
Divergent Directionality of Immune Cell-Specific Protein Expression between Bipolar Lithium Responders and Non-Responders Revealed by Enhanced Flow Cytometry


 and
and
Abstract
:1. Introduction
2. Methods
2.1. Study Design
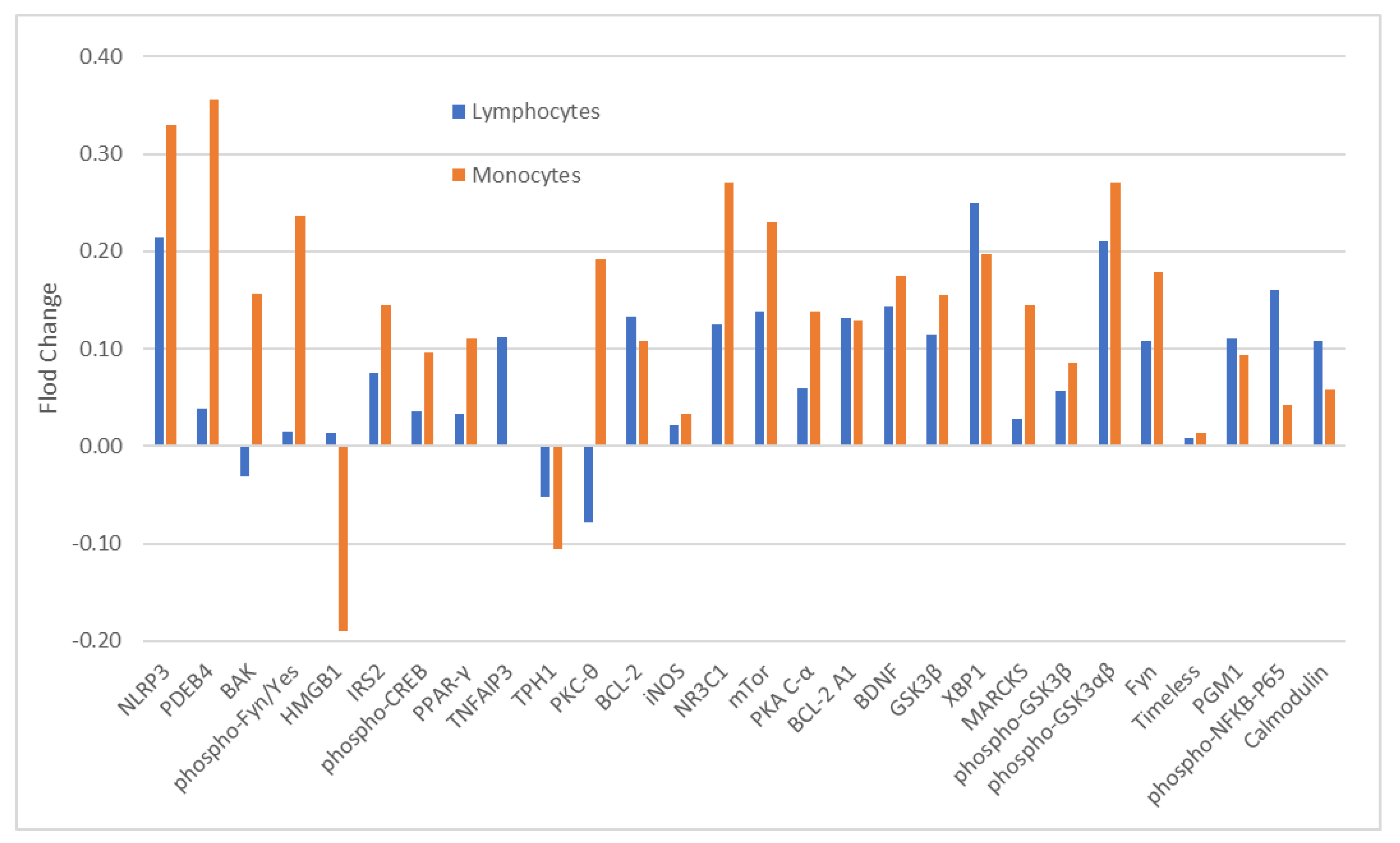
2.2. Rationale of Using Monocytes and CD4+ Lymphocytes
2.3. Antibodies and Cytometric Analyses
2.4. Normalization of Raw Data
2.5. Statistical Analysis
3. Results
3.1. MFR of Analytes at Baseline: ITT-Responders vs. ITT-Non-Responders
3.2. MFR of Analytes at Baseline and at the End of Study: Completed Responders vs. Completed Non-Responders
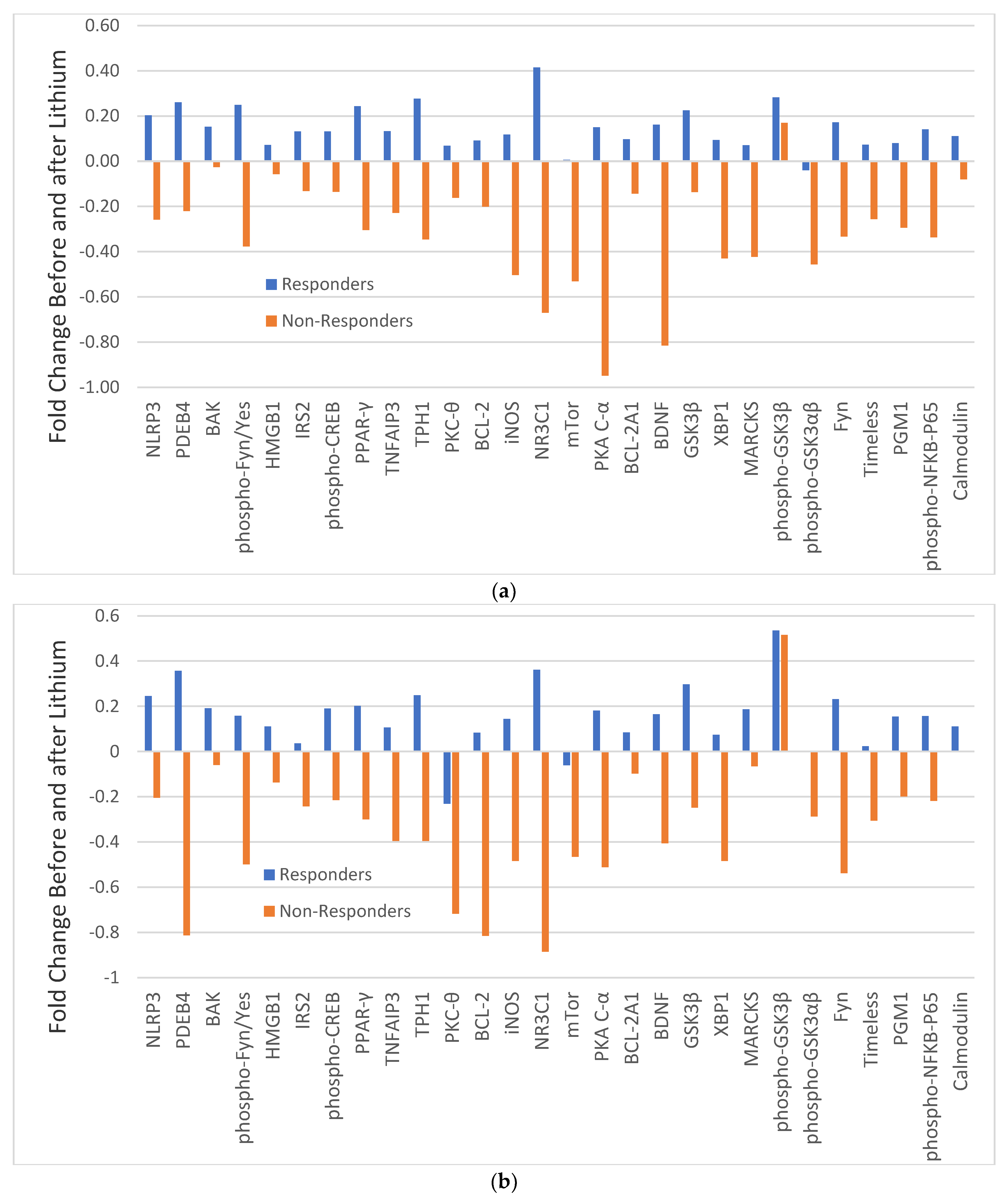
3.3. MFR of Analytes before and after Lithium in Completed Responders and Completed Non-Responders
3.4. Fold Change in MFR of Analytes before and after Lithium in Responders and Non-Responders
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yatham, L.N.; Kennedy, S.H.; Parikh, S.V.; Schaffer, A.; Bond, D.J.; Frey, B.N.; Sharma, V.; Goldstein, B.I.; Rej, S.; Beaulieu, S.; et al. Canadian Network for Mood and Anxiety Treatments (CANMAT) and International Society for Bipolar Disorders (ISBD) 2018 guidelines for the management of patients with bipolar disorder. Bipolar Disord. 2018, 20, 97–170. [Google Scholar] [CrossRef]
- Gao, K.; Goto, T.; Yuan, C.; Brownrigg, B.; Conroy, C.; Chan, P.K.; Serrano, M.B.; Ganocy, S.J.; Fang, F.; Calabrese, J.R. A pilot study of the effectiveness of lithium versus quetiapine immediate release monotherapy in patients with bipolar spectrum disorders. J. Clin. Psychopharmacol. 2018, 38, 422–434. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Maihofer, A.X.; Stapp, E.; Ritchey, M.; Alliey-Rodriguez, N.; Anand, A.; Balaraman, Y.; Berrettini, W.H.; Bertram, H.; Bhattacharjee, A.; et al. Clinical predictors of non-response to lithium treatment in the Pharmacogenomics of Bipolar Disorder (PGBD) study. Bipolar Disord. 2021, 23, 821–831. [Google Scholar] [CrossRef] [PubMed]
- van der Loos, M.L.; Mulder, P.G.; Hartong, E.G.; Blom, M.B.; Vergouwen, A.C.; de Keyzer, H.J.; Notten, P.J.; Luteijn, M.L.; Timmermans, M.A.; Vieta, E.; et al. Efficacy and safety of lamotrigine as add-on treatment to lithium in bipolar depression: A multicenter, double-blind, placebo-controlled trial. J. Clin. Psychiatry 2009, 70, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Young, A.H.; McElroy, S.L.; Bauer, M.; Philips, N.; Chang, W.; Olausson, B.; Paulsson, B.; Brecher, M.; EMBOLDEN I (Trial 001) Investigators. A double-blind, placebo-controlled study of quetiapine and lithium monotherapy in adults in the acute phase of bipolar depression (EMBOLDEN I). J. Clin. Psychiatry 2010, 71, 150–162. [Google Scholar] [CrossRef]
- Gao, K.; Calabrese, J.R. The mechanisms of action of lithium in bipolar disorder. In Neurobiology of Bipolar Disorder Road to Novel Therapeutics; Quevedo, J., Carvalho, A.F., Vieta, E., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 357–364. [Google Scholar]
- Hui, T.P.; Kandola, A.; Shen, L.; Lewis, G.; Osborn, D.P.J.; Geddes, J.R.; Hayes, J.F. A systematic review and meta-analysis of clinical predictors of lithium response in bipolar disorder. Acta Psychiatr. Scand. 2019, 140, 94–115. [Google Scholar] [CrossRef] [Green Version]
- Rybakowski, J.K. Challenging the negative perception of lithium and optimizing its long-term administration. Front. Mol. Neurosci. 2018, 11, 349. [Google Scholar] [CrossRef]
- Can, A.; Schulze, T.G.; Gould, T.D. Molecular actions and clinical pharmacogenetics of lithium therapy. Pharmacol. Biochem. Behav. 2014, 123, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Tondo, L.; Baldessarini, R.J. Antisuicidal effects in mood disorders: Are they unique to lithium? Pharmacopsychiatry 2018, 51, 177–188. [Google Scholar] [CrossRef]
- Rybakowski, J.K. Response to lithium in bipolar disorder: Clinical and genetic findings. ACS Chem. Neurosci. 2014, 5, 413–421. [Google Scholar] [CrossRef]
- Budde, M.; Degner, D.; Brockmöller, J.; Schulze, T. Pharmacogenomic aspects of bipolar disorder: An update. Eur. Neuropsychopharmacol. 2017, 27, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Amare, A.T.; Schubert, K.O.; Baune, B.T. Pharmacogenomics in the treatment of mood disorders: Strategies and opportunities for personalized psychiatry. EPMA J. 2017, 8, 211–227. [Google Scholar] [CrossRef] [Green Version]
- International Consortium on Lithium Genetics (ConLi+Gen); Amare, A.T.; Schubert, K.O.; Hou, L.; Clark, S.R.; Papiol, S.; Heilbronner, U.; Degenhardt, F.; Tekola-Ayele, F.; Hsu, Y.H.; et al. Association of polygenic score for schizophrenia and HLA antigen and inflammation genes with response to lithium in bipolar affective disorder: A genome-wide association study. JAMA Psychiatry 2018, 75, 65–74. [Google Scholar]
- Beech, R.D.; Leffert, J.J.; Lin, A.; Sylvia, L.G.; Umlauf, S.; Mane, S.; Zhao, H.; Bowden, C.; Calabrese, J.R.; Friedman, E.S.; et al. Gene-expression differences in peripheral blood between lithium responders and non-responders in the Lithium Treatment-Moderate dose Use Study (LiTMUS). Pharm. J. 2013, 14, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Anand, A.; McClintick, J.N.; Murrell, J.; Karne, H.; Nurnberger, J.I.; Edenberg, H.J. Effects of lithium monotherapy for bipolar disorder on gene expression in peripheral lymphocytes. Complex Psychiatry 2016, 2, 115–123. [Google Scholar] [CrossRef] [Green Version]
- Kittel-Schneider, S.; Hilscher, M.; Scholz, C.-J.; Weber, H.; Grünewald, L.; Schwarz, R.; Chiocchetti, A.G.; Reif, A. Lithium-induced gene expression alterations in two peripheral cell models of bipolar disorder. World J. Biol. Psychiatry 2019, 20, 462–475. [Google Scholar] [CrossRef]
- Papadima, E.M.; Niola, P.; Melis, C.; Pisanu, C.; Congiu, D.; Cruceanu, C.; Lopez, J.P.; Turecki, G.; Ardau, R.; Severino, G.; et al. Evidence towards RNA Binding Motif (RNP1, RRM) Protein 3 (RBM3) as a potential biomarker of lithium response in bipolar disorder patients. J. Mol. Neurosci. 2017, 62, 304–308. [Google Scholar] [CrossRef]
- Costa, M.; Squassina, A.; Piras, I.S.; Pisanu, C.; Congiu, D.; Niola, P.; Angius, A.; Chillotti, C.; Ardau, R.; Severino, G.; et al. Preliminary transcriptome analysis in lymphoblasts from cluster headache and bipolar disorder patients implicates dysregulation of circadian and serotonergic genes. J. Mol. Neurosci. 2015, 56, 688–695. [Google Scholar] [CrossRef]
- Pisanu, C.; Papadima, E.M.; Melis, C.; Congiu, D.; Loizedda, A.; Orrù, N.; Calza, S.; Carcassi, C.; Severino, G.; Ardau, R.; et al. Whole genome expression analyses of miRNAs and mRNAs suggest the involvement of miR-320a and miR-155-3p and their targeted genes in lithium response in bipolar disorder. Int. J. Mol. Sci. 2019, 20, 6040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soeiro-De-Souza, M.G.; Gold, P.W.; Brunoni, A.R.; de Sousa, R.T.; Zanetti, M.V.; Carvalho, A.F.; Gattaz, W.F.; Machado-Vieira, R.; Teixeira, A.L. Lithium decreases plasma adiponectin levels in bipolar depression. Neurosci. Lett. 2014, 564, 111–114. [Google Scholar] [CrossRef]
- Teixeira, A.L.; de Sousa, R.T.; Zanetti, M.V.; Brunoni, A.R.; Busatto, G.F.; Zarate, C.A., Jr.; Gattaz, W.F.; Machado-Vieira, R. Increased plasma levels of soluble TNF receptors 1 and 2 in bipolar depression and impact of lithium treatment. Hum. Psychopharmacol. 2015, 30, 52–56. [Google Scholar] [CrossRef]
- Loch, A.A.; Zanetti, M.V.; de Sousa, R.T.; Chaim, T.M.; Serpa, M.H.; Gattaz, W.F.; Teixeira, A.L.; Machado-Vieira, R. Elevated neurotrophin-3 and neurotrophin 4/5 levels in unmedicated bipolar depression and the effects of lithium. Prog. Neuropsychopharmacol. Biol. Psychiatry 2015, 56, 243–246. [Google Scholar] [CrossRef]
- van den Ameele, S.; van Diermen, L.; Staels, W.; Coppens, V.; Dumont, G.; Sabbe, B.; Morrens, M. The effect of mood-stabilizing drugs on cytokine levels in bipolar disorder: A systematic review. J. Affect. Disord. 2016, 203, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, S.D.; Williams, A.J.; Blacker, C.J.; Voort, J.L.V.; Schak, K.M.; Nemeroff, C.B.; Widge, A.S.; Tohen, M. Putative biological predictors of treatment response in bipolar disorders. Pers. Med. Psychiatry 2017, 1, 39–58. [Google Scholar] [CrossRef] [Green Version]
- Hibar, D.P.; The ENIGMA Bipolar Disorder Working Group; Westlye, L.T.; Doan, N.T.; Jahanshad, N.; Cheung, J.W.; Ching, C.R.K.; Versace, A.; Bilderbeck, A.C.; Uhlmann, A.; et al. Cortical abnormalities in bipolar disorder: An MRI analysis of 6503 individuals from the ENIGMA Bipolar Disorder Working Group. Mol. Psychiatry 2018, 23, 932–942. [Google Scholar] [CrossRef] [PubMed]
- Atagün, M.I. Brain oscillations in bipolar disorder and lithium-induced changes. Neuropsychiatr. Dis. Treat. 2016, 12, 589–601. [Google Scholar] [CrossRef] [Green Version]
- Gao, K.; Calabrese, J.R. Pharmacogenetics of lithium response: Close to clinical practice? Lancet 2016, 387, 1034–1036. [Google Scholar] [CrossRef] [PubMed]
- Vieta, E.; Berk, M.; Schulze, T.G.; Carvalho, A.F.; Suppes, T.; Calabrese, J.R.; Gao, K.; Miskowiak, K.W.; Grande, I. Bipolar disorders. Nat. Rev. Dis. Prim. 2018, 4, 18008. [Google Scholar] [CrossRef]
- Mertens, J.; Wang, Q.W.; Kim, Y.; Yu, D.X.; Pham, S.; Yang, B.; Zheng, Y.; Diffenderfer, K.E.; Zhang, J.; Soltani, S.; et al. Differential responses to lithium in hyperexcitable neurons from patients with bipolar disorder. Nature 2015, 527, 95–99. [Google Scholar] [CrossRef] [Green Version]
- Stern, S.; Santos, R.; Marchetto, M.C.; Mendes, A.P.D.; Rouleau, G.A.; Biesmans, S.; Wang, Q.-W.; Yao, J.; Charnay, P.; Bang, A.G.; et al. Neurons derived from patients with bipolar disorder divide into intrinsically different sub-populations of neurons, predicting the patients’ responsiveness to lithium. Mol. Psychiatry 2017, 23, 1453–1465. [Google Scholar] [CrossRef] [PubMed]
- Tobe, B.T.D.; Crain, A.M.; Winquist, A.M.; Calabrese, B.; Makihara, H.; Zhao, W.N.; Lalonde, J.; Nakamura, H.; Konopaske, G.; Sidor, M.; et al. Proc Probing the lithium-response pathway in hiPSCs implicates the phosphoregulatory set-point for a cytoskeletal modulator in bipolar pathogenesis. Proc. Natl. Acad. Sci. USA 2017, 114, E4462–E4471. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, M.J.; Wei, H.; Nievergelt, C.M.; Stautland, A.; Maihofer, A.X.; Welsh, D.K.; Shilling, P.; Alda, M.; Alliey-Rodriguez, N.; Anand, A.; et al. Chronotype and cellular circadian rhythms predict the clinical response to lithium maintenance treatment in patients with bipolar disorder. Neuropsychopharmacology 2019, 44, 620–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Federoff, M.; McCarthy, M.J.; Anand, A.; Berrettini, W.H.; Bertram, H.; Bhattacharjee, A.; Calkin, C.V.; Conroy, C.; Coryell, W.H.; D’Arcangelo, N.; et al. Correction of depression-associated circadian rhythm abnormalities is associated with lithium response in bipolar disorder. Bipolar Disord. 2022, 24, 521–529. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, J.; Wang, X.; Zhu, J.; Liu, Q.; Shi, Z.; Chambers, M.C.; Zimmerman, L.J.; Shaddox, K.F.; Kim, S.; et al. Proteogenomic characterization of human colon and rectal cancer. Nature 2014, 513, 382–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hukelmann, J.L.; Anderson, K.E.; Sinclair, L.V.; Grzes, K.M.; Murillo, A.B.; Hawkins, P.T.; Stephens, L.R.; Lamond, A.I.; Cantrell, D.A. The cytotoxic T cell proteome and its shaping by the kinase mTOR. Nat. Immunol. 2016, 17, 104–112. [Google Scholar] [CrossRef] [Green Version]
- Darmanis, S.; Gallant, C.J.; Marinescu, V.D.; Niklasson, M.; Segerman, A.; Flamourakis, G.; Fredriksson, S.; Assarsson, E.; Lundberg, M.; Nelander, S.; et al. Simultaneous multiplexed measurement of RNA and proteins in single cells. Cell. Rep. 2016, 14, 380–389. [Google Scholar] [CrossRef]
- Barbosa, I.G.; Nogueira, C.R.; Rocha, N.P.; Queiroz, A.L.; Vago, J.P.; Tavares, L.P.; Assis, F.; Fagundes, C.T.; Huguet, R.B.; Bauer, M.E.; et al. Altered intracellular signaling cascades in peripheral blood mononuclear cells from BD patients. J. Psychiatr. Res. 2013, 47, 1949–1954. [Google Scholar] [CrossRef] [PubMed]
- Brietzke, E.; Stertz, L.; Fernandes, B.S.; Kauer-Sant’Anna, M.; Mascarenhas, M.; Vargas, A.E.; Chies, J.A.; Kapczinski, F. Comparison of cytokine levels in depressed, manic and euthymic patients with bipolar disorder. J. Affect. Disord. 2009, 116, 214–217. [Google Scholar] [CrossRef] [PubMed]
- do Prado, C.H.; Rizzo, L.B.; Wieck, A.; Lopes, R.P.; Teixeira, A.L.; Grassi-Oliveira, R.; Bauer, M.E. Reduced regulatory T cells are associated with higher levels of Th1/TH17 cytokines and activated MAPK in type 1 bipolar disorder. Psychoneuroendocrinology 2013, 38, 667–676. [Google Scholar] [CrossRef]
- Guloksuz, S.; Cetin, E.A.; Cetin, T.; Deniz, G.; Oral, E.T.; Nutt, D.J. Cytokine levels in euthymic bipolar patients. J. Affect. Disord. 2010, 126, 458–462. [Google Scholar] [CrossRef]
- Wieck, A.; Grassi-Oliveira, R.; Prado, C.H.D.; Rizzo, L.B.; de Oliveira, A.S.; Kommers-Molina, J.; Viola, T.W.; Teixeira, A.L.; Bauer, M.E. Differential neuroendocrine and immune responses to acute psychosocial stress in women with type 1 bipolar disorder. Brain Behav. Immun. 2013, 34, 47–55. [Google Scholar] [CrossRef]
- Han, Y.; Gu, Y.; Zhang, A.C.; Lo, Y.-H. Review: Imaging technologies for flow cytometry. Lab Chip 2016, 16, 4639–4647. [Google Scholar] [CrossRef] [Green Version]
- McKinnon, K.M. Flow Cytometry: An Overview. Curr. Protoc. Immunol. 2018, 120, 5.1.1–5.1.11. [Google Scholar] [CrossRef]
- Gao, K.; Ayati, M.; Koyuturk, M.; Calabrese, J.R.; Ganocy, S.J.; Kaye, N.M.; Lazarus, H.M.; Christian, E.; Kaplan, D. Protein biomarkers in monocytes and CD4 + Lymphocytes for predicting lithium treatment response of bipolar disorder: A feasibility study with tyramine-based signal-amplified flow cytometry. Psychopharmacol. Bull. 2022, 52, 8–35. [Google Scholar]
- Kaplan, D.; Smith, D. Enzymatic amplification staining for flow cytometric analysis of cell surface molecules. Cytometry 2000, 40, 81–85. [Google Scholar] [CrossRef]
- Kaplan, D.; Husel, W.; Meyerson, H. Immunophenotypic analysis with enhanced sensitivity of detection by enzymatic amplification staining. Clin. Lab. Med. 2001, 21, 763–778. [Google Scholar]
- Kaplan, D.; Meyerson, H.; Lewandowska, K. High resolution immunophenotypic analysis of chronic lymphocytic leukemic cells by enzymatic amplification staining. Am. J. Clin. Pathol. 2001, 116, 429–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, D. Enzymatic amplification staining for single cell analysis: Applied to in situ hybridization. J. Immunol. Methods 2003, 283, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, D. Enzymatic Amplification Staining for Cell Surface Antigens. Curr. Protoc. Cytom. 2003, 23, 6.14.1–6.14.10. [Google Scholar] [CrossRef]
- Kaplan, D.; Meyerson, H.; Husel, W.; Lewandowska, K.; MacLennan, G. D cyclins in lymphocytes. Cytom. Part A 2004, 63A, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Meyerson, H.J.; MacLennan, G.; Husel, W.; Tse, W.; Lazarus, H.M.; Kaplan, D. D Cyclins in CD5+ B-Cell lymphoproliferative disorders: Cyclin D1 and Cyclin D2 identify diagnostic groups and Cyclin D1 correlates with ZAP-70 expression in chronic lymphocytic leukemia. Am. J. Clin. Pathol. 2006, 125, 241–250. [Google Scholar] [CrossRef]
- Liew, C.-C.; Ma, J.; Tang, H.-C.; Zheng, R.; Dempsey, A.A. The peripheral blood transcriptome dynamically reflects system wide biology: A potential diagnostic tool. J. Lab. Clin. Med. 2006, 147, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, P.F.; Fan, C.; Perou, C. Evaluating the comparability of gene expression in blood and brain. Am. J. Med Genet. Part B Neuropsychiatr. Genet. 2006, 141B, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Rollins, B.; Martin, M.V.; Morgan, L.; Vawter, M.P. Analysis of whole genome biomarker expression in blood and brain. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2010, 153B, 919–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tylee, D.S.; Kawaguchi, D.M.; Glatt, S.J. On the outside, looking in: A review and evaluation of the comparability of blood and brain “-omes”. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2013, 162B, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Spiliotaki, M.; Salpeas, V.; Malitas, P.; Alevizos, V.; Moutsatsou, P. Altered glucocorticoid receptor signaling cascade in lymphocytes of bipolar disorder patients. Psychoneuroendocrinology 2006, 31, 748–760. [Google Scholar] [CrossRef]
- Bei, E.; Salpeas, V.; Pappa, D.; Anagnostara, C.; Alevizos, V.; Moutsatsou, P. Phosphorylation status of glucocorticoid receptor, heat shock protein 70, cytochrome c and Bax in lymphocytes of euthymic, depressed and manic bipolar patients. Psychoneuroendocrinology 2009, 34, 1162–1175. [Google Scholar] [CrossRef]
- Hunsberger, J.G.; Chibane, F.L.; Elkahloun, A.G.; Henderson, R.; Singh, R.; Lawson, J.; Cruceanu, C.; Nagarajan, V.; Turecki, G.; Squassina, A.; et al. Novel integrative genomic tool for interrogating lithium response in bipolar disorder. Transl. Psychiatry 2015, 5, e504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marie-Claire, C.; Lejeune, F.X.; Mundwiller, E.; Ulveling, D.; Moszer, I.; Bellivier, F.; Etain, B. A DNA methylation signature discriminates between excellent and non-response to lithium in patients with bipolar disorder type 1. Sci. Rep. 2020, 10, 12239. [Google Scholar] [CrossRef]
- Eugene, A.R.; Masiak, J.; Eugene, B. Predicting lithium treatment response in bipolar patients using gender-specific gene expression biomarkers and machine learning. F1000Research 2018, 7, 474. [Google Scholar] [CrossRef]
- Munkholm, K.; Peijs, L.; Vinberg, M.; Kessing, L.V. A composite peripheral blood gene expression measure as a potential diagnostic biomarker in bipolar disorder. Transl. Psychiatry 2015, 5, e614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- So, J.; Warsh, J.J.; Li, P.P. Impaired endoplasmic reticulum stress response in B-Lymphoblasts from patients with Bipolar-I disorder. Biol. Psychiatry 2007, 62, 141–147. [Google Scholar] [CrossRef]
- Wieck, A.; Grassi-Oliveira, R.; Prado, C.H.D.; Viola, T.W.; Petersen, L.E.; Porto, B.; Teixeira, A.L.; Bauer, M.E. Toll-like receptor expression and function in type I bipolar disorder. Brain Behav. Immun. 2016, 54, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Amoruso, A.; Bardelli, C.; Cattaneo, C.I.; Fresu, L.G.; Manzetti, E.; Brunelleschi, S. Neurokinin (NK)-1 receptor expression in monocytes from bipolar disorder patients: A pilot study. J. Affect. Disord. 2015, 178, 188–192. [Google Scholar] [CrossRef]
- Pfaffenseller, B.; Wollenhaupt-Aguiar, B.; Fries, G.R.; Colpo, G.D.; Burque, R.K.; Bristot, G.; Ferrari, P.; Ceresér, K.M.M.; Rosa, A.R.; Klamt, F.; et al. Impaired endoplasmic reticulum stress response in bipolar disorder: Cellular evidence of illness progression. Int. J. Neuropsychopharmacol. 2014, 17, 1453–1463. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, I.G.; Rocha, N.P.; Assis, F.; Vieira, E.L.M.; Soares, J.C.; Bauer, M.E.; Teixeira, A.L. Monocyte and lymphocyte activation in bipolar disorder: A new piece in the puzzle of immune dysfunction in mood disorders. Int. J. Neuropsychopharmacol. 2014, 18, pyu021. [Google Scholar] [CrossRef] [Green Version]
- Maecker, H.T.; McCoy, J.P.; Nussenblatt, R. Standardizing immunophenotyping for the Human Immunology Project. Nat. Rev. Immunol. 2012, 12, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Miklowitz, D.J.; Portnoff, L.C.; Armstrong, C.C.; Keenan-Miller, D.; Breen, E.C.; Muscatell, K.A.; Eisenberger, N.I.; Irwin, M.R. Inflammatory cytokines and nuclear factor-kappa B activation in adolescents with bipolar and major depressive disorders. Psychiatry Res. 2016, 241, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Lambert, C.; Preijers, F.W.M.B.; Yanikkaya Demirel, G.; Sack, U. Monocytes and macrophages in flow: An ESCCA initiative on advanced analyses of monocyte lineage using flow cytometry. Cytom. B Clin. Cytom. 2017, 92, 180–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Sousa, R.T.; Zanetti, M.V.; Talib, L.L.; Serpa, M.H.; Chaim, T.M.; Carvalho, A.F.; Brunoni, A.R.; Busatto, G.F.; Gattaz, W.F.; Machado-Vieira, R. Lithium increases platelet serine-9 phosphorylated GSK-3β levels in drug-free bipolar disorder during depressive episodes. J. Psychiatr. Res. 2015, 62, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Ladeira, R.B.; Joaquim, H.P.; Talib, L.L.; Nunes, P.V.; Forlenza, O.V. Higher proportion of inactive Gsk3β in platelets of elderly patients with bipolar disorder: An effect of treatment? Braz. J. Psychiatry 2013, 35, 274–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Liu, M.; Cai, Z.; Wang, G.; Li, X. Regulation of glycogen synthase kinas 3 during bipolar mania treatment. Bipolar Disord. 2010, 12, 741–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Friedman, A.B.; Zhu, W.; Wang, L.; Boswell, S.; May, R.S.; Davis, L.L.; Jope, R.S. Lithium regulates glycogen synthase kinase-3beta in human peripheral blood mononuclear cells: Implication in the treatment of bipolar disorder. Biol. Psychiatry 2007, 61, 216–222. [Google Scholar] [CrossRef]
- Pandey, G.N.; Ren, X.; Rizavi, H.S.; Dwivedi, Y.J. Glycogen synthase kinase-3beta in the platelets of patients with mood disorders: Effect of treatment. J. Psychiatr. Res. 2010, 44, 143–148. [Google Scholar] [CrossRef]
- Hoeflich, K.P.; Luo, J.; Rubie, E.A.; Tsao, M.S.; Jin, O.; Woodgett, J.R. Requirement for glycogen synthase kinase-3beta in cell survival and NF-kappaB activation. Nature 2000, 406, 86–90. [Google Scholar] [CrossRef]
- Hofmann, C.; Dunger, N.; Schölmerich, J.; Falk, W.; Obermeier, F. Glycogen synthase kinase 3-β: A master regulator of toll-like receptor-mediated chronic intestinal inflammation. Inflamm. Bowel. Dis. 2010, 16, 1850–1858. [Google Scholar] [CrossRef] [PubMed]
- Sakrajda, K.; Szczepankiewicz, A. Inflammation-related changes in mood disorders and the immunomodulatory role of lithium. Int. J. Mol. Sci. 2021, 22, 1532. [Google Scholar] [CrossRef]
- Wu, T.-N.; Lee, C.-S.; Wu, B.-J.; Sun, H.-J.; Chang, C.-H.; Chen, C.-Y.; Chen, C.-K.; Wu, L.S.-H.; Cheng, A.T.-A. Immunophenotypes associated with bipolar disorder and lithium treatment. Sci. Rep. 2019, 9, 17453. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Huang, K.; Liu, X.; Liu, J.; Lu, X.; Tao, K.; Wang, G.; Wang, J. Lithium chloride suppresses colorectal cancer cell survival and proliferation through ROS/GSK-3beta/NF-kappaB signaling pathway. Oxidative Med. Cell. Longev. 2014, 2014, 241864. [Google Scholar] [CrossRef] [Green Version]
- Nassar, A.; Azab, A.N. Effects of Lithium on Inflammation. ACS Chem. Neurosci. 2014, 5, 451–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troib, A.; Azab, A.N. Effects of psychotropic drugs on Nuclear Factor kappa B. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 1198–1208. [Google Scholar] [PubMed]
- Xia, Y.; Rao, J.; Yao, A.; Zhang, F.; Li, G.; Wang, X.; Lu, L. Lithium exacerbates hepatic ischemia/reperfusion injury by inhibiting GSK-3β/NF-κB-mediated protective signaling in mice. Eur. J. Pharmacol. 2012, 697, 117–125. [Google Scholar] [CrossRef]
- Christian, F.; Smith, E.L.; Carmody, R.J. The regulation of NF-κB subunits by phosphorylation. Cells 2016, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.; Vargas, J.; Hoffmann, A. Signaling via the NFκB system. Wiley Interdiscip. Rev. Syst. Biol. Med. 2016, 8, 227–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarai, S.K.; Mekala, H.M.; Lippmann, S. Lithium suicide prevention: A brief review and reminder. Innov. Clin. Neurosci. 2018, 15, 30–32. [Google Scholar]
- Scott, J.; Hidalgo-Mazzei, D.; Strawbridge, R.; Young, A.; Resche-Rigon, M.; Etain, B.; Andreassen, O.A.; Bauer, M.; Bennabi, D.; Blamire, A.M.; et al. Prospective cohort study of early biosignatures of response to lithium in bipolar-I-disorders: Overview of the H2020-funded R-LiNK initiative. Int. J. Bipolar Disord. 2019, 7, 20. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Tolliver, B.K.; Kemp, D.E.; Verduin, M.L.; Ganocy, S.J.; Bilali, S.; Brady, K.T.; Shim, S.S.; Findling, R.L.; Calabrese, J.R. Differential interactions between comorbid anxiety disorders and substance use disorder in rapid cycling bipolar I or II disorder. J. Affect. Disord. 2008, 110, 167–173. [Google Scholar]
- Sheehan, D.V. M.I.N.I. Mini International Neuropsychiatric Interview, Version 7.02, for DSM-5. 2016. Available online: https://harmresearch.org/index.php/mini-international-neuropsychiatric-interview-mini/#How%20to%20Cite%20the%20MINI (accessed on 1 December 2022).
- Spearing, M.K.; Post, R.M.; Leverich, G.S.; Brandt, D.; Nolen, W. Modification of the Clinical Global Impressions (CGI) scale for use in bipolar illness (BP): The CGI-BP. Psychiatry Res. 1997, 73, 159–171. [Google Scholar]
- Rush, A.; Trivedi, M.H.; Ibrahim, H.M.; Carmody, T.J.; Arnow, B.; Klein, D.N.; Markowitz, J.C.; Ninan, P.T.; Kornstein, S.; Manber, R.; et al. The 16-Item quick inventory of depressive symptomatology (QIDS), clinician rating (QIDS-C), and self-report (QIDS-SR): A psychometric evaluation in patients with chronic major depression. Biol. Psychiatry 2003, 54, 573–583. [Google Scholar]
- Linn, B.S.; Linn, M.W.; Gurel, L. Cumulative Illness Rating Scale. J. Am. Geriatr. Soc. 1968, 16, 622–626. [Google Scholar]
- Montgomery, S.A.; Åsberg, M. A New Depression Scale Designed to be Sensitive to Change. Br. J. Psychiatry 1979, 134, 382–389. [Google Scholar]
- Young, R.C.; Biggs, J.T.; Ziegler, V.E.; Meyer, D.A. A Rating Scale for Mania: Reliability, Validity and Sensitivity. Br. J. Psychiatry 1978, 133, 429–435. [Google Scholar]
- Guy, W. ECDEU Assessment Manuel for Psychopharmacology, Revised; U.S. Government Printing Office: Washington, DC, USA, 1976.
- Hamilton, M. The Assessment of Anxiety States by Rating. Psychol. Psychother. Theory Res. Pr. 1959, 32, 50–55. [Google Scholar]
- Snaith, R.P.; Hamilton, M.; Morley, S.; Humayan, A.; Hargreaves, D.; Trigwell, P. A Scale for the Assessment of Hedonic Tone the Snaith–Hamilton Pleasure Scale. Br. J. Psychiatry 1995, 167, 99–103. [Google Scholar]
- Leon, A.C.; Olfson, M.; Portera, L.; Farber, L.; Sheehan, D.V. Assessing Psychiatric Impairment in Primary Care with the Sheehan Disability Scale. Int. J. Psychiatry Med. 1997, 27, 93–105. [Google Scholar]
- Endicott, J.; Nee, J.; Harrison, W.; Blumenthal, R. Quality of Life Enjoyment and Satisfaction Questionnaire: A new measure. Psychopharmacol. Bull. 1993, 29, 321–326. [Google Scholar]
- Hartz, A.; Bentler, S.; Watson, D. Measuring fatigue severity in primary care patients. J. Psychosom. Res. 2003, 54, 515–521. [Google Scholar]
- Sobell, L.C.; Sobell, M.B. Timeline Followback: A technique for assessing self-reported alcohol consumption. In Measuring Alcohol Consumption: Psychosocial and Biological Methods; Litten, R.Z., Allen, J.P., Eds.; Humana Press: Totowa, NJ, USA, 1996; pp. 41–72. [Google Scholar]
- Wisniewski, S.R.; Rush, A.J.; Balasubramani, G.K.; Trivedi, M.H.; Nierenberg, A.A. STARD Investigators. Self-rated global measure of the frequency, intensity, and burden of side effects. J. Psychiatr. Pract. 2006, 12, 71–79. [Google Scholar]
- Posner, K.; Oquendo, M.A.; Gould, M.; Stanley, B.; Davies, M. Columbia Classification Algorithm of Suicide Assessment (C-CASA): Classification of Suicidal Events in the FDA’s Pediatric Suicidal Risk Analysis of Antidepressants. Am. J. Psychiatry 2007, 164, 1035–1043. [Google Scholar]

{kind=link}
{kind=link}
| Before Lithium | After Lithium | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Analytes | Intent-to-Treat-Responders (ITT-R) | Intent-to-Treat -Non-Responders (ITT-NR) | ITT-R vs. ITT-NR | Completed-Responders (C-R) | Completed-Non-Responders (C-NR) | C-R vs. C-NR | ||||
| n | Mean ± SD | n | Mean ± SD | p-value | n | Mean ± SD | n | Mean ± SD | p-Value | |
| BAK | 12 | 23.1 ± 5.2 | 11 | 25.3 ± 6.0 | 0.367 | 9 | 26.5 ± 3.0 | 4 | 19.6 ± 5.9 | 0.134 |
| BCL-2 | 11 | 21.4 ± 4.6 | 11 | 23.3 ± 9.0 | 0.551 | 9 | 23.5 ± 3.0 | 4 | 13.0 ± 8.4 | 0.117 |
| BCL-2 A1 | 12 | 1.4 ± 0.4 | 11 | 1.5 ± 0.3 | 0.380 | 9 | 1.5 ± 0.1 | 4 | 1.4 ± 0.2 | 0.298 |
| BDNF | 12 | 48.1 ± 12.5 | 10 | 54.6 ± 9.3 | 0.208 | 9 | 56.1 ± 8.7 | 4 | 36.2 ± 13.3 | 0.157 |
| Calmodulin | 12 | 46.4 ± 11.6 | 11 | 48.3 ± 10.3 | 0.712 | 9 | 52.2 ± 8.7 | 4 | 41.6 ± 17.5 | 0.377 |
| Fyn | 12 | 34.1 ± 7.5 | 11 | 38.4 ± 6.7 | 0.188 | 9 | 40.4 ± 3.9 | 4 | 24.3 ± 7.7 | 0.031 |
| GSK3β | 12 | 27.5 ± 10.7 | 10 | 29.8 ± 10.4 | 0.625 | 9 | 36.6 ± 5.4 | 4 | 24.8 ± 5.8 | 0.028 |
| HMGB1 | 11 | 26.2 ± 8.9 | 8 | 21.3 ± 5.8 | 0.240 | 8 | 27.0 ± 9.8 | 4 | 16.4 ± 6.1 | 0.070 |
| iNOS | 12 | 27.9 ± 12.9 | 11 | 27.6 ± 6.1 | 0.944 | 9 | 29.3 ± 6.5 | 4 | 17.2 ± 5.5 | 0.021 |
| IRS2 | 12 | 42.3 ± 13.7 | 11 | 46.2 ± 8.7 | 0.464 | 9 | 44.7 ± 10.8 | 4 | 38.7 ± 13.8 | 0.525 |
| MARCKS | 12 | 37.0 ± 8.8 | 11 | 40.2 ± 10.0 | 0.440 | 9 | 43.1 ± 6.5 | 4 | 35.4 ± 16.1 | 0.477 |
| mTor | 12 | 21.5 ± 10.7 | 11 | 24.9 ± 7.9 | 0.433 | 9 | 22.1 ± 4.4 | 4 | 20.2 ± 7.8 | 0.719 |
| NR3C1 | 8 | 7.0 ± 1.5 | 4 | 8.5 ± 2.6 | 0.228 | 9 | 39.7 ± 6.5 | 4 | 30.5 ± 14.6 | 0.358 |
| NLRP3 | 12 | 32.3 ± 10.7 | 10 | 41.2 ± 8.4 | 0.062 | 9 | 9.0 ± 1.5 | 4 | 4.6 ± 1.1 | 0.001 |
| PDEB4 | 10 | 22.4 ± 8.1 | 5 | 27.8 ± 9.8 | 0.308 | 9 | 26.5 ± 3.0 | 3 | 15.8 ± 10.4 | 0.172 |
| phospho-CREB | 12 | 46.4 ± 9.2 | 11 | 48.4 ± 8.2 | 0.596 | 9 | 46.2 ± 6.7 | 4 | 34.7 ± 11.1 | 0.168 |
| phospho-Fyn Yes | 12 | 23.9 ± 10.9 | 11 | 26.6 ± 6.7 | 0.517 | 9 | 52.8 ± 5.3 | 4 | 40.3 ± 15.5 | 0.256 |
| phospho-GSK3β | 12 | 4.7 ± 1.4 | 11 | 4.5 ± 1.1 | 0.815 | 9 | 27.6 ± 7.3 | 4 | 18.8 ± 4.5 | 0.041 |
| phospho-GSK3αβ | 12 | 3.0 ± 1.0 | 11 | 3.7 ± 1.0 | 0.146 | 9 | 6.6 ± 2.7 | 4 | 7.2 ± 4.6 | 0.826 |
| Phospho-NFkB-P65 | 12 | 8.5 ± 5.2 | 11 | 8.2 ± 2.9 | 0.861 | 9 | 3.2 ± 1.0 | 4 | 3.6 ± 1.0 | 0.552 |
| PGM1 | 12 | 39.8 ± 10.4 | 11 | 41.8 ± 8.0 | 0.628 | 9 | 10.9 ± 2.6 | 4 | 7.5 ± 1.2 | 0.014 |
| PKA C-α | 12 | 39.5 ± 11.01 | 11 | 43.0 ± 8.0 | 0.437 | 9 | 47.9 ± 6.7 | 4 | 29.1 ± 11.2 | 0.130 |
| PKC-θ | 9 | 5.2 ± 2.0 | 5 | 6.1 ± 3.0 | 0.566 | 9 | 4.4 ± 1.0 | 4 | 3.7 ± 1.6 | 0.486 |
| PPAR-γ | 12 | 39.2 ± 11.3 | 11 | 41.8 ± 7.0 | 0.548 | 9 | 24.5 ± 7.0 | 4 | 32.4 ± 10.7 | 0.102 |
| Timeless | 12 | 1.79 ± 0.48 | 11 | 1.8 ± 0.2 | 0.936 | 9 | 1.8 ± 0.2 | 4 | 1.4 ± 0.4 | 0.209 |
| TNFAIP3 | 12 | 24.71 ± 11.58 | 11 | 24.43 ± 10.78 | 0.9564 | 9 | 46.4 ± 6.1 | 4 | 14.8 ± 3.9 | 0.014 |
| TPH1 | 12 | 16.75 ± 5.65 | 11 | 15.29 ± 2.95 | 0.4897 | 9 | 19.1 ± 3.9 | 4 | 12.2 ± 3.4 | 0.028 |
| XBP1 | 12 | 1.21 ± 0.31 | 11 | 1.39 ± 0.48 | 0.3027 | 9 | 1.3 ± 0.2 | 4 | 1.2 ± 0.1 | 0.130 |
| Completed-Responders | Completed-Non-Responders | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Analytes | Before Lithium | After Lithium | Before vs. After Lithium | Before Lithium | After Lithium | Before vs. After Lithium | ||||
| n | Mean ± SD | n | Mean ± SD | p-Value | n | Mean ± SD | n | Mean ± SD | p-Value | |
| BAK | 9 | 23.2 ± 5.0 | 9 | 26.5 ± 3.2 | 0.118 | 4 | 20.5 ± 2.6 | 4 | 19.6 ± 6.8 | 0.826 |
| BCL-2 | 8 | 22.1 ± 5.0 | 8 | 22.8 ± 2.8 | 0.742 | 4 | 22.9 ± 5.4 | 4 | 13.0 ± 9.7 | 0.124 |
| BCL-2 A1 | 9 | 1.4 ± 0.3 | 9 | 1.5 ± 0.1 | 0.474 | 4 | 1.5 ± 0.2 | 4 | 1.4 ± 0.2 | 0.526 |
| BDNF | 9 | 50.1 ± 13.3 | 9 | 56.1 ± 9.2 | 0.277 | 3 | 44.3 ± 6.2 | 3 | 36.2 ± 16.3 | 0.461 |
| Calmodulin | 9 | 48.4 ± 12.8 | 9 | 52.2 ± 9.2 | 0.474 | 4 | 41.6 ± 10.6 | 4 | 41.6 ± 20.2 | 0.999 |
| Fyn | 9 | 34.4 ± 8.5 | 9 | 40.4 ± 4.1 | 0.074 | 4 | 35.3 ± 7.3 | 4 | 24.3 ± 8.9 | 0.106 |
| GSK3β | 9 | 29.8 ± 10.3 | 9 | 36.6 ± 5.8 | 0.103 | 4 | 29.4 ± 7.6 | 4 | 24.8 ± 6.7 | 0.393 |
| HMGB1 | 8 | 25.8 ± 9.6 | 8 | 27.0 ± 10.5 | 0.806 | 4 | 18.1 ± 3.2 | 4 | 16.4 ± 7.1 | 0.689 |
| iNOS | 9 | 26.5 ± 13.8 | 9 | 29.3 ± 6.9 | 0.598 | 4 | 24.0 ± 5.8 | 4 | 17.2 ± 6.4 | 0.164 |
| IRS2 | 9 | 43.7 ± 14.0 | 9 | 44.7 ± 11.4 | 0.860 | 4 | 45.7 ± 7.9 | 4 | 38.7 ± 16.0 | 0.456 |
| MARCKS | 9 | 37.9 ± 9.9 | 9 | 43.1 ± 6.9 | 0.210 | 4 | 27.9 ± 10.2 | 4 | 20.2 ± 9.0 | 0.300 |
| mTor | 9 | 23.0 ± 11.5 | 9 | 22.1 ± 4.7 | 0.821 | 4 | 37.1 ± 11.6 | 4 | 35.4 ± 18.6 | 0.885 |
| NLRP3 | 9 | 33.5 ± 11.0 | 9 | 39.7 ± 6.9 | 0.172 | 3 | 35.1 ± 7.9 | 3 | 23.6 ± 12.0 | 0.236 |
| NR3C1 | 8 | 7.0 ± 1.5 | 8 | 8.6 ± 1.2 | 0.029 | 4 | 8.4 ± 2.6 | 4 | 4.6 ± 1.2 | 0.037 |
| PDEB4 | 9 | 20.7 ± 6.4 | 9 | 26.5 ± 3.2 | 0.027 | 3 | 30.8 ± 9.4 | 3 | 21.1 ± 6.9 | 0.223 |
| PGM1 | 9 | 41.5 ± 11.2 | 9 | 46.2 ± 7.1 | 0.303 | 4 | 39.8 ± 7.8 | 4 | 34.7 ± 12.8 | 0.518 |
| phospho-CREB | 9 | 46.3 ± 9.7 | 9 | 52.8 ± 5.6 | 0.101 | 4 | 46.8 ± 6.9 | 4 | 40.3 ± 17.9 | 0.524 |
| phospho-Fyn/Yes | 9 | 24.7 ± 10.5 | 9 | 27.6 ± 7.7 | 0.519 | 4 | 26.5 ± 5.6 | 4 | 18.8 ± 5.2 | 0.089 |
| phospho-GSK3β | 9 | 4.5 ± 1.4 | 9 | 6.6 ± 2.8 | 0.073 | 4 | 5.1 ± 1.2 | 4 | 7.2 ± 5.3 | 0.458 |
| phospho-GSK3αβ | 9 | 3.1 ± 1.1 | 9 | 3.2 ± 1.1 | 0.987 | 4 | 4.4 ± 1.2 | 4 | 3.6 ± 1.1 | 0.373 |
| phospho-NFkB-P65 | 9 | 9.7 ± 5.4 | 9 | 10.9 ± 2.8 | 0.590 | 4 | 8.7 ± 3.2 | 4 | 7.5 ± 1.4 | 0.510 |
| PKA C-α | 9 | 42.3 ± 10.9 | 9 | 47.9 ± 7.1 | 0.213 | 4 | 38.8 ± 3.2 | 4 | 29.1 ± 13.7 | 0.296 |
| PKC-θ | 9 | 5.2 ± 2.0 | 9 | 4.4 ± 1.0 | 0.322 | 4 | 6.0 ± 3.0 | 4 | 3.7 ± 1.9 | 0.229 |
| PPAR-γ | 9 | 40.3 ± 11.5 | 9 | 46.4 ± 7.4 | 0.206 | 4 | 39.9 ± 9.0 | 4 | 32.4 ± 12.3 | 0.364 |
| Timeless | 9 | 1.8 ± 0.5 | 9 | 1.8 ± 0.3 | 0.887 | 4 | 1.8 ± 0.1 | 4 | 1.4 ± 0.5 | 0.253 |
| TNFAIP3 | 9 | 22.8 ± 9.6 | 9 | 24.5 ± 6.5 | 0.659 | 4 | 19.5 ± 2.8 | 4 | 14.8 ± 4.5 | 0.128 |
| TPH1 | 9 | 16.1 ± 5.9 | 9 | 19.1 ± 4.2 | 0.230 | 4 | 16.1 ± 4.4 | 4 | 12.2 ± 3.9 | 0.240 |
| XBP1 | 9 | 1.3 ± 0.3 | 9 | 1.3 ± 0.2 | 0.572 | 4 | 1.6 ± 0.7 | 4 | 1.2 ± 0.2 | 0.218 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, K.; Kaye, N.M.; Ayati, M.; Koyuturk, M.; Calabrese, J.R.; Christian, E.; Lazarus, H.M.; Kaplan, D. Divergent Directionality of Immune Cell-Specific Protein Expression between Bipolar Lithium Responders and Non-Responders Revealed by Enhanced Flow Cytometry. Medicina 2023, 59, 120. https://doi.org/10.3390/medicina59010120
Gao K, Kaye NM, Ayati M, Koyuturk M, Calabrese JR, Christian E, Lazarus HM, Kaplan D. Divergent Directionality of Immune Cell-Specific Protein Expression between Bipolar Lithium Responders and Non-Responders Revealed by Enhanced Flow Cytometry. Medicina. 2023; 59(1):120. https://doi.org/10.3390/medicina59010120
Chicago/Turabian StyleGao, Keming, Nicholas M. Kaye, Marzieh Ayati, Mehmet Koyuturk, Joseph R. Calabrese, Eric Christian, Hillard M. Lazarus, and David Kaplan. 2023. "Divergent Directionality of Immune Cell-Specific Protein Expression between Bipolar Lithium Responders and Non-Responders Revealed by Enhanced Flow Cytometry" Medicina 59, no. 1: 120. https://doi.org/10.3390/medicina59010120