
Relative Importance of Landscape and Climate Factors to the Species Diversity of Plant Growth Forms along an East Asian Archipelago


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Plant Diversity and Environmental Variables
2.3. Statistical Analysis
3. Results
3.1. Plant Species Composition
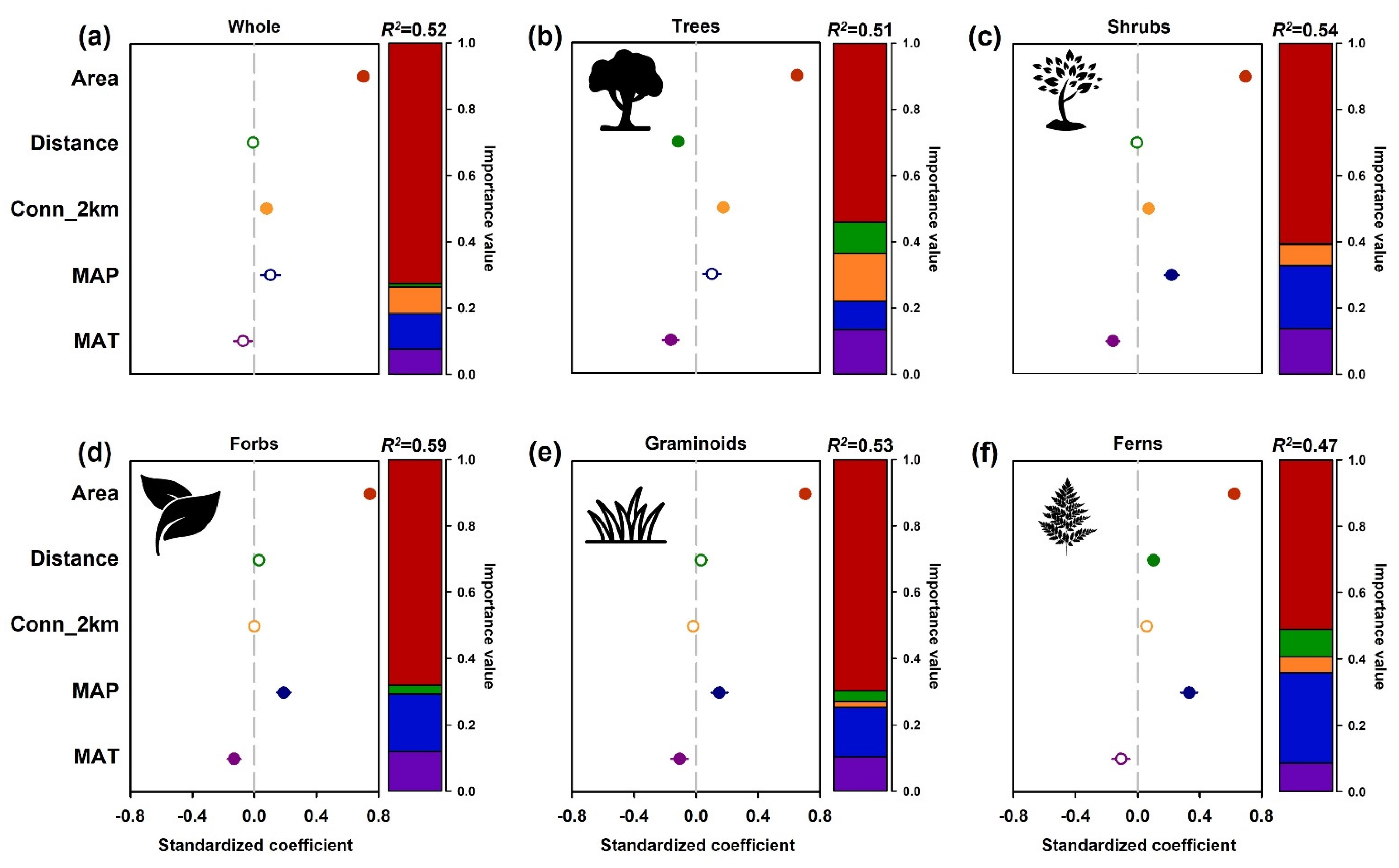
3.2. Species Richness Patterns of Plant Growth Forms with Environmental Variables in Overall Islands
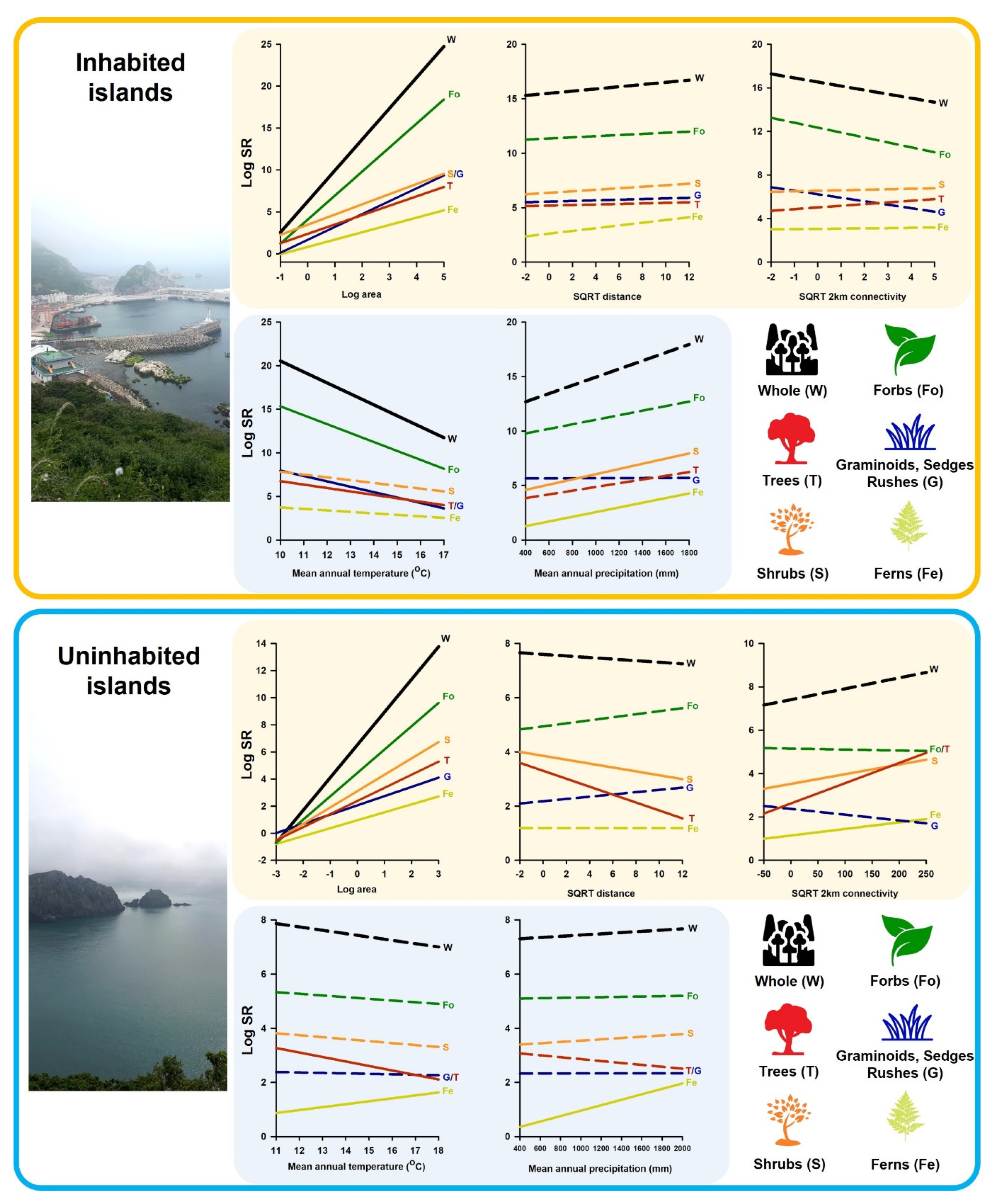
3.3. Species Richness Patterns of Plant Growth Forms with Environmental Variables between Inhabited and Uninhabited Islands
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, S.D. Studies of Avian species in the islands of Korea based on the theory of island biogeography. Environ. Impact Assess. 2010, 19, 547–552. [Google Scholar]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Whittaker, R.J.; Fernández-Palacios, J.M. Island Biogeography: Ecology, Evolution, and Conservation; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Dale, S. Islands in the forest: Effects of patch size and isolation on farmland bird species richness and community composition of farmland patches in forest landscapes. Landsc. Ecol. 2019, 34, 2823–2836. [Google Scholar] [CrossRef]
- Wallace, A.R. The Malay Archipelago: The Land of the Orang-Utan and the Bird of Paradise; a Narrative of Travel, with Studies of Man and Nature; Courier Corporation: North Chelmsford, MA, USA, 1962. [Google Scholar]
- Whittaker, R.H.; Niering, W.A. Vegetation of the Santa Catalina Mountains, Arizona. V. Biomass, production, and diversity along the elevation gradient. Ecology 1975, 56, 771–790. [Google Scholar] [CrossRef] [Green Version]
- Connor, E.F.; McCoy, E.D. The statistics and biology of the species-area relationship. Am. Nat. 1979, 113, 791–833. [Google Scholar] [CrossRef]
- Lomolino, M.V. Ecology’s most general, yet protean pattern: The species-area relationship. J. Biogeogr. 2000, 27, 17–26. [Google Scholar] [CrossRef]
- Aggemyr, E.; Auffret, A.G.; Jädergård, L.; Cousins, S.A. Species richness and composition differ in response to landscape and biogeography. Landsc. Ecol. 2018, 33, 2273–2284. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.H.; Lee, C.B. On the relative importance of landscape variables to plant diversity and phylogenetic community structure on uninhabited islands, South Korea. Landsc. Ecol. 2021, 36, 209–221. [Google Scholar] [CrossRef]
- Lindgren, J.P.; Cousins, S.A. Island biogeography theory outweighs habitat amount hypothesis in predicting plant species richness in small grassland remnants. Landsc. Ecol. 2017, 32, 1895–1906. [Google Scholar] [CrossRef]
- Schrader, J.; Moeljono, S.; Keppel, G.; Kreft, H. Plants on small islands revisited: The effects of spatial scale and habitat quality on the species–area relationship. Ecography 2019, 42, 1405–1414. [Google Scholar] [CrossRef] [Green Version]
- De Kort, H.; Prunier, J.G.; Ducatez, S.; Honnay, O.; Baguette, M.; Stevens, V.M.; Blanchet, S. Life history, climate and biogeography interactively affect worldwide genetic diversity of plant and animal populations. Nat. Commun. 2021, 12, 516. [Google Scholar] [CrossRef]
- Ottaviani, G.; Keppel, G.; Götzenberger, L.; Harrison, S.; Opedal, Ø.H.; Conti, L.; Liancourt, P.; Klimešová, J.; Silveira, F.A.; Jiménez-Alfaro, B. Linking plant functional ecology to island biogeography. Trends Plant Sci. 2020, 25, 329–339. [Google Scholar] [CrossRef]
- Ross, S.R.J.; Friedman, N.R.; Janicki, J.; Economo, E.P. A test of trophic and functional island biogeography theory with the avifauna of a continental archipelago. J. Anim. Ecol. 2019, 88, 1392–1405. [Google Scholar] [CrossRef]
- Li, S.P.; Wang, P.; Chen, Y.; Wilson, M.C.; Yang, X.; Ma, C.; Lu, J.; Chen, X.Y.; Wu, J.; Shu, W.S. Island biogeography of soil bacteria and fungi: Similar patterns, but different mechanisms. ISME J. 2020, 14, 1886–1896. [Google Scholar] [CrossRef] [PubMed]
- Negoita, L.; Fridley, J.D.; Lomolino, M.V.; Mittelhauser, G.; Craine, J.M.; Weiher, E. Isolation-driven functional assembly of plant communities on islands. Ecography 2016, 39, 1066–1077. [Google Scholar] [CrossRef]
- Auffret, A.G.; Aggemyr, E.; Plue, J.; Cousins, S.A. Spatial scale and specialization affect how biogeography and functional traits predict long-term patterns of community turnover. Funct. Ecol. 2017, 31, 436–443. [Google Scholar] [CrossRef]
- Ottaviani, G.; Keppel, G.; Marcantonio, M.; Mucina, L.; Wardell-Johnson, G. Woody species in resource-rich microrefugia of granite outcrops display unique functional signatures. Austral. Ecol. 2019, 44, 575–580. [Google Scholar] [CrossRef]
- Wullschleger, S.D.; Epstein, H.E.; Box, E.O.; Euskirchen, E.S.; Goswami, S.; Iversen, C.M.; Kattge, J.; Norby, R.J.; van Bodegom, P.M.; Xu, X. Plant functional types in Earth system models: Past experiences and future directions for application of dynamic vegetation models in high-latitude ecosystems. Ann. Bot. 2014, 114, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Šímová, I.; Violle, C.; Svenning, J.C.; Kattge, J.; Engemann, K.; Sandel, B.; Peet, R.K.; Wiser, S.K.; Blonder, B.; McGill, B.J. Spatial patterns and climate relationships of major plant traits in the New World differ between woody and herbaceous species. J. Biogeogr. 2018, 45, 895–916. [Google Scholar] [CrossRef] [Green Version]
- Chi, Y.; Sun, J.; Fu, Z.; Xie, Z. Spatial pattern of plant diversity in a group of uninhabited islands from the perspectives of island and site scales. Sci. Total Environ. 2019, 664, 334–346. [Google Scholar] [CrossRef]
- Russell, J.C.; Abrahão, C.R.; Silva, J.C.; Dias, R.A. Management of cats and rodents on inhabited islands: An overview and case study of Fernando de Noronha, Brazil. Perspect. Ecol. Conserv. 2018, 16, 193–200. [Google Scholar] [CrossRef]
- Nogué, S.; de Nascimento, L.; Froyd, C.A.; Wilmshurst, J.M.; de Boer, E.J.; Coffey, E.E.; Whittaker, R.J.; Fernández-Palacios, J.M.; Willis, K.J. Island biodiversity conservation needs palaeoecology. Nat. Ecol. Evol. 2017, 1, 0181. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.H.; Kim, Y.J.; Lim, H.M.; Han, Y.G.; Choi, M.J.; Nam, S.H. A faunistic study of insects of uninhabited islands in the Docho-myeon, Sinan-gun, Jeollanam-do, Korea. Korean J. Environ. Ecol. 2011, 25, 673–684. [Google Scholar]
- Jung, S.H.; Cho, Y.C. Redefining floristic zones in the Korean Peninsula using high-resolution georeferenced specimen data and self-organizing maps. Ecol. Evol. 2020, 10, 11549–11564. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Cho, K.M.; Hong, S.K.; Kim, J.E.; Kim, K.W.; Lee, K.A.; Moon, K.O. Management plan for UNESCO Shinan Dadohae Biosphere Reserve (SDBR), Republic of Korea: Integrative perspective on ecosystem and human resources. J. Ecol. Environ. 2010, 33, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.M.; Kang, C.W.; Jeon, C.H.; Oh, M.R. A Study on the Distribution of Fairy Pitta (Pitta nympha) and the Forest Type on Inhabited Islands in the Southern Coast of Jeollanamdo Province, Republic of Korea. J. Korean Isl. 2016, 28, 259–271. [Google Scholar]
- National Institute of Environmental Research. National Natural Environment Survey of Uninhabited Islands: Yeosu II.; National Institute of Environmental Research: Incheon, Korea, 2003. [Google Scholar]
- Lee, C.B.; Kim, H.H. Elevational patterns of plant species richness and relative importance of climatic and topographic factors on the mt. seorak, South Korea. J. Agricult. Life Sci. 2018, 52, 702. [Google Scholar] [CrossRef]
- Spanowicz, A.G.; Jaeger, J.A. Measuring landscape connectivity: On the importance of within-patch connectivity. Landsc. Ecol. 2019, 34, 2261–2278. [Google Scholar] [CrossRef]
- Yun, J.I. Agroclimatic maps augmented by a GIS technology. Korean J. Agric. For. Meteorol. 2010, 12, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Bumham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar]
- Symonds, M.R.; Moussalli, A. A brief guide to model selection, multimodel inference and model averaging in behavioural ecology using Akaike’s information criterion. Behav. Ecol. Sociobiol. 2011, 65, 13–21. [Google Scholar] [CrossRef]
- Graham, M.H. Confronting multicollinearity in ecological multiple regression. Ecology 2003, 84, 2809–2815. [Google Scholar] [CrossRef] [Green Version]
- Mason, N.W.; Mouillot, D.; Lee, W.G.; Wilson, J.B. Functional richness, functional evenness and functional divergence: The primary components of functional diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- Schrader, J.; König, C.; Triantis, K.A.; Trigas, P.; Kreft, H.; Weigelt, P. Species–area relationships on small islands differ among plant growth forms. Global Ecol. Biogeogr. 2020, 29, 814–829. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.K.; Chun, J.H.; Lee, C.B. Prediction of distribution changes of Carpinus laxiflora and C. tschonoskii based on climate change scenarios using MaxEnt model. Korean J. Agric. For. Meteorol. 2021, 23, 55–67. [Google Scholar]
- Chase, J.M.; Knight, T.M. Scale-dependent effect sizes of ecological drivers on biodiversity: Why standardised sampling is not enough. Ecol. Lett. 2013, 16, 17–26. [Google Scholar] [CrossRef]
- Marx, H.E.; Dentant, C.; Renaud, J.; Delunel, R.; Tank, D.C.; Lavergne, S. Riders in the sky (islands): Using a mega-phylogenetic approach to understand plant species distribution and coexistence at the altitudinal limits of angiosperm plant life. J. Biogeogr. 2017, 44, 2618–2630. [Google Scholar] [CrossRef]
- Portillo, J.T.d.M.; Ouchi-Melo, L.S.; Crivellari, L.B.; de Oliveira, T.A.L.; Sawaya, R.J.; Duarte, L.d.S. Area and distance from mainland affect in different ways richness and phylogenetic diversity of snakes in Atlantic Forest coastal islands. Ecol. Evol. 2019, 9, 3909–3917. [Google Scholar] [CrossRef]
- An, J.S.; Choi, S.W. Study on the pattern and ecological variables of butterfly species diversity on 19 uninhabited islands around Imja-myeon, Shinan-gun, Jeonnam, Korea. Korean J. Environ. Biol. 2012, 30, 185–192. [Google Scholar]
- Choi, S.W.; An, J.S.; Yang, H.S. Effect of island geography on plant species on uninhabited islands in southeastern South Korea. J. Ecol. Environ. 2015, 38, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Baum, K.A.; Haynes, K.J.; Dillemuth, F.P.; Cronin, J.T. The matrix enhances the effectiveness of corridors and stepping stones. Ecology 2004, 85, 2671–2676. [Google Scholar] [CrossRef]
- Auffret, A.G.; Rico, Y.; Bullock, J.M.; Hooftman, D.A.; Pakeman, R.J.; Soons, M.B.; Suárez-Esteban, A.; Traveset, A.; Wagner, H.H.; Cousins, S.A. Plant functional connectivity—integrating landscape structure and effective dispersal. J. Ecol. 2017, 105, 1648–1656. [Google Scholar] [CrossRef]
- Lee, C.B.; Chun, J.H.; Ahn, H.H. Elevational patterns of plant richness and their drivers on an Asian mountain. Nord. J. Bot. 2014, 32, 347–357. [Google Scholar] [CrossRef]
- Levey, D.J.; Moermond, T.C.; Denslow, J.S. Frugivory: An overview. In La Selva: Ecology and Natural History of a Neotropical Rain Forest; University of Chicago Press: Chicago, IL, USA, 1994; pp. 282–294. [Google Scholar]
- Muller-Landau, H.C.; Hardesty, D. Seed dispersal of woody plants in tropical forests: Concepts, examples and future directions. In Biotic Interactions in the Tropics: Their Role in the Maintenance of Species Diversity; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Harrison, S.; Spasojevic, M.J.; Li, D. Climate and plant community diversity in space and time. Proc. Nat. Acad. Sci. 2020, 117, 4464–4470. [Google Scholar] [CrossRef] [PubMed]
- Vranckx, G.; Jacquemyn, H.; Muys, B.; Honnay, O. Meta-analysis of susceptibility of woody plants to loss of genetic diversity through habitat fragmentation. Conserv. Biol. 2012, 26, 228–237. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, R.H.; MacArthur, J.W. On bird species diversity. Ecology 1961, 42, 594–598. [Google Scholar] [CrossRef]
- Chi, Y.; Shi, H.; Wang, X.; Li, J.; Feng, A. The spatial distribution and impact factors of net primary productivity in island ecosystem of five Southern Islands of Miaodao archipelago. Acta Ecol. Sin. 2015, 35, 8094–8106. [Google Scholar]
- Chen, S.; Wang, W.; Xu, W.; Wang, Y.; Wan, H.; Chen, D.; Tang, Z.; Tang, X.; Zhou, G.; Xie, Z. Plant diversity enhances productivity and soil carbon storage. Proc. Nat. Acad. Sci. 2018, 115, 4027–4032. [Google Scholar] [CrossRef] [Green Version]
- Chun, J.H.; Lee, C.B. Temporal changes in species, phylogenetic, and functional diversity of temperate tree communities: Insights from assembly patterns. Front. Plant Sci. 2019, 10, 294. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.-K.; Lee, H.-S.; Lee, H.-I.; Lee, S.-W.; Lee, Y.-J.; Lee, C.-B. Relative Importance of Landscape and Climate Factors to the Species Diversity of Plant Growth Forms along an East Asian Archipelago. Forests 2022, 13, 218. https://doi.org/10.3390/f13020218
Lee M-K, Lee H-S, Lee H-I, Lee S-W, Lee Y-J, Lee C-B. Relative Importance of Landscape and Climate Factors to the Species Diversity of Plant Growth Forms along an East Asian Archipelago. Forests. 2022; 13(2):218. https://doi.org/10.3390/f13020218
Chicago/Turabian StyleLee, Min-Ki, Ho-Sang Lee, Hae-In Lee, Sang-Wook Lee, Yong-Ju Lee, and Chang-Bae Lee. 2022. "Relative Importance of Landscape and Climate Factors to the Species Diversity of Plant Growth Forms along an East Asian Archipelago" Forests 13, no. 2: 218. https://doi.org/10.3390/f13020218