
Isolation and Identification of Salinity-Tolerant Rhizobia and Nodulation Phenotype Analysis in Different Soybean Germplasms


 and
and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Isolation and Characterization of Rhizobia from Different Regions
2.2. Identification of Rhizobium Resistance
2.3. Identification of Rhizobium Host Compliance with Cultivars
2.4. Data Collection
2.5. Statistical Analysis
3. Results
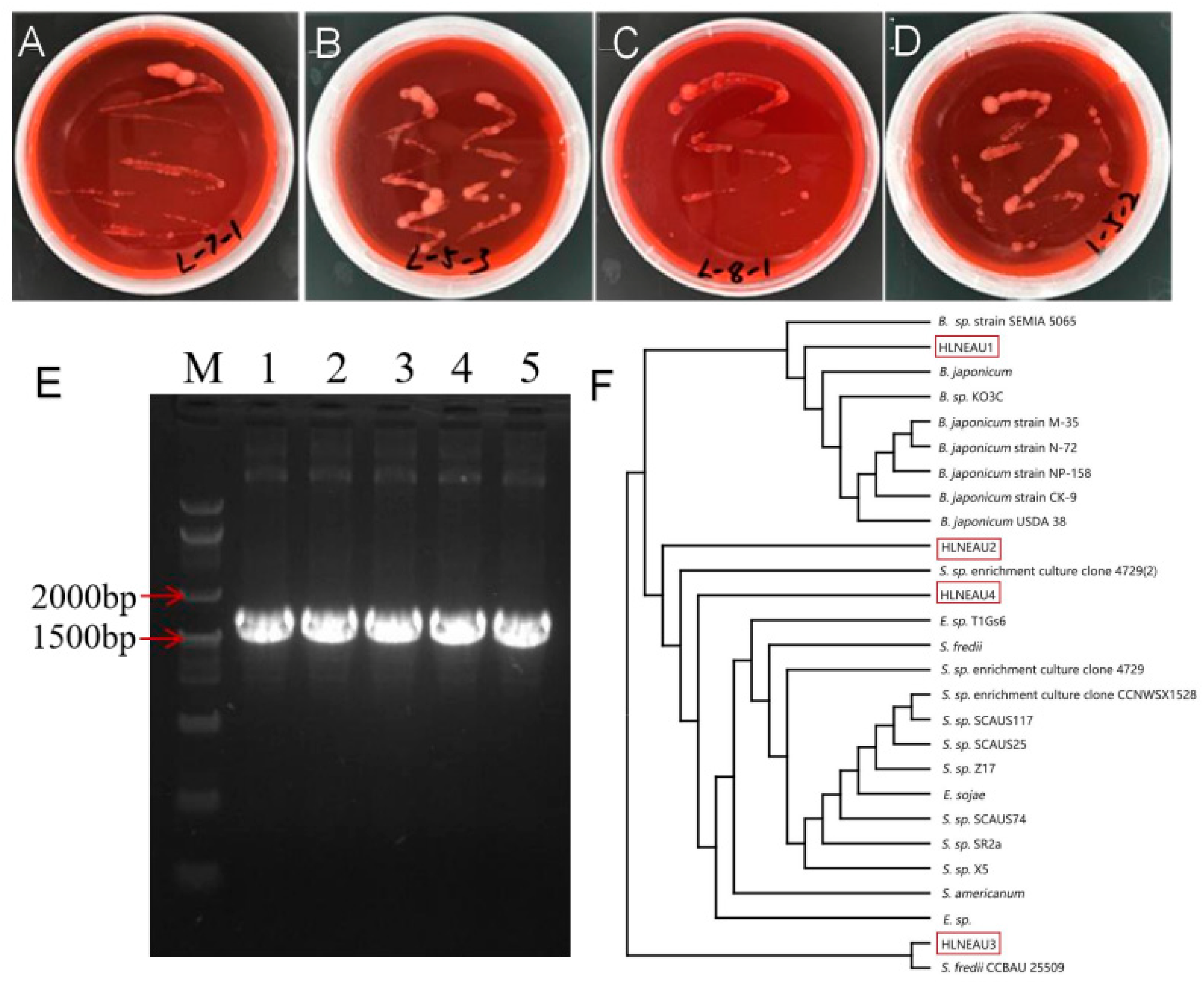
3.1. Isolation and Identification of Rhizobia from Different Regions
3.2. Identification of Rhizobium Antibiotic Resistance
3.2.1. Antibiotic Resistance Identification of Rhizobia
3.2.2. Salt Resistance, Alkali Resistance, and Acid Resistance Identification
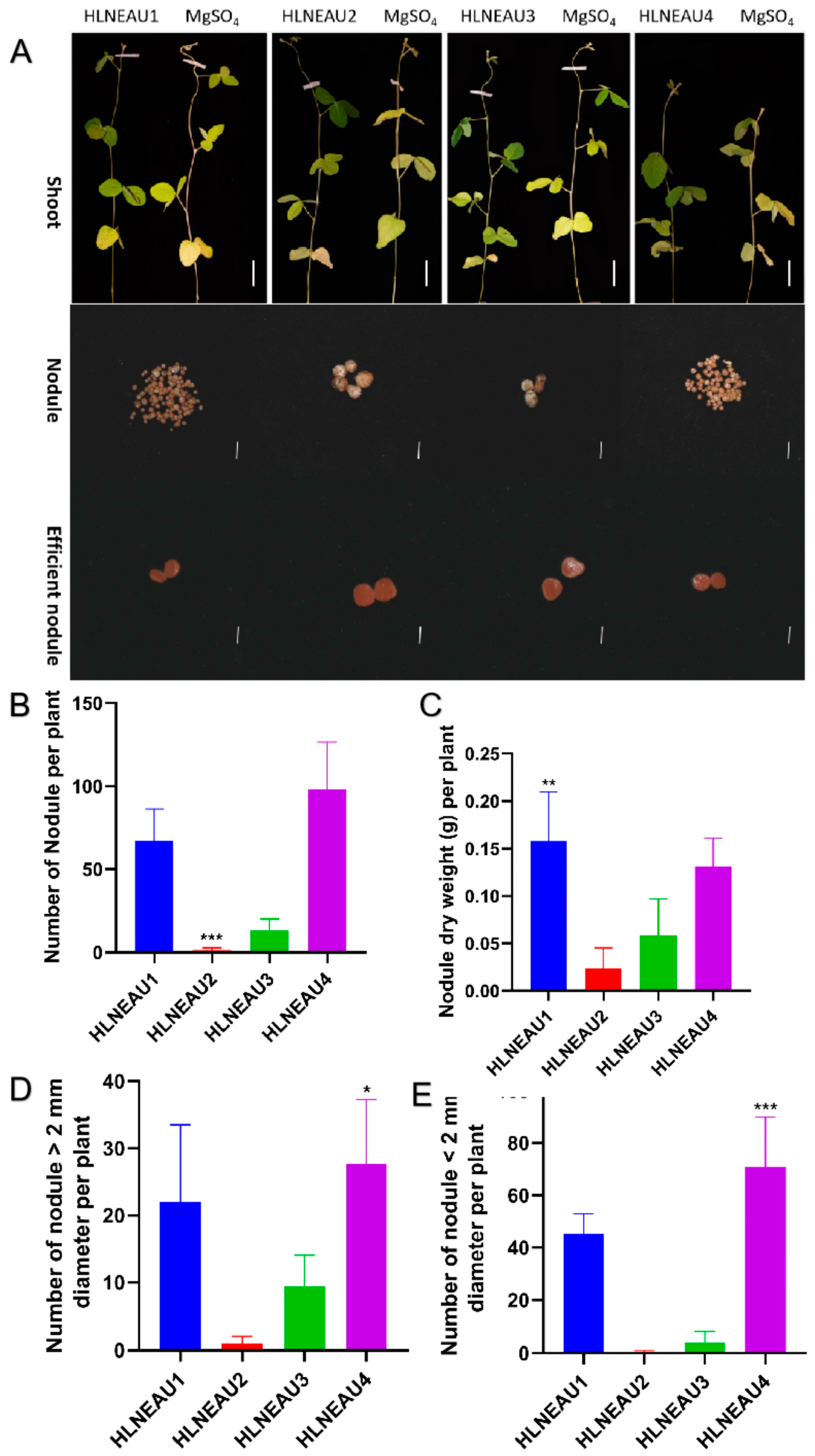
3.3. Identification of Nodulation Phenotypes of Isolated Rhizobia and Compatibilities with Cultivated Soybean Varieties
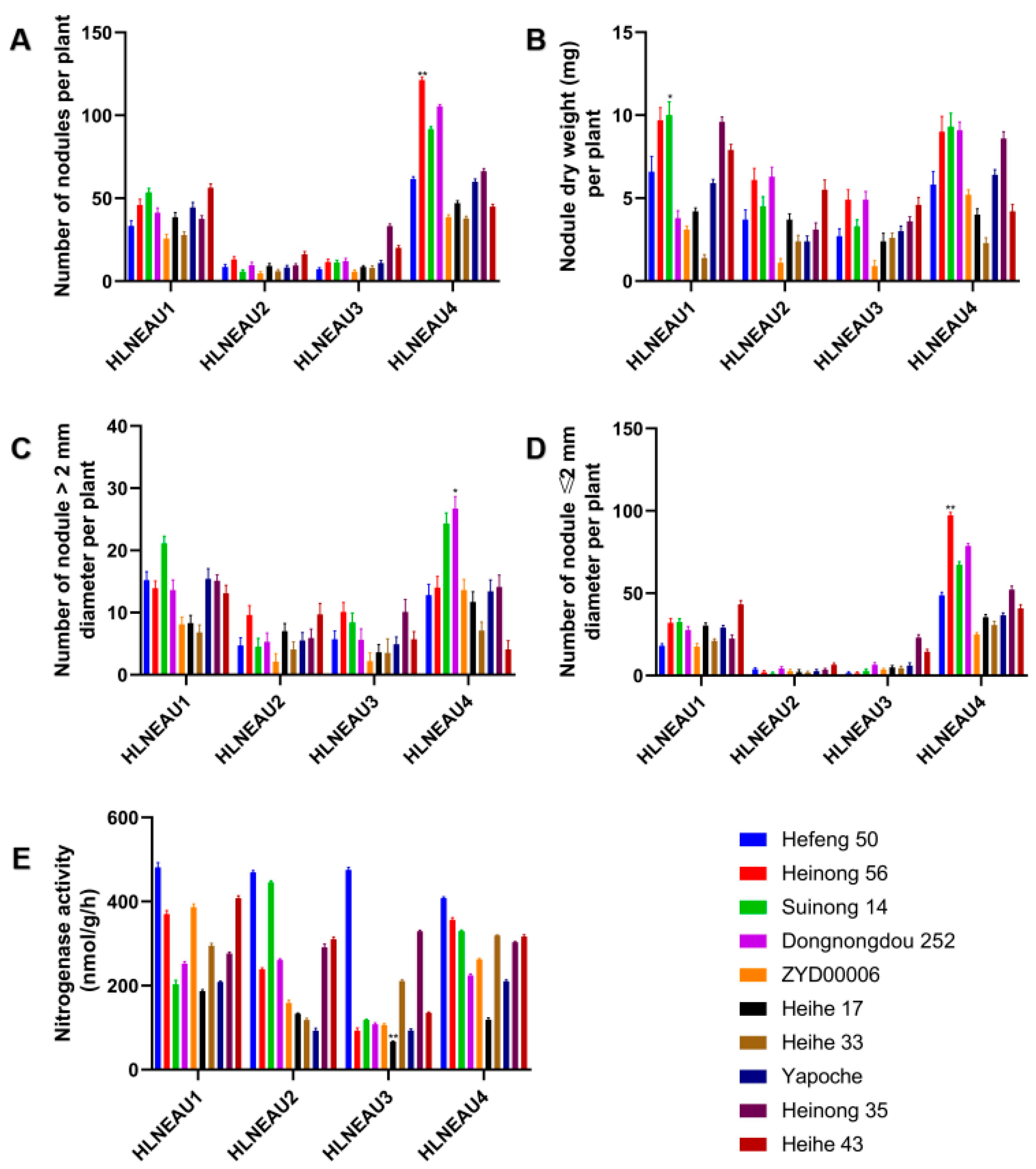
3.4. Identification of Rhizobium Host Compliances with Cultivars
4. Discussion
4.1. Application Analysis of Rhizobia from Different Areas
4.2. The Effects of Rhizobia on the Environment
4.3. Effects of Compatibility between Soybean and Rhizobacteria on Symbiotic Nitrogen Fixation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Balbinot Junior, A.A.; Debiasi, H.; Franchini, J.C.; de Oliveira, M.A.; Coelho, A.E.; de Moraes, M.T. Soybean yield, seed protein and oil concentration, and soil fertility affected by off-season crops. Eur. J. Agron. 2024, 153, 127039. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, Y.; Zhao, M.; Wang, K.; Li, S.; Gao, Z.; Shi, X.; Chu, Q. Prospects for soybean production increase by closing yield gaps in the Northeast Farming Region, China. Field Crops Res. 2023, 293, 108843. [Google Scholar] [CrossRef]
- Serafin-Andrzejewska, M.; Jama-Rodzeńska, A.; Helios, W.; Kozak, M.; Lewandowska, S.; Zalewski, D.; Kotecki, A. Influence of nitrogen fertilization, seed inoculation and the synergistic effect of these treatments on soybean yields under conditions in south-western Poland. Sci. Rep. 2024, 14, 6672. [Google Scholar] [CrossRef]
- Ye, J.Y.; Tian, W.H.; Jin, C.W. Nitrogen in plants: From nutrition to the modulation of abiotic stress adaptation. Stress Biol. 2022, 2, 4. [Google Scholar] [CrossRef]
- Zhang, Y.; Ye, C.; Su, Y.; Peng, W.; Lu, R.; Liu, Y.; Huang, H.; He, X.; Yang, M.; Zhu, S. Soil Acidification caused by excessive application of nitrogen fertilizer aggravates soil-borne diseases: Evidence from literature review and field trials. Agric. Ecosyst. Environ. 2022, 340, 108176. [Google Scholar] [CrossRef]
- Sinharoy, S.; Tian, C.-F.; Montiel, J. Editorial: Plant-rhizobia symbiosis and nitrogen fixation in legumes. Front. Plant Sci. 2024, 15, 1392006. [Google Scholar] [CrossRef]
- Poole, P. Symbiosis for rhizobia is not an easy ride. Nat. Microbiol. 2024, 9, 314–315. [Google Scholar] [CrossRef]
- Burghardt, L.T.; diCenzo, G.C. The evolutionary ecology of rhizobia: Multiple facets of competition before, during, and after symbiosis with legumes. Curr. Opin. Microbiol. 2023, 72, 102281. [Google Scholar] [CrossRef]
- Kohlen, W.; Ng, J.L.P.; Deinum, E.E.; Mathesius, U. Auxin transport, metabolism, and signalling during nodule initiation: Indeterminate and determinate nodules. J. Exp. Bot. 2017, 69, 229–244. [Google Scholar] [CrossRef]
- Mahmud, K.; Missaoui, A.; Lee, K.; Ghimire, B.; Presley, H.W.; Makaju, S. Rhizosphere microbiome manipulation for sustainable crop production. Curr. Plant Biol. 2021, 27, 100210. [Google Scholar] [CrossRef]
- Mng′ong′o, M.E.; Ojija, F.; Aloo, B.N. The role of Rhizobia toward food production, food and soil security through microbial agro-input utilization in developing countries. Case Stud. Chem. Environ. Eng. 2023, 8, 100404. [Google Scholar] [CrossRef]
- Xu, H.; Yang, Y.; Tian, Y.; Xu, R.; Zhong, Y.; Liao, H. Rhizobium Inoculation Drives the Shifting of Rhizosphere Fungal Community in a Host Genotype Dependent Manner. Front. Microbiol. 2019, 10, 3135. [Google Scholar] [CrossRef]
- Jabborova, D.; Kannepalli, A.; Davranov, K.; Narimanov, A.; Enakiev, Y.; Syed, A.; Elgorban, A.M.; Bahkali, A.H.; Wirth, S.; Sayyed, R.Z.; et al. Co-inoculation of rhizobacteria promotes growth, yield, and nutrient contents in soybean and improves soil enzymes and nutrients under drought conditions. Sci. Rep. 2021, 11, 22081. [Google Scholar] [CrossRef]
- Omari, R.A.; Yuan, K.; Anh, K.T.; Reckling, M.; Halwani, M.; Egamberdieva, D.; Ohkama-Ohtsu, N.; Bellingrath-Kimura, S.D. Enhanced Soybean Productivity by Inoculation with Indigenous Bradyrhizobium Strains in Agroecological Conditions of Northeast Germany. Front. Plant Sci. 2022, 12, 707080. [Google Scholar] [CrossRef]
- Wei, W.; Guan, D.; Ma, M.; Jiang, X.; Fan, F.; Meng, F.; Li, L.; Zhao, B.; Zhao, Y.; Cao, F.; et al. Long-term fertilization coupled with rhizobium inoculation promotes soybean yield and alters soil bacterial community composition. Front. Microbiol. 2023, 14, 1161983. [Google Scholar] [CrossRef]
- Kumar, K.S.; Pindi, P.K. Data on host specificity and symbiotic association between indigenous Rhizobium BD1 strain and Vigna radiata (green gram). Data Brief 2021, 39, 107520. [Google Scholar] [CrossRef]
- Menéndez, E.; Robledo, M.; Jiménez-Zurdo, J.I.; Velázquez, E.; Rivas, R.; Murray, J.D.; Mateos, P.F. Legumes display common and host-specific responses to the rhizobial cellulase CelC2 during primary symbiotic infection. Sci. Rep. 2019, 9, 13907. [Google Scholar] [CrossRef]
- Schoebitz, M.; López Belchí, M.D. Encapsulation Techniques for Plant Growth-Promoting Rhizobacteria. In Bioformulations: For Sustainable Agriculture; Arora, N.K., Mehnaz, S., Balestrini, R., Eds.; Springer: New Delhi, India, 2016; pp. 251–265. [Google Scholar] [CrossRef]
- Lavudi, H.N.; Jakinala, P.; Kumar, J.S.; Babu, B.N.; Srinivas, M.; Katika, M.R. Chapter 6—Plant growth promoting rhizobacteria (PGPR): An overview for sustainable agriculture and development. In Rhizobiome; Parray, J.A., Shameem, N., Egamberdieva, D., Sayyed, R.Z., Eds.; Academic Press: Waltham, MA, USA, 2023; pp. 95–125. [Google Scholar] [CrossRef]
- Damanhuri; Erdiansyah, I.; Eliyatiningsih, E.; Sari, V.; Pratama, A.; Wiharto, K. Utilization of Rhizobium spp as substitution agent of nitrogen chemical fertilizer on soybean cultivation. IOP Conf. Ser. Earth Environ. Sci. 2020, 411, 012065. [Google Scholar] [CrossRef]
- De Souza-Torres, A.; Govea-Alcaide, E.; Gómez-Padilla, E.; Masunaga, S.H.; Effenberger, F.B.; Rossi, L.M.; López-Sánchez, R.; Jardim, R.F. Fe3O4 nanoparticles and Rhizobium inoculation enhance nodulation, nitrogen fixation and growth of common bean plants grown in soil. Rhizosphere 2021, 17, 100275. [Google Scholar] [CrossRef]
- Poole, P.; Ramachandran, V.; Terpolilli, J. Rhizobia: From saprophytes to endosymbionts. Nat. Rev. Microbiol. 2018, 16, 291–303. [Google Scholar] [CrossRef]
- Tan, Q.; Liu, Y.; Dai, L.; Pan, T. Shortened key growth periods of soybean observed in China under climate change. Sci. Rep. 2021, 11, 8197. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Ma, Y.; Luo, S. Spatiotemporal Evolution and Influencing Factors of Soybean Production in Heilongjiang Province, China. Land 2023, 12, 2090. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, P.; Marley, B.; Liu, W. The Factors Affecting Farmers’ Soybean Planting Behavior in Heilongjiang Province, China. Agriculture 2019, 9, 188. [Google Scholar] [CrossRef]
- Naik, V.; Rahi, P. Methods for Isolation and Identification of RhizobiaRhizobia. In Practical Handbook on Agricultural Microbiology; Amaresan, N., Patel, P., Amin, D., Eds.; Springer: New York, NY, USA, 2022; pp. 3–14. [Google Scholar] [CrossRef]
- Naqqash, T.; Malik, K.A.; Imran, A.; Hameed, S.; Shahid, M.; Hanif, M.K.; Majeed, A.; Arshad, M.; van Elsas, J.D. Isolation and characterization of Rhizobium from non-leguminous potato plants: New frontiers in Rhizobium research. Biol. Fertil. Soils 2024, 60, 307–325. [Google Scholar] [CrossRef]
- Peng, C.; Sun, W.; Dong, X.; Zhao, L.; Hao, J. Isolation, identification and utilization of lactic acid bacteria from silage in a warm and humid climate area. Sci. Rep. 2021, 11, 12586. [Google Scholar] [CrossRef]
- Skirvin, R.M.; Chu, M.C.; Mann, M.L.; Young, H.; Sullivan, J.; Fermanian, T. Stability of tissue culture medium pH as a function of autoclaving, time, and cultured plant material. Plant Cell Rep. 1986, 5, 292–294. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhu, Z.; Deng, X.; Zou, J.; Ma, C.; Li, C.; Yin, T.; Liu, C.; Wang, J.; Chen, Q.; et al. GmPBS1, a Hub Gene Interacting with Rhizobial Type-III Effectors NopT and NopP, Regulates Soybean Nodulation. Agronomy 2023, 13, 1242. [Google Scholar] [CrossRef]
- Brensha, W.; Kantartzi, S.K.; Meksem, K.; Grier, R.L.; Bara, A.; Lightfoot, D.A. Genetic Analysis of Root and Shoot Traits in the ‘Essex’ By ‘Forrest’ Recombinant Inbred Line (RIL) Population of Soybean [Glycine max (L.) Merr.]. J. Plant Genome Sci. 2012, 1, 1–9. [Google Scholar] [CrossRef]
- Wang, J.; Huang, X.; Zhong, T.; Chen, Z. Review on sustainable utilization of salt-affected land. Acta Geogr. Sin. 2011, 66, 673–684. [Google Scholar]
- Wang, Z.; Zhao, G.; Gao, M.; Chang, C.; Jia, J.; Li, J. Characteristics and Spatial Variability of Saline-Alkaline Soil Degradation in the Typical Yellow River Delta Area of Kenli County, China. J. Environ. Prot. 2014, 05, 1053–1063. [Google Scholar] [CrossRef]
- Zongfan, B.; Ling, H.; Huiqun, L.; Xuhai, J.; Liangzhi, L. Spatiotemporal change and driving factors of ecological status in Inner Mongolia based on the modified remote sensing ecological index. Environ. Sci. Pollut. Res. 2023, 30, 52593–52608. [Google Scholar] [CrossRef]
- Lu, X.; Wu, Z.; Xue, Z.; Feng, W.; Meng, R.; Sun, X.; Li, Z.; Wang, X. Study on Salt Tolerance Identification for 144 Soybean Resources. Hans J. Agric. Sci. 2023, 13, 481–487. [Google Scholar] [CrossRef]
- Li, P.; Teng, C.; Zhang, J.; Liu, Y.; Wu, X.; He, T. Characterization of drought stress-mitigating Rhizobium from faba bean (Vicia faba L.) in the Chinese Qinghai-Tibet Plateau. Front. Microbiol. 2023, 14, 1212996. [Google Scholar] [CrossRef]
- Abd-Alla, M.H.; Morsy, F.M.; El-Enany, A.-W.E.; Ohyama, T. Isolation and characterization of a heavy-metal-resistant isolate of Rhizobium leguminosarum bv. viciae potentially applicable for biosorption of Cd2+ and Co2+. Int. Biodeterior. Biodegrad. 2012, 67, 48–55. [Google Scholar] [CrossRef]
- Sobti, S.; Belhadj, H.A.; Djaghoubi, A. Isolation and Characterization of the Native Rhizobia Under Hyper-Salt Edaphic Conditions in Ouargla (southeast Algeria). Energy Procedia 2015, 74, 1434–1439. [Google Scholar] [CrossRef]
- Saranraj, P.; Sayyed, R.Z.; Sivasakthivelan, P.; Kokila, M.; Al-Tawaha, A.R.M.; Amala, K.; Yasmin, H. Symbiotic Effectiveness of Rhizobium Strains in Agriculture. In Plant Growth Promoting Microorganisms of Arid Region; Mawar, R., Sayyed, R.Z., Sharma, S.K., Sattiraju, K.S., Eds.; Springer Nature: Singapore, 2023; pp. 389–421. [Google Scholar] [CrossRef]
- Wadhwa, Z.; Srivastava, V.; Rani, R.; Bhatia, T.; Makkar, K.; Jangra, S. Isolation and Characterization of Rhizobium from Chickpea (Cicer arietinum). Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 2880–2893. [Google Scholar] [CrossRef]
- Dong, R.; Zhang, J.; Huan, H.; Bai, C.; Chen, Z.; Liu, G. High Salt Tolerance of a Bradyrhizobium Strain and Its Promotion of the Growth of Stylosanthes guianensis. Int. J. Mol. Sci. 2017, 18, 1625. [Google Scholar] [CrossRef]
- Scholla, M.H.; Elkan, G.H. Rhizobium fredii sp. nov., a Fast-Growing Species That Effectively Nodulates Soybeans. Int. J. Syst. Evol. Microbiol. 1984, 34, 484–486. [Google Scholar] [CrossRef]
- Yang, S.S.; Bellogín, R.A.; Buendía, A.; Camacho, M.; Chen, M.; Cubo, T.; Daza, A.; Díaz, C.L.; Espuny, M.R.; Gutiérrez, R.; et al. Effect of pH and soybean cultivars on the quantitative analyses of soybean rhizobia populations. J. Biotechnol. 2001, 91, 243–255. [Google Scholar] [CrossRef]
- Tian, C.F.; Zhou, Y.J.; Zhang, Y.M.; Li, Q.Q.; Zhang, Y.Z.; Li, D.F.; Wang, S.; Wang, J.; Gilbert, L.B.; Li, Y.R.; et al. Comparative genomics of rhizobia nodulating soybean suggests extensive recruitment of lineage-specific genes in adaptations. Proc. Natl. Acad. Sci. USA 2012, 109, 8629–8634. [Google Scholar] [CrossRef]
- Sugawara, M.; Takahashi, S.; Umehara, Y.; Iwano, H.; Tsurumaru, H.; Odake, H.; Suzuki, Y.; Kondo, H.; Konno, Y.; Yamakawa, T.; et al. Variation in bradyrhizobial NopP effector determines symbiotic incompatibility with Rj2-soybeans via effector-triggered immunity. Nat. Commun. 2018, 9, 3139. [Google Scholar] [CrossRef]
- Ramongolalaina, C. Dynamics of symbiotic relationship of soybean with Bradyrhizobium diazoefficiens and involvements of root-secreted daidzein behind the continuous cropping. Eur. J. Soil Biol. 2019, 93, 103098. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, T.; Wu, X.; Song, Y.; Lv, H.; Zhang, G.; Tang, W.; Zheng, Z.; Wang, X.; Gu, Y.; Zhou, X.; et al. Isolation and Identification of Salinity-Tolerant Rhizobia and Nodulation Phenotype Analysis in Different Soybean Germplasms. Curr. Issues Mol. Biol. 2024, 46, 3342-3352. https://doi.org/10.3390/cimb46040209
Yu T, Wu X, Song Y, Lv H, Zhang G, Tang W, Zheng Z, Wang X, Gu Y, Zhou X, et al. Isolation and Identification of Salinity-Tolerant Rhizobia and Nodulation Phenotype Analysis in Different Soybean Germplasms. Current Issues in Molecular Biology. 2024; 46(4):3342-3352. https://doi.org/10.3390/cimb46040209
Chicago/Turabian StyleYu, Tong, Xiaodong Wu, Yunshan Song, Hao Lv, Guoqing Zhang, Weinan Tang, Zefeng Zheng, Xiaohan Wang, Yumeng Gu, Xin Zhou, and et al. 2024. "Isolation and Identification of Salinity-Tolerant Rhizobia and Nodulation Phenotype Analysis in Different Soybean Germplasms" Current Issues in Molecular Biology 46, no. 4: 3342-3352. https://doi.org/10.3390/cimb46040209
APA StyleYu, T., Wu, X., Song, Y., Lv, H., Zhang, G., Tang, W., Zheng, Z., Wang, X., Gu, Y., Zhou, X., Li, J., Tian, S., Hou, X., Chen, Q., Xin, D., & Ni, H. (2024). Isolation and Identification of Salinity-Tolerant Rhizobia and Nodulation Phenotype Analysis in Different Soybean Germplasms. Current Issues in Molecular Biology, 46(4), 3342-3352. https://doi.org/10.3390/cimb46040209