The Novel Nature Microtubule Inhibitor Ivalin Induces G2/M Arrest and Apoptosis in Human Hepatocellular Carcinoma SMMC-7721 Cells In Vitro

 and
and 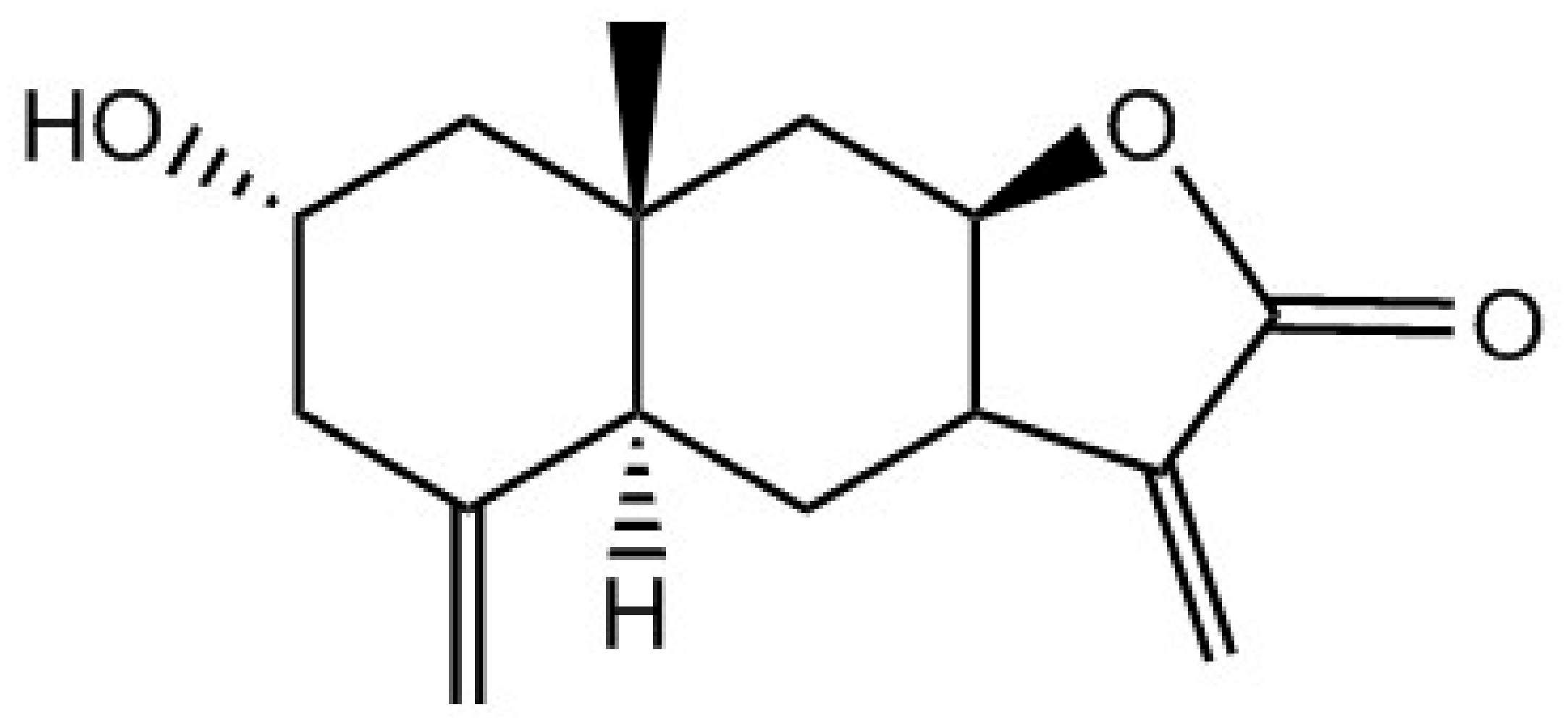{kind=link}
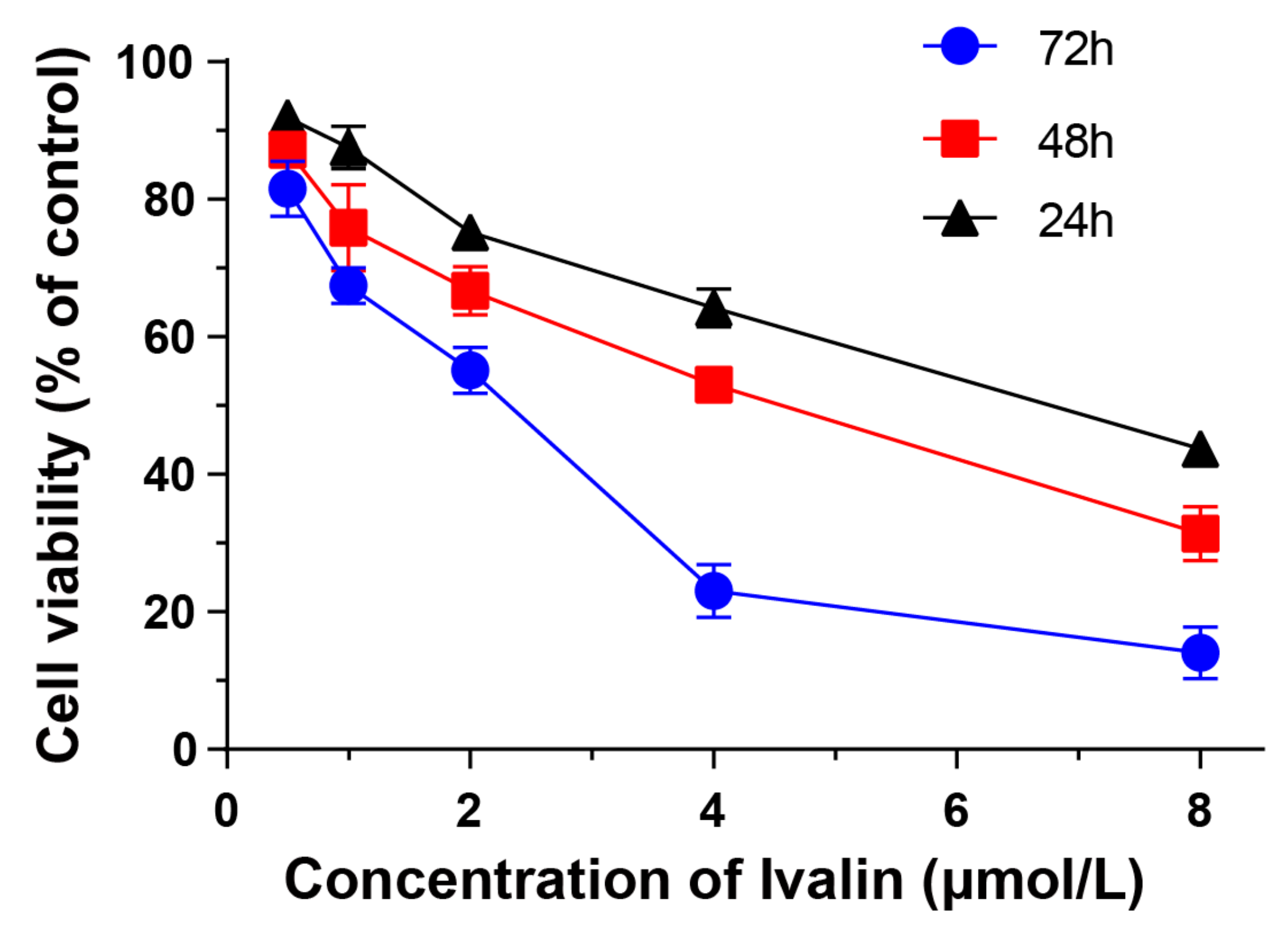{kind=link}
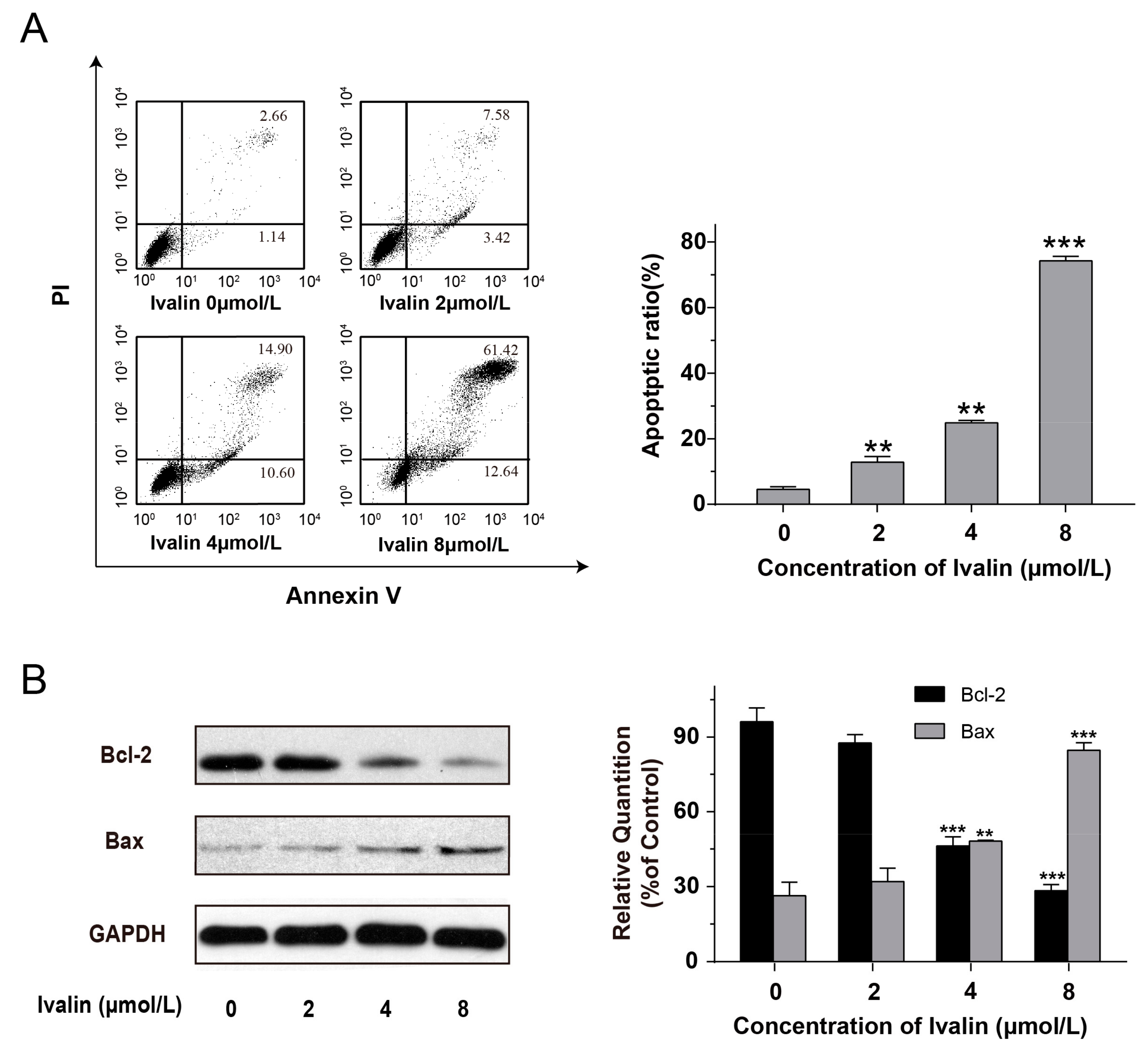{kind=link}
{kind=link}
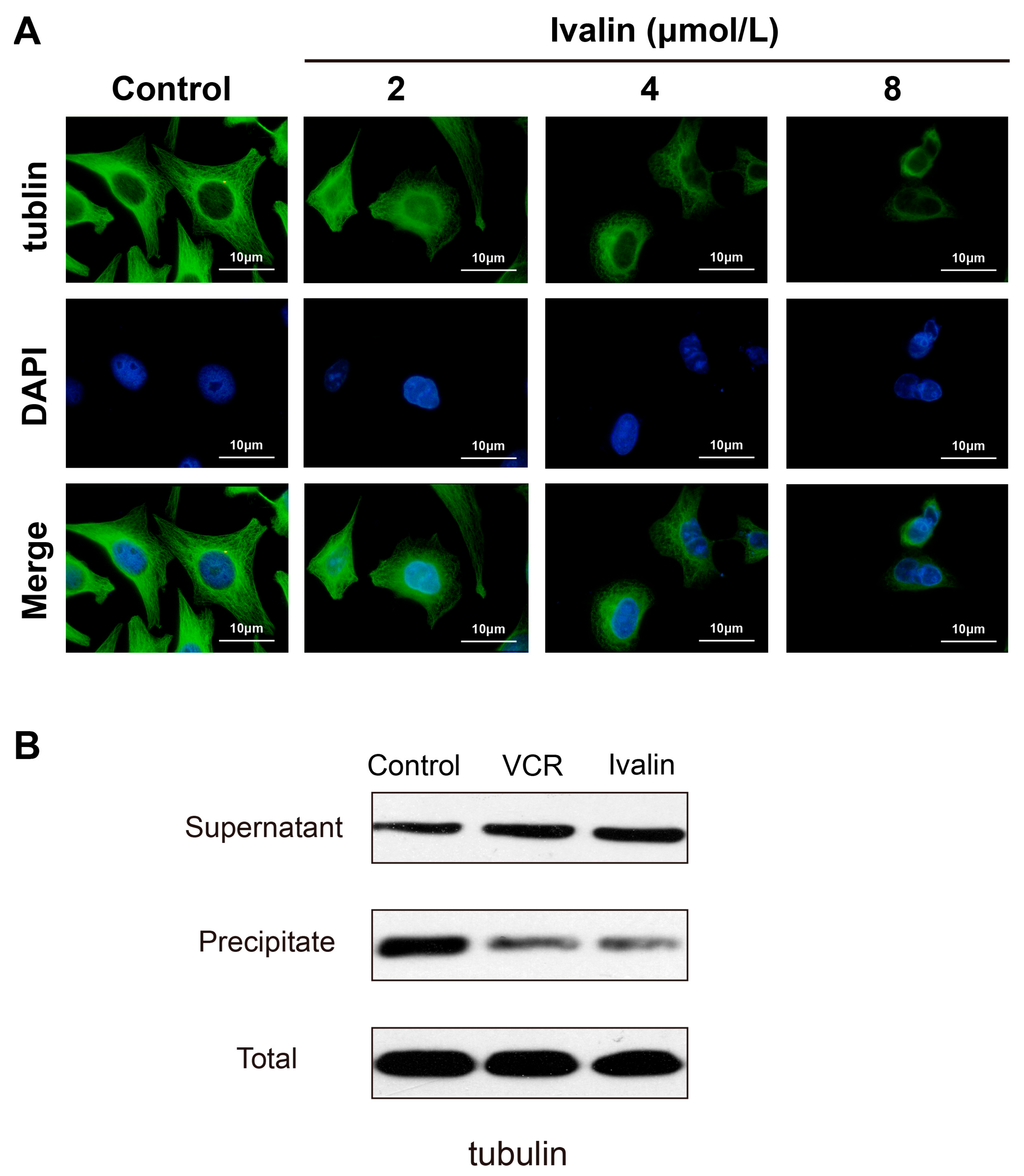{kind=link}
Abstract
Share and Cite
Liu, F.; Lin, S.; Zhang, C.; Ma, J.; Han, Z.; Jia, F.; Xie, W.; Li, X. The Novel Nature Microtubule Inhibitor Ivalin Induces G2/M Arrest and Apoptosis in Human Hepatocellular Carcinoma SMMC-7721 Cells In Vitro. Medicina 2019, 55, 470. https://doi.org/10.3390/medicina55080470
Liu F, Lin S, Zhang C, Ma J, Han Z, Jia F, Xie W, Li X. The Novel Nature Microtubule Inhibitor Ivalin Induces G2/M Arrest and Apoptosis in Human Hepatocellular Carcinoma SMMC-7721 Cells In Vitro. Medicina. 2019; 55(8):470. https://doi.org/10.3390/medicina55080470
Chicago/Turabian StyleLiu, Fangyuan, Shiqi Lin, Caiyun Zhang, Jiahui Ma, Zhuo Han, Fujuan Jia, Weidong Xie, and Xia Li. 2019. "The Novel Nature Microtubule Inhibitor Ivalin Induces G2/M Arrest and Apoptosis in Human Hepatocellular Carcinoma SMMC-7721 Cells In Vitro" Medicina 55, no. 8: 470. https://doi.org/10.3390/medicina55080470
APA StyleLiu, F., Lin, S., Zhang, C., Ma, J., Han, Z., Jia, F., Xie, W., & Li, X. (2019). The Novel Nature Microtubule Inhibitor Ivalin Induces G2/M Arrest and Apoptosis in Human Hepatocellular Carcinoma SMMC-7721 Cells In Vitro. Medicina, 55(8), 470. https://doi.org/10.3390/medicina55080470