
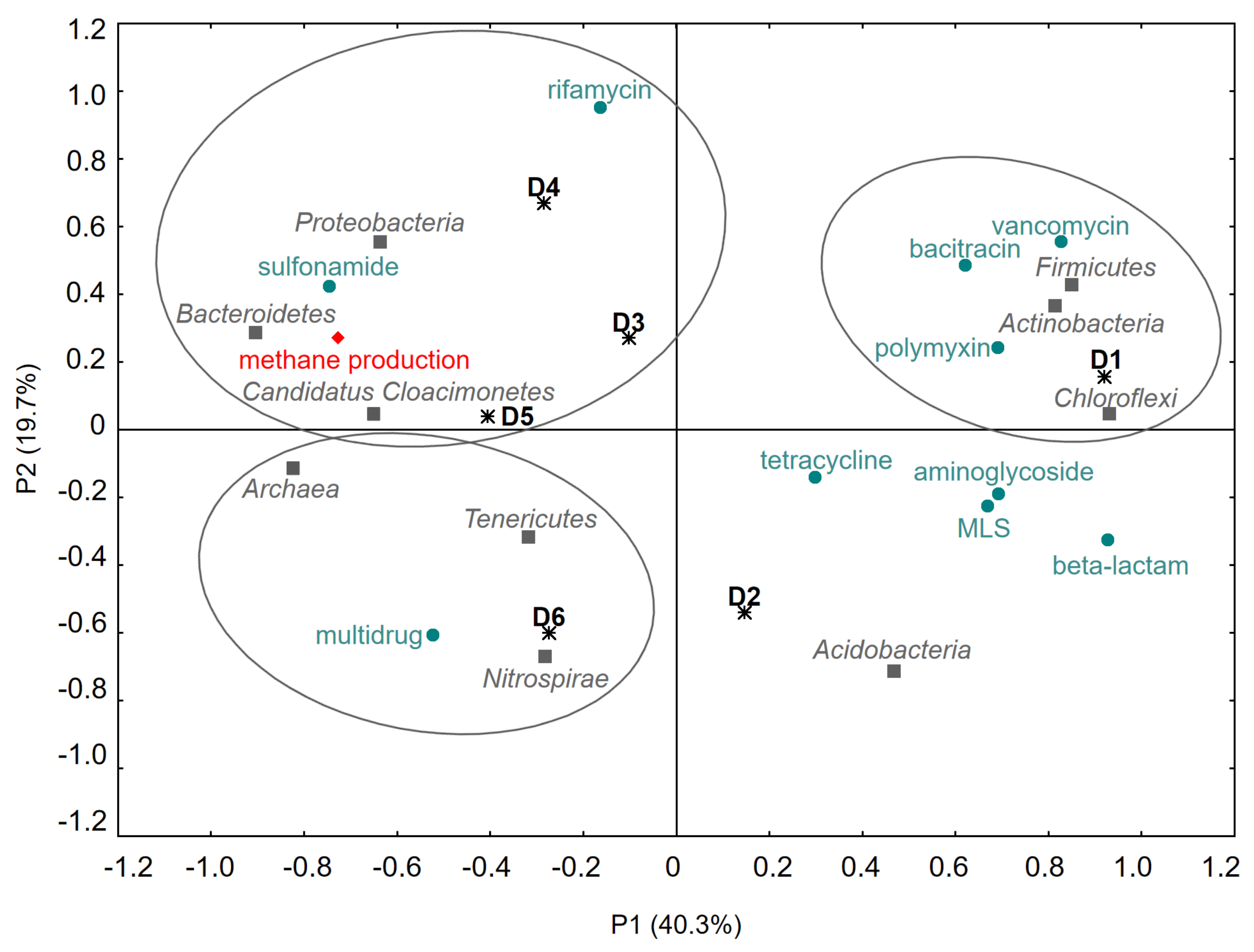
Long-Term, Simultaneous Impact of Antimicrobials on the Efficiency of Anaerobic Digestion of Sewage Sludge and Changes in the Microbial Community

 , ,
, ,  , , , ,
, , , ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Share and Cite
Czatzkowska, M.; Harnisz, M.; Korzeniewska, E.; Wolak, I.; Rusanowska, P.; Paukszto, Ł.; Jastrzębski, J.P.; Bajkacz, S. Long-Term, Simultaneous Impact of Antimicrobials on the Efficiency of Anaerobic Digestion of Sewage Sludge and Changes in the Microbial Community. Energies 2022, 15, 1826. https://doi.org/10.3390/en15051826
Czatzkowska M, Harnisz M, Korzeniewska E, Wolak I, Rusanowska P, Paukszto Ł, Jastrzębski JP, Bajkacz S. Long-Term, Simultaneous Impact of Antimicrobials on the Efficiency of Anaerobic Digestion of Sewage Sludge and Changes in the Microbial Community. Energies. 2022; 15(5):1826. https://doi.org/10.3390/en15051826
Chicago/Turabian StyleCzatzkowska, Małgorzata, Monika Harnisz, Ewa Korzeniewska, Izabela Wolak, Paulina Rusanowska, Łukasz Paukszto, Jan P. Jastrzębski, and Sylwia Bajkacz. 2022. "Long-Term, Simultaneous Impact of Antimicrobials on the Efficiency of Anaerobic Digestion of Sewage Sludge and Changes in the Microbial Community" Energies 15, no. 5: 1826. https://doi.org/10.3390/en15051826
APA StyleCzatzkowska, M., Harnisz, M., Korzeniewska, E., Wolak, I., Rusanowska, P., Paukszto, Ł., Jastrzębski, J. P., & Bajkacz, S. (2022). Long-Term, Simultaneous Impact of Antimicrobials on the Efficiency of Anaerobic Digestion of Sewage Sludge and Changes in the Microbial Community. Energies, 15(5), 1826. https://doi.org/10.3390/en15051826