Assessing the Sensitizing and Allergenic Potential of the Albumin and Globulin Fractions from Amaranth (Amaranthus hypochondriacus) Grains before and after an Extrusion Process

 , , ,
, , ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Amaranth Flour and Extrusion Process
2.2. Protein Extraction
2.3. Ovalbumin and Potato Acid Phosphatase Preparations
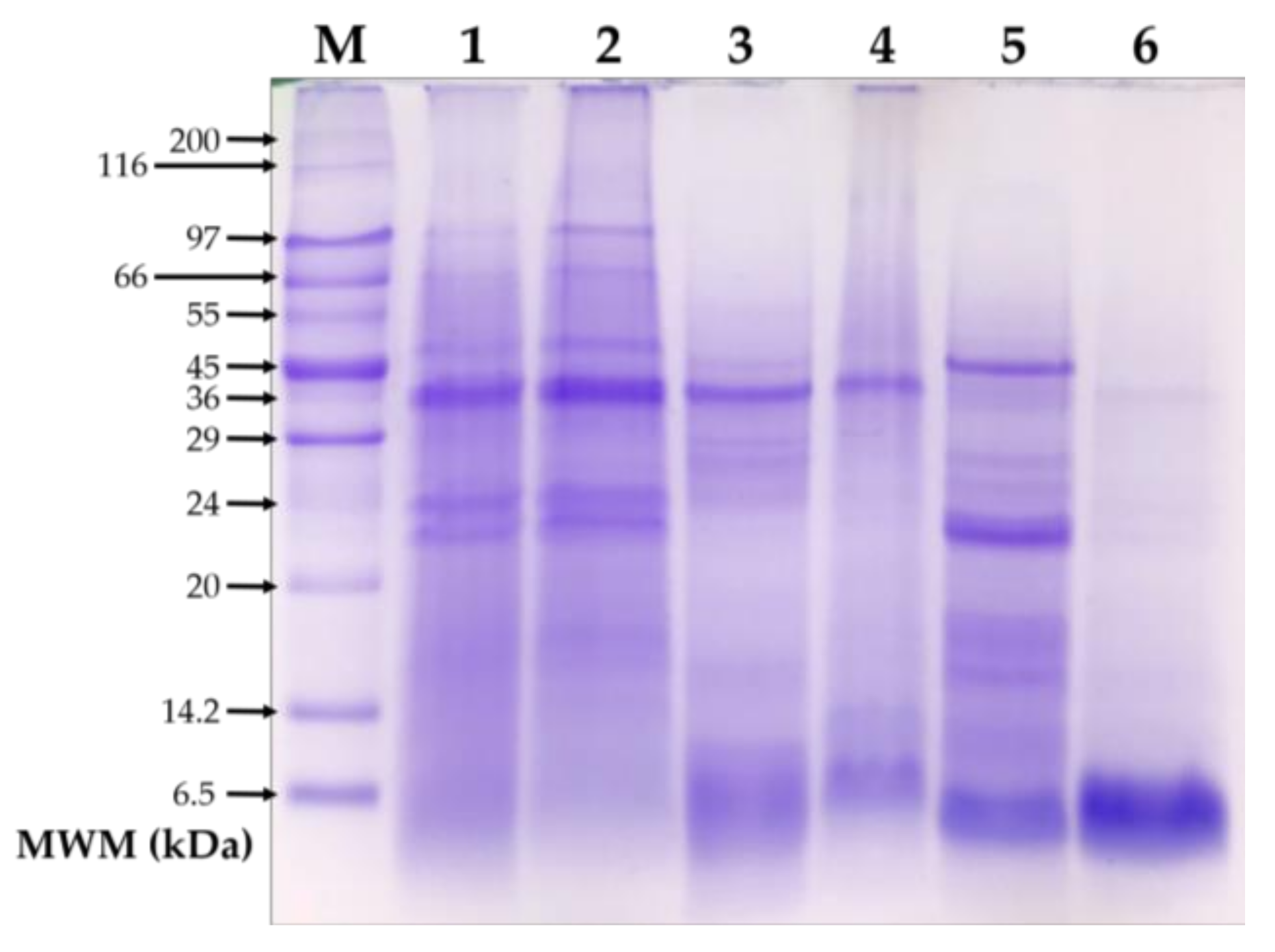
2.4. SDS-PAGE Analysis
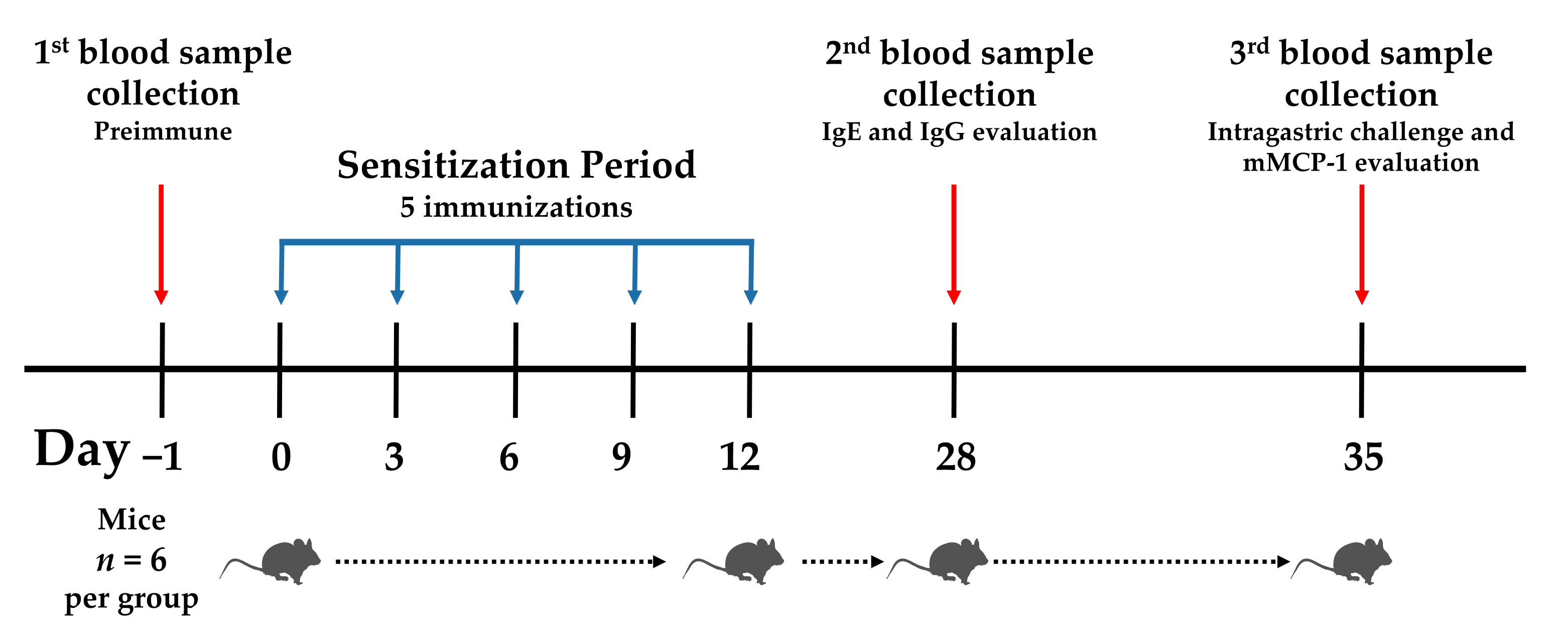
2.5. Sensitization Protocol and Ethical Aspects
2.6. Enzyme-Linked Immunosorbent Assay
2.7. mMCP-1 Serum Evaluation
2.8. Statistical Analysis
3. Results
3.1. SDS-PAGE
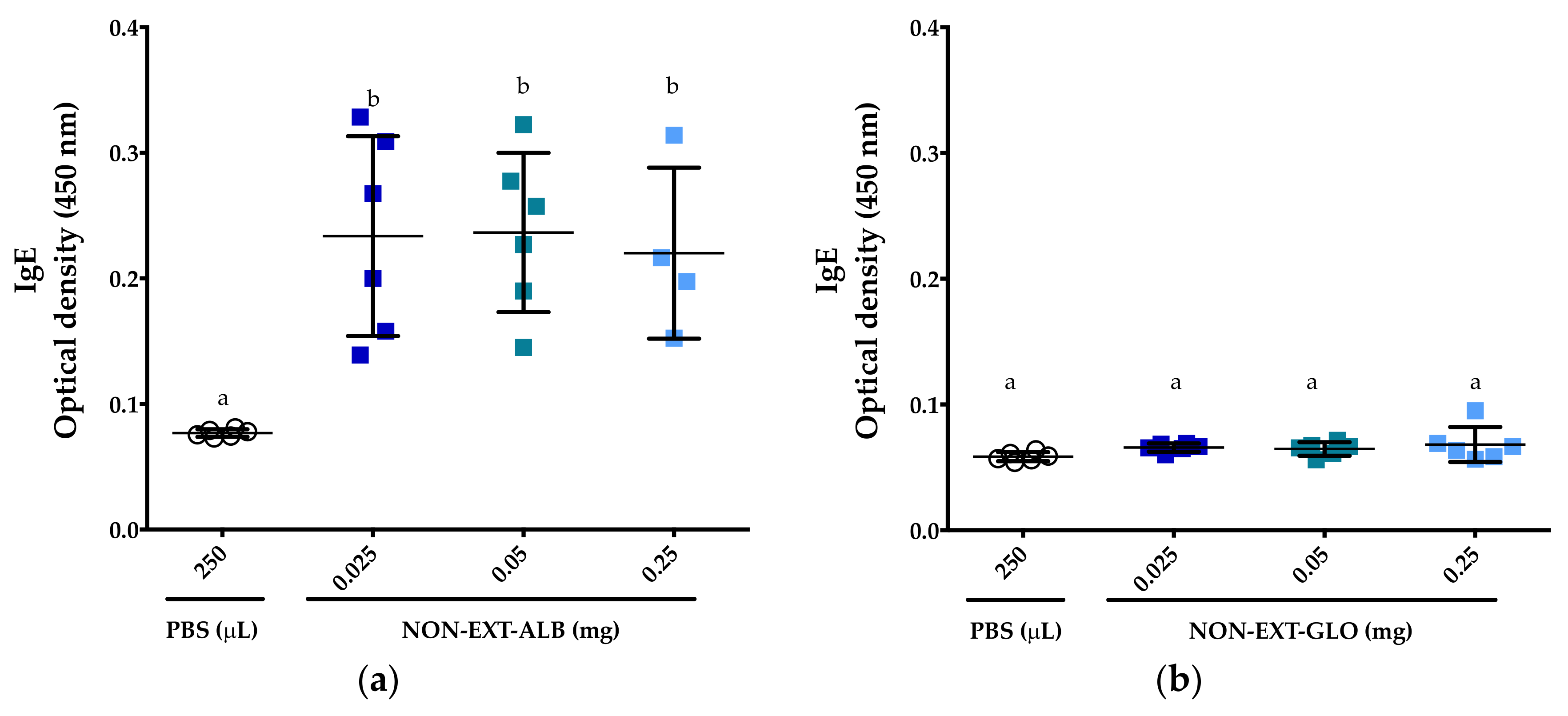
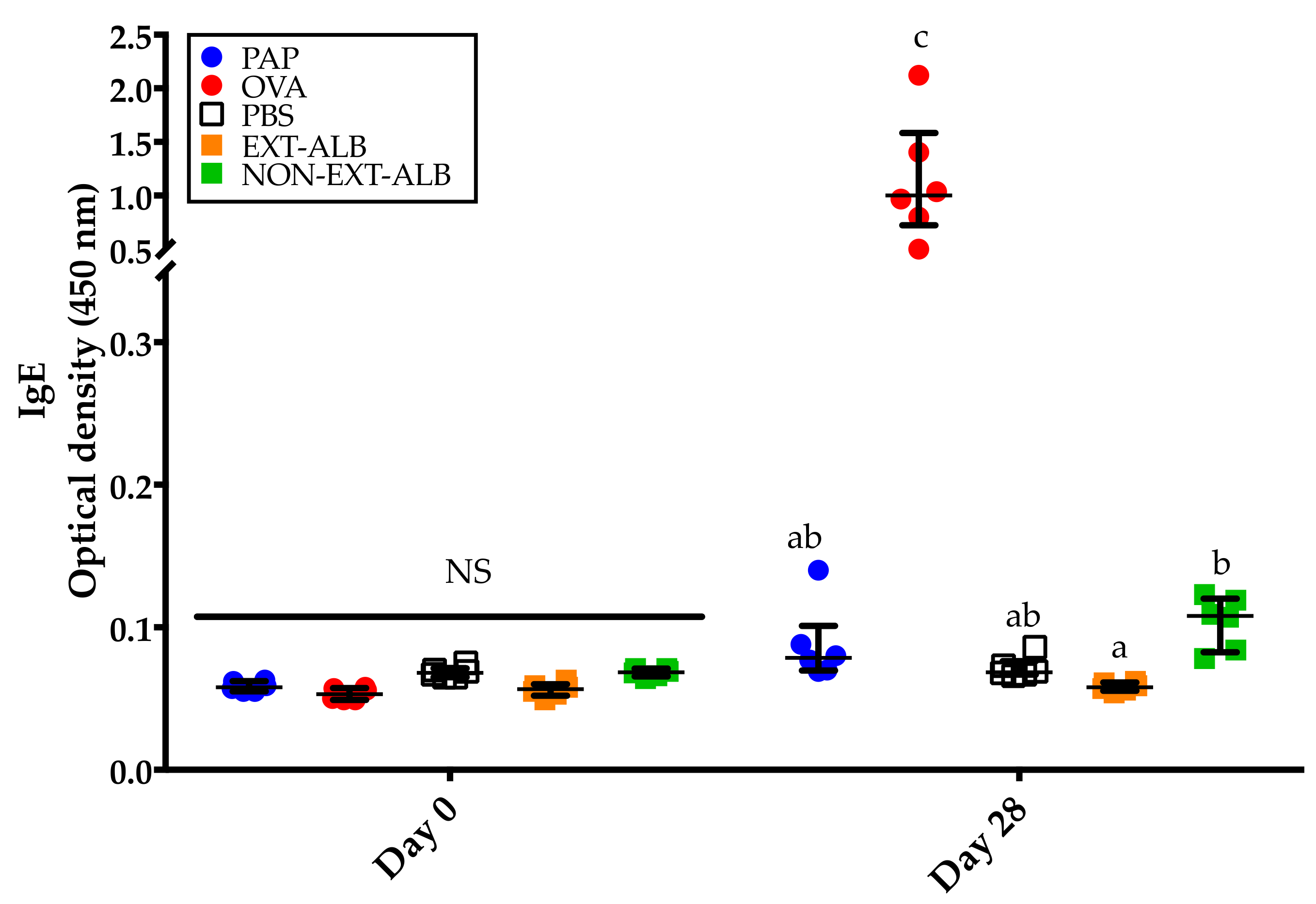
3.2. Albumins, But Not Globulins, Trigger Consistent IgE Responses in BALB/c Mice
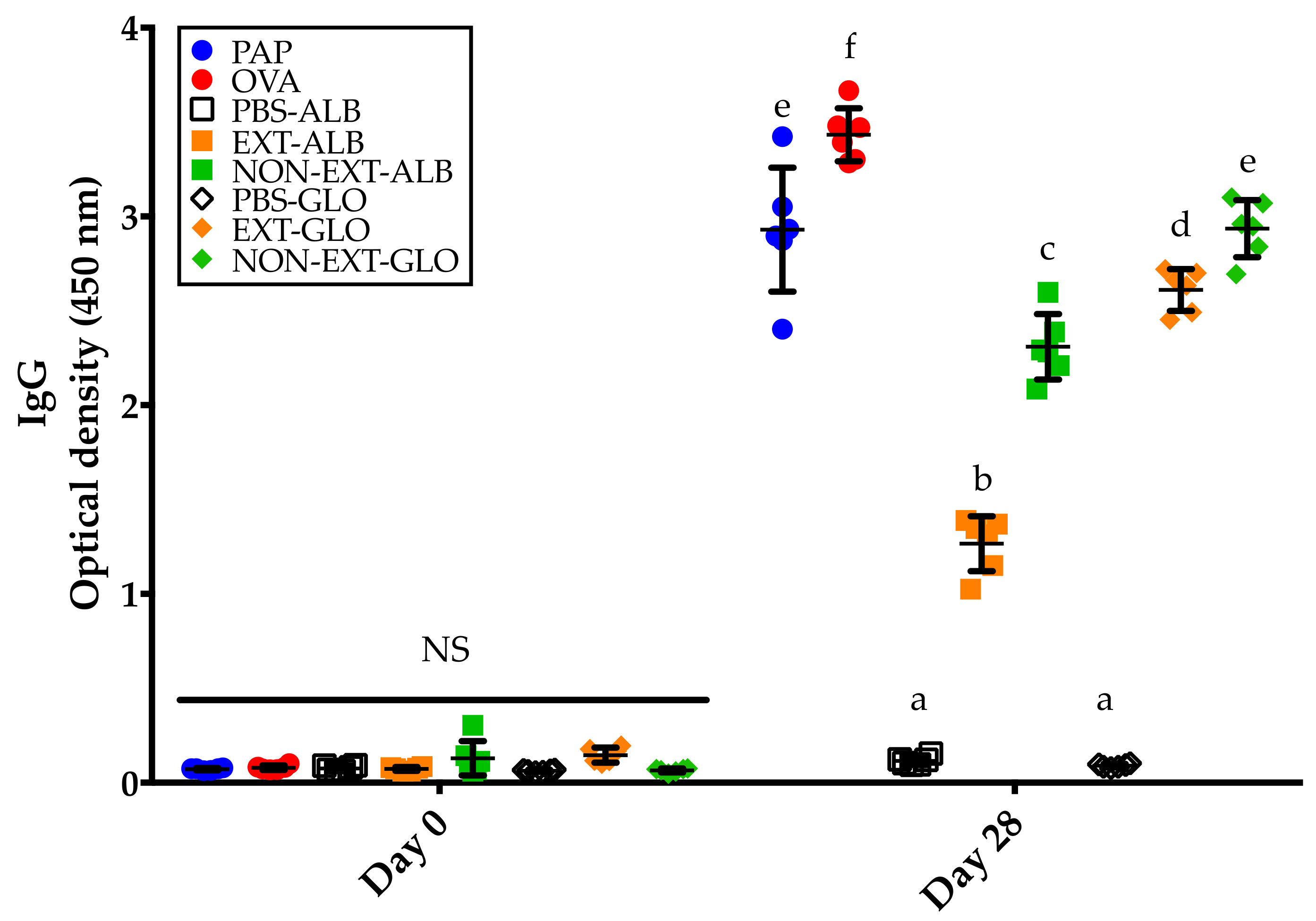
3.3. Amaranth Protein Fractions, Ovalbumin, and Potato Acid Phosphatase Trigger IgG Responses
3.4. The Extrusion Process Abolishes the IgE Immune Response Triggered by Amaranth Albumins
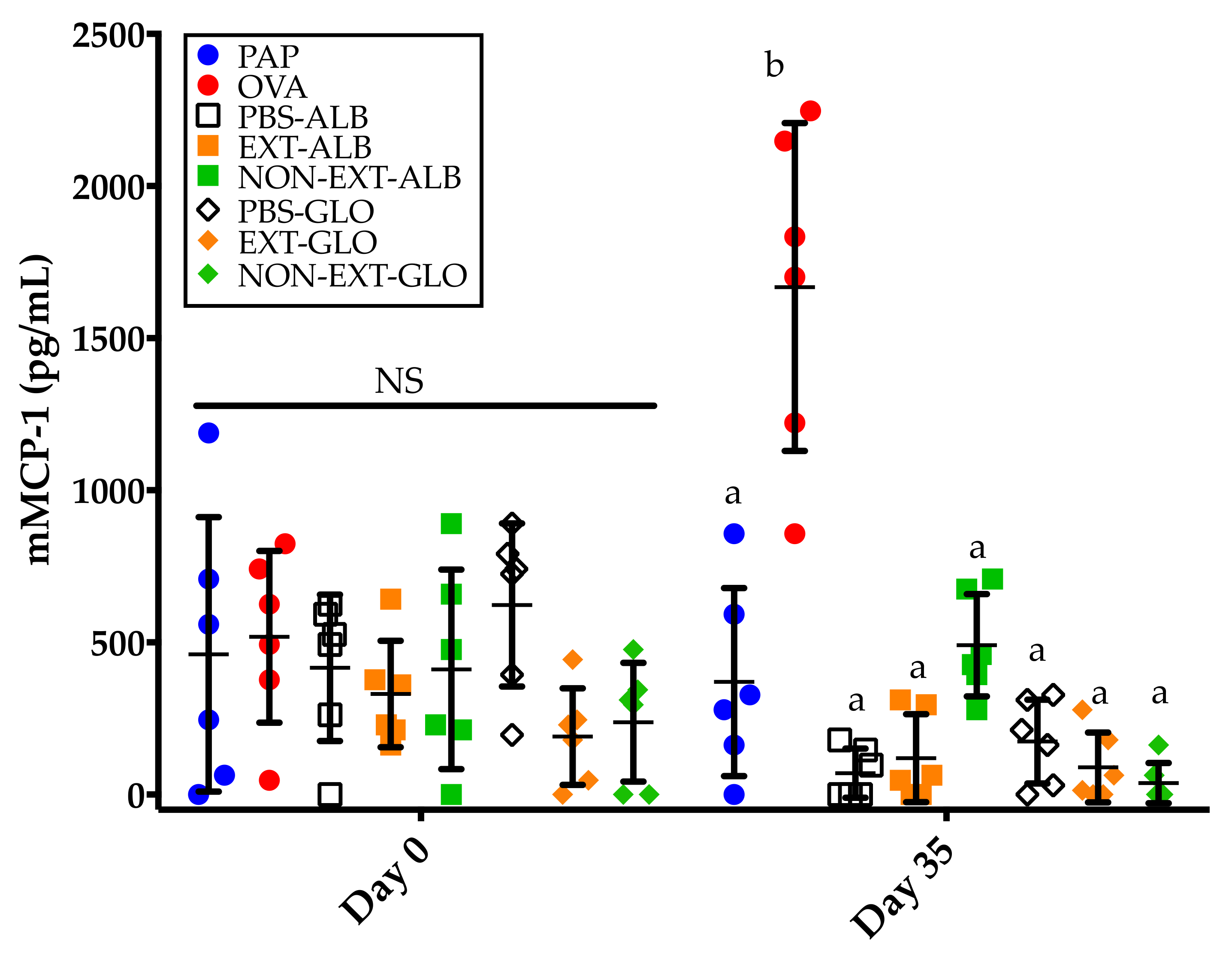
3.5. Mice Sensitized to Amaranth Albumins Fail to Trigger mMCP-1 Responses
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Casañal, A.; Zander, U.; Dupeux, F.; Valpuesta, V.; Marquez, J.A. Purification, crystallization and preliminary X-ray analysis of the strawberry allergens Fra a 1E and Fra a 3 in the presence of catechin. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2013, 69, 510–514. [Google Scholar] [CrossRef]
- Waserman, S.; Watson, W. Food allergy. Allergy Asthma Clin. Immunol. 2011, 7, S7. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Zhang, H.; Zeng, X.; Yang, P. Self-Amplification Mechanisms of Mast Cell Activation: A New Look in Allergy. Curr. Mol. Med. 2012, 12, 1329–1339. [Google Scholar] [CrossRef] [PubMed]
- Wastling, J.M.; Knight, P.; Ure, J.; Wright, S.; Thornton, E.M.; Scudamore, C.L.; Mason, J.; Smith, A.; Miller, H.R.P. Histochemical and ultrastructural modification of mucosal mast cell granules in parasitized mice lacking the β-chymase, mouse mast cell protease-1. Am. J. Pathol. 1998, 153, 491–504. [Google Scholar] [CrossRef]
- Sicherer, S.H.; Muñoz-Furlong, A.; Burks, A.W.; Sampson, H.A. Prevalence of peanut and tree nut allergy in the US determined by a random digit dial telephone survey. J. Allergy Clin. Immunol. 1999, 103, 559–562. [Google Scholar] [CrossRef]
- Ontiveros, N.; Flores-Mendoza, L.; Canizalez-Roman, V.; Cabrera-Chavez, F. Food Allergy: Prevalence and Food Technology Approaches for the Control of IgE-mediated Food Allergy. Austin J. Nutr. Food Sci. 2014, 2, 1029. [Google Scholar]
- Food and Agriculture Organization (FAO). Foods Derived from Modern Biotechnology; Codex Alimentarius; Food and Agriculture Organization of the United Nations: Rome, Italy, 2009. [Google Scholar]
- Matías-Luis, G.; Hernández-Hernández, B.R.; Peña-Caballero, V.; Torres-López, N.G.; Espinoza-Martínez, V.A.; Ramírez-Pacheco, L. Current and potential uses of Amaranth (Amaranthus spp.). J. Negat. No Posit. Results 2018, 3, 423–436. [Google Scholar] [CrossRef]
- Kasera, R.; Niphadkar, P.V.; Saran, A.; Mathur, C.; Singh, A.B. First case report of anaphylaxis caused by Rajgira seed flour (Amaranthus paniculatus) from India: A clinico-immunologic evaluation. Asian Pac. J. Allergy Inmmonol. 2013, 31, 79–83. [Google Scholar]
- Pföhler, C.; Merkoureas, A.; Müller, C.S.; Vogt, T. Fading Due the Unfading: Repeated Anaphylaxis Caused by Amaranth Grains. J. Allergy Ther. 2015, 6, 5–6. [Google Scholar] [CrossRef]
- Deluze, C.; Beaumont, P. Allergie alimentaire à la graine d’amarante. Rev. Fr. Allergol. 2017, 57, 337–340. [Google Scholar] [CrossRef]
- Tehrani, M.; Sankian, M.; Assarehzadegan, M.A.; Falak, R.; Noorbakhsh, R.; Moghadam, M.; Jabbari, F.; Varasteh, A. Identification of a new allergen from Amaranthus retroflexus pollen, Ama r 2. Allergol. Int. 2011, 60, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Villalba, M.; Barderas, R.; Mas, S.; Colás, C.; Batanero, E.; Rodríguez, R. Amaranthaceae pollens: Review of an emerging allergy in the mediterranean area. J. Investig. Allergol. Clin. Immunol. 2014, 24, 371–381. [Google Scholar]
- Arámburo-Galvez, J.G.; Sotelo-Cruz, N.; Flores-Mendoza, L.K.; Gracia-Valenzuela, M.H.; Chiquete-Elizalde, F.I.R.; Espinoza-Alderete, J.G.; Trejo-Martínez, H.; Canizalez-Román, V.A.; Ontiveros, N.; Cabrera-Chávez, F. Assessment of the Sensitizing Potential of Proteins in BALB/c Mice: Comparison of Three Protocols of Intraperitoneal Sensitization. Nutrients 2018, 10, 903. [Google Scholar] [CrossRef] [PubMed]
- Besler, M.; Steinhart, H.; Paschke, A. A Stability of food allergens and allergenicity of processed foods. J. Chromatogr. B. Biomed. Sci. Appl. 2001, 756, 207–228. [Google Scholar] [CrossRef]
- Chen, L.; Phillips, R.D. Effects of twin-screw extrusion of peanut flour on in vitro digestion of potentially allergenic peanut proteins. J. Food Prot. 2005, 68, 1712–1719. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, T.; Para, R.; Gonipeta, B.; Reitmeyer, M.; He, Y.; Srkalovic, I.; Ng, P.K.W.; Gangur, V. Effect of extrusion processing on immune activation properties of hazelnut protein in a mouse model. Int. J. Food Sci. Nutr. 2016, 67, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Álvarez, J.; Guillamón, E.; Crespo, J.F.; Cuadrado, C.; Burbano, C.; Rodríguez, J.; Fernández, C.; Muzquiz, M. Effects of extrusion, boiling, autoclaving, and microwave heating on lupine allergenicity. J. Agric. Food Chem. 2005, 53, 1294–1298. [Google Scholar] [CrossRef] [PubMed]
- Milán-Carrillo, J.; Montoya-Rodríguez, A.; Gutiérrez-Dorado, R.; Perales-Sánchez, X.; Reyes-Moreno, C. Optimization of Extrusion Process for Producing High Antioxidant Instant Amaranth (Amaranthus hypochondriacus L.) Flour Using Response Surface Methodology. Appl. Math. 2012, 3, 1516–1525. [Google Scholar] [CrossRef]
- Osborne, T.B. The vegetable proteins. In Monographs on Biochemistry; Longmans Green and Co.: London, UK, 1924; Volume 43. [Google Scholar]
- Dearman, R.J.; Kimber, I. Determination of protein allergenicity: Studies in mice. Toxicol. Lett. 2001, 120, 181–186. [Google Scholar] [CrossRef]
- Dearman, R.J.; Caddick, H.; Stone, S.; Basketter, D.A.; Kimber, I. Characterization of antibody responses induced in rodents by exposure to food proteins: Influence of route of exposure. Toxicology 2001, 167, 217–231. [Google Scholar] [CrossRef]
- Chen, C.; Sun, N.; Li, Y.; Jia, X. A BALB/c mouse model for assessing the potential allergenicity of proteins: Comparison of allergen dose, sensitization frequency, timepoint and sex. Food Chem. Toxicol. 2013, 62, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Chávez, F.; Calderón de la Barca, A.M.; Islas-Rubio, A.R.; Marti, A.; Marengo, M.; Pagani, M.A.; Bonomi, F.; Iametti, S. Molecular rearrangements in extrusion processes for the production of amaranth-enriched, gluten-free rice pasta. LWT—Food Sci. Technol. 2012, 47, 421–426. [Google Scholar] [CrossRef]
- Morin, S.; Bernard, H.; Przybylski-Nicaise, L.; Corthier, G.; Rabot, S.; Wal, J.M.; Hazebrouck, S. Allergenic and immunogenic potential of cow’s milk β-lactoglobulin and caseins evidenced without adjuvant in germ-free mice. Mol. Nutr. Food Res. 2011, 55, 1700–1707. [Google Scholar] [CrossRef] [PubMed]
- Ladics, G.S.; Knippels, L.M.J.; Penninks, A.H.; Bannon, G.A.; Goodman, R.E.; Herouet-Guicheney, C. Review of animal models designed to predict the potential allergenicity of novel proteins in genetically modified crops. Regul. Toxicol. Pharmacol. 2010, 56, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Micinski, J.; KowaIski, I.M.; Zwierzchowski, G.; Szarek, J.; Pieroyñski, B.; Zablocka, E. Characteristics of cow’s milk proteins including allergenic properties and methods for its reduction. Polish Ann. Med. 2013, 20, 69–76. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cárdenas-Torres, F.I.; Reyes-Moreno, C.; Vergara-Jiménez, M.d.J.; Cuevas-Rodríguez, E.O.; Milán-Carrillo, J.; Gutiérrez-Dorado, R.; Arámburo-Gálvez, J.G.; Ontiveros, N.; Cabrera-Chávez, F. Assessing the Sensitizing and Allergenic Potential of the Albumin and Globulin Fractions from Amaranth (Amaranthus hypochondriacus) Grains before and after an Extrusion Process. Medicina 2019, 55, 72. https://doi.org/10.3390/medicina55030072
Cárdenas-Torres FI, Reyes-Moreno C, Vergara-Jiménez MdJ, Cuevas-Rodríguez EO, Milán-Carrillo J, Gutiérrez-Dorado R, Arámburo-Gálvez JG, Ontiveros N, Cabrera-Chávez F. Assessing the Sensitizing and Allergenic Potential of the Albumin and Globulin Fractions from Amaranth (Amaranthus hypochondriacus) Grains before and after an Extrusion Process. Medicina. 2019; 55(3):72. https://doi.org/10.3390/medicina55030072
Chicago/Turabian StyleCárdenas-Torres, Feliznando Isidro, Cuauhtémoc Reyes-Moreno, Marcela de Jesús Vergara-Jiménez, Edith Oliva Cuevas-Rodríguez, Jorge Milán-Carrillo, Roberto Gutiérrez-Dorado, Jesús Gilberto Arámburo-Gálvez, Noé Ontiveros, and Francisco Cabrera-Chávez. 2019. "Assessing the Sensitizing and Allergenic Potential of the Albumin and Globulin Fractions from Amaranth (Amaranthus hypochondriacus) Grains before and after an Extrusion Process" Medicina 55, no. 3: 72. https://doi.org/10.3390/medicina55030072