 | 1 | 16408 |
| Tectocepheus sp. 1 | 0 | 0 | 0 | 0 | 1 | 0 | 1 |
| Tectocepheus velatus (Michael, 1880) | 814 | 0 | 2 | 187 | 1 | 0 | 1004 |
| Trhypochthonius americanus (Ewing, 1908) | 55 | 0 | 0 | 473 | 0 | 0 | 528 |
| Total Number of Individuals | 12441 | 172 | 73 | 22990 | 526 | 19 | 36221 |
| Total Number of Species | 14 | 15 | 10 | 16 | 17 | 6 | 23 |
| Oribatid Mite Species | Soil Depth: 0-2 cm | Soil Depth: 0-5 cm | Total Numbers | ||||
|---|---|---|---|---|---|---|---|
| Without Worms | Without Anecic Worms | With Anecic Worms | Without Worms | Without Anecic Worms | With Anecic Worms | ||
| Belba sp. 1 | 0.00 | 1.41 | 1.41 | 0.00 | 0.00 | 0.00 | 2.83 |
| Carabodes niger (Banks, 1895) | 0.00 | 0.00 | 1.41 | 0.00 | 0.00 | 0.00 | 1.41 |
| Ceratozetes sp. 1 | 11.31 | 7.07 | 5.66 | 2.94 | 6.85 | 5.87 | 39.70 |
| Damaeus angustipes (Banks, 1905) | 0.00 | 0.00 | 0.00 | 0.00 | 0.98 | 0.00 | 0.98 |
| Eulohmannia ribagai (Berlese, 1910) | 0.00 | 0.00 | 1.41 | 0.98 | 0.98 | 2.94 | 6.31 |
| Furcoribula furcillata (Nordenskiold, 1901) | 24.04 | 4.24 | 33.94 | 1.96 | 0.98 | 3.92 | 69.08 |
| Galumna sp. 1 | 0.00 | 0.00 | 1.41 | 0.00 | 0.00 | 0.00 | 1.41 |
| Galumna sp. 2 | 0.00 | 1.41 | 4.24 | 0.00 | 0.00 | 0.00 | 5.66 |
| Gemmazetes crosbyi (Berlese, 1908) | 57.98 | 330.91 | 114.55 | 0.00 | 37.20 | 24.48 | 565.11 |
| Hypochthonius sp. 1 | 0.00 | 7.07 | 0.00 | 0.00 | 0.00 | 0.00 | 7.07 |
| Nothrus anauniensis (Canestrini and Fanzago, 1876) | 11.31 | 0.00 | 0.00 | 0.98 | 0.00 | 0.98 | 13.27 |
| Oppia sp. 1 | 12.73 | 4.24 | 0.00 | 0.00 | 0.00 | 0.00 | 16.97 |
| Oppiella nova (Oudemans, 1902) | 97.58 | 29.70 | 50.91 | 0.98 | 1.96 | 13.71 | 194.83 |
| Oribatula tibialis (Nicolet, 1855) | 14.14 | 5.66 | 0.00 | 1.96 | 0.00 | 0.00 | 21.76 |
| Parachipteria sp. 1 | 14.14 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 14.14 |
| Phthiracarus sp. 1 | 1.41 | 9.90 | 1.41 | 0.00 | 0.00 | 0.00 | 12.73 |
| Phthiracarus sp. 2 | 1.41 | 35.35 | 4.24 | 0.00 | 5.87 | 12.73 | 59.61 |
| Plasmobates sp. 1 | 19.80 | 0.00 | 0.00 | 0.00 | 0.00 | 1.96 | 21.76 |
| Platynothrus peltifer (Koch, 1839) | 2.83 | 1.41 | 0.00 | 0.00 | 0.00 | 0.00 | 4.24 |
| Rhysotritia ardua (Koch, 1841) | 1.41 | 0.00 | 0.00 | 0.00 | 0.98 | 0.98 | 3.37 |
| Scheloribates sp. 1 | 158.38 | 66.46 | 76.36 | 8.81 | 22.52 | 23.50 | 356.04 |
| Tectocepheus velatus (Michael, 1880) | 9.90 | 7.07 | 19.80 | 0.00 | 0.00 | 0.98 | 37.75 |
| Trhypochthonius americanus (Ewing, 1908) | 1.41 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.41 |
| Total Number of Individuals | 439.80 | 511.92 | 316.77 | 18.60 | 78.32 | 92.03 | 1457.44 |
| Total Number of Species | 16 | 14 | 13 | 7 | 9 | 11 | 23 |
Acknowledgments
We thank L. Ogden, V. Kinney, J. DeVore, T. Luhring, and K. Cecala for assistance in the field and laboratory. J. Battigelli (Earthworks, Edmonton) provided guidance about slide preparations of oribatid mites. We are especially grateful to R. Norton (State University of New York, Syracuse) for verifying our species identifications, L. Ogden (University of Georgia) for formatting the manuscript, and anonymous reviewers for providing useful comments that improved the manuscript. Funding was provided by the National Science Foundation, and Daniel B. Warnell School of Forestry and Natural Resources, the University of Georgia, Athens.
References
- Lovett, G.M.; Canham, C.D.; Arthur, M.A.; Weathers, K.C.; Fitzhugh, R.D. Forest ecosystem responses to exotic pests and pathogens in eastern North America. Biosci. 2006, 56, 395–405. [Google Scholar]
- Liebhold, A.M.; MacDonald, W.L.; Bergdahl, D.; Mastro, V.C. Invasion by exotic forest pests: a threat to forest ecosystems. For. Sci. Mono. 1995, 30, 1–49. [Google Scholar]
- Vitousek, P.; D'antonio, C.; Loope, L.; Westbrooks, R.L. Biological invasions as global environmental change. Am. Sci. 1996, 84, 468–478. [Google Scholar]
- Gandhi, K.J.K.; Herms, D.A. Direct and indirect effects of invasive exotic insect herbivores on ecological processes and interactions in forests of eastern North America. Biol. Inv. 2010, 12, 389–405. [Google Scholar]
- Brown, G.G. How do Earthworms Affect Microfloral and Faunal Community Diversity? Plant Soil. 1995, 170, 209–231. [Google Scholar]
- Bohlen, P.J.; Groffman, P.M.; Fahey, T.J.; Fisk, M.C.; Suearez, E.; Pelletier, D.M.; Fahey, R.T. Ecosystem consequences of exotic earthworm invasion of north temperate forests. Ecosystems 2004, 7, 1–12. [Google Scholar]
- Bohlen, P.J.; Scheu, S.; Hale, C.M.; McLean, M.A.; Migge-Kleian, P.M.; Groffman, P.M.; Parkinson, D. Non-native invasive earthworms as agents of change in northern temperate forests. Front. Ecol. Environ. 2004, 2, 427–435. [Google Scholar]
- Gates, G.E. Farewell to North American megadriles. Mega 1982, 4, 12–77. [Google Scholar]
- Frelich, L.E.; Hale, C.M.; Scheu, S.; Holdsworth, A.R.; Heneghan, L.; Bohlen, P.J.; Reich, P.B. Earthworm invasion into previously earthworm-free temperate and boreal forests. Biol. Inv. 2006, 8, 1235–1245. [Google Scholar]
- Migge-Kleian, S.; McLean, M.A.; Maerz, J.C.; Heneghan, L. The influence of invasive earthworms on indigenous fauna in ecosystems previously uninhabited by earthworms. Biol. Inv. 2006, 8, 1275–1285. [Google Scholar]
- Eisenhauer, N.S.; Partsch, S.; Parkinson, D.; Scheu, S. Invasion of a deciduous forest by earthworms: Changes in soil chemistry, microflora, microarthropods, and vegetation. Soil Biol. Biochem. 2007, 39, 1099–1110. [Google Scholar]
- Maerz, J.; Nuzzo, V.; Blossey, B. Declines in woodland salamander abundance associated with non-native earthworm and plant invasions. Conserv. Biol. 2009, 23, 975–981. [Google Scholar]
- Nuzzo, V.A.; Maerz, J.C.; Blossey, B. Earthworm invasion as the driving force behind plant invasion and community change in northeastern North American Forests. Conserv. Biol. 2009, 23, 966–974. [Google Scholar]
- Hale, C.L.; Frelich, L.; Reich, P. Exotic European earthworm invasion dynamics in northern hardwood forests of Minnesota, USA. Ecol. Appl. 2005, 15, 848–860. [Google Scholar]
- Eisenhauer, N. The action of animal ecosystem engineer: identification of the main mechanisms of earthworm impacts on soil microarthropods. Pedobiologia 2010, 53, 343–352. [Google Scholar]
- Staube, D.; Johnson, E.A.; Parkinson, D.; Scheu, S.; Eisenhauer, N. Nonlinearity of effects of invasive ecosystem engineers on abiotic soil properties and soil biota. Oikos 2009, 118, 885–896. [Google Scholar]
- Wallwork, J.A. Oribatids in forest ecosystems. Annu. Rev. Entomol. 1983, 28, 109–130. [Google Scholar]
- Norton, R.; Behan-Pelletier, V. Suborder Oribatida. In A manual of Acarology; Krantz, G.W., Walter, D.E., Eds.; Texas Tech University Press: Texas, TX, USA, 2009; pp. 430–564. [Google Scholar]
- Renker, C.P.; Otto, P.; Schneider, K.; Zimdars, B.; Maraun, M.; Buscot, F. Oribatid mites as potential vectors for soil microfungi: study of mite-associated fungal species. Micro. Ecol. 2005, 50, 518–528. [Google Scholar]
- Burton, T.M. An analysis of the feeding ecology of salamanders (Amphibia, Urodela) of the Hubbard Brook Experimental Forest, New Hampshire. J. Herp. 1976, 10, 187–204. [Google Scholar]
- Maglia, A.M. Ontogeny and feeding ecology of the red-backed salamander. Plethodon cinereus. Copeia. 1996, 3, 576–586. [Google Scholar]
- Wickenbrock, L.; Heisler, C. Influence of earthworm activity on the abundance of collembola in soil. Soil Biol. Biochem. 1997, 29, 517–521. [Google Scholar]
- Loranger, G.; Ponge, J.F.; Blanchart, E.; Lavelle, P. Impact of earthworms on the diversity of microarthropods in a vertisol (Martinique). Biol. Fertil. Soils. 1998, 27, 21–26. [Google Scholar]
- Salmon, S. Earthworm excreta (mucus and urine) affect the distribution of springtails in forest soils. Biol. Fertil. Soils. 2001, 34, 304–310. [Google Scholar]
- Salmon, S. The impact of earthworms on the abundance of Collembola: improvement of food resources or of habitat? Biol. Fertil. Soils. 2004, 40, 323–333. [Google Scholar]
- Salmon, S.; Geoffroy, J.J.; Ponge, J.F. Earthworms and collembola relationships: effects of predatory centipedes and humus forms. Soil Biol. Biochem. 2005, 37, 487–495. [Google Scholar]
- Gutiérrez López, M.; Matesanz, M.R.; Lidon, J.B.J.; Cosin, D.J.D. The effect of Hormogaster elisae (Hormogastridae) on the abundance of soil Collembola and Acari in laboratory cultures. Biol. Fert. Soils. 2003, 37, 231–236. [Google Scholar]
- Salmon, S.; Ponge, J.F. Earthworm excreta attract soil springtails: laboratory experiments on Heteromurus nitidus (Collembola: Entomobryidae). Soil Biol. Biochem. 2001, 33, 1959–1969. [Google Scholar]
- McLean, M.A.; Parkinson, D. Impacts of the epigeic earthworm Dendrobaena octaedra on oribatid mite community diversity and microarthropod abundances in pine forest floor: a mesocosm study. Appl. Soil Ecol. 1998, 7, 125–136. [Google Scholar]
- McLean, M.A.; Parkinson, D. Introduction of the epigeic earthworm Dendrobaena octaedra changes the oribatid community and microarthropod abundances in a pine forest. Soil Biol. Biochem. 2000, 32, 1671–1681. [Google Scholar]
- Parkinson, D.; McLean, M.A. Impacts of earthworms on the community structure of other biota in forest soils. In Earthworm Ecology; Edwards, C.A., Ed.; Soil and Water Conservation Society: Ankeny, IA, USA, 1998; pp. 213–226. [Google Scholar]
- Fain, J.J.; Volk, T.A.; Fahey, T.J. Fifty years of change in an upland forest in south-central New York: general patterns. Bull. Torr. Bot. Club. 1994, 121, 130–139. [Google Scholar]
- Cline, M. Soils and soil associations of New York; Extension Bulletin 930: New York, NY, USA, 1970. [Google Scholar]
- Odell, A.L.; Lassoie, J.P.; Murrow, R.R. A History of Cornell University's Arnot Forest; Cornell Univiversity: New York, NY, USA, 1980. [Google Scholar]
- Lawrence, B.M.; Fisk, M.C.; Fahey, T.J.; Suarez, E.R. Influence of nonnative earthworms on mycorrhizal colonization of sugar maple (Acer saccharum). New Phytol. 2003, 157, 145–153. [Google Scholar]
- Magurran, A. Measuring Biological Diversity; Black. Pub. Co.: Oxford, UK, 2004. [Google Scholar]
- Scheiner, S.M. MANOVA: Multiple response variables and multispecies interactions. In Design and Analysis of Ecological Experiments; Scheiner, S., Gurevitch, J., Eds.; Chapman & Hall/CRC: New York, NY, USA, 1993; pp. 96–112. [Google Scholar]
- McCune, B.; Mefford, M. PC-Ord. Multivariate analysis of ecological data, version 4; 1999. [Google Scholar]
- McCune, B.; Grace, J. Analysis of Ecological Communities; MjM software design: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- Maraun, M.; Alphei, J.; Beste, P.; Bonkowski, M.; Buryn, R.; Migge, S.; Peter, M.; Schaefer, M.; Scheu, S. Indirect effects of carbon and nutrient amendments on the soil meso-and microfauna of a beechwood. Biol. Fertil. Soils. 2001, 34, 222–229. [Google Scholar]
- Hamilton, W.E.; Sillman, D.Y. Influence of Earthworm Middens on the Distribution of Soil Microarthropods. Biol. Fertil. Soils. 1989, 8, 279–284. [Google Scholar]
- Mortensen, S.; Mortensen, C. A new angle on earthworms. Minn. Conserv. 1998, 20–29. [Google Scholar]
- Li, X.; Fisk, M.C.; Fahey, T.J.; Bohlen, P.J. Influence of earthworm invasion on soil microbial biomass and activity in a northern hardwood forest. Soil Biol. Biochem. 2002, 34, 1929–1937. [Google Scholar]
- Mitchell, M. Vertical and horizontal distributions of oribatid mites (Acari: Cryptostigmata) in an aspen woodland soil. Ecology. 1978, 59, 516–525. [Google Scholar]
- Norton, R. Evolutionary aspects of oribatid mite life histories and consequences for the origin of the Astigmata. In Mites: Ecological and Evolutionary Analyses of Life-history Patterns; Houck, M.A., Ed.; Chapman & Hall, Inc.: New York, NY, USA, 1994; pp. 99–135. [Google Scholar]
- Schneider, K.; Renker, C.; Maraun, M. Oribatid mite (Acari, Oribatida) feeding on ectomycorrhizal fungi. Mycorrhiza. 2005, 16, 67–72. [Google Scholar]
- Went, J. Influence of earthworms on the number of bacteria in the soil. In Soil Organisms; Doeksen, J., Van der Drift, J., Eds.; North-Holland: Amsterdam, Holland, 1963; pp. 260–265. [Google Scholar]
- Scheu, S.; Parkinson, D. Effects of earthworms on nutrient dynamics, carbon turnover and microorganisms in soils from cool temperate forests of the Canadian Rocky Mountains-Laboratory Studies. Appl. Soil Ecol. 1994, 1, 113–125. [Google Scholar]
- Marshall, V.; Reeves, R.; Norton, R.; Ewen, A. Catalogue of the Oribatida (Acari) of continental United States and Canada. Entomol. Soc. Can. 1987. [Google Scholar]
- Hansen, R.A. Effects of habitat complexity and composition on a diverse litter microarthropod assemblage. Ecology. 2000, 81, 1120–1132. [Google Scholar]
- Huhta, V.; Niemi, R. Communities of soil mites (Acarina) in planted birch stands compared with natural forests in central Finland. Can. J. For. Res. 2003, 33, 171–180. [Google Scholar]
- Battigelli, J.P.; Spence, J.R.; Langor, D.L.; Berch, S.M. Short-term impact of forest soil compaction and organic matter removal on soil mesofauna density and oribatid mite diversity. Can. J. For. Res. 2004, 34, 1136–1149. [Google Scholar]
- Peck, R.; Niwa, C. Longer-term effects of selective thinning on microarthropod communities in a late-successional coniferous forest. Environ. Entomol. 2005, 34, 646–655. [Google Scholar]
- Zaitsev, A.; Chauvat, M.; Pflug, A.; Wolters, V. Oribatid mite diversity and community dynamics in a spruce chronosequence. Soil Biol. Biochem. 2002, 34, 1919–1927. [Google Scholar]
- Callaham, M.; Gonzalez, G.; Hale, C.; Heneghan, L.; Lachnicht, S.; Zou, X. Policy and management responses to earthworm invasions in North America. Biol. Inv. 2006, 8, 1317–1329. [Google Scholar]
- Addison, J.A. Distribution and impacts of invasive earthworms in Canadian forest ecosystems. Biol. Inv. 2009, 11, 59–79. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the CreativeCommons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Burke, J.L.; Maerz, J.C.; Milanovich, J.R.; Fisk, M.C.; Gandhi, K.J.K. Invasion by Exotic Earthworms Alters Biodiversity and Communities of Litter- and Soil-dwelling Oribatid Mites. Diversity 2011, 3, 155-175. https://doi.org/10.3390/d3010155
Burke JL, Maerz JC, Milanovich JR, Fisk MC, Gandhi KJK. Invasion by Exotic Earthworms Alters Biodiversity and Communities of Litter- and Soil-dwelling Oribatid Mites. Diversity. 2011; 3(1):155-175. https://doi.org/10.3390/d3010155
Chicago/Turabian StyleBurke, Jordan L., John C. Maerz, Joseph R. Milanovich, Melany C. Fisk, and Kamal J.K. Gandhi. 2011. "Invasion by Exotic Earthworms Alters Biodiversity and Communities of Litter- and Soil-dwelling Oribatid Mites" Diversity 3, no. 1: 155-175. https://doi.org/10.3390/d3010155
APA StyleBurke, J. L., Maerz, J. C., Milanovich, J. R., Fisk, M. C., & Gandhi, K. J. K. (2011). Invasion by Exotic Earthworms Alters Biodiversity and Communities of Litter- and Soil-dwelling Oribatid Mites. Diversity, 3(1), 155-175. https://doi.org/10.3390/d3010155