
Synhelminthosporium gen. et sp. nov. and Two New Species of Helminthosporium (Massarinaceae, Pleosporales) from Sichuan Province, China

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Isolation, and Morphological Examination
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Phylogenetic Analyses
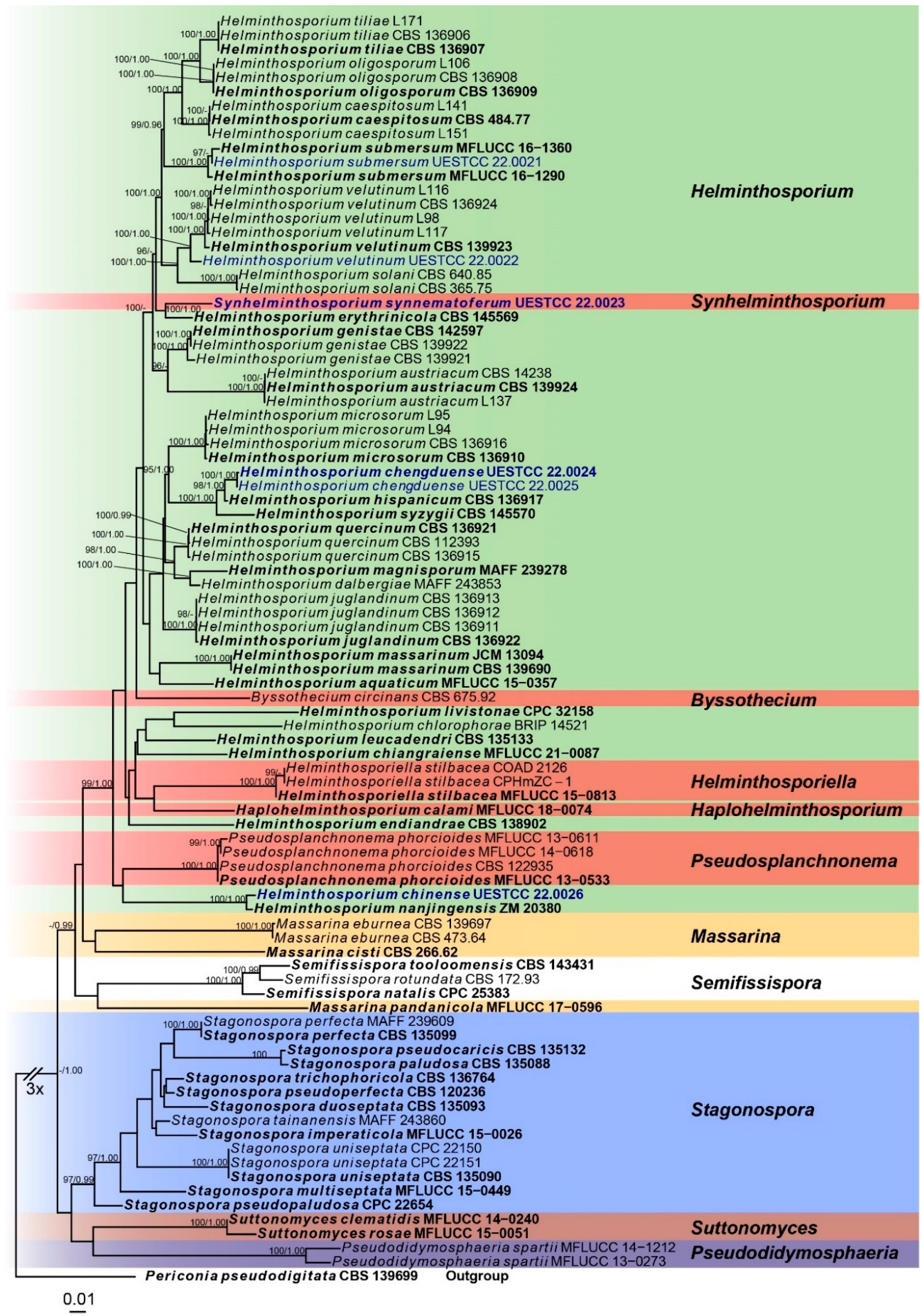
3. Results
3.1. Molecular Phylogeny
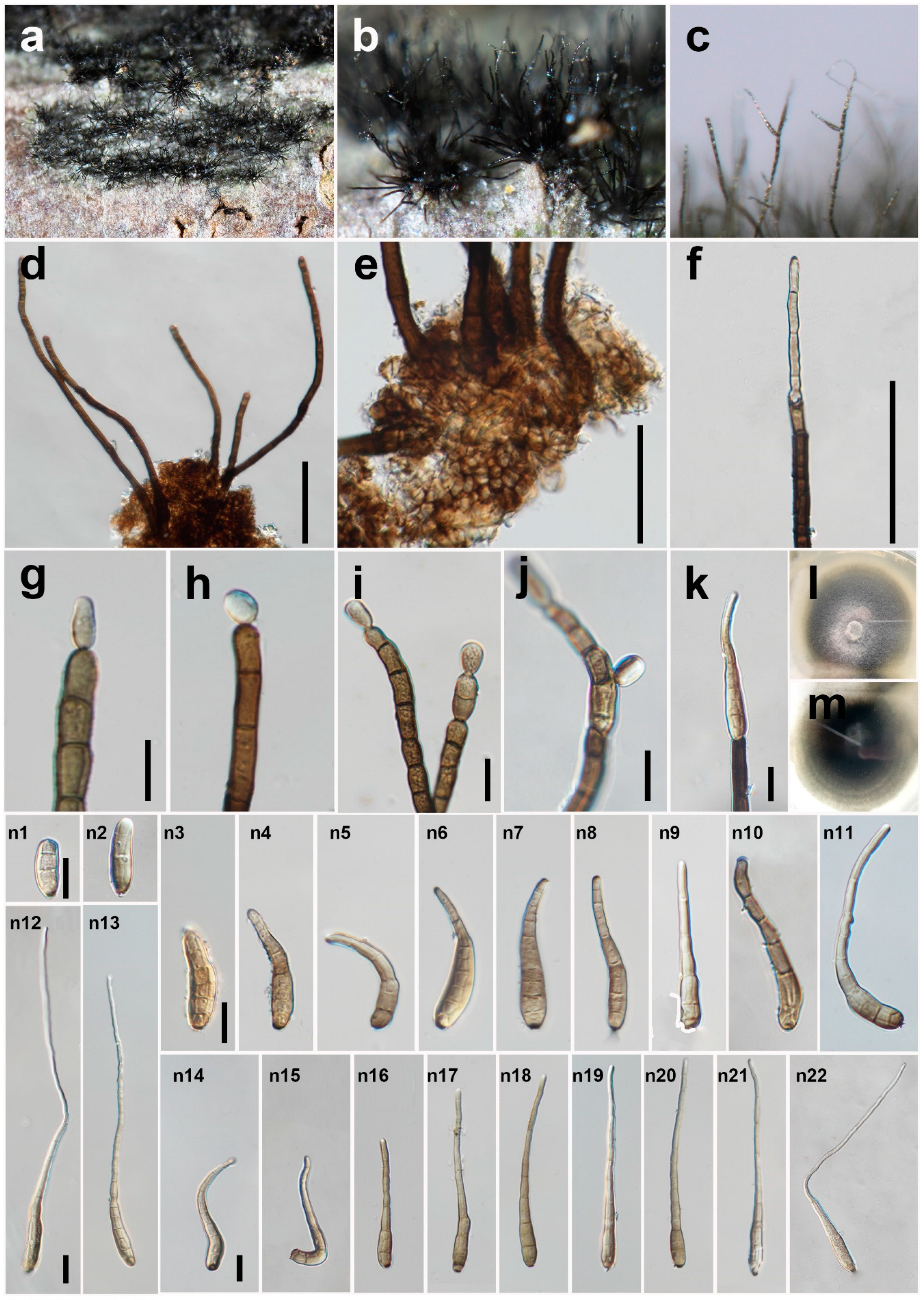
3.2. Taxonomy





4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hawksworth, D.L.; Lücking, R.; Heitman, J.; James, T.Y. Fungal Diversity Revisited: 2.2 to 3.8 Million Species. Microbiol. Spectr. 2017, 5, 79–95. [Google Scholar] [CrossRef] [PubMed]
- Celio, G.J.; Padamsee, M.; Dentinger, B.T.M.; Bauer, R.; McLaughlin, D.J. Assembling the Fungal Tree of Life: Constructing the Structural and Biochemical Database. Mycologia 2006, 98, 850–859. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.J.; McCluskey, K.; Verkleij, G.; Robert, V.; Smith, D. Fungal biological resources to support international development: Challenges and opportunities. World J. Microbiol. Biotechnol. 2019, 35, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Link, H.F. Observationes in Ordines plantarum naturales. Dissertatio prima complectens Anandrarum ordines Epiphytas, Mucedines, Gastromycos et Fungos. Ges. Natur 1809, 3, 3–42. [Google Scholar]
- Hongsanan, S.; Hyde, K.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Lücking, R.; Boonmee, S.; Bhat, J.D.; Liu, N.G.; et al. Refined families of Dothideomycetes: Orders and families incertae sedis in Dothideomycetes. Fungal Divers. 2020, 105, 17–318. [Google Scholar] [CrossRef]
- Wijayawardene, N.; Hyde, K.; Dai, D.; Sánchez-García, M.; Goto, B.; Saxena, R.; Erdogdu, M.; Selçuk, F.; Rajeshkumar, K.; Aptroot, A. Outline of Fungi and fungus-like taxa-2021. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- Konta, S.; Hyde, K.; Karunarathna, S.C.; Mapook, A.; Senwanna, C.; Dauner, L.A.P.; Nanayakkara, C.M.; Xu, J.; Tibpromma, S.; Lumyong, S. Multi-gene phylogeny and morphology reveal Haplohelminthosporium gen. nov. and Helminthosporiella gen. nov. associated with palms in Thailand and a checklist for Helminthosporium reported worldwide. Life 2021, 11, 454. [Google Scholar] [CrossRef]
- Voglmayr, H.; Jaklitsch, W.M. Corynespora, Exosporium and Helminthosporium revisited—New species and generic reclassification. Stud. Mycol. 2017, 87, 43–76. [Google Scholar] [CrossRef]
- Errampalli, D.; Saunders, J.M.; Holley, J.D. Emergence of silver scurf (Helminthosporium solani) as an economically important disease of potato. Plant Pathol. 2001, 50, 141–153. [Google Scholar] [CrossRef]
- Tian, S.M.; Chen, Y.C.; Zou, M.Q.; Xue, Q. First report of Helminthosporium solani causing silver scurf of potato in Hebei Province, North China. Plant Dis. 2007, 91, 460. [Google Scholar] [CrossRef]
- Tanaka, K.; Hirayama, K.; Yonezawa, H.; Sato, G.; Toriyabe, A.; Kudo, H.; Hashimoto, A.; Matsumura, M.; Harada, Y.; Kurihara, Y.; et al. Revision of the Massarineae (Pleosporales, Dothideomycetes). Stud. Mycol. 2015, 82, 75–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, D.; Luo, Z.L.; Baht, D.J.; Mckenzie, E.H.; Bahkali, A.H.; Zhou, D.Q.; Su, H.Y.; Hyde, K. Helminthosporium velutinum and H. aquaticum sp. nov. from aquatic habitats in Yunnan Province, China. Phytotaxa 2016, 253, 179–190. [Google Scholar] [CrossRef] [Green Version]
- Luttrell, E.S. Systematics of Helminthosporium and related genera. Mycologia 1964, 56, 119–132. [Google Scholar] [CrossRef]
- Subramanian, C.V.; Sekar, G. Three bitunicate ascomycetes and their tretic anamorphs. Kavaka 1987, 15, 87–97. [Google Scholar]
- Liang, Z.Q.; Wang, B.; Kang, J.C. Several rare entopathogenic fungi from the Western Sichuan mountains. Fungal Divers. 2003, 12, 129–134. [Google Scholar]
- Zhou, Y.; He, S.; Gong, G.; Zhang, S.; Chang, X.; Liu, N.; Sun, X.; Qi, X.; Ye, K.; Wang, Y. Soil fungal diversity in three nature reserves of Jiuzhaigou County, Sichuan Province, China. Ann. Microbiol. 2014, 64, 1275–1290. [Google Scholar] [CrossRef]
- Zhou, W.; Wei, Q.; Feng, R.; Liu, Y.; Liang, H.; Li, J.; Yan, K. Diversity and spatial distribution of endophytic fungi in Cinnamomum longepaniculatum of Yibin, China. Arch. Microbiol. 2021, 203, 3361–3372. [Google Scholar] [CrossRef]
- Yan, K.; Abbas, M.; Meng, L.; Cai, H.; Peng, Z.; Li, Q.; El-Sappah, A.H.; Yan, L.; Zhao, X. Analysis of the fungal diversity and community structure in Sichuan dark tea during pile-fermentation. Front. Microbiol. 2021, 12, 706714. [Google Scholar] [CrossRef]
- Toju, H.; Tanabe, A.S.; Yamamoto, S.; Sato, H. High-coverage ITS primers for the DNA-based identification of Ascomycetes and Basidiomycetes in environmental samples. PLoS ONE 2012, 7, e40863. [Google Scholar] [CrossRef] [Green Version]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Cubeta, M.A.; Echandi, E.; Abernethy, T.; Vilgalys, R. Characterization of anastomosis groups of binucleate Rhizoctonia species using restriction analysis of an amplified ribosomal RNA gene. Phytopathology 1991, 81, 1395–1400. [Google Scholar] [CrossRef]
- Hibbett, D.S. Phylogenetic evidence for horizontal transmission of group I introns in the nuclear ribosomal DNA of mushroom-forming fungi. Mol. Biol. Evol. 1996, 13, 903–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-alpha sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Jaklitsch, W.M.; Komon, M.; Kubicek, C.P.; Druzhinina, I.S. Hypocrea voglmayrii sp. nov. from the Austrian Alps represents a new phylogenetic clade in Hypocrea/Trichoderma. Mycologia 2005, 97, 1365–1378. [Google Scholar] [CrossRef]
- Voglmayr, H.; Akulov, O.Y.; Jaklitsch, W.M. Reassessment of Allantonectria, phylogenetic position of Thyronectroidea, and Thyronectria caraganae sp. nov. Mycol. Prog. 2016, 15, 921–937. [Google Scholar] [CrossRef] [Green Version]
- Sung, G.H.; Sung, J.M.; Hywel-Jones, N.L.; Spatafora, J.W. A multi-gene phylogeny of Clavicipitaceae (Ascomycota, Fungi): Identification of localized incongruence using a combinational bootstrap approach. Mol. Phylogenet. Evol. 2007, 44, 1204–1223. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of Phylogenetics and Evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.i.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2016, 34, 772–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2014, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [Green Version]
- Yu, G. Using ggtree to Visualize Data on Tree-Like Structures. Curr. Protoc. Bioinform. 2020, 69, e96. [Google Scholar] [CrossRef]
- Cooke, M.C. Ravenel’s American fungi. Grevillea 1878, 6, 129–146. [Google Scholar]
- Matsushima, T. Matsushima Mycological Memoirs No. 7; Matsushima Fungus: Kobe, Japan, 1993; Volume 7, 141p. [Google Scholar]
- Sydow, P.; Sydow, H. Weitere neue Micromyceten der Philippinen-Inseln. Ann. Mycol. 1920, 18, 98–104. [Google Scholar]
- Wang, X.J.; Wu, H.Y.; Zhang, M. A new species of Helminthosporium from Jiangsu, China. Mycotaxon 2014, 127, 1–4. [Google Scholar] [CrossRef]
- Zhao, N.; Luo, Z.L.; Hyde, K.D.; Su, H.Y.; Bhat, D.J.; Liu, J.K.; Bao, D.F.; Hao, Y.E. Helminthosporium submersum sp. nov. (Massarinaceae) from submerged wood in north-western Yunnan Province, China. Phytotaxa 2018, 348, 269–278. [Google Scholar] [CrossRef]
- Farr, D.F.; Rossman, A.Y. Fungal Databases; U.S. National Fungus Collections, USDA: Washingtin, DC, USA, 2022. [Google Scholar]
- Crous, P.W.; Wingfield, M.J.; Richardson, D.M.; Leroux, J.J.; Strasberg, D.; Edwards, J.; Roets, F.; Hubka, V.; Taylor, P.W.J.; Heykoop, M.; et al. Fungal Planet description sheets: 400–468. Persoonia 2016, 36, 316–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crous, P.W.; Carnegie, A.J.; Wingfield, M.J.; Sharma, R.; Mughini, G.; Noordeloos, M.E.; Santini, A.; Shouche, Y.S.; Bezerra, J.D.P.; Dima, B.; et al. Fungal Planet description sheets: 868–950. Persoonia 2019, 42, 291–473. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, R.A. Nomenclature of Drechslera and Bipolaris, grass parasites segregated from ‘Helminthosporium’. Can. J. Plant Pathol. 2006, 28, S212–S220. [Google Scholar] [CrossRef]
- Manamgoda, D.S.; Cai, L.; McKenzie, E.H.C.; Crous, P.W.; Madrid, H.; Chukeatirote, E.; Shivas, R.G.; Tan, Y.P.; Hyde, K. A phylogenetic and taxonomic re-evaluation of the Bipolaris-Cochliobolus-Curvularia complex. Fungal Divers. 2012, 56, 131–144. [Google Scholar] [CrossRef]
- Castañeda-Ruiz, R.F.; Li, D.W.; Zhang, X.G.; Kendrick, B.; Ramos-García, B.; Pérez-Martínez, S.; Sosa, D. Ellismarsporium gen. nov. and Stanhughesiella gen. nov. to accommodate atypical Helminthosporium and Corynesporella species. Mycotaxon 2018, 132, 759–766. [Google Scholar] [CrossRef]
- Cooke, M.C. Some Extra—European Fungi. Grevillea 1878, 7, 13–15. [Google Scholar]
- Thümen, V. Die Pilze des Weinstockes; W. Braumüller: Vienna, Austria, 1878. [Google Scholar]
- Gilman, J.C.; Abbott, E.V. A Summary of the Soil Fungi. Iowa State Coll. J. Sci. 1927, 1, 225–343. [Google Scholar]
- Curzi, M. Helminthosporium gibberosporum. C. R. Accad. Lincei. 1931, 6, 146. [Google Scholar]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Boonmee, S.; Wanasinghe, D.N.; Calabon, M.S.; Huanraluek, N.; Chandrasiri, S.K.U.; Jones, G.E.B.; Rossi, W.; Leonardi, M.; Singh, S.K.; Rana, S.; et al. Fungal diversity notes 1387–1511: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2021, 111, 1–335. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | PCR Primers | PCR: Thermal Cycles | Reference |
|---|---|---|---|
| ITS | ITS9mun/ITS4_KYO1 | (94 °C: 30 s, 56 °C: 30 s, 72 °C: 30 s) × 35 cycles | [19] |
| LSU | LR0R/LR5 | (94 °C: 30 s, 56 °C: 30 s, 72 °C: 1 min) × 35 cycles | [20,21] |
| SSU | PNS1/NS41 | (94 °C: 30 s, 56 °C: 30 s, 72 °C: 1 min) × 35 cycles | [22] |
| TEF1 | EF1-728F or EF1-983/EF1-2218R or TEF1LLErev | (94 °C: 30 s, 52 °C: 30 s, 72 °C: 1 min) × 35 cycles | [23,24,25] |
| RPB2 | dRPB2-5f or RPB2-5F2/dRPB2-7r or fRPB2-7cR | (94 °C: 30 s, 52 °C: 30 s, 72 °C: 1 min) × 35 cycles | [26,27,28] |
| Organism | Culture/Specimen No. 1 | SSU 2,3 | LSU | ITS | RPB2 | TEF1 |
|---|---|---|---|---|---|---|
| Byssothecium circinans | CBS 675.92 | GU205235 | GU205217 | OM337536 | DQ767646 | GU349061 |
| Haplohelminthosporium calami | MFLUCC 18-0074 HT | MT928160 | MT928156 | MT928158 | – 2 | – |
| Helminthosporiella stilbacea | COAD 2126 | – | – | MG668862 | – | MG682500 |
| H. stilbacea | MFLUCC 15-0813 HT | MT928161 | MT928157 | MT928159 | – | MT928151 |
| H. stilbacea | CPHmZC-01 | – | KX228355 | KX228298 | – | – |
| Helminthosporium aquaticum | MFLUCC 15-0357 = S-096 HT | KU697310 | KU697306 | KU697302 | – | – |
| H. austriacum | CBS 139924 = L132 HT | KY984420 | KY984301 | KY984301 | KY984365 | KY984437 |
| H. austriacum | CBS 14238 = L169 | – | KY984303 | KY984303 | KY984367 | KY984439 |
| H. austriacum | L137 | – | KY984302 | KY984302 | KY984366 | KY984438 |
| H. caespitosum | CBS 484.77 = L99 ET | KY984421 | JQ044448 | JQ044429 | KY984370 | KY984440 |
| H. caespitosum | L141 | – | KY984305 | KY984305 | KY984368 | – |
| H. caespitosum | L151 | – | KY984306 | KY984306 | KY984369 | – |
| H. chengduense | UESTC 22.0024 = YQ 071048 = CGMCC 3.23575 HT | ON557757 | ON557745 | ON557751 | ON563073 | ON600598 |
| H. chengduense | UESTC 22.0025 = YQ 071047 | ON557756 | ON557744 | ON557750 | ON563072 | ON600597 |
| H. chiangraiense | MFLUCC 21-0087 HT | – | MZ538538 | MZ538504 | – | – |
| H. chlorophorae | BRIP 14521 | – | – | AF120259 | – | – |
| H. dalbergiae | MAFF 243853 = H 4628 = TS 36 | AB797231 | AB807521 | LC014555 | – | AB808497 |
| H. endiandrae | CBS 138902 = CPC 22194 HT | – | KP004478 | KP004450 | – | – |
| H. erythrinicola | CPC 35291 = CBS 145569 HT | – | MK876432 | NR_165563 | MK876486 | – |
| H. genistae | CBS 142597 = L142 ET | – | KY984310 | KY984310 | KY984374 | – |
| H. genistae | CBS 139922 = L129 | KY984423 | KY984309 | KY984309 | KY984373 | – |
| H. genistae | CBS 139921 = L128 | KY984422 | KY984308 | KY984308 | KY984372 | – |
| H. hispanicum | CBS 136917 = L109 HT | KY984424 | KY984318 | KY984318 | KY984381 | KY984441 |
| H. juglandinum | CBS 136922 = L118 HT | – | KY984321 | KY984321 | KY984384 | KY984444 |
| H. juglandinum | CBS 136911 = L97 | KY984425 | KY984322 | KY984322 | KY984385 | KY984445 |
| H. juglandinum | CBS 136912 = L101 | – | KY984319 | KY984319 | KY984382 | KY984442 |
| H. juglandinum | CBS 136913 = L102 | – | KY984320 | KY984320 | KY984383 | KY984443 |
| H. leucadendri | CBS 135133 = CPC 19345 HT | – | KF251654 | KF251150 | KF252159 | KF253110 |
| H. livistonae | CPC 32158 = CBS 144413 HT | – | NG_064539 | NR_160348 | – | – |
| H. magnisporum | MAFF 239278 = H 4627 = TS 33 HT | AB797232 | AB807522 | AB811452 | – | AB808498 |
| H. massarinum | CBS 139690 = JCM 13095 = MAFF 239605 = KT 1564 HT | AB797234 | AB807524 | AB809629 | – | AB808500 |
| H. massarinum | JCM 13094 = MAFF 239604 = KT 838 EP | AB797233 | AB807523 | AB809628 | – | AB808499 |
| H. microsorum | CBS 136910 = L96 ET | KY984427 | KY984329 | KY984329 | KY984390 | KY984448 |
| H. microsorum | L94 | KY984426 | KY984327 | KY984327 | KY984388 | KY984446 |
| H. microsorum | CBS 136916 = L108 | – | KY984323 | KY984323 | KY984386 | – |
| H. microsorum | L95 | – | KY984328 | KY984328 | KY984389 | KY984447 |
| H. nanjingensis | HHAUF020380 = ZM020380 | – | – | KF192322 | – | – |
| H. oligosporum | CBS 136909 = L93 ET | – | KY984333 | KY984333 | KY984394 | KY984451 |
| H. oligosporum | CBS 136908 = L92 | KY984428 | KY984332 | KY984332 | KY984393 | KY984450 |
| H. oligosporum | L106 | – | KY984330 | KY984330 | KY984391 | KY984449 |
| H. quercinum | CBS 136921 = L90 HT | KY984429 | KY984339 | KY984339 | KY984400 | KY984453 |
| H. quercinum | CBS 112393 | – | KY984334 | KY984334 | KY984395 | KY984452 |
| H. quercinum | CBS 136915 = L107 | – | KY984336 | KY984336 | KY984397 | – |
| H. solani | CBS 365.75 | KY984430 | KY984341 | KY984341 | KY984402 | KY984455 |
| H. solani | CBS 640.85 | – | KY984342 | KY984342 | KY984403 | – |
| H. submersum | MFLUCC 16-1360 HT | MG098796 | MG098787 | – | – | MG098586 |
| H. submersum | MFLUCC 16-1290 PT | MG098797 | MG098788 | MG098780 | MG098592 | MG098587 |
| H. submersum | UESTCC 22.0021 = Sara 08_3 = CGMCC 3.23571 | ON557759 | ON557747 | ON557753 | ON563075 | ON600600 |
| H. syzygii | CPC 35312 = CBS 145570 HT | – | MK876433 | NR_165564 | MK876487 | – |
| H. tiliae | CBS 136907 = L88 ET | KY984431 | KY984345 | KY984345 | KY984406 | KY984457 |
| H. tiliae | CBS 136906 = L87 | – | KY984344 | KY984344 | KY984405 | – |
| H. tiliae | L171 | – | KY984343 | KY984343 | KY984404 | KY984456 |
| H. velutinum | CBS 139923 = L131 ET | KY984432 | KY984352 | KY984352 | KY984413 | KY984463 |
| H. velutinum | L98 | KY984433 | KY984359 | KY984359 | KY984417 | KY984466 |
| H. velutinum | CBS 136924 = L115 | – | KY984347 | KY984347 | KY984408 | KY984458 |
| H. velutinum | L116 | – | KY984348 | KY984348 | KY984409 | KY984459 |
| H. velutinum | L117 | – | KY984349 | KY984349 | KY984410 | KY984460 |
| H. velutinum | UESTCC 22.0022 = BY 14_2 = CGMCC 3.23572 | ON557761 | ON557749 | ON557755 | – | ON600602 |
| H. chinense | UESTCC 22.0026 = YQ 071,005 = CGMCC 3.23570 HT | ON557760 | ON557748 | ON557754 | – | ON600601 |
| Massarina cisti | CBS 266.62 = JCM 14140 HT | AB797249 | AB807539 | LC014568 | FJ795464 | AB808514 |
| M. eburnea | CBS 473.64 | GU296170 | GU301840 | OM337528 | GU371732 | GU349040 |
| M. eburnea | CBS 139697 = JCM 14422 = H 3953 | AB521718 | AB521735 | LC014569 | – | AB808517 |
| M. pandanicola | MFLUCC 17-0596 = KUMCC 17-0293 HT | MG646979 | MG646947 | MG646958 | – | MG646986 |
| Periconia pseudodigitata | KT 1395 = HHUF 29370 = CBS 139699 = JCM 13166 = MAFF 239676 HT | NG_064850 | NG_059396 | NR_153490 | – | AB808540 |
| Pseudodidymosphaeria spartii | MFLUCC 13-0273 | KP325438 | KP325436 | KP325434 | – | – |
| P. spartii | MFLUCC 14-1212 | KP325439 | KP325437 | KP325435 | – | – |
| Pseudosplanchnonema phorcioides | L16 = CBS 122935 | KY984434 | KY984360 | KY984360 | KY984418 | KY984467 |
| P. phorcioides | MFLUCC 13-0533 = CGMCC 3.17583 | KM875455 | KM875454 | – | – | – |
| P. phorcioides | MFLUCC 13-0611 | KP683377 | KP683376 | KP683375 | – | – |
| P. phorcioides | MFLUCC 14-0618 | KP683374 | KP683373 | KP683372 | – | – |
| Semifissispora natalis | CPC 25383 = CBS 140659 HT | – | KT950858 | KT950846 | – | KT950878 |
| S. rotundata | CBS 172.93 = CPC 549 | – | KT950859 | KT950847 | – | – |
| S. tooloomensis | CBS 143431 = CPC 31680 HT | – | NG_058526 | NR_156674 | – | – |
| Stagonospora duoseptata | CBS 135093 = S618 HT | – | KF251758 | KF251255 | KF252260 | KF253205 |
| S. imperaticola | MFLUCC 15-0026 = ICMP 21563 HT | KY706138 | KY706133 | KY706143 | KY706149 | KY706146 |
| S. multiseptata | MFLUCC 15-0449 = ICMP 21562 HT | – | NG_068239 | NR_165854 | – | – |
| S. paludosa | CBS 135088 = S601 NT | – | KF251760 | KF251257 | KF252262 | KF253207 |
| S. perfecta | KT 1726A = JCM 13099 = MAFF 239609 | AB797289 | AB807579 | AB809642 | – | AB808555 |
| S. perfecta | CBS 135099 = S656 HT | – | KF251761 | KF251258 | KF252263 | – |
| S. pseudocaricis | CBS 135132 = S610 HT | – | KF251763 | KF251259 | KF252265 | KF253210 |
| S. pseudopaludosa | CPC 22654 = CBS 136424 HT | – | NG_058052 | NR_137840 | – | – |
| S. pseudoperfecta | CBS 120236 = JCM 13097 = MAFF 239607 HT | AB797287 | AB807577 | AB809641 | – | AB808553 |
| S. tainanensis | KT 1866 = MAFF 243860 | AB797290 | AB807580 | AB809643 | – | AB808556 |
| S. trichophoricola | CBS 136764 = D652 HT | – | NG_058081 | NR_156586 | KJ869232 | – |
| S. uniseptata | CBS 135090 = S611 HT | – | KF251767 | KF251264 | KF252269 | – |
| S. uniseptata | S607 = CPC 22151 | – | KF251768 | KF251265 | KF252270 | KF253213 |
| S. uniseptata | S608 = CPC 22150 | – | KF251769 | KF251266 | KF252271 | KF253214 |
| Suttonomyces clematidis | MFLUCC 14-0240 = GUCC 18 | KP842920 | KP842917 | – | – | – |
| S. rosae | MFLUCC 15-0051 HT | MG829185 | MG829085 | MG828973 | – | – |
| Synhelminthosporium synnematoferum | UESTCC 22.0023 = HLG 072894 = CGMCC 3.23574 HT | ON557758 | ON557746 | ON557752 | ON563074 | ON600599 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Tian, W.; Guo, Y.; Madrid, H.; Maharachchikumbura, S.S.N. Synhelminthosporium gen. et sp. nov. and Two New Species of Helminthosporium (Massarinaceae, Pleosporales) from Sichuan Province, China. J. Fungi 2022, 8, 712. https://doi.org/10.3390/jof8070712
Chen Y, Tian W, Guo Y, Madrid H, Maharachchikumbura SSN. Synhelminthosporium gen. et sp. nov. and Two New Species of Helminthosporium (Massarinaceae, Pleosporales) from Sichuan Province, China. Journal of Fungi. 2022; 8(7):712. https://doi.org/10.3390/jof8070712
Chicago/Turabian StyleChen, Yanpeng, Wenhui Tian, Yaobin Guo, Hugo Madrid, and Sajeewa S. N. Maharachchikumbura. 2022. "Synhelminthosporium gen. et sp. nov. and Two New Species of Helminthosporium (Massarinaceae, Pleosporales) from Sichuan Province, China" Journal of Fungi 8, no. 7: 712. https://doi.org/10.3390/jof8070712
APA StyleChen, Y., Tian, W., Guo, Y., Madrid, H., & Maharachchikumbura, S. S. N. (2022). Synhelminthosporium gen. et sp. nov. and Two New Species of Helminthosporium (Massarinaceae, Pleosporales) from Sichuan Province, China. Journal of Fungi, 8(7), 712. https://doi.org/10.3390/jof8070712