
Improved Productivity of Astaxanthin from Photosensitive Haematococcus pluvialis Using Phototaxis Technology


 ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
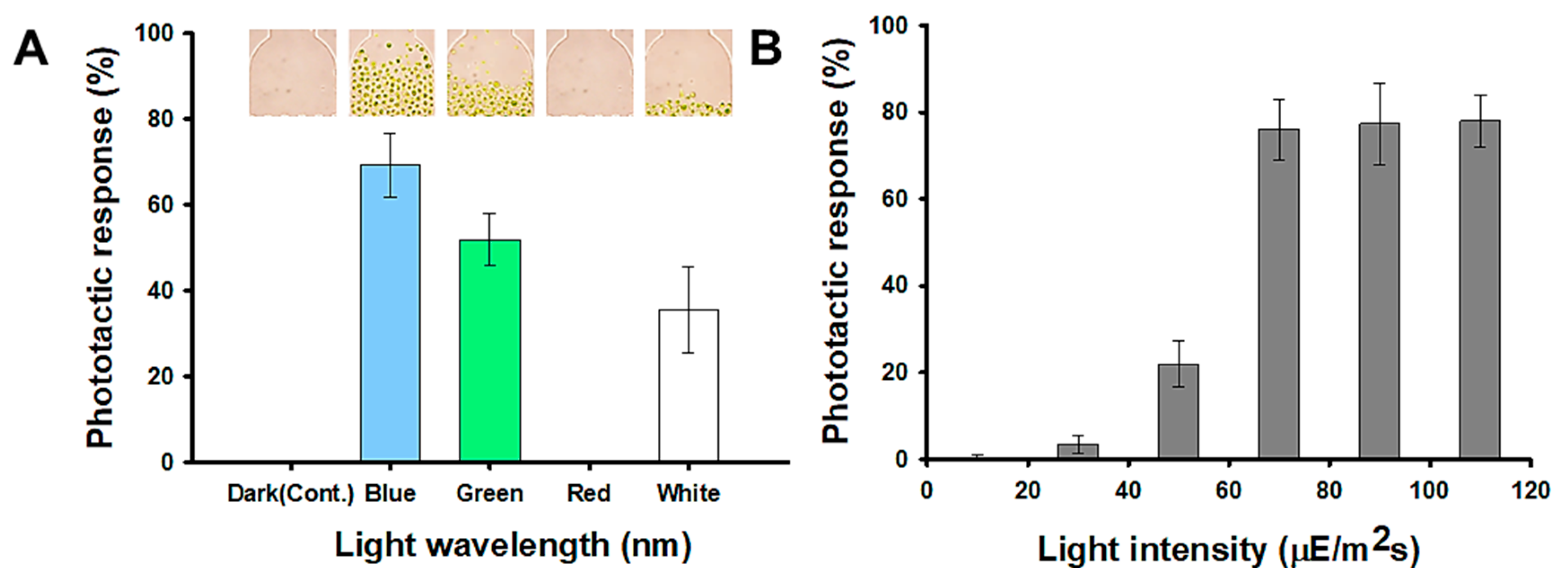
2.1. Investigation of Phototactic Properties of H. pluvialis
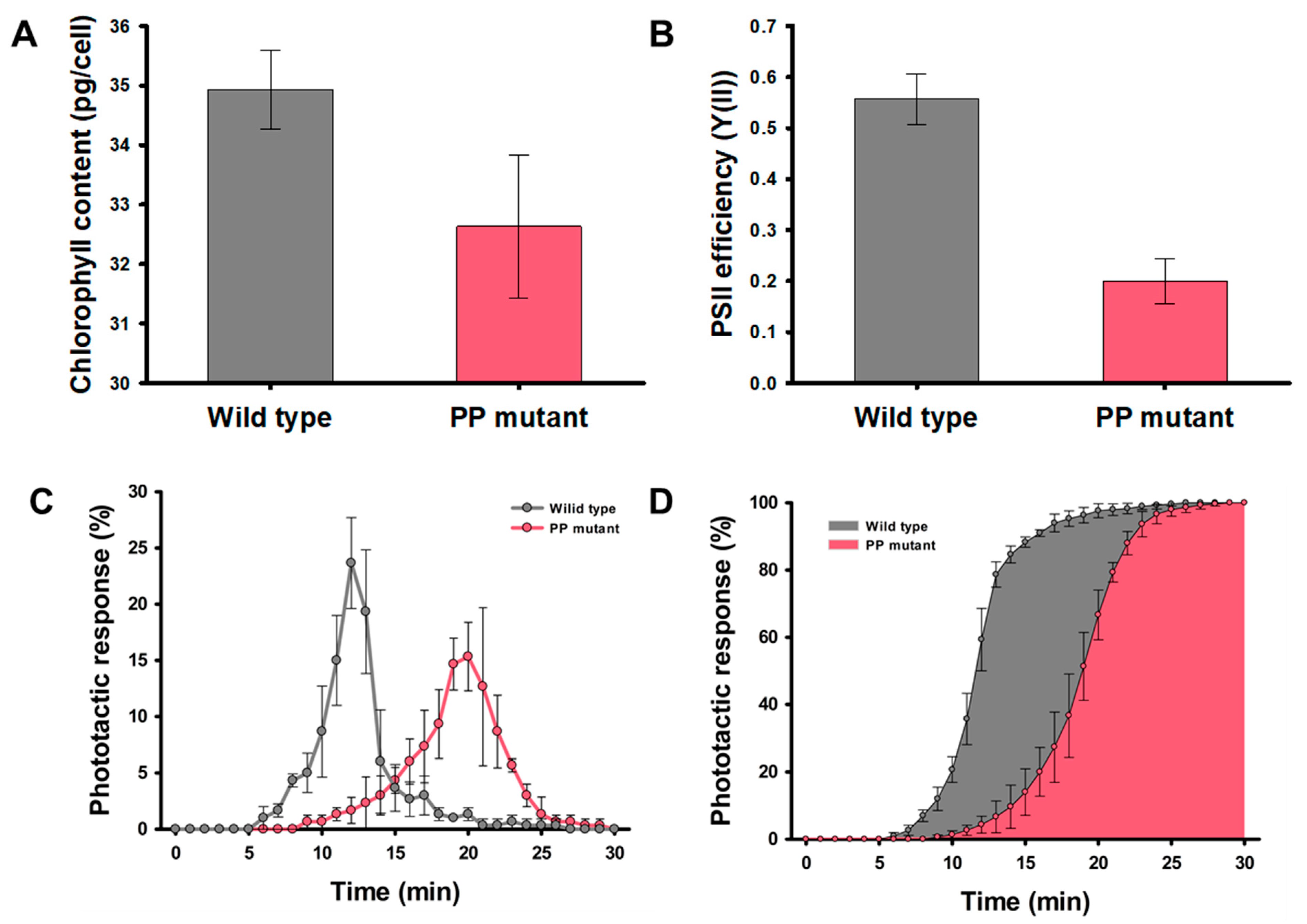
2.2. Correlation between Photosynthesis and Phototaxis
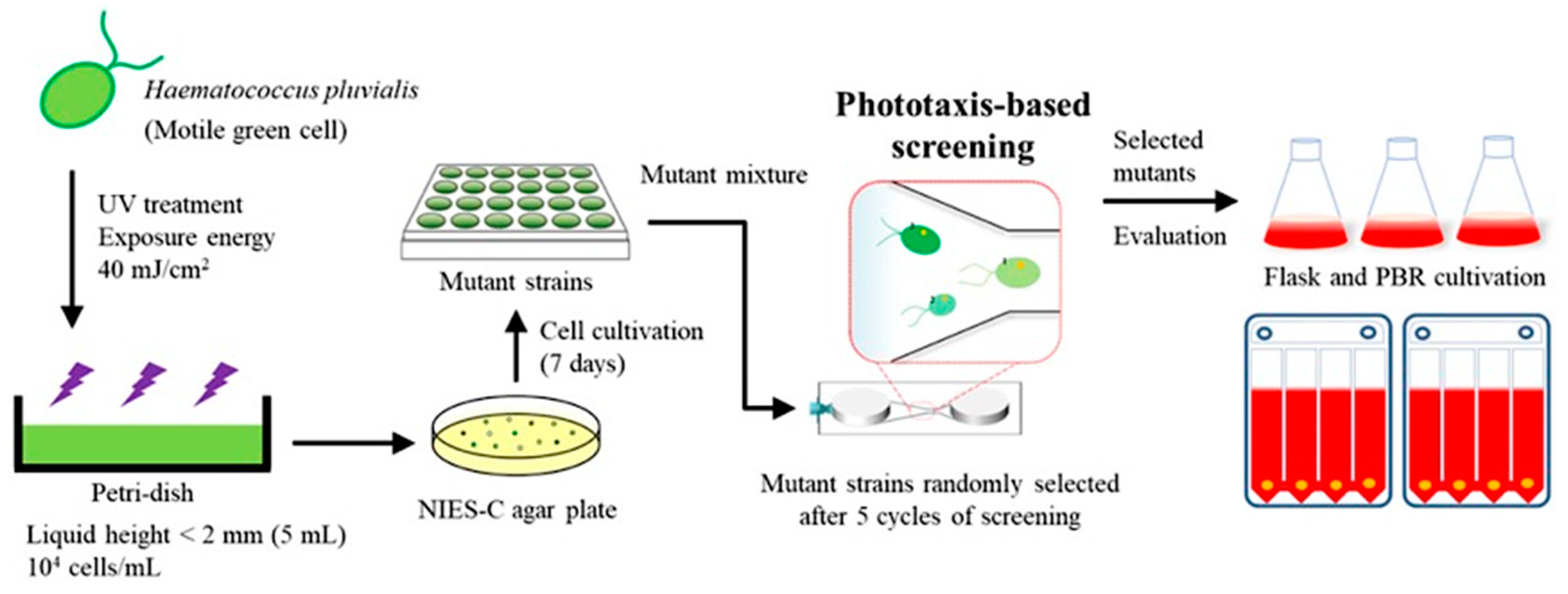
2.3. Strain Selection Using the Microfluidic Device and Flask Cultivation
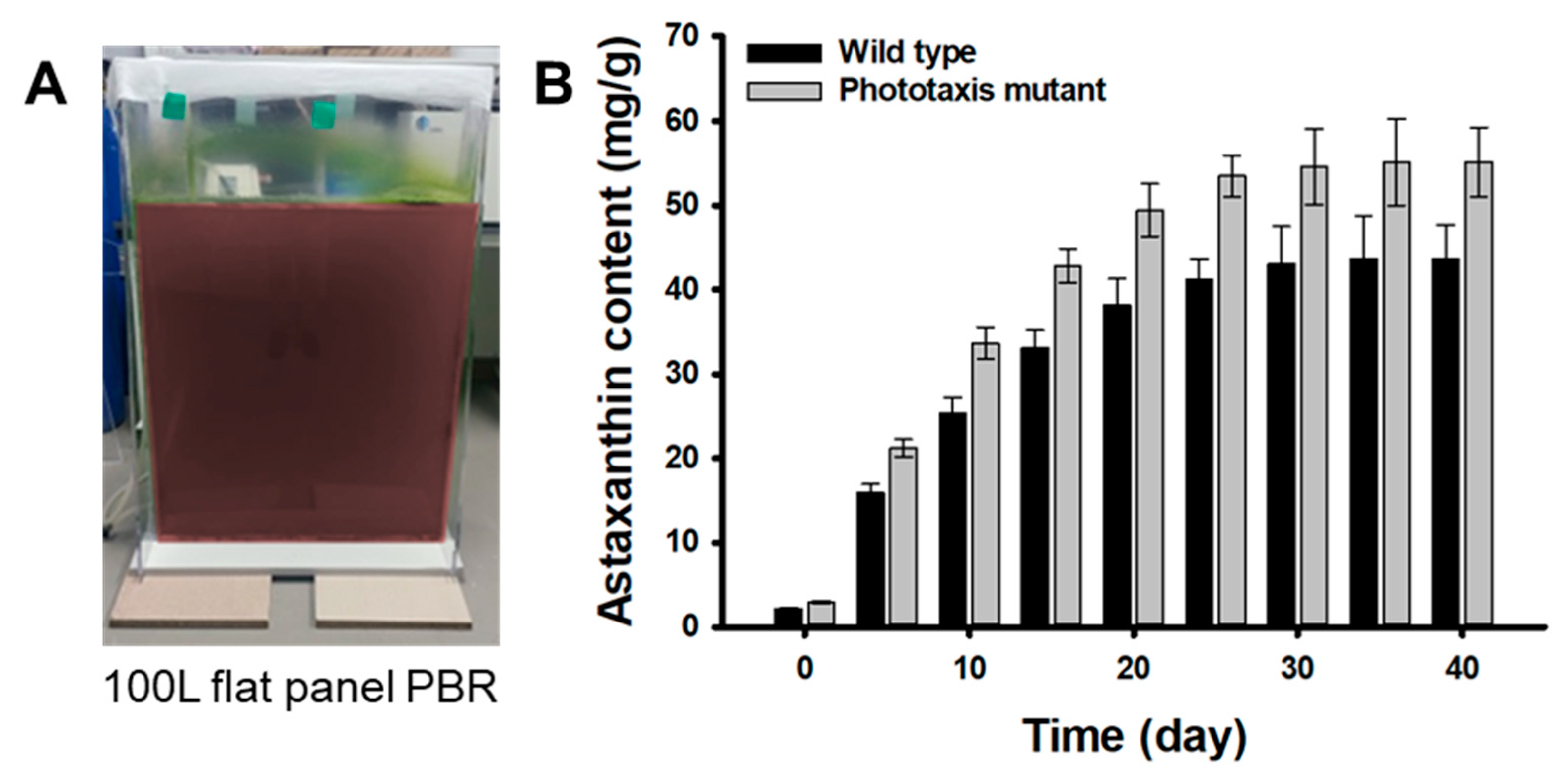
2.4. Validation of the M1 Mutant Performance in Large Scale Cultivation
3. Materials and Methods
3.1. Algal Strains and Culture Conditions
3.2. UV Irradiation Procedure for Mutagenesis
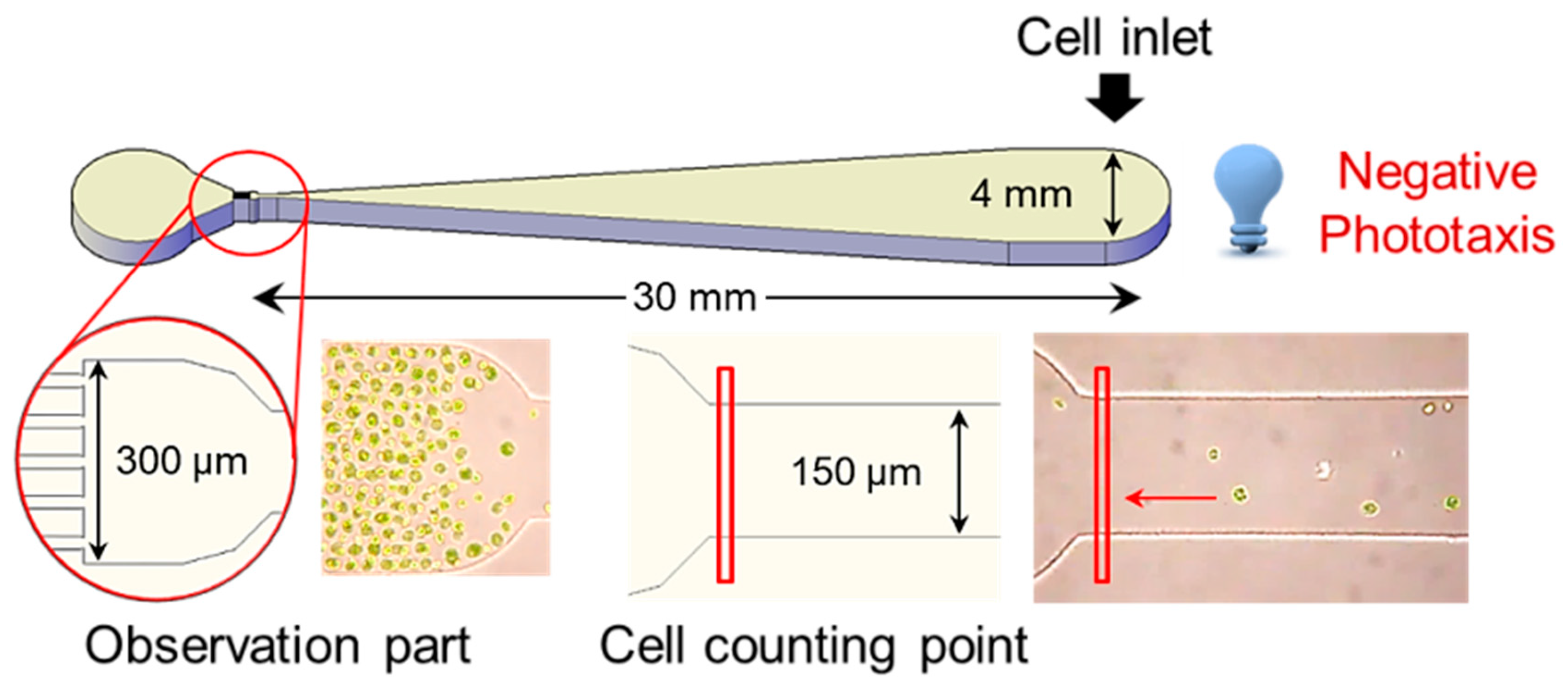
3.3. Fabrication of Microfluidic Device
3.4. Analysis of Phototactic Response in Microfluidic Device
3.5. Phototaxis-Based Screening
3.6. Analytical Methods
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, X.; Osawa, T. Cis astaxanthin and especially 9-cis astaxanthin exhibits a higher antioxidant activity in vitro compared to the all-trans isomer. Biochem. Biophys. Res. Commun. 2007, 357, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Ranga Rao, A.; Harshvardhan Reddy, A.; Aradhya, S.M. Antibacterial properties of Spirulina platensis, Haematococcus pluvialis, Botryococcus braunii microalgal extracts. Curr. Trends Biotechnol. Pharm. 2010, 4, 809–819. [Google Scholar]
- Pérez-López, P.; González-Garcia, S.; Jeffryes, C.; Agathos, S.N.; McHugh, E.; Walsh, D.; Murray, P.; Moane, S.; Feijoo, G.; Moreira, M.T. Life cycle assessment of the production of the red antioxidant carotenoid astaxanthin by microalgae: From lab to pilot scale. J. Clea. Prod. 2014, 64, 332–344. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.M.R.; Liang, Y.; Cheng, J.J.; Daroch, M. Astaxanthin-producing green microalga Haematococcus pluvialis: From single cell to high value commercial products. Front. Plant Sci. 2016, 7, 531. [Google Scholar] [PubMed] [Green Version]
- Panis, G.; Rosales Carreon, J. Commercial astaxanthin production derived by green alga Haematococcus pluvialis: A microalgae process model and a techno-economic assessment all through production line. Algal Res. 2016, 18, 175–190. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhu, D.L.; Niu, J.; Shen, S.D.; Wang, G. An economic assessment of astaxanthin production by large scale cultivation of Haematococcus pluvialis. Biotechnol. Adv. 2011, 29, 568–574. [Google Scholar] [CrossRef]
- Fassett, R.G.; Combes, J.S. Astaxanthin: A potential therapeutic agent in cardiovascular disease. Mar. Drugs 2011, 9, 447–465. [Google Scholar] [CrossRef] [Green Version]
- McWilliams, A. The Global Market for Carotenoids, BCC Research: Market Research Reports. 2018. FOD025F. Available online: https://www.bccresearch.com (accessed on 19 January 2021).
- Martínez-Álvarez, Ó.; Calvo, M.M.; Gómez-Estaca, J. Recent Advances in Astaxanthin Micro/Nanoencapsulation to Improve Its Stability and Functionality as a Food Ingredient. Mar. Drugs 2020, 18, 406. [Google Scholar] [CrossRef]
- Rammuni, M.N.; Ariyadasa, T.U.; Nimarshana, P.H.V.; Attalage, R.A. Comparative assessment on the extraction of carotenoids from microalgal sources: Astaxanthin from H. pluvialis and β-carotene from D. salina. Food Chem. 2019, 277, 128–134. [Google Scholar] [CrossRef]
- Khoo, K.S.; Lee, S.Y.; Ooi, C.W.; Fu, X.; Miao, X.; Ling, T.C.; Show, P.L. Recent advances in biorefinery of astaxanthin from Haematococcus pluvialis. Bioresour. Technol. 2019, 288, 121606. [Google Scholar] [CrossRef]
- Higuera-Ciapara, I.; Feliz-Valenzuela, L.; Goycoolea, F.M. Astaxanthin: A review of its chemistry and applications. Crit. Rev. Food Sci. Nutr. 2006, 46, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Ranga Rao, A.; Siew Moi, P.; Ravi, S.; Aswathanarayana, R.G. Astaxanthin: Sources, extraction, stability, biological activities and its commercial applications—A review. Mar. Drugs 2014, 12, 128–152. [Google Scholar]
- Koller, M.; Muhr, A.; Braunegg, G. Microalgae as versatile cellular factories for valued products. Algal Res. 2014, 6, 52–63. [Google Scholar] [CrossRef]
- Lu, Q.; Li, H.; Zou, Y.; Liu, H.; Yang, L. Astaxanthin as a microalgal metabolite for aquaculture: A review on the synthetic mechanisms, production techniques, and practical application. Algal Res. 2021, 54, 102178. [Google Scholar] [CrossRef]
- Pham, H.M.; Kwak, H.S.; Hong, M.E.; Lee, J.W.; Chang, W.S.; Sim, S.J. Development of an X-shape airlift photobioreactor for increasing algal biomass and biodiesel production. Bioresour. Technol. 2017, 239, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.C.; Ferreira, I.C.F.R.; Dias, M.M.; Barreiro, M.F. Microalgae-Derived Pigments: A 10-Year Bibliometric Review and Industry and Market Trend Analysis. Molecules 2020, 25, 3406. [Google Scholar] [CrossRef] [PubMed]
- Kwak, H.S.; Kim, J.Y.H.; Sim, S.J. A microreactor system for cultivation of Haematococcus pluvialis and astaxanthin production. J. Nanosci. Nanotechnol. 2015, 15, 1618–1623. [Google Scholar] [CrossRef]
- Boussiba, S.; Bing, W.; Yuan, J.P.; Zarka, A.; Chen, F. Changes in pigments profile in the green alga Haematococcus pluvialis exposed to environmental stresses. Biotechnol. Lett. 1999, 21, 601–604. [Google Scholar] [CrossRef]
- Wang, N.; Guan, B.; Kong, Q.; Sun, H.; Geng, Z.; Duan, L. Enhancement of astaxanthin production from Haematococcus pluvialis mutants by three-stage mutagenesis breeding. J. Biotechnol. 2016, 236, 71–77. [Google Scholar] [CrossRef]
- Pereira, S.; Otero, A. Haematococcus pluvialis bioprocess optimization: Effect of light quality, temperature and irradiance on growth, pigment content and photosynthetic response. Algal Res. 2020, 51, 102027. [Google Scholar] [CrossRef]
- Abomohra, A.E.F.; El-Sheekh, M.; Hanelt, D. Protoplast fusion and genetic recombination between Ochromonas danica (chrysophyta) and Haematococcus pluvialis (chlorophyta). Phycologia 2016, 55, 65–71. [Google Scholar] [CrossRef]
- Nasri, N.; Keyhanfar, M.; Behbahani, M.; Dini, G. Enhancement of astaxanthin production in Haematococcus pluvialis using zinc oxide nanoparticles. J. Biotechnol. 2021, 342, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, X.; Duan, C.; Yi, S.; Gao, Z.; Xiao, C.; Agathos, S.N.; Wang, G.; Li, J. Biotechnological production of astaxanthin from the microalga Haematococcus pluvialis. Biotechnol. Adv. 2020, 43, 107602. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Li, Y.; Wu, B.; Hu, W.; He, M.; Hu, G. Novel mutagenesis and screening technologies for food microorganisms: Advances and prospects. Appl. Microbiol. Biotechnol. 2020, 104, 1517–1531. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.E.; Choi, S.P.; Park, Y.I.; Kim, Y.K.; Chang, W.S.; Kim, B.W.; Sim, S.J. Astaxanthin production by a highly photosensitive Haematococcus mutant. Process Biochem. 2012, 47, 1972–1979. [Google Scholar] [CrossRef]
- Sandesh Kamath, B.; Vidhyavathi, R.; Sarada, R.; Ravishankar, G.A. Enhancement of carotenoids by mutation and stress induced carotenogenic genes in Haematococcus pluvialis mutants. Bioresour. Technol. 2008, 99, 8667–8673. [Google Scholar] [CrossRef]
- Hu, Z.; Li, Y.; Sommerfeld, M.; Chen, F.; Hu, Q. Enhanced protection against oxidative stress in an astaxanthin-overproduction Haematococcus mutant (Chlorophyceae). Eur. J. Phycol. 2008, 43, 365–376. [Google Scholar] [CrossRef]
- Sung, Y.J.; Kwak, H.S.; Hong, M.E.; Choi, H.I.; Sim, S.J. Two-dimensional microfluidic system for the simultaneous quantitative analysis of phototactic/chemotactic responses of microalgae. Anal. Chem. 2018, 90, 14029–14038. [Google Scholar] [CrossRef]
- Kim, J.Y.H.; Kwak, H.S.; Sung, Y.J.; Choi, H.I.; Hong, M.E.; Lim, H.S.; Lee, J.H.; Lee, S.Y.; Sim, S.J. Microfluidic high-throughput selection of microalgal strains with superior photosynthetic productivity using competitive phototaxis. Sci. Rep. 2016, 6, 21155. [Google Scholar] [CrossRef] [Green Version]
- Tamaki, S.; Tanno, Y.; Kato, S.; Ozasa, K.; Wakazaki, M.; Sato, M.; Toyooka, K.; Maoka, T.; Ishikawa, T.; Maeda, M.; et al. Carotenoid accumulation in the eyespot apparatus required for phototaxis is independent of chloroplast development in Euglena gracilis. Plant Sci. 2020, 298, 110564. [Google Scholar] [CrossRef]
- Ma, Z.E.; Helbling, E.W.; Li, W.; Villafañe, V.E.; Gao, K. Motility and photosynthetic responses of the green microalga Tetraselmis subcordiformis to visible and UV light levels. J. Appl. Phycol. 2012, 24, 1613–1621. [Google Scholar] [CrossRef]
- Choi, H.I.; Kim, J.Y.H.; Kwak, H.S.; Sung, Y.J.; Sim, S.J. Quantitative analysis of the chemotaxis of a green alga, Chlamydomonas reinhardtii, to bicarbonate using diffusion-based microfluidic device. Biomicrofluidics 2016, 10, 014121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ristori, T.; Rosati, G. The eyespot membranes of Haematococcus pluvialis flotow (Chlorophyceae): Their ultrastructure and possible significance in phototaxis. Ital. J. Zool. 1983, 17, 401–408. [Google Scholar]
- Tamaki, S.; Mochida, K.; Suzuki, K. Diverse Biosynthetic Pathways and Protective Functions against Environmental Stress of Antioxidants in Microalgae. Plants 2021, 10, 1250. [Google Scholar] [CrossRef]
- Ma, R.; Thomas-Hall, S.R.; Chua, E.T.; Eltanahy, E.; Netzel, M.E.; Netzel, G.; Lu, Y.; Schenk, P.M. Blue light enhances astaxanthin biosynthesis metabolism and extraction efficiency in Haematococcus pluvialis by inducing haematocyst germination. Algal Res. 2018, 35, 215–222. [Google Scholar] [CrossRef]
- Wan, M.X.; Zhang, Z.; Wang, J.; Huang, J.K.; Fan, J.H.; Yu, A.Q.; Wang, W.L.; Li, Y.G. Sequential Heterotrophy–Dilution–Photoinduction Cultivation of Haematococcus pluvialis for efficient production of astaxanthin. Bioresour. Technol. 2015, 198, 557–563. [Google Scholar] [CrossRef]
- Kwak, H.S.; Kim, J.Y.H.; Na, S.C.; Jeon, N.L.; Sim, S.J. Multiplex microfluidic system integrating sequential operations of microalgal lipid production. Analyst 2016, 141, 1218–1225. [Google Scholar] [CrossRef]
- Meyvantsson, I.; Beebe, D.J. Cell culture models in microfluidic systems. Annu. Rev. Anal. Chem. 2008, 1, 423–449. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Yue, C.; Ding, W.; Li, T.; Xu, J.-W.; Zhao, P.; Ma, H.; Yu, X. Butylated hydroxytoluene induces astaxanthin and lipid production in Haematococcus pluvialis under high-light and nitrogen-deficiency conditions. Bioresour. Technol. 2018, 266, 315–321. [Google Scholar] [CrossRef]
- Kobayashi, M.; Kakizono, T.; Nagai, S. Enhanced carotenoid biosynthesis by oxidative stress in acetate-induced cyst cells of a green unicellular alga, Haematococcus pluvialis. Appl. Environ. Microbiol. 1993, 59, 867–873. [Google Scholar] [CrossRef] [Green Version]
- He, Q.; Yang, H.; Wu, L.; Hu, C. Effect of light intensity on physiological changes, carbon allocation and neutral lipid accumulation in oleaginous microalgae. Bioresour. Technol. 2015, 191, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Zuluaga, M.; Gueguen, V.; Letourneur, D.; Pavon-Djavid, G. Astaxanthin-antioxidant impact on excessive Reactive Oxygen Species generation induced by ischemia and reperfusion injury. Chem. Biol. Interact. 2018, 279, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, D.; Lu, W.; Xing, J.; Hui, B.; Han, Y. Screening and characterization of astaxanthin-hyperproducing mutants of Haematococcus pluvialis. Biotechnol. Lett 2003, 25, 527–529. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, J.; Zhang, X.; Lin, W. Strain H2-419-4 of Haematococcus pluvialis induced by ethyl methanesulphonate and ultraviolet radiation. Chin. J. Oceanol. Limnol. 2008, 26, 152–156. [Google Scholar] [CrossRef]
- Hong, M.E.; Choi, H.; Kwak, S.; Hwang, S.W.; Sung, J.Y.; Chang, S.; Sim, S.J. Rapid selection of astaxanthin-hyperproducing Haematococcus mutant via azide-based colorimetric assay combined with oil-based astaxanthin extraction. Bioresour. Technol. 2018, 267, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Li, K.; Yang, Z.; Zhou, J.; Cen, K. Enhancing the growth rate and astaxanthin yield of Haematococcus pluvialis by nuclear irradiation and high concentration of carbon dioxide stress. Bioresour. Technol. 2016, 204, 49–54. [Google Scholar] [CrossRef]
- Liu, J.; Chen, J.; Chen, Z.; Qin, S.; Huang, Q. Isolation and characterization of astaxanthin-hyperproducing mutants of Haematococcus pluvialis (Chlorophyceae) produced by dielectric barrier discharge plasma. Phycologia 2016, 55, 650–658. [Google Scholar] [CrossRef]
- Gómez, P.I.; Inostroza, I.; Pizarro, M.; Pérez, J. From genetic improvement to commercial-scale mass culture of a chilean strain of the green microalga Haematococcus pluvialis with enhanced productivity of the red ketocarotenoid astaxanthin. AoB Plants 2013, 5, plt026. [Google Scholar] [CrossRef] [Green Version]
- Kang, C.D.; Lee, J.S.; Park, T.H.; Sim, S.J. Comparison of heterotrophic and photoautotrophic induction on astaxanthin production by Haematococcus pluvialis. Appl. Microbiol. Biotechnol. 2005, 68, 237–241. [Google Scholar] [CrossRef]
- Bišová, K.; Zachleder, V. Cell-cycle regulation in green algae dividing by multiple fission. J. Exp. Bot. 2014, 65, 2585–2602. [Google Scholar] [CrossRef]
- Qin, D.; Xia, Y.; Whitesides, G.M. Soft lithography for micro- and nanoscale patterning. Nat. Protoc. 2010, 5, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirst, H.; Garcia-Cerdan, J.G.; Zurbriggen, A.; Melis, A. Assembly of the light-harvesting chlorophyll antenna in the green alga Chlamydomonas reinhardtii requires expression of the TLA2-CpFTSY gene. Plant Physiol. 2012, 158, 930–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baroli, I.; Do, A.D.; Yamane, T.; Niyogi, K.K. Zeaxanthin accumulation in the absence of a functional xanthophyll cycle protects Chlamydomonas reinhardtii from photooxidative stress. Plant Cell 2003, 15, 992–1008. [Google Scholar] [CrossRef] [Green Version]
- Lamb, M.R.; Dutcher, S.K.; Worley, C.K.; Dieckmann, C.L. Eyespot-assembly mutants in Chlamydomonas reinhardtii. Genetics 1999, 153, 721–729. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Mutagenesis Strategy | Isolation Strategy | Astaxanthin Production | Growth Rate Improvement | Ref | |
|---|---|---|---|---|---|---|
| Unit | Improvement | |||||
| DPA12–1 | UV, EMS 1 | DPA 2 | 47.2 mg g−1 | 1.7-fold | 1.4-fold | [20] |
| EU3 | UV, EMS | Nicotine | 25.0 mg g−1 | 1.3-fold | Similar | [44] |
| H2–419–4 | UV, EMS | 37 pg cell−1 | 1.3-fold | 1.7-fold | [45] | |
| M13 | UV | Azide | 174.7 mg L−1 | 1.6-fold | Similar | [46] |
| Not specified | γ–ray | 70.8 mg g−1 | 2.4-fold | 1.2-fold | [47] | |
| M3 | DBD 3 plasma | DPA | 33.5 mg g−1 | 1.5-fold | 1.6-fold | [48] |
| B24 | EMS | DPA | 26.4 mg g−1 | 1.3-fold | 1.6-fold | [49] |
| M1 | UV | Phototaxis | 55.1 mg g−1 | 1.3-fold | 1.2-fold | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, K.H.; Chun, Y.; Lee, J.H.; Park, C.; Yoo, H.Y.; Kwak, H.S. Improved Productivity of Astaxanthin from Photosensitive Haematococcus pluvialis Using Phototaxis Technology. Mar. Drugs 2022, 20, 220. https://doi.org/10.3390/md20040220
Lee KH, Chun Y, Lee JH, Park C, Yoo HY, Kwak HS. Improved Productivity of Astaxanthin from Photosensitive Haematococcus pluvialis Using Phototaxis Technology. Marine Drugs. 2022; 20(4):220. https://doi.org/10.3390/md20040220
Chicago/Turabian StyleLee, Kang Hyun, Youngsang Chun, Ja Hyun Lee, Chulhwan Park, Hah Young Yoo, and Ho Seok Kwak. 2022. "Improved Productivity of Astaxanthin from Photosensitive Haematococcus pluvialis Using Phototaxis Technology" Marine Drugs 20, no. 4: 220. https://doi.org/10.3390/md20040220
APA StyleLee, K. H., Chun, Y., Lee, J. H., Park, C., Yoo, H. Y., & Kwak, H. S. (2022). Improved Productivity of Astaxanthin from Photosensitive Haematococcus pluvialis Using Phototaxis Technology. Marine Drugs, 20(4), 220. https://doi.org/10.3390/md20040220