

Genome-Wide Identification and Expression Analysis of the Sweet Cherry Whirly Gene Family
Abstract
1. Introduction
2. Materials and Methods
2.1. Genome-Wide Whirly Gene Members Identification
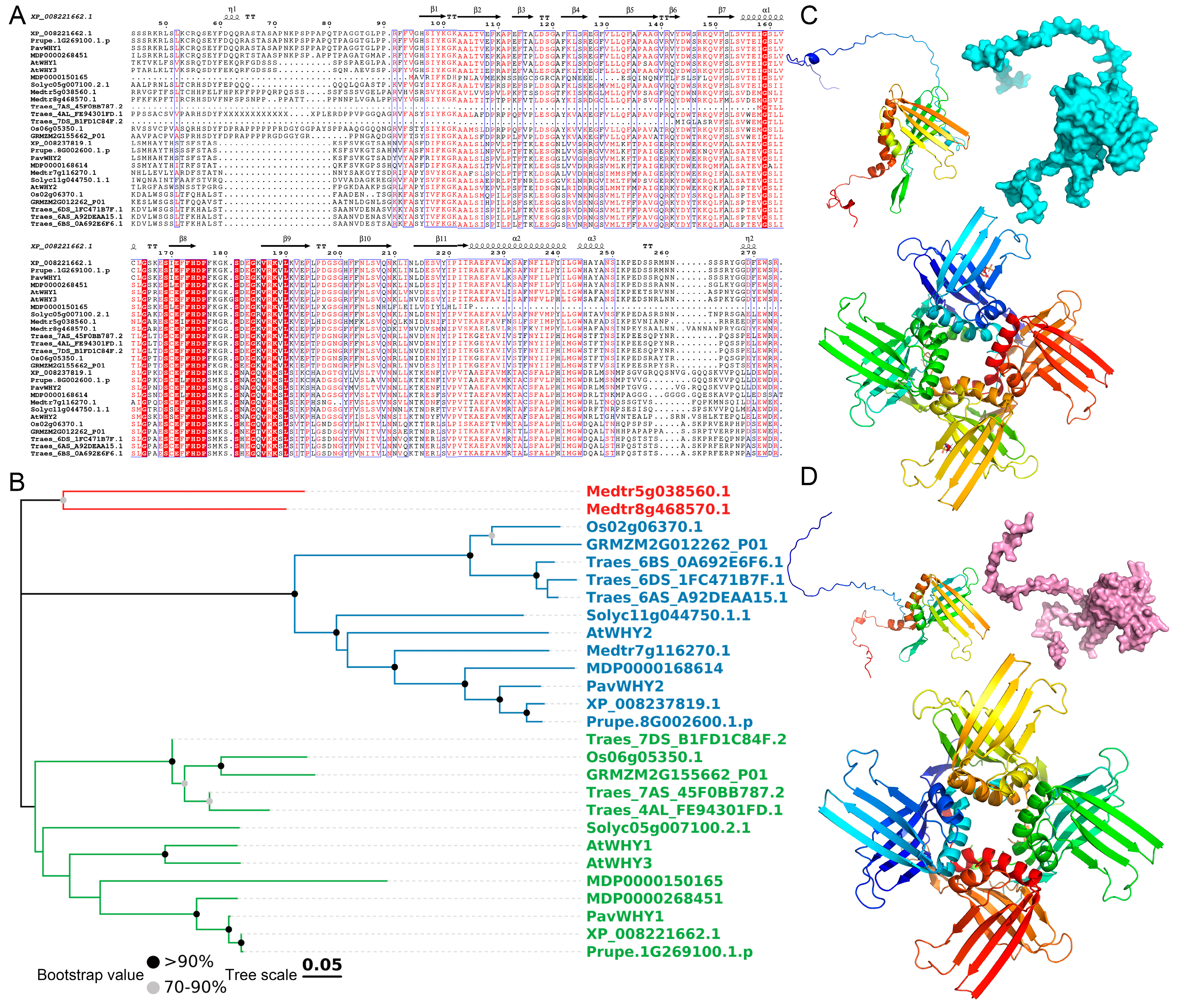
2.2. Phylogenetic Tree, Gene Structure and Conserved Motifs
2.3. Gene Localization and Collinearity Analysis
2.4. Promoter Analysis and Expression in RNA-Seq
2.5. Gene Expression under Different Treatments
2.6. Promoter Cloning and Activity Analysis
2.7. Protein Interaction Prediction
3. Results
3.1. Two WHY Members Exist in the Sweet Cherry Genome
3.2. Gene Structure of PavWHYs
3.3. Phylogenetic and Sequence Characteristics of Whirly
3.4. Chromosomal Location and Collinearity of PavWHYs
3.5. Promoter Cis-Acting Elements and Expression Profiles of PavWHYs
3.6. Tissue-Specific Expression of PavWHYs
3.7. Expression of PavWHYs during Cold Stress Treatment
3.8. Expression of PavWHYs during NaCl Treatment
3.9. Expression of PavWHYs during ABA Treatment
3.10. The Promoters of PavWHY1 and PavWHY2 Were Active
3.11. Interaction of PavWHY1 and PavWHY2 with Other Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, J.; Liu, W.; Zhu, D.; Hong, P.; Zhang, S.; Xiao, S.; Tan, Y.; Chen, X.; Xu, L.; Zong, X.; et al. Chromosome-scale genome assembly of sweet cherry (Prunus avium L.) cv. Tieton obtained using long-read and Hi-C sequencing. Hortic. Res. 2020, 7, 122. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Ediger, D.; Sharifi, M. Horticultural Practices in Early Spring to Mitigate the Adverse Effect of Low Temperature on Fruit Set in ‘Lapins’ Sweet Cherry. Plants 2023, 12, 468. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Yang, S. Surviving and thriving: How plants perceive and respond to temperature stress. Dev. Cell 2022, 57, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhao, X.; Bürger, M.; Chory, J.; Wang, X. The role of ethylene in plant temperature stress response. Trends Plant Sci. 2023, 28, 808–824. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Luan, Y.; Meng, J.; Sun, J.; Tao, J.; Zhao, D. WRKY transcription factor response to high-temperature stress. Plants 2021, 10, 2211. [Google Scholar] [CrossRef] [PubMed]
- Hou, Q.; Qiu, Z.; Wen, Z.; Zhang, H.; Li, Z.; Hong, Y.; Qiao, G. Genome-Wide Identification of ARF Gene Family Suggests a Functional Expression Pattern during Fruitlet Abscission in Prunus avium L. Int. J. Mol. Sci. 2021, 22, 11968. [Google Scholar] [CrossRef] [PubMed]
- Hou, Q.; Yu, R.; Shang, C.; Deng, H.; Wen, Z.; Qiu, Z.; Qiao, G. Molecular characterization and evolutionary relationships of DOFs in four cherry species and functional analysis in sweet cherry. Int. J. Biol. Macromol. 2024, 263, 130346. [Google Scholar] [CrossRef] [PubMed]
- Bonthala, V.S.; Mayes, K.; Moreton, J.; Blythe, M.; Wright, V.; May, S.T.; Massawe, F.; Mayes, S.; Twycross, J. Identification of gene modules associated with low temperatures response in bambara groundnut by network-based analysis. PLoS ONE 2016, 11, 148771. [Google Scholar] [CrossRef] [PubMed]
- Maréchal, A.; Parent, J.S.; Véronneau-Lafortune, F.; Joyeux, A.; Lang, B.F.; Brisson, N. Whirly proteins maintain plastid genome stability in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 14693–14698. [Google Scholar] [CrossRef]
- Taylor, R.E.; West, C.E.; Foyer, C.H. WHIRLY protein functions in plants. Food Energy Secur. 2023, 12, e379. [Google Scholar] [CrossRef]
- Lee, J.M.; Cho, C.H.; Park, S.I.; Choi, J.W.; Song, H.S.; West, J.A.; Bhattacharya, D.; Yoon, H.S. Parallel evolution of highly conserved plastid genome architecture in red seaweeds and seed plants. BMC Biol. 2016, 14, 75. [Google Scholar] [CrossRef] [PubMed]
- Desveaux, D.; Maréchal, A.; Brisson, N. Whirly transcription factors: Defense gene regulation and beyond. Trends Plant Sci. 2005, 10, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Desveaux, D.; Despres, C.; Joyeux, A.; Subramaniam, R.; Brisson, N. PBF-2 is a novel single-stranded DNA binding factor implicated in PR-10a gene activation in potato. Plant Cell 2000, 12, 1477–1489. [Google Scholar] [CrossRef]
- Isemer, R.; Mulisch, M.; Schäfer, A.; Kirchner, S.; Koop, H.U.; Krupinska, K. Recombinant Whirly1 translocates from transplastomic chloroplasts to the nucleus. FEBS Lett. 2012, 586, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Q.; Wang, Y.; Xu, H.; Wang, B.; Zhu, X.; Wei, B.; Wei, X. Genome-wide identification, phylogenetic, and expression analysis under abiotic stress conditions of Whirly (WHY) gene family in Medicago sativa L. Sci. Rep. 2022, 12, 18676. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Karpinska, B.; Krupinska, K. The functions of WHIRLY1 and Redoxresponsive Transcription Factor 1 in cross tolerance responses in plants: A hypothesis. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 15–17. [Google Scholar] [CrossRef] [PubMed]
- Maréchal, A.; Parent, J.S.; Sabar, M.; Véronneau-Lafortune, F.; Abou-Rached, C.; Brisson, N. Overexpression of mtDNA-associated AtWhy2 compromises mitochondrial function. BMC Plant Biol. 2008, 8, 42. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.E.; Waterworth, W.; West, C.E.; Foyer, C.H. WHIRLY proteins maintain seed longevity by effects on seed oxygen signalling during imbibition. Biochem. J. 2023, 480, 941–956. [Google Scholar] [CrossRef] [PubMed]
- Desveaux, D.; Subramaniam, R.; Després, C.; Mess, J.N.; Lévesque, C.; Fobert, P.R.; Dangl, J.L.; Brisson, N. A “Whirly” transcription factor is required for salicylic acid-dependent disease resistance in Arabidopsis. Dev. Cell 2004, 6, 229–240. [Google Scholar] [CrossRef]
- Hou, Q.D.; Hong, Y.; Wen, Z.; Shang, C.Q.; Li, Z.C.; Cai, X.W.; Qiao, G.; Wen, X.P. Molecular characterization of the SAUR gene family in sweet cherry and functional analysis of PavSAUR55 in the process of abscission. J. Integr. Agric. 2023, 22, 1720–1739. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Xie, J.; Chen, Y.; Cai, G.; Cai, R.; Hu, Z.; Wang, H. Tree Visualization by One Table (tvBOT): A web application for visualizing, modifying and annotating phylogenetic trees. Nucleic Acids Res. 2023, 51, W587–W592. [Google Scholar] [CrossRef]
- Wei, H.; Chen, X.; Zong, X.; Shu, H.; Gao, D.; Liu, Q. Comparative transcriptome analysis of genes involved in anthocyanin biosynthesis in the red and yellow fruits of sweet cherry (Prunus avium L.). PLoS ONE 2015, 10, e0121164. [Google Scholar] [CrossRef] [PubMed]
- Bing, L.; Rojas, P.; Perez, J.; Villar, L.; Lienqueo, I.; Correa, F.; Sagredo, B.; Masciarelli, O.; Luna, V. Comparative transcriptomic analysis reveals novel roles of transcription factors and hormones during the flowering induction and floral bud differentiation in sweet cherry trees. PLoS ONE 2020, 15, e0230110. [Google Scholar] [CrossRef]
- Ionescu, I.A.; López-Ortega, G.; Burow, M.; Bayo-Canha, A.; Junge, A.; Gericke, O.; Møller, B.L.; Sánchez-Pérez, R. Transcriptome and metabolite changes during hydrogen cyanamide-induced floral bud break in sweet cherry. Front. Plant Sci. 2017, 8, 1233. [Google Scholar] [CrossRef]
- Rothkegel, K.; Sandoval, P.; Soto, E.; Ulloa, L.; Riveros, A.; Lillo-Carmona, V.; Cáceres-Molina, J.; Almeida, A.M.; Meneses, C. Dormant but Active: Chilling Accumulation Modulates the Epigenome and Transcriptome of Prunus avium during Bud Dormancy. Front Plant Sci. 2020, 17, 1115. [Google Scholar] [CrossRef] [PubMed]
- Jacob, A.G.; Smith, C.W.J. Intron retention as a component of regulated gene expression programs. Hum. Genet. 2017, 9, 1043–1057. [Google Scholar] [CrossRef]
- Liu, H.; Wang, X.; Yang, W.; Liu, W.; Wang, Y.; Wang, Q.; Zhao, Y. Identification of Whirly transcription factors in Triticeae species and functional analysis of TaWHY1-7D in response to osmotic stress. Front. Plant Sci. 2023, 14, 1297228. [Google Scholar] [CrossRef]
- Akbudak, M.A.; Filiz, E. Whirly (Why) transcription factors in tomato (Solanum lycopersicum L.): Genome-wide identification and transcriptional profiling under drought and salt stresses. Mol. Biol. Rep. 2019, 46, 4139–4150. [Google Scholar] [CrossRef]
- Lai, C.; Que, Q.; Pan, R.; Wang, Q.; Gao, H.; Guan, X.; Che, J.; Lai, G. The Single-Stranded DNA-Binding Gene Whirly (Why1) with a Strong Pathogen-Induced Promoter from Vitis pseudoreticulata Enhances Resistance to Phytophthora capsici. Int. J. Mol. Sci. 2022, 23, 8052. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Takusagawa, M.; Harada, N.; Fukao, Y.; Yamaoka, S.; Kohchi, T.; Hori, K.; Ohta, H.; Shikanai, T.; Nishimura, Y. Eukaryotic Components Remodeled Chloroplast Nucleoid Organization during the Green Plant Evolution. Genome Biol. Evol. 2016, 8, 1–16. [Google Scholar] [CrossRef]
- Grabowski, E.; Miao, Y.; Mulisch, M.; Krupinska, K. Single-stranded DNA-binding protein whirly1 in barley leaves is located in plastids and the nucleus of the same cell. Plant Physiol. 2008, 147, 1800–1804. [Google Scholar] [CrossRef]
- Huang, C.; Yu, J.; Cai, Q.; Chen, Y.; Li, Y.; Ren, Y.; Miao, Y. Triple-localized WHIRLY2 influences leaf senescence and silique development via carbon allocation. Plant Physiol. 2020, 184, 1348–1362. [Google Scholar] [CrossRef]
- Cappadocia, L.; Parent, J.S.; Zampini, É.; Lepage, É.; Sygusch, J.; Brisson, N. A conserved lysine residue of plant Whirly proteins is necessary for higher order protein assembly and protection against DNA damage. Nucleic Acids Res. 2012, 40, 258–269. [Google Scholar] [CrossRef]
- Chitnis, V.R.; Gao, F.; Yao, Z.; Jordan, M.C.; Park, S.; Ayele, B.T. After-ripening induced transcriptional changes of hormonal genes in wheat seeds: The cases of brassinosteroids, ethylene, cytokinin and salicylic acid. PLoS ONE 2014, 9, e0087543. [Google Scholar] [CrossRef] [PubMed]
- Meng, C.; Yang, M.; Wang, Y.; Chen, C.; Sui, N.; Meng, Q.; Zhuang, K.; Lv, W. SlWHY2 interacts with SlRECA2 to maintain mitochondrial function under drought stress in tomato. Plant Sci. 2020, 301, 110674. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Liu, W.; Wei, Y.; Shi, H. MeCIPK23 interacts with Whirly transcription factors to activate abscisic acid biosynthesis and regulate drought resistance in cassava. Plant Biotechnol. J. 2020, 18, 1504–1506. [Google Scholar] [CrossRef] [PubMed]
- Karpinska, B.; Razak, N.; James, E.K.; Morris, J.A.; Verrall, S.R.; Hedley, P.E.; Hancock, R.D.; Foyer, C.H. WHIRLY1 functions in the nucleus to regulate barley leaf development and associated metabolite profiles. Biochem. J. 2022, 479, 641–659. [Google Scholar] [CrossRef]
- Tian, T.; Qiao, G.; Wen, Z.; Deng, B.; Qiu, Z.; Hong, Y.; Wen, X. Comparative transcriptome analysis reveals the molecular regulation underlying the adaptive mechanism of cherry (Cerasus pseudocerasus Lindl.) to shelter covering. BMC Plant Biol. 2020, 1, 27. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | ID in Genome | aa | MW | pI | II | AI | GAH | Location |
|---|---|---|---|---|---|---|---|---|
| PavWHY1 | FUN_039943-T1 | 272 | 29.91092 | 9.68 | 51.12 | 71.03 | −0.371 | Plastid |
| PavWHY2 | FUN_027824-T1 | 231 | 25.21904 | 9.79 | 42.01 | 79.31 | −0.07 | Plastid, Mitochondrion |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Hou, Q.; Qiao, G. Genome-Wide Identification and Expression Analysis of the Sweet Cherry Whirly Gene Family. Curr. Issues Mol. Biol. 2024, 46, 8015-8030. https://doi.org/10.3390/cimb46080474
Wang L, Hou Q, Qiao G. Genome-Wide Identification and Expression Analysis of the Sweet Cherry Whirly Gene Family. Current Issues in Molecular Biology. 2024; 46(8):8015-8030. https://doi.org/10.3390/cimb46080474
Chicago/Turabian StyleWang, Lili, Qiandong Hou, and Guang Qiao. 2024. "Genome-Wide Identification and Expression Analysis of the Sweet Cherry Whirly Gene Family" Current Issues in Molecular Biology 46, no. 8: 8015-8030. https://doi.org/10.3390/cimb46080474
APA StyleWang, L., Hou, Q., & Qiao, G. (2024). Genome-Wide Identification and Expression Analysis of the Sweet Cherry Whirly Gene Family. Current Issues in Molecular Biology, 46(8), 8015-8030. https://doi.org/10.3390/cimb46080474