
Effects of Litter Decomposition on Soil N in Picea mongolica Forest at Different Forest Ages

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Sample Plot Setting and Collection and Pre-Treatment of Litter
2.3. Sample Analysis and Data Statistics
3. Results and Analysis
3.1. Chemical Composition of P. mongolica Forest Litter in Different Stands
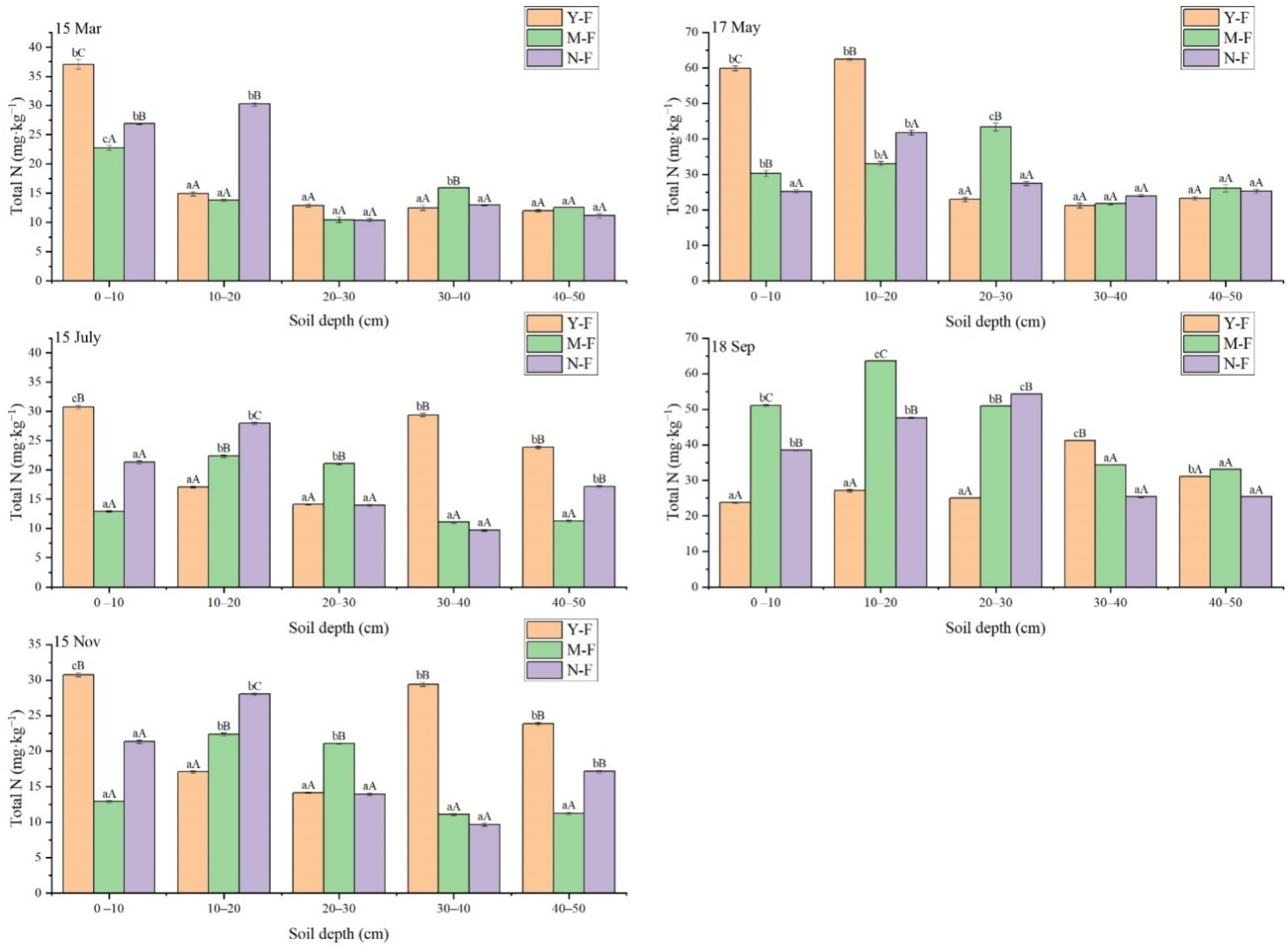
3.2. Seasonal Variation of Soil Total N Content with Soil Depth and Stand Age
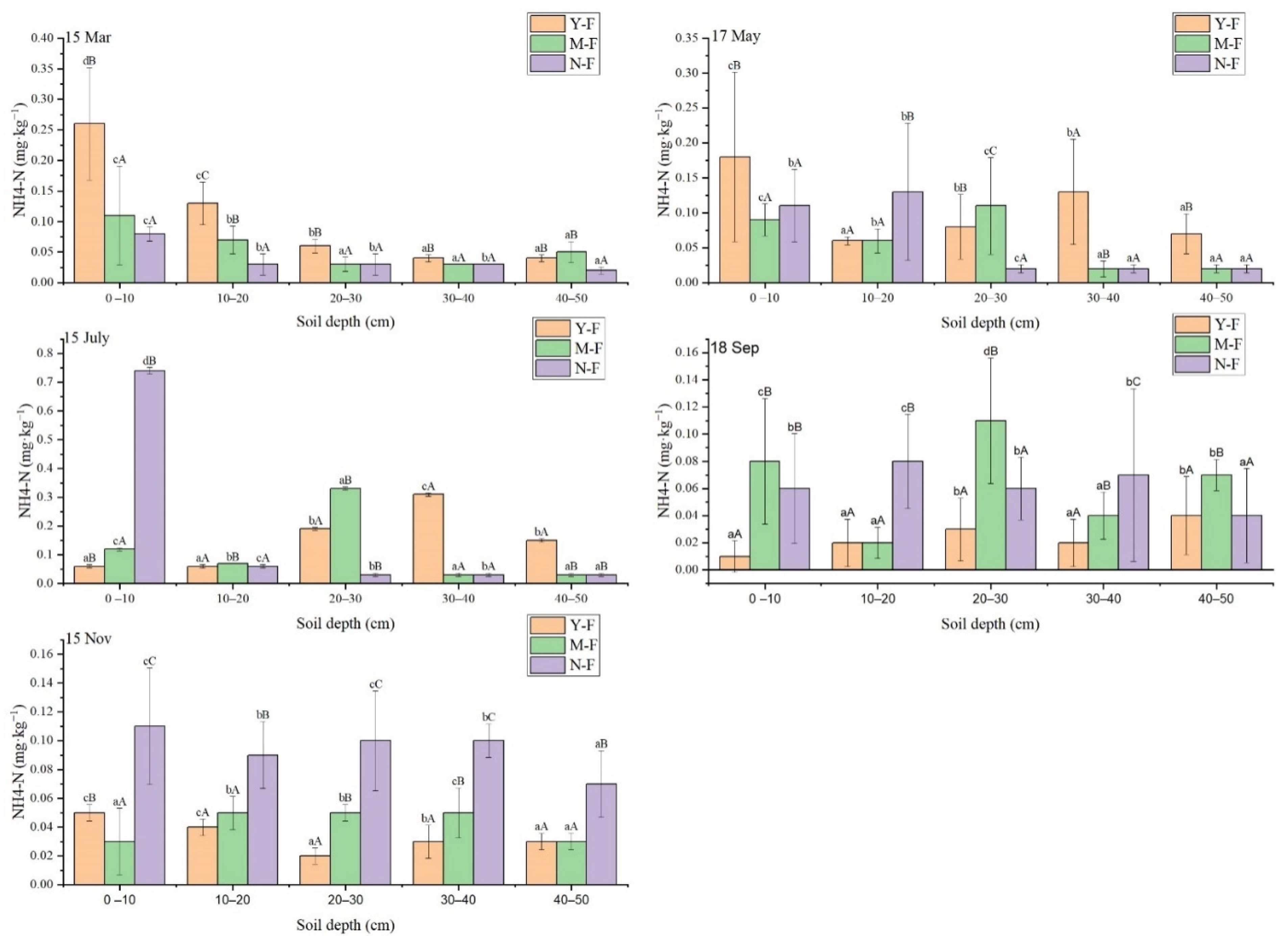
3.3. Characteristics of Seasonal Variation of Soil NH4+—N Content with Soil Depth and Stand Age
3.4. Characteristics in Seasonal Dynamic Variation of Soil NO3−—N Content with Soil Depth and Stand Age
3.5. Correlation between Soil Total N, NH4+—N, NO3−—N Content and Month, Soil Depth, and Forest Age
4. Discussion
4.1. Effects of Litter Addition on the Contents of Total N, NH4+—N, and NO3−—N in the Soil of P. mongolica Forest
4.2. Effects of Environmental Factors on the Contents of Total N, NH4+—N, and NO3−—N the Soil of P. mongolica Forest
4.3. Effects of Seasonal Variation on the Contents of total N, NH4+—N, and NO3−—N in the Soil of P. mongolica Forest
4.4. Effect of Soil Depth on the Contents of Total N, NH4+—N, and NO3−—N in the Soil of P. mongolica Forest
4.5. Effects of Forest Age on the Contents of Total N, NH4+—N, and NO3−—N in the Soil of P. mongolica Forest
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grant, R.F. Nitrogen Mineralization Drives the Response of Forest Productivity to Soil Warming: Modelling in Ecosys vs. Measurements from the Harvard Soil Heating Experiment. Ecol. Model. 2014, 288, 38–46. [Google Scholar] [CrossRef]
- Ma, H.F.; Xie, M.Y.; Hu, H.; Guo, Y.X.; Ren, C.J.; Zhao, F.Z. Effects of Stoichiometric Characteristics of Soil-plant-litter on soil nitrogen components in different forests along an elevational gradient of Qinling Mountains. Chin. J. Ecol. 2020, 39, 749–757. Available online: http://www.cje.net.cn/CN/Y2020/V39/I3/749 (accessed on 25 December 2021).
- Xu, C.Q.; Chen, L.X.; Yan, Y.Q.; Ji, X. Seasonal dynamic characteristics of ammonium-nitrogen and nitrate nitrogen contents in soil in temperate forests. J. Northeast For. Univ. 2008, 10, 19–21. [Google Scholar]
- Pan, F.J.; Liang, Y.M.; Ma, J.M.; Yang, Z.Q.; Ling, T.W.; Li, M.J.; Lu, S.H.; Zhong, F.Y. Stand Age and Density Affected Litter Nutrient Changes in Planted Pinus Massoniana Forests. Guihaia 2020, 40, 237–246. [Google Scholar] [CrossRef]
- Yang, X.; Chen, L.H.; Zheng, X.L. Ecological Stoichiometry Characterization of Soil Carbon, Nitrogen, and Phosphorus of Pinus tabuliformis Plantations at Different Stand Ages. Sci. Soil Water Conserv. 2021, 19, 108–116. [Google Scholar] [CrossRef]
- Fang, L.N.; Yang, X.D.; Du, J. Effects of Land Use Pattern on Soil Microbial Biomass Carbon in Xishuangbanna. Chin. J. Appl. Ecol. 2011, 22, 837–844. [Google Scholar] [CrossRef]
- Xu, J.F.; Zhao, J.H.; Yuan, Z.X.; Jin, X.M.; Chen, B.; Guan, Q.W.; Gao, Y. Effects of Tree Species and Land use Patterns on Soil Microbial Biomass Carbon & Nitrogen. J. Cent. South Univ. For. Technol. 2018, 38, 95–100. [Google Scholar] [CrossRef]
- Hu, Y.S.; Yao, X.Y.; Liu, Y.H. N and P Stoichiometric Traits of Plant and Soil in Different Forest Succession Stages in Changbai Mountains. Chin. J. Appl. Ecol. 2014, 25, 632–638. Available online: http://www.cjae.net/CN/Y2014/V25/I3/632 (accessed on 25 December 2021).
- Wang, X.M.; Yan, B.G.; Zhao, G.; Shi, L.T.; Liu, G.C.; Fang, H.D. Effects of Microorganism on Carbon, Nitrogen and Phosphorus of Dodonaea viscosa and the Soils from Different Elevations in Yuanmou, Yunnan, China. Chin. J. Plant Ecol. 2017, 41, 311–324. [Google Scholar] [CrossRef]
- Zhang, S.S.; Li, A.Q.; Wang, H.R.; Wang, J.J.; Xu, X.N. Ecological Stoichiometry of Soil Carbon, Nitrogen and Phosphorus in Cunninghamia lanceolata Plantation Across an Elevation Gradient. Ecol. Environ. Sci. 2020, 29, 97–104. [Google Scholar] [CrossRef]
- Zhao, W.J.; Liu, X.D.; Jing, M.; Zhang, X.L.; Che, Z.X.; Jing, W.M.; Wang, S.L.; Niu, Y.; Qi, P.; Li, W.J. Ecological Stoichiometric Characteristics of Carbon, Nitrogen and Phosphorus in Leaf-Litter-Soil System of Picea crassifolia Forest in the Qilian Mountains. Acta Pedol. Sin. 2016, 53, 477–489. [Google Scholar] [CrossRef]
- Meng, Y.; Xue, J.Y.; Sha, L.Q.; Tang, J.W. Variations of Soil NH4-N, NO3−—Nand N Mineralization under Different Forests in Xishuang-banna, Southwest China. Chin. J. Plant Ecol. 2001, 25, 99–104. Available online: https://www.plant-ecology.com/CN/Y2001/V25/I1/99 (accessed on 25 December 2021).
- Castro, H.F.; Classen, A.T.; Austin, E.E.; Norby, R.J.; Schadt, C.W. Soil Microbial Community Responses to Multiple Experimental Climate Change Drivers. Appl. Environ. Microbiol. 2010, 76, 999–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, X.F.; Xu, F.L.; Wang, W.L.; Zhao, Y.F.; Wang, L.L.; Sun, P.Y. Ecological Stoichiometry of Soil Carbon, Nitrogen and Phosphorus in a Larix Principis Rupprechtii Plantation. Sci. Soil Water Conserv. 2015, 13, 68–75. [Google Scholar] [CrossRef]
- Magill, A.H.; Aber, J.D.; Berntson, G.M.; McDowell, W.H.; Nadelhoffer, K.J.; Melillo, J.M.; Steudler, P. Long-Term Nitrogen Additions and Nitrogen Saturation in Two Temperate Forests. Ecosystems 2000, 3, 238–253. [Google Scholar] [CrossRef]
- Lv, R.H.; Li, G.L.; Liu, Y.; Jia, L.M.; Jiang, P.; Lin, N. Decomposition Characteristics of coniferous Litter under Different Site Conditions in a Larix principis-rupprechtii Plantation. Sci. Silvae Sin. 2012, 48, 31–37. [Google Scholar] [CrossRef]
- Liu, Z.J.; Lin, W.S.; Yang, Z.R.; Lin, T.W.; Liu, X.F.; Chen, Y.M.; Yang, Y.S. Effects of Soil Warming and Nitrogen Deposition on Available Nitrogen in a Young Cunninghamia lanceolata Stand in Mid-Subtropical China. Acta Ecol. Sin. 2017, 37, 44–53. [Google Scholar] [CrossRef]
- Zhao, W.J.; Liu, Y.T.; Tan, C.J.; Cui, Y.C.; Wu, P.; Hou, Y.J.; Liu, Y.H.; Ding, J.J. Effects of simulated nitrogen deposition on soil nitrogen mineralization in karst primary forest. J. Cent. South Univ. For. Technol. 2021, 41, 124–133. [Google Scholar] [CrossRef]
- Zhang, G.S.; Deng, H.J.; Du, K.; Lin, Y.M.; Ma, R.F.; Yu, W.; Wang, D.J.; Wu, C.Z.; Hong, W. Soil Stoichiometry Characteristics at Different Elevation Gradients of a Mountain in an Area with High Frequency Debris Flow: A Case Study in Xiaojiang Watershed, Yunnan. Acta Ecol. Sin. 2016, 36, 675–687. [Google Scholar] [CrossRef]
- Chen, L.X.; Huang, L.Y.; Qiao, L.; Duan, W.B.; Ji, X.; Yu, Y.C. Influence of Simulated Nitrogen Deposition Mineralization Rate under Different Temperate Forest Stands. J. Soil Water Conserv. 2012, 26, 139–146. [Google Scholar] [CrossRef]
- Zhang, L.; Huang, J.H.; Bai, Y.F.; Han, X.G. Effects of Nitrogen Addition on Net Nitrogen Mineralization in LeymuschinensisGraassland, Inner Mongolia, China. Chin. J. Plant Ecol. 2009, 33, 563–569. Available online: https://www.plant-ecology.com/CN/Y2009/V33/I3/56 (accessed on 25 December 2021). [CrossRef]
- Fang, Y.T.; Mo, J.M.; Zhou, G.Y.; Per Gundersen, L.D.J.; Jiang, Y.Q. The Short-Term Responses of Soil Available Nitrogen of Dinghushan Forests to Simulated N Deposition in Subtropical China. Acta Ecol. Sin. 2004, 24, 73–80. Available online: https://www.ecologica.cn/stxb/article/abstract/040730?st=search (accessed on 25 December 2021).
- Hester, E.R.; Harpenslager, S.F.; van Diggelen, J.M.H.; Lamers, L.L.; Jetten, M.S.M.; Lüke, C.; Lücker, S.; Welte, C.U. Linking Nitrogen Load to the Structure and Function of Wetland Soil and Rhizosphere Microbial Communities. mSystems 2018, 3, e00214-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barton, L.; Murphy, D.V.; Butterbach-Bahl, K. Influence of Crop Rotation and Liming on Greenhouse Gas Emissions from a Semi-Arid Soil. Agric. Ecosyst. Environ. 2013, 167, 23–32. [Google Scholar] [CrossRef]
- ŠImek, M.; Cooper, J.E. The Influence of Soil pH on Denitrification: Progress Towards the Understanding of This Interaction over the Last 50 Years. Eur. J. Soil Sci. 2002, 53, 345–354. [Google Scholar] [CrossRef]
- Deng, X.H.; Cai, X.; Tian, F.; Cai, Y.F.; Tian, M.H.; Zhang, L.M.; Zhang, M.F. pH and Main Nutrients of Tobacco-Growing Soils and Their Relations in Western Hunan. Soils 2017, 49, 49–56. [Google Scholar] [CrossRef]
- Chen, G.T.; Li, S.; Peng, T.C.; Qiu, X.R.; Chen, Y.Q.; Ma, H.Y.; Tu, L.H. Effect of Root Isolation and Nitrogen Addition on Soil Carbon and Nitrogen Component in a Moist Evergreen Broad-Leaved Forest. Chin. J. Appl. Environ. Biol. 2018, 24, 125–131. [Google Scholar] [CrossRef]
- Xu, Y.B.; Xu, Z.H.; Cai, Z.C. Progresses in Research on Denitrification in Tropical and Subtropical Soils of Terrestrial Ecosystems. Ecol. Environ. Sci. 2014, 23, 1557–1566. Available online: http://www.jeesci.com/CN/Y2014/V23/I9/1557 (accessed on 25 December 2021).
- Wang, X.; Gao, M.D.; Yang, F.; Guo, Y.P.; Ma, C.M. Litter Decomposition and Nutrient Dynamics of LarixPrincipis-Rupprechtii Plantations of Different Ages. J. Northeast For. Univ. 2012, 40, 56–60, 66. [Google Scholar] [CrossRef]
- Glass, A.D.M.; Siddiqi, M.Y. Nitrogen Absorption by Plant Roots; Srivastava, H.S., Singh, R.P., Eds.; Nitrogen Nutrition in Higher Plants; Associated Publishing Co.: New Delhi, India, 1995; pp. 21–56. [Google Scholar]
- Li, X.Y.; Zhang, W.Y.; Liu, F.; Zhang, Z.M.; He, T.B.; Lin, C.H. The Distribution Characteristics of Soil Carbon, Nitrogen and Phosphorus at Different Altitudes in Fanjing Mountain. Res. Soil Water Conserv. 2016, 23, 19–24. [Google Scholar] [CrossRef]
- Wang, Y.L.; Zi, H.B.; Cheng, R.X.; Tang, L.T.; Suoer Azhi, L.X.P.; Li, J.; Wang, C.T. Forest Soil Organic Carbon and Nitrogen Storage and Characteristics of Vertical Distribution in Qinghai Province. Acta Ecol. Sin. 2019, 39, 4096–4105. [Google Scholar] [CrossRef]
- Wang, S.P.; Zhou, G.S.; Lu, Y.C.; Zou, J.J. Distribution of Soil Carbon, Nitrogen and Phosphorus Along Northeast China Transect (NECT) and Their Relationships with Climatic Factors. Chin. J. Plant Ecol. 2002, 26, 513–517. Available online: https://www.plant-ecology.com/CN/Y2002/V26/I5/513 (accessed on 25 December 2021).
- Qin, J.; Kong, H.Y.; Liu, H. Stoichiometric Characteristics of Soil C, N, P and K in Different Pinus Massoniana Forests. J. Northwest. Agric. For. Univ. 2016, 44, 68–76. [Google Scholar] [CrossRef]
- Wang, P.A.; Ren, X.M.; Bo, F.J.; Zhang, Y.X.; Guo, J.P. Analysis on Soil Stoichiometric Characteristics and Soil Physico Chemical Properties of Larix principis-rupprechtii Plantations in Different Forest Ages in Guandi Mountain. J. Shanxi Agric. Sci. 2020, 48, 956–963. Available online: https://sxlx.cbpt.cnki.net/WKB3/WebPublication/paperDigest.aspx?paperID=ff65f658-9c16-4de5-aa05-11d0093ce5e3 (accessed on 25 December 2021).
- Cui, N.J.; Zhang, D.J.; Liu, Y.; Zhang, J.; Ou, J.; Zhang, J.; Deng, C.; Ji, T.W. Plant Diversity and Soil Physicochemical Properties under Different Aged Pinus Massoniana Plantations. Chin. J. Ecol. 2014, 33, 2610–2617. Available online: http://www.cje.net.cn/CN/Y2014/V33/I10/2610 (accessed on 25 December 2021).
- Wang, D.; Dai, W.; Wang, B.; Li, P.; Deng, Z.F.; Zhao, C. Changes of Soil Properties at Different Developmental Stages of Chinese Fir Plantations. J. Beijing For. Univ. 2010, 32, 59–63. [Google Scholar] [CrossRef]
- Peng, Y.H.; Tan, C.Q.; Shen, W.H.; Zheng, W.; He, Q.F.; He, F.; Chen, S.G. Effects of Differt Forest Age on Soil Physical and Chemical Properties of Cunninghamia lanceolata Plantations. Guangxi For. Sci. 2020, 49, 361–367. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Forest Type | Soil Layer (0–30 cm) | |||||
|---|---|---|---|---|---|---|
| Age of Stand | Elevation Gradient | Average DBH | Average Height | Soil pH | Soil Moisture/ | Soil Temperature |
| (a) | m | cm | m | % | °C | |
| Young forest (0–5a) | 1344 | 1.79 ± 2.15 | 2.06 ± 0.71 | 7.01 ± 0.12 | 35.95 ± 0.45 | −15.78 ± 2.49 |
| Middle-aged forest (5–30a) | 1352 | 8.03 ± 4.81 | 8.15 ± 1.86 | 6.67 ± 0.03 | 42.45 ± 4.13 | −14.59 ± 2.61 |
| Near-mature forest (30–40a) | 1342 | 14.26 ± 4.12 | 13.94 ± 2.15 | 6.69 ± 0.01 | 17.28 ± 2.61 | −15.33 ± 2.42 |
| Months | Monthly Average Temperature (°C) | Monthly Precipitation (mm) | Soil pH Value | ||
|---|---|---|---|---|---|
| Y-F | M-F | N-F | |||
| 17 March | −10.4 | 1.38 | 7.28 | 6.72 | 6.97 |
| 17 May | 14.5 | 39.91 | 7.06 | 6.72 | 6.79 |
| 17 July | 22.62 | 95.12 | 6.37 | 5.89 | 6.49 |
| 17 September | 13.16 | 59.11 | 6.95 | 6.54 | 6.63 |
| 17 November | −6.48 | 3.93 | 7.14 | 6.26 | 6.54 |
| Sampling Time | Forest Type | Total C (mg·kg−1) | Total N (mg·kg−1) | C/N |
|---|---|---|---|---|
| 15 November 2016 | Litters | 262.74 | 4.29 | 61.21 |
| 15 March 2017 | Y-F | 261.96 ± 0.06aA | 4.33 ± 0.01aB | 60.49 ± 0.3aA |
| M-F | 257.69 ± 0.35aA | 4.37 ± 0.02aA | 58.97 ± 0.1aA | |
| N-F | 254.88 ± 0.13aB | 4.43 ± 0.01aB | 57.55 ± 0.10aA | |
| 17 May 2017 | Y-F | 246.99 ± 0.02bA | 4.55 ± 0.02bA | 54.27 ± 0.01bA |
| M-F | 239.13 ± 0.06bB | 4.68 ± 0.01bA | 51.10 ± 0.02bA | |
| N-F | 236.01 ± 0.42bB | 4.69 ± 0.05bA | 50.32 ± 0.47ba | |
| 15 July 2017 | Y-F | 233.49 ± 4.09bA | 4.77 ± 0.01bA | 48.95 ± 0.13bA |
| M-F | 226.46 ± 5.53bA | 4.84 ± 0.01bA | 46.79 ± 0.74bA | |
| N-F | 224.35 ± 16.09bA | 4.98 ± 0.01bA | 45.05 ± 1.11ba | |
| 18 September 2017 | Y-F | 228.59 ± 1.86bA | 5.09 ± 0.01bA | 44.91 ± 3.25cA |
| M-F | 204.22 ± 1.86bA | 5.37 ± 0.04bA | 38.03 ± 0.34ca | |
| N-F | 198.81 ± 1.36bA | 5.35 ± 0.04bA | 37.16 ± 0.54ca | |
| 15 November 2017 | Y-F | 203.12 ± 0.79aA | 5.28 ± 0.01aA | 38.47 ± 0.15cA |
| M-F | 193.43 ± 2.59aA | 5.42 ± 0.02aA | 35.69 ± 0.42cA | |
| N-F | 187.28 ± 1.89aA | 5.28 ± 0.07aA | 35.47 ± 0.16cA |
| Times | Soil Layer (cm) | Total N (mg·kg−1) | NNH4N (mg·kg−1) | NNO3N (mg·kg−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Y-F | M-F | N-F | Y-F | M-F | N-F | Y-F | M-F | N-F | ||
| 15 March | 0–10 | 37.01 ± 1.43bC | 22.73 ± 0.66cA | 26.88 ± 0.23bB | 0.26 ± 0.16dB | 0.11 ± 0.14cA | 0.08 ± 0.02cA | 21.93 ± 11.75cA | 24.03 ± 10.39cA | 25.19 ± 3.29cA |
| 10–20 | 14.92 ± 0.64aA | 13.82 ± 0.29aA | 30.25 ± 0.47bB | 0.13 ± 0.06cC | 0.07 ± 0.04bB | 0.03 ± 0.03bA | 12.81 ± 3.01bA | 11.87 ± 1.63bA | 15.47 ± 10.85bB | |
| 20–30 | 12.91 ± 0.52aA | 10.46 ± 0.83aA | 10.38 ± 0.55aA | 0.06 ± 0.02bB | 0.03 ± 0.02aA | 0.03 ± 0.03bA | 9.41 ± 0.58aA | 8.91 ± 0.56aA | 9.98 ± 1.80aA | |
| 30–40 | 12.52 ± 0.81aA | 15.96 ± 0.00bB | 12.96 ± 0.06aA | 0.04 ± 0.01aB | 0.03 ± 0.00aA | 0.03 ± 0.00bA | 8.69 ± 0.54aA | 12.86 ± 1.54bB | 9.89 ± 1.12aA | |
| 40–50 | 12.02 ± 0.41aA | 12.55 ± 0.00aA | 11.13 ± 0.64aA | 0.04 ± 0.01aB | 0.05 ± 0.03aB | 0.02 ± 0.01aA | 10.37 ± 1.08aA | 10.25 ± 1.33aA | 10.61 ± 3.04aA | |
| 17 May | 0–10 | 59.95 ± 1.21bC | 30.32 ± 1.44bB | 25.19 ± 0.76aA | 0.18 ± 0.21cB | 0.09 ± 0.04cA | 0.11 ± 0.09bA | 33.57 ± 14.81bB | 22.59 ± 6.16aA | 36.09 ± 12.77cB |
| 10–20 | 62.42 ± 0.64bB | 33.03 ± 1.11bA | 41.75 ± 1.21bA | 0.06 ± 0.01aA | 0.06 ± 0.03bA | 0.13 ± 0.17bB | 38.12 ± 16.38bB | 34.47 ± 15.62bA | 42.67 ± 24.25dC | |
| 20–30 | 22.92 ± 1.01aA | 43.37 ± 1.96cB | 27.42 ± 0.92aA | 0.08 ± 0.08bB | 0.11 ± 0.12cC | 0.02 ± 0.01cA | 21.01 ± 1.64aA | 36.09 ± 9.98bB | 40.24 ± 15.46dB | |
| 30–40 | 21.23 ± 1.25aA | 21.75 ± 0.35aA | 23.97 ± 0.62aA | 0.13 ± 0.13bA | 0.02 ± 0.02aB | 0.02 ± 0.01aA | 22.47 ± 2.84aA | 30.61 ± 18.72bB | 31.86 ± 14.58bB | |
| 40–50 | 23.25 ± 0.87aA | 26.08 ± 1.67aA | 25.32 ± 0.98aA | 0.07 ± 0.05aB | 0.02 ± 0.01aA | 0.02 ± 0.01aA | 20.05 ± 0.54aA | 21.96 ± 2.85aA | 24.12 ± 1.77aA | |
| 15 July | 0 –10 | 59.55 ± 0.52bC | 31.42 ± 0.46bB | 25.79 ± 0.27aA | 0.06 ± 0.01aB | 0.12 ± 0.01cA | 0.74 ± 0.02dB | 54.39 ± 0.01bC | 31.25 ± 0.01bB | 22.42 ± 0.01aA |
| 10–20 | 62.45 ± 0.69bC | 33.34 ± 0.58cA | 41.16 ± 0.20bB | 0.06 ± 0.01aA | 0.07 ± 0.00bB | 0.06 ± 0.01cA | 59.95 ± 0.01bB | 33.41 ± 0.02bA | 36.19 ± 0.01bA | |
| 20–30 | 23.82 ± 0.55aA | 44.43 ± 0.12dB | 26.99 ± 0.17aA | 0.19 ± 0.01bA | 0.33 ± 0.01aB | 0.03 ± 0.01bB | 21.15 ± 0.02aA | 42.59 ± 0.01cB | 23.49 ± 0.03aA | |
| 30–40 | 22.15 ± 0.35aA | 21.57 ± 0.04aA | 24.6 ± 0.48aA | 0.31 ± 0.01cA | 0.03 ± 0.01aA | 0.03 ± 0.01bA | 20.55 ± 0.01aA | 21.19 ± 0.00aA | 22.31 ± 0.01aA | |
| 40–50 | 23.90 ± 0.27aA | 27.28 ± 0.40bA | 25.60 ± 0.50aA | 0.15 ± 0.01bA | 0.03 ± 0.01aB | 0.03 ± 0.01aB | 19.99 ± 0.01aA | 20.66 ± 0.01aA | 21.49 ± 0.01aA | |
| 18 September | 0 –10 | 23.75 ± 0.35aA | 51.03 ± 0.52bC | 38.51 ± 0.28bB | 0.01 ± 0.02aA | 0.08 ± 0.08cB | 0.06 ± 0.07bB | 25.97 ± 5.34aA | 42.31 ± 16.01cB | 27.90 ± 8.46aA |
| 10–20 | 27.13 ± 0.65aA | 63.61 ± 0.01cC | 47.61 ± 0.30bB | 0.02 ± 0.03aA | 0.02 ± 0.02aA | 0.08 ± 0.06cB | 33.05 ± 9.34bA | 39.16 ± 17.72cB | 37.72 ± 12.24cB | |
| 20–30 | 25.06 ± 0.10aA | 50.955 ± 0.02bB | 54.35 ± 0.13cB | 0.03 ± 0.04bA | 0.11 ± 0.08dB | 0.06 ± 0.04bA | 22.92 ± 1.90aA | 36.99 ± 12.81bB | 38.53 ± 18.15cB | |
| 30–40 | 41.26 ± 0.15cB | 34.39 ± 0.01aA | 25.35 ± 0.29aA | 0.02 ± 0.03aA | 0.04 ± 0.03aB | 0.07 ± 0.11bC | 31.85 ± 7.71bA | 31.59 ± 8.13aA | 33.12 ± 8.73bA | |
| 40–50 | 31.13 ± 0.11bA | 33.14 ± 0.03aA | 25.43 ± 0.05aA | 0.04 ± 0.05bA | 0.07 ± 0.02bB | 0.04 ± 0.06aA | 33.08 ± 3.51bB | 34.20 ± 6.24cB | 24.84 ± 9.97aA | |
| 15 November | 0 –10 | 30.76 ± 0.56cB | 12.91 ± 0.22aA | 21.34 ± 0.42aA | 0.05 ± 0.01cB | 0.03 ± 0.04aA | 0.11 ± 0.07cC | 17.25 ± 10.92bA | 20.69 ± 9.41bB | 26.82 ± 5.85bC |
| 10–20 | 17.08 ± 0.30aA | 22.36 ± 0.37bB | 28.00 ± 0.29bC | 0.04 ± 0.01cA | 0.05 ± 0.02bA | 0.09 ± 0.04bB | 9.62 ± 4.24aA | 15.11 ± 4.80aA | 35.73 ± 9.73cB | |
| 20–30 | 14.14 ± 0.15aA | 21.05 ± 0.17bB | 13.94 ± 0.36aA | 0.02 ± 0.01aA | 0.05 ± 0.01bB | 0.10 ± 0.06cC | 11.90 ± 1.79aA | 14.55 ± 5.40aA | 11.24 ± 1.95aA | |
| 30–40 | 29.39 ± 0.54bB | 11.07 ± 0.24aA | 9.68 ± 0.32aA | 0.03 ± 0.02bA | 0.05 ± 0.03cB | 0.10 ± 0.02bC | 16.79 ± 10.23bA | 11.51 ± 6.26aA | 13.31 ± 4.25aA | |
| 40–50 | 23.84 ± 0.33bB | 11.23 ± 0.28aA | 17.13 ± 0.26bB | 0.03 ± 0.01aA | 0.03 ± 0.01aA | 0.07 ± 0.04aB | 15.59 ± 5.06bA | 12.32 ± 1.65aA | 27.18 ± 25.29bB | |
| Model | Sum of Squares | Freedom | Mean Square | F | Significance | |
|---|---|---|---|---|---|---|
| 1 | Regression | 632,256,641 | 10 | 63,225,664 | 2.319 | 0.013 |
| Residual | 7,062,498,135 | 259 | 27,268,333 | |||
| Total | 7,694,754,776 | 269 |
| Model | Nonstandard Coefficient | Standardization Coefficient | t | Significance | |
|---|---|---|---|---|---|
| (Constant) | 52,347.16 | 27,613.11 | 1.90 | 0.06 | |
| Particular year | −3899.58 | 1154.61 | −1.25 | −3.38 | 0.01 |
| Forest type | −1327.88 | 550.65 | −0.20 | −2.41 | 0.02 |
| Soil depth | 5.12 | 24.72 | 0.01 | 0.21 | 0.08 |
| pH value | −118.22 | 1006.91 | −0.01 | −0.12 | 0.19 |
| Humidity | −367.86 | 390.42 | −0.11 | −0.94 | 0.03 |
| Temperature | −24.02 | 78.97 | −0.05 | −0.30 | 0.04 |
| Litter N content | −3047.91 | 3006.83 | −0.22 | −1.01 | 0.31 |
| Litter C content | 255.93 | 165.93 | 1.21 | 1.54 | 0.12 |
| Litter CN ratio | −1617.78 | 569.09 | −2.82 | −2.84 | 0.01 |
| Soil organic matter content | 8.19 | 6.66 | 0.09 | 1.23 | 0.02 |
| Model | Sum of Squares | Freedom | Mean Square | F | Significance | |
|---|---|---|---|---|---|---|
| 1 | Regression | 0.681 | 10 | 0.068 | 8.488 | 0.001 |
| Residual | 2.079 | 259 | 0.008 | |||
| Total | 2.761 | 269 |
| Model | Nonstandard Coefficient Beta | Standardized Number Beta | t | Significance | |
|---|---|---|---|---|---|
| Particular year | 0.04 | 0.02 | 0.59 | 2.75 | 0.05 |
| Forest type | 0.02 | 0.01 | 0.12 | 2.54 | 0.02 |
| Soil depth | 0.01 | 0.01 | −0.11 | −2.82 | 0.07 |
| pH value | 0.02 | 0.02 | 0.09 | 1.11 | 0.27 |
| Humidity | 0.03 | 0.01 | 0.53 | 4.81 | 0.01 |
| Temperature | 0.00 | 0.00 | −0.03 | −1.23 | 0.02 |
| Litter N content | 0.01 | 0.05 | 0.05 | 0.25 | 0.80 |
| Litter C content | −0.01 | 0.00 | −1.65 | −2.32 | 0.02 |
| Litter CN ratio | 0.03 | 0.01 | 2.39 | 2.66 | 0.01 |
| Soil organic matter content | 0.00 | 0.00 | 0.16 | 2.33 | 0.02 |
| Model | Sum of Squares | Freedom | Mean Square | F | Significance | |
|---|---|---|---|---|---|---|
| 1 | Regression | 1,348,848.91 | 10 | 134,884.89 | 2.98 | 0.001 |
| Residual | 11,727,318.02 | 259 | 45,279.22 | |||
| Total | 13,076,166.93 | 269 |
| Model | Nonstandard Coefficient Beta | Standardized Number Beta | t | Significance | |
|---|---|---|---|---|---|
| Sampling time | 13.24 | 47.05 | 0.10 | 0.28 | 0.08 |
| Forest type | 4.79 | 22.44 | 0.02 | 0.21 | 0.08 |
| Soil depth | 1.74 | 1.01 | 0.11 | 1.72 | 0.08 |
| pH value | −39.97 | 41.03 | −0.08 | −0.97 | 0.33 |
| Humidity | 54.79 | 15.91 | 0.41 | 3.44 | 0.01 |
| Temperature | −2.95 | 3.22 | −0.14 | −0.92 | 0.03 |
| Litter N content | −30.05 | 122.53 | −0.05 | −0.25 | 0.08 |
| Litter C content | −3.92 | 6.76 | −0.45 | −0.58 | 0.06 |
| Litter CN ratio | 12.71 | 23.19 | 0.54 | 0.55 | 0.05 |
| Soil organic matter content | −0.29 | 0.27 | −0.08 | −1.06 | 0.02 |
| Sampling Time | Soil Depth | Forest Type | Total N | NNH4N | NNO3N | pH | Temperature | Humidity | Litter C Content | Litter N Content | Litter C/N | Soil Organic Matter Content | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sampling time | 1 | 0 | 0 | 0.394 ** | 0.11 | 0.44 ** | −0.164 ** | 0.251 ** | 0.258 ** | −0.957 ** | 0.919 ** | −0.973 ** | 0.052 |
| Soil depth | 1 | 0 | −0.001 | −0.228 ** | 0.06 | 0.248 ** | −0.057 | −0.165 ** | −0.76 | −0.62 | −0.71 | −0.359 ** | |
| Forest type | 1 | 0.21 | 0.47 ** | 0.006 | −0.285 ** | 0 | −0.047 | −0.196 ** | 0.124 * | −0.158 ** | 0.166 ** | ||
| Total N | 1 | 0.037 | −0.005 | −0.029 | 0.078 | 0.008 | −0.120 * | 0.093 | −0.129 * | 0.066 | |||
| NNH4N | 1 | −0.037 | −0.222 ** | 0.236 ** | 0.388 ** | −0.002 | −0.013 | −0.005 | 0.264 ** | ||||
| NNO3N | 1 | −0.140 * | 0.157 ** | 0.265 ** | −0.038 | 0.024 | −0.044 | −0.012 | |||||
| pH value | 1 | −0.246 ** | −0.413 ** | 0.237 ** | −0.280 ** | 0.229 ** | −0.531 ** | ||||||
| Temperature | 1 | 0.819 ** | −0.185 ** | 0.072 | −0.266 ** | 0.205 ** | |||||||
| Humidity | 1 | −0.197 ** | 0.130 * | −0.255 ** | 0.233 ** | ||||||||
| Litter C content | 1 | −0.941 ** | 0.993 ** | −0.086 | |||||||||
| Litter N content | 1 | −0.934 ** | 0.119 | ||||||||||
| Litter C/N | 1 | −0.085 | |||||||||||
| Soil organic matter content | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Chen, L.; Duan, W.; Bai, Y.; Li, X. Effects of Litter Decomposition on Soil N in Picea mongolica Forest at Different Forest Ages. Forests 2022, 13, 520. https://doi.org/10.3390/f13040520
Liu Y, Chen L, Duan W, Bai Y, Li X. Effects of Litter Decomposition on Soil N in Picea mongolica Forest at Different Forest Ages. Forests. 2022; 13(4):520. https://doi.org/10.3390/f13040520
Chicago/Turabian StyleLiu, Yunchao, Lixin Chen, Wenbiao Duan, Yongan Bai, and Xiaolan Li. 2022. "Effects of Litter Decomposition on Soil N in Picea mongolica Forest at Different Forest Ages" Forests 13, no. 4: 520. https://doi.org/10.3390/f13040520
APA StyleLiu, Y., Chen, L., Duan, W., Bai, Y., & Li, X. (2022). Effects of Litter Decomposition on Soil N in Picea mongolica Forest at Different Forest Ages. Forests, 13(4), 520. https://doi.org/10.3390/f13040520