
Moderate Exercise Combined with Enriched Environment Enhances Learning and Memory through BDNF/TrkB Signaling Pathway in Rats


Abstract
:1. Introduction
2. Experimental Procedures
2.1. Animals
2.2. Groups
2.2.1. Control Group
2.2.2. Treadmill Exercise
2.2.3. Enriched Environment
2.2.4. Treadmill Exercise Combined with Enriched Environment
2.3. Experimental Design
2.4. Morris Water Maze
2.5. ELISA for Plasma BDNF and TrkB
2.6. Statistical Analysis
3. Results
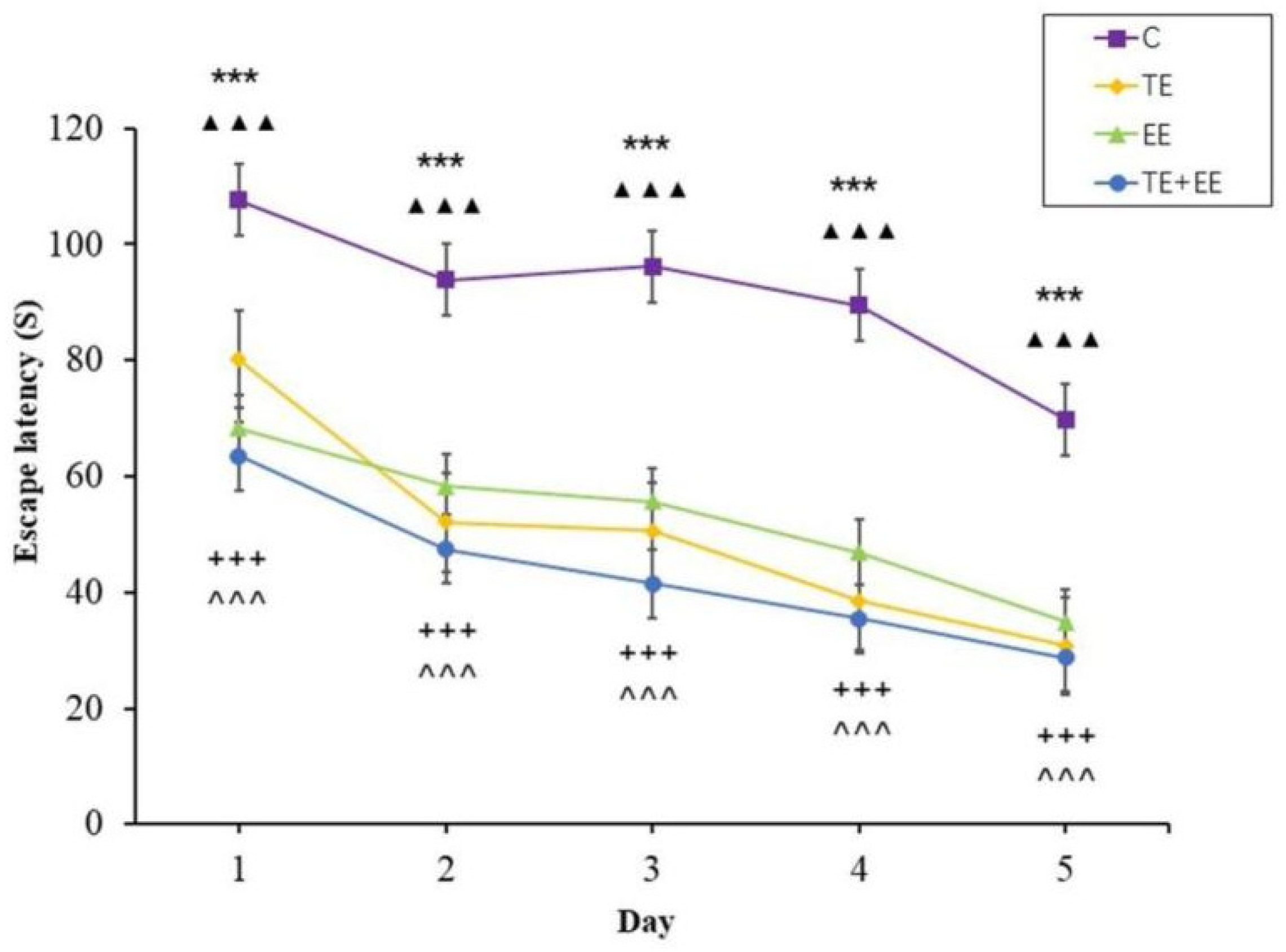
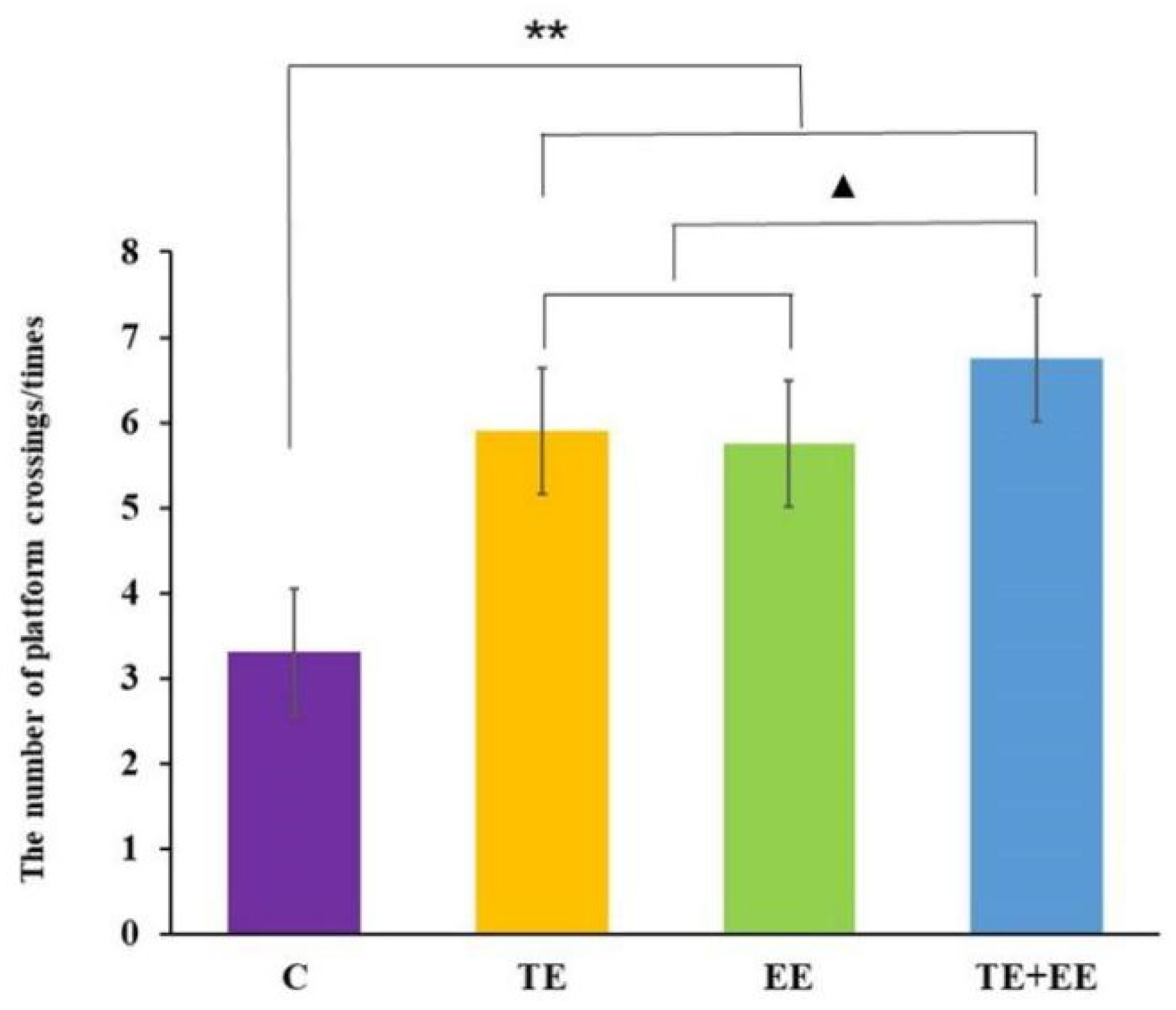
3.1. Behavioral Performance: The Morris Water Maze (MWM)
3.2. BDNF and TrkB in Plasma
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in neuronal development and function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef] [Green Version]
- McAllister, A.K.; Katz, L.C.; Lo, D.C. Neurotrophins and synaptic plasticity. Annu. Rev. Neurosci. 1999, 22, 295–318. [Google Scholar] [CrossRef] [Green Version]
- Sartorius, A.; Hellweg, R.; Litzke, J.; Vogt, M.; Dormann, C.; Vollmayr, B.; Danker-Hopfe, H.; Gass, P. Correlations and discrepancies between serum and brain tissue levels of neurotrophins after electroconvulsive treatment in rats. Pharmacopsychiatry 2009, 42, 270–276. [Google Scholar] [CrossRef]
- Karege, F.; Schwald, M.; Cisse, M. Postnatal developmental profile of brain-derived neurotrophic factor in rat brain and platelets. Neurosci. Lett. 2002, 328, 261–264. [Google Scholar] [CrossRef]
- Lang, U.E.; Hellweg, R.; Seifert, F.; Schubert, F.; Gallinat, J. Correlation between serum brain-derived neurotrophic factor level and an in vivo marker of cortical integrity. Biol. Psychiatry 2007, 62, 530–535. [Google Scholar] [CrossRef]
- Tyler, W.J. From acquisition to consolidation: On the role of brain-derived neurotrophic factor signaling in hippocampal-dependent learning. Learn. Mem. 2002, 9, 224–237. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.M.; Leem, Y.H. Chronic stress-induced memory deficits are reversed by regular exercise via ampk-mediated BDNF induction. Neuroscience 2016, 324, 271–285. [Google Scholar] [CrossRef]
- Sanz-García, A.; Knafo, S.; Pereda-Pérez, I.; Esteban, J.A.; Venero, C.; Armario, A. Administration of the TrkB receptor agonist 7,8-dihydroxyflavone prevents traumatic stress-induced spatial memory deficits and changes in synaptic plasticity. Hippocampus 2016, 26, 1179–1188. [Google Scholar] [CrossRef]
- Sabouri, M.; Kordi, M.; Shabkhiz, F.; Taghibeikzadehbadr, P.; Geramian, Z.S. Moderate treadmill exercise improves spatial learning and memory deficits possibly via changing PDE-5, IL-1 beta and pCREB expression. Exp. Gerontol. 2020, 139, 111056. [Google Scholar] [CrossRef] [PubMed]
- Erickson, K.I.; Voss, M.W.; Prakash, R.S.; Basak, C.; Szabo, A.; Chaddock, L.; Kim, J.S.; Heo, S.; Alves, H.; White, S.M.; et al. Exercise training increases size of hippocampus and improves memory. Proc. Natl. Acad. Sci. USA 2011, 108, 3017–3022. [Google Scholar] [CrossRef] [Green Version]
- van Dongen, E.V.; Kersten, I.H.P.; Wagner, I.C.; Morris, R.G.M.; Fernández, G. Physical exercise performed four hours after learning improves memory retention and increases hippocampal pattern similarity during retrieval. Curr. Biol. 2016, 26, 1722–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joubert, C.; Chainay, H. Aging brain: The effect of combined cognitive and physical training on cognition as compared to cognitive and physical training alone—A systematic review. Clin. Interv. Aging 2018, 13, 1267–1301. [Google Scholar] [CrossRef] [Green Version]
- Lago, T.R.; Hsiung, A.; Leitner, B.P.; Duckworth, C.J.; Balderston, N.L.; Chen, K.Y.; Grillon, C.; Ernst, M. Exercise modulates the interaction between cognition and anxiety in humans. Cogn. Emot. 2019, 33, 863–870. [Google Scholar] [CrossRef]
- Vaynman, S.; Ying, Z.; Gómez-Pinilla, F. Exercise induces BDNF and synapsin I to specific hippocampal subfields. J. Neurosci. Res. 2004, 76, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, A.J.; Tuon, T.; Pinho, C.A.; Silva, L.A.; Andreazza, A.C.; Kapczinski, F.; Quevedo, J.; Streck, E.L.; Pinho, R.A. Intense exercise induces mitochondrial dysfunction in mice brain. Neurochem. Res. 2008, 33, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Azimi, M.; Gharakhanlou, R.; Naghdi, N.; Khodadadi, D.; Heysieattalab, S. Moderate treadmill exercise ameliorates amyloid-beta-induced learning and memory impairment, possibly via increasing AMPK activity and up-regulation of the PGC-1alpha/FNDC5/BDNF pathway. Peptides 2018, 102, 78–88. [Google Scholar] [CrossRef]
- Tsai, S.F.; Ku, N.W.; Wang, T.F.; Yang, Y.H.; Shih, Y.H.; Wu, S.Y.; Lee, C.W.; Yu, M.; Yang, T.T.; Kuo, Y.M. Long-Term Moderate Exercise Rescues Age-Related Decline in Hippocampal Neuronal Complexity and Memory. Gerontology 2018, 64, 551–561. [Google Scholar] [CrossRef]
- Shafia, S.; Vafaei, A.A.; Samaei, S.A.; Bandegi, A.R.; Rafiei, A.; Valadan, R.; Hosseini-Khah, Z.; Mohammadkhani, R.; Rashidy-Pour, A. Effects of moderate treadmill exercise and fluoxetine on behavioural and cognitive deficits, hypothalamic-pituitary-adrenal axis dysfunction and alternations in hippocampal BDNF and mRNA expression of apoptosis-related proteins in a rat model of post-traumatic stress disorder. Neurobiol. Learn. Mem. 2017, 139, 165–178. [Google Scholar] [CrossRef]
- Just-Borras, L.; Hurtado, E.; Cilleros-Mane, V.; Biondi, O.; Charbonnier, F.; Tomas, M.; Garcia, N.; Tomas, J.; Lanuza, M.A. Running and swimming prevent the deregulation of the BDNF/TrkB neurotrophic signalling at the neuromuscular junction in mice with amyotrophic lateral sclerosis. Cell Mol. Life Sci. 2020, 77, 3027–3040. [Google Scholar] [CrossRef]
- Almeida, M.F.; Chaves, R.S.; Silva, C.M.; Chaves, J.C.S.; Melo, K.P.; Ferrari, M.F.R. BDNF trafficking and signaling impairment during early neurodegeneration is prevented by moderate physical activity. IBRO Rep. 2016, 1, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Rosenzweig, M.R.; Bennett, E.L.; Hebert, M.; Morimoto, H. Social grouping cannot account for cerebral effects of enriched environments. Brain Res. 1978, 153, 563–576. [Google Scholar] [CrossRef]
- Robison, L.S.; Francis, N.; Popescu, D.L.; Anderson, M.E.; Hatfield, J.; Xu, F.; Anderson, B.J.; Van Nostrand, W.E.; Robinson, J.K. Environmental enrichment: Disentangling the influence of novelty, social, and physical activity on cerebral amyloid angiopathy in a transgenic mouse model. Int. J. Mol. Sci. 2020, 21, 843. [Google Scholar] [CrossRef] [Green Version]
- Mora-Gallegos, A.; Rojas-Carvajal, M.; Salas, S.; Saborio-Arce, A.; Fornaguera-Trias, J.; Brenes, J.C. Age-dependent effects of environmental enrichment on spatial memory and neurochemistry. Neurobiol. Learn. Mem. 2015, 118, 96–104. [Google Scholar] [CrossRef]
- Stratta, P.; Sanità, P.; Bonanni, R.L.; de Cataldo, S.; Angelucci, A.; Rossi, R.; Origlia, N.; Domenici, L.; Carmassi, C.; Piccinni, A.; et al. Clinical correlates of plasma brain-derived neurotrophic factor in post-traumatic stress disorder spectrum after a natural disaster. Psychiatry Res. 2016, 244, 165–170. [Google Scholar] [CrossRef]
- Solinas, M.; Thiriet, N.; El, R.R.; Lardeux, V.; Jaber, M. Environmental enrichment during early stages of life reduces the behavioral, neurochemical, and molecular effects of cocaine. Neuropsychopharmacology 2009, 34, 1102–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austin, M.W.; Ploughman, M.; Glynn, L.; Corbett, D. Aerobic exercise effects on neuroprotection and brain repair following stroke: A systematic review and perspective. Neurosci. Res. 2014, 87, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Bondi, C.O.; Klitsch, K.C.; Leary, J.B.; Kline, A.E. Environmental enrichment as a viable neurorehabilitation strategy for experimental traumatic brain injury. J. Neurotraum. 2014, 31, 873–888. [Google Scholar] [CrossRef] [Green Version]
- Gajhede Gram, M.; Gade, L.; Wogensen, E.; Mogensen, J.; Malá, H. Equal effects of typical environmental and specific social enrichment on posttraumatic cognitive functioning after fimbria-fornix transection in rats. Brain Res. 2015, 1629, 182–195. [Google Scholar] [CrossRef]
- Ahmadalipour, A.; Ghodrati-Jaldbakhan, S.; Samaei, S.A.; Rashidy-Pour, A. Deleterious effects of prenatal exposure to morphine on the spatial learning and hippocampal BDNF and long-term potentiation in juvenile rats: Beneficial influences of postnatal treadmill exercise and enriched environment. Neurobiol. Learn. Mem. 2018, 147, 54–64. [Google Scholar] [CrossRef]
- Xie, H.; Wu, Y.; Jia, J.; Liu, G.; Zhang, Q.; Yu, K.; Guo, Z.; Shen, L.; Hu, R. Enrichment-induced exercise to quantify the effect of different housing conditions: A tool to standardize enriched environment protocols. Behav. Brain Res. 2013, 249, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Fabel, K.; Wolf, S.A.; Ehninger, D.; Babu, H.; Leal-Galicia, P.; Kempermann, G. Additive effects of physical exercise and environmental enrichment on adult hippocampal neurogenesis in mice. Front. Neurosci. 2009, 3, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapgal, V.; Prem, N.; Hegde, P.; Laxmi, T.R.; Kutty, B.M. Long term exposure to combination paradigm of environmental enrichment, physical exercise and diet reverses the spatial memory deficits and restores hippocampal neurogenesis in ventral subicular lesioned rats. Neurobiol. Learn. Mem. 2016, 130, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Bedford, T.G.; Tipton, C.M.; Wilson, N.C.; Oppliger, R.A.; Gisolfi, C.V. Maximum oxygen consumption of rats and its changes with various experimental procedures. J. Appl. Physiol. 1979, 47, 1278–1283. [Google Scholar] [CrossRef] [PubMed]
- Jing, L.; Zhongming, L.; Haijun, L.; Yan, Z.; Yongliang, W. Effects of different exercise loads on cognitive ability and expression of VEGF/VEGI in hippocampus of aged rats. Chin. J. Neuroanat. 2018, 34, 572–578. [Google Scholar]
- Frye, C.A. Estrus-associated decrements in a water maze task are limited to acquisition. Physiol. Behav. 1995, 57, 5–14. [Google Scholar] [CrossRef]
- Sakata, A.; Mogi, M.; Iwanami, J.; Tsukuda, K.; Min, L.J.; Jing, F.; Iwai, M.; Ito, M.; Horiuchi, M. Female exhibited severe cognitive impairment in type 2 diabetes mellitus mice. Life Sci. 2010, 86, 638–645. [Google Scholar] [CrossRef]
- Chen, W.Q.; Zhao, X.L.; Hou, Y.; Li, S.T.; Hong, Y.; Wang, D.L.; Cheng, Y.Y. Protective effects of green tea polyphenols on cognitive impairments induced by psychological stress in rats. Behav. Brain Res. 2009, 202, 71–76. [Google Scholar] [CrossRef]
- Pietrelli, A.; Matkovic, L.; Vacotto, M.; Lopez-Costa, J.J.; Basso, N.; Brusco, A. Aerobic exercise upregulates the BDNF-Serotonin systems and improves the cognitive function in rats. Neurobiol. Learn. Mem. 2018, 155, 528–542. [Google Scholar] [CrossRef] [PubMed]
- Pietrelli, A.; López-Costa, J.J.; Goñi, R.; López, E.M.; Brusco, A.; Basso, N. Effects of moderate and chronic exercise on the nitrergic system and behavioral parameters in rats. Brain Res. 2011, 1389, 71–82. [Google Scholar] [CrossRef]
- Wang, X.Q.; Wang, G.W. Effects of treadmill exercise intensity on spatial working memory and long-term memory in rats. Life Sci. 2016, 149, 96–103. [Google Scholar] [CrossRef]
- Okudan, N.; Belviranlı, M. Long-term voluntary exercise prevents post-weaning social isolation-induced cognitive impairment in rats. Neuroscience 2017, 360, 1–8. [Google Scholar] [CrossRef]
- Berchtold, N.C.; Castello, N.; Cotman, C.W. Exercise and time-dependent benefits to learning and memory. Neuroscience 2010, 167, 588–597. [Google Scholar] [CrossRef] [Green Version]
- Gradari, S.; Palle, A.; McGreevy, K.R.; Fontan-Lozano, A.; Trejo, J.L. Can Exercise Make You Smarter, Happier, and Have More Neurons? A Hormetic Perspective. Front. Neurosci. 2016, 10, 93. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Pinilla, F.; Ying, Z.; Roy, R.R.; Molteni, R.; Edgerton, V.R. Voluntary exercise induces a bdnf-mediated mechanism that promotes neuroplasticity. J. Neurophysiol. 2002, 88, 2187–2195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Pinilla, F.; Ying, Z.; Opazo, P.; Roy, R.R.; Edgerton, V.R. Differential regulation by exercise of BDNF and NT-3 in rat spinal cord and skeletal muscle. Eur. J. Neurosci. 2001, 13, 1078–1084. [Google Scholar] [CrossRef]
- Hauser, S.L.; Johnston, S.C. Of ghosts and sirens: The subtlest lures of industry. Ann. Neurol. 2007, 61, A11–A12. [Google Scholar] [CrossRef]
- Lou, S.J.; Liu, J.Y.; Chang, H.; Chen, P.J. Hippocampal neurogenesis and gene expression depend on exercise intensity in juvenile rats. Brain Res. 2008, 1210, 48–55. [Google Scholar] [CrossRef]
- Shih, P.; Yang, Y.; Wang, R. Effects of exercise intensity on spatial memory performance and hippocampal synaptic plasticity in transient brain ischemic rats. PLoS ONE 2013, 8, e78163. [Google Scholar] [CrossRef] [Green Version]
- Sheikhzadeh, F.; Etemad, A.; Khoshghadam, S.; Asl, N.A.; Zare, P. Hippocampal BDNF content in response to short- and long-term exercise. Neurol. Sci. 2015, 36, 1163–1166. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.S.; Ardais, A.P.; Fioreze, G.T.; Mioranzza, S.; Botton, P.H.S.; Souza, D.O.; Rocha, J.B.T.; Porciúncula, L.O. The impact of the frequency of moderate exercise on memory and brain-derived neurotrophic factor signaling in young adult and middle-aged rats. Neuroscience 2012, 222, 100–109. [Google Scholar] [CrossRef]
- Dalise, S.; Cavalli, L.; Ghuman, H.; Wahlberg, B.; Gerwig, M.; Chisari, C.; Ambrosio, F.; Modo, M. Biological effects of dosing aerobic exercise and neuromuscular electrical stimulation in rats. Sci. Rep. 2017, 7, 10830. [Google Scholar] [CrossRef]
- Li, X.; Wu, Q.; Xie, C.; Wang, C.; Wang, Q.; Dong, C.; Fang, L.; Ding, J.; Wang, T. Blocking of BDNF-TrkB signaling inhibits the promotion effect of neurological function recovery after treadmill training in rats with spinal cord injury. Spinal Cord 2019, 57, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Schapiro, S.J. Effects of social manipulations and environmental enrichment on behavior and cell-mediated immune responses in rhesus macaques. Pharm. Biochem. Behav. 2002, 73, 271–278. [Google Scholar] [CrossRef]
- Ambrogini, P.; Lattanzi, D.; Ciuffoli, S.; Betti, M.; Fanelli, M.; Cuppini, R. Physical exercise and environment exploration affect synaptogenesis in adult-generated neurons in the rat dentate gyrus: Possible role of BDNF. Brain Res. 2013, 1534, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, P.; Brassard, P.; Adser, H.; Pedersen, M.V.; Leick, L.; Hart, E.; Secher, N.H.; Pedersen, B.K.; Pilegaard, H. Evidence for a release of brain-derived neurotrophic factor from the brain during exercise. Exp. Physiol. 2009, 94, 1062–1069. [Google Scholar] [CrossRef] [PubMed]
- Branchi, I.; D Andrea, I.; Fiore, M.; Di Fausto, V.; Aloe, L.; Alleva, E. Early social enrichment shapes social behavior and nerve growth factor and brain-derived neurotrophic factor levels in the adult mouse brain. Biol. Psychiatry 2006, 60, 690–696. [Google Scholar] [CrossRef]
- Zhu, S.; Yee, B.K.; Nyffeler, M.; Winblad, B.; Feldon, J.; Mohammed, A.H. Influence of differential housing on emotional behaviour and neurotrophin levels in mice. Behav. Brain Res. 2006, 169, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Lu, B. BDNF and activity-dependent synaptic modulation. Learn. Mem. 2003, 10, 86–98. [Google Scholar] [CrossRef] [Green Version]
- Klein, A.B.; Williamson, R.; Santini, M.A.; Clemmensen, C.; Ettrup, A.; Rios, M.; Knudsen, G.M.; Aznar, S. Blood BDNF concentrations reflect brain-tissue BDNF levels across species. Int. J. Neuropsychopharmacol. 2011, 14, 347–353. [Google Scholar] [CrossRef] [Green Version]
- Pan, W.; Banks, W.A.; Fasold, M.B.; Bluth, J.; Kastin, A.J. Transport of brain-derived neurotrophic factor across the blood-brain barrier. Neuropharmacology 1998, 37, 1553–1561. [Google Scholar] [CrossRef]
- Bliss, T.V.; Collingridge, G.L. A synaptic model of memory: Long-term potentiation in the hippocampus. Nature 1993, 361, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.L. The Effects of Swimming Movement and Rich Environment on Autism Rats’ Learning and Memory Ability and Mechanism Research. Master’s Thesis, Jilin University, Changchun, China, 2016. [Google Scholar]
- Du, M.; Guan, Q.; Wang, X. Effects of Memantine combined with enriched environment on the expression of brain derived neurophic factor and tyrosine kinase receptor B in hippocampus and the learning and memory ability of senescence accelerated mouse prone 8. J. Clin. Neurol. 2013, 26, 278–281. [Google Scholar]
- Moradi-Kor, N.; Ghanbari, A.; Rashidipour, H.; Yousefi, B.; Bandegi, A.R.; Rashidy-Pour, A. Beneficial effects of Spirulina platensis, voluntary exercise and environmental enrichment against adolescent stress induced deficits in cognitive functions, hippocampal BDNF and morphological remolding in adult female rats. Horm. Behav. 2019, 112, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Warren, S.G.; Juraska, J.M. Spatial and nonspatial learning across the rat estrous cycle. Behav. Neurosci. 1997, 111, 259–266. [Google Scholar] [CrossRef]
- Galea, L.A.; Kavaliers, M.; Ossenkopp, K.P.; Hampson, E. Gonadal hormone levels and spatial learning performance in the Morris water maze in male and female meadow voles, Microtus pennsylvanicus. Horm. Behav. 1995, 29, 106–125. [Google Scholar] [CrossRef]
- Kennard, J.; Woodruff-Pak, D.S. Aging and exercise effects on motor learning and spatial memory. Ageing Res. 2011, 2, 2. [Google Scholar] [CrossRef] [Green Version]
- Qingwei, Y.; Bo, X.; Qing, T. Treadmill exercise improves hippocampal mitochondrial function and learning and memory ability of APP/PS1 mice. Chin. J. Biochem. Mol. Biol. 2019, 35, 669–677. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Group | N | BDNF | TrkB |
|---|---|---|---|
| C | 10 | 492.22 ± 77.03 | 1538.34 ± 133.86 |
| TE | 10 | 707.19 ± 63.63 **▲ | 2049.94 ± 122.38 **▲ |
| EE | 10 | 702.05 ± 106.30 **▲ | 1941.09 ± 239.68 **▲ |
| TE + EE | 10 | 790.49 ± 102.57 ** | 2213.22 ± 156.81 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, L.; Zhu, L.; Zhu, L.; Chen, D.; Cai, K.; Liu, Z.; Chen, A. Moderate Exercise Combined with Enriched Environment Enhances Learning and Memory through BDNF/TrkB Signaling Pathway in Rats. Int. J. Environ. Res. Public Health 2021, 18, 8283. https://doi.org/10.3390/ijerph18168283
Xu L, Zhu L, Zhu L, Chen D, Cai K, Liu Z, Chen A. Moderate Exercise Combined with Enriched Environment Enhances Learning and Memory through BDNF/TrkB Signaling Pathway in Rats. International Journal of Environmental Research and Public Health. 2021; 18(16):8283. https://doi.org/10.3390/ijerph18168283
Chicago/Turabian StyleXu, Liyuan, Linna Zhu, Lina Zhu, Dandan Chen, Kelong Cai, Zhimei Liu, and Aiguo Chen. 2021. "Moderate Exercise Combined with Enriched Environment Enhances Learning and Memory through BDNF/TrkB Signaling Pathway in Rats" International Journal of Environmental Research and Public Health 18, no. 16: 8283. https://doi.org/10.3390/ijerph18168283