
Elicitation of Secondary Metabolites in Aquilaria malaccensis Lam. Callus Culture by Crude Mycelial Extract of Fusarium solani and Methyl Jasmonate

 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
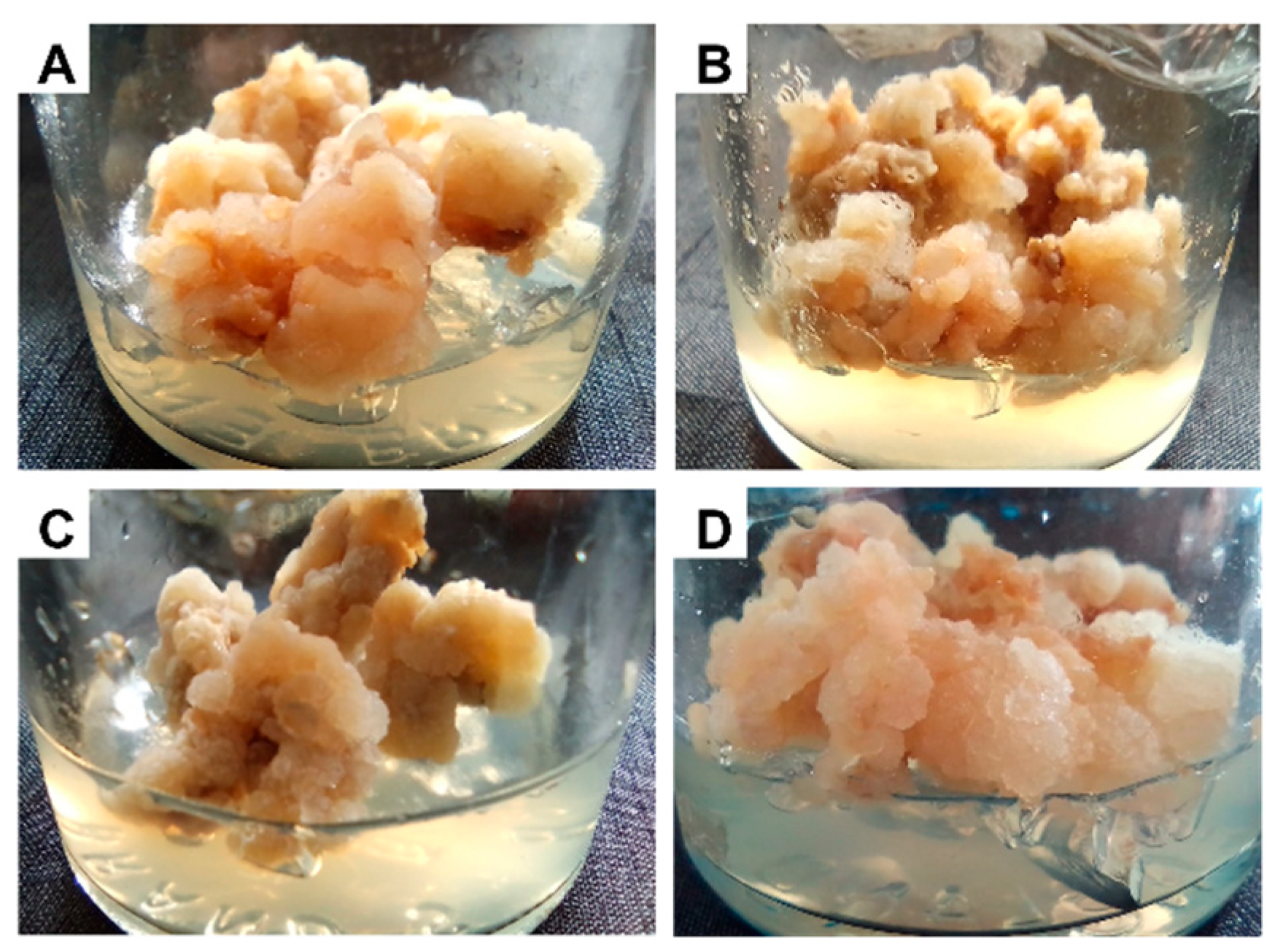
2.1. Generation of Callus
2.2. Preparation of MeJA Solution and Crude Mycelial Extract of F. solani
2.3. Callus Treatment
2.4. Estimation of Growth Rate and Doubling Time of Callus
2.5. Measurement of Cell Viability
2.6. Sample Extraction and Gas Chromatography–Mass Spectrometry Analysis
3. Results
3.1. The Growth Rate of Callus Treated with Elicitors or MeJA Was Less Than That of Untreated Callus
3.2. The Viability of A. malaccensis Callus Cells in Culture Decreased after Treatment with Elicitors
3.3. Chromone Derivatives Were Detected in Samples Treated with Methyl Jasmonate, and the Elicitors Derived from F. solani Strain GSL2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Turjaman, M.; Hidayat, A.; Santoso, E. Development of Agarwood Induction Technology Using Endophytic Fungi. In Agarwood: Science Behind the Fragrance; Mohamed, R., Ed.; Springer: Singapore, 2016; pp. 57–71. [Google Scholar]
- Maman, T.; Asep, H. Agarwood-planted tree inventory in Indonesia. IOP Conf. Ser. Earth Environ. Sci. 2017, 54, 012062. [Google Scholar]
- Nasution, A.A.; Siregar, U.J.; Miftahudin; Turjaman, M. Identification of chemical compounds in agarwood-producing species Aquilaria malaccensis and Gyrinops versteegii. J. For. Res. 2020, 31, 1371–1380. [Google Scholar] [CrossRef]
- Du, T.-Y.; Dao, C.-J.; Mapook, A.; Stephenson, S.L.; Elgorban, A.M.; Al-Rejaie, S.; Suwannarach, N.; Karunarathna, S.C.; Tibpromma, S. Diversity and biosynthetic activities of agarwood associated fungi. Diversity 2022, 14, 211. [Google Scholar] [CrossRef]
- Faizal, A.; Hermawaty, D.; Junita, E.; Rahmawati, A.; Azar, A.W.P.; Makajanma, M.M.; Turjaman, M. Evaluation of biotic and abiotic stressors to artificially induce agarwood production in Gyrinops versteegii (Gilg.) Domke seedlings. Symbiosis 2022, 86, 229–239. [Google Scholar] [CrossRef]
- Subasinghe, S.M.C.U.P.; Hitihamu, H.I.D.; Fernando, K.M.E.P. Use of two fungal species to induce agarwood resin formation in Gyrinops walla. J. For. Res. 2019, 30, 721–726. [Google Scholar] [CrossRef]
- Faizal, A.; Azar, A.W.P.; Turjaman, M.; Esyanti, R.R. Fusarium solani induces the formation of agarwood in Gyrinops versteegii (Gilg.) Domke branches. Symbiosis 2020, 81, 15–23. [Google Scholar] [CrossRef]
- Chen, X.; Sui, C.; Liu, Y.; Yang, Y.; Liu, P.; Zhang, Z.; Wei, J. Agarwood formation induced by fermentation liquid of Lasiodiplodia theobromae, the dominating fungus in wounded wood of Aquilaria sinensis. Curr. Microbiol. 2017, 74, 460–468. [Google Scholar] [CrossRef]
- Chen, X.; Liu, Y.; Yang, Y.; Feng, J.; Liu, P.; Sui, C.; Wei, J. Trunk surface agarwood-inducing technique with Rigidoporus vinctus: An efficient novel method for agarwood production. PLoS ONE 2018, 13, e0198111. [Google Scholar] [CrossRef]
- Naef, R. The volatile and semi-volatile constituents of agarwood, the infected heartwood of Aquilaria species: A review. Flavour Frag. J. 2011, 26, 73–87. [Google Scholar] [CrossRef]
- Chen, H.; Wei, J.; Yang, J.; Zhang, Z.; Yang, Y. Chemical constituents of agarwood originating from the endemic genus Aquilaria plants. Chem. Biodivers. 2012, 9, 236–250. [Google Scholar] [CrossRef]
- Wang, S.; Yu, Z.; Wang, C.; Wu, C.; Guo, P.; Wei, J. Chemical constituents and pharmacological activity of agarwood and Aquilaria plants. Molecules 2018, 23, 342. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-y.; Wei, J.-h.; Gao, Z.-h.; Zhang, Z.; Lyu, J.-c. A review of quality assessment and grading for agarwood. Chin. Herb. Med. 2017, 9, 22–30. [Google Scholar] [CrossRef]
- Mohamed, R.; Lee, S.Y. Keeping up appearances: Agarwood grades and quality. In Agarwood; Mohamed, R., Ed.; Springer: Singapore, 2016; pp. 149–167. [Google Scholar]
- Okudera, Y.; Ito, M. Production of agarwood fragrant constituents in Aquilaria calli and cell suspension cultures. Plant Biotechnol. 2009, 26, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Munasinghe, S.; Somaratne, S.; Weerakoon, S.; Ranasinghe, C. Sustainable utilization of Gyrinops walla Gaetner: In vitro production of sesquiterpenes by chemical and biological elicitation. J. Genet. Eng. Biotechnol. 2021, 19, 134. [Google Scholar] [CrossRef] [PubMed]
- Faizal, A.; Esyanti, R.R.; Adn’ain, N.; Rahmani, S.; Azar, A.W.P.; Iriawati; Turjaman, M. Methyl jasmonate and crude extracts of Fusarium solani elicit agarwood compounds in shoot culture of Aquilaria malaccensis Lamk. Heliyon 2021, 7, e06725. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, S.; Mohamed, R. Crude extract of Trichoderma elicits agarwood substances in cell suspension culture of the tropical Tree, Aquilaria malaccensis Lam. Turk. J. Agric. For. 2015, 39, 163–173. [Google Scholar] [CrossRef]
- Esyanti, R.R.; Fadholi, M.; Rizki, R.M.; Faizal, A. Shoot multiplication and growth rates of Aquilaria malaccensis Lamk. shoot cultures in temporary immersion system (TIS)-RITA® and bubble column bioreactors. Pak. J. Bot. 2019, 51, 1317–1321. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Dong, X.; Gao, B.; Feng, Y.; Liu, X.; Wang, J.; Wang, J.; Tu, P.; Wang, X.; Shi, S. Production of 2-(2-phenylethyl)chromones in Aquilaria sinensis calli under different treatments. Plant Cell Tissue Organ Cult. 2018, 135, 53–62. [Google Scholar] [CrossRef]
- Godoy-Hernández, G.; Vázquez-Flota, F.A. Growth measurements: Estimation of cell division and cell expansion. In Plant Cell Culture Protocols; Loyola-Vargas, V.M., Ochoa-Alejo, N., Eds.; Humana Press: Totowa, NJ, USA, 2012; pp. 41–48. [Google Scholar]
- Huang, J.-Q.; Liao, Y.-C.; Chen, H.-J.; Zhang, Z. Chemical solution is an efficient method to induce the formation of 2-(2-phenylethyl) chromone derivatives in Aquilaria sinensis. Phytochem. Lett. 2017, 19, 64–70. [Google Scholar] [CrossRef]
- Qi, S.-Y.; He, M.-L.; Lin, L.-D.; Zhang, C.-H.; Hu, L.-J.; Zhang, H.-Z. Production of 2-(2-phenylethyl) chromones in cell suspension cultures of Aquilaria sinensis. Plant Cell Tissue Organ Cult. 2005, 83, 217–221. [Google Scholar] [CrossRef]
- Bourgaud, F.; Gravot, A.; Milesi, S.; Gontier, E. Production of plant secondary metabolites: A historical perspective. Plant Sci. 2001, 161, 839–851. [Google Scholar] [CrossRef]
- Künstler, A.; Bacsó, R.; Gullner, G.; Hafez, Y.M.; Király, L. Staying alive—Is cell death dispensable for plant disease resistance during the hypersensitive response? Physiol. Mol. Plant Pathol. 2016, 93, 75–84. [Google Scholar] [CrossRef]
- Kumar, J.; Ramlal, A.; Kumar, K.; Rani, A.; Mishra, V. Signaling pathways and downstream effectors of host innate immunity in plants. Int. J. Mol. Sci. 2021, 22, 9022. [Google Scholar] [CrossRef]
- Chamkhi, I.; Benali, T.; Aanniz, T.; El Menyiy, N.; Guaouguaou, F.-E.; El Omari, N.; El-Shazly, M.; Zengin, G.; Bouyahya, A. Plant-microbial interaction: The mechanism and the application of microbial elicitor induced secondary metabolites biosynthesis in medicinal plants. Plant Physiol. Biochem. 2021, 167, 269–295. [Google Scholar] [CrossRef]
- Ito, M.; Okimoto, K.-I.; Yagura, T.; Honda, G.; Kiuchi, F.; Shimada, Y. Induction of sesquiterpenoid production by methyl jasmonate in Aquilaria sinensis cell suspension culture. J. Essent. Oil Res. 2005, 17, 175–180. [Google Scholar] [CrossRef]
- Sen, S.; Dehingia, M.; Talukdar, N.C.; Khan, M. Chemometric analysis reveals links in the formation of fragrant bio-molecules during agarwood (Aquilaria malaccensis) and fungal interactions. Sci. Rep. 2017, 7, 44406. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Arif, Y.; Miszczuk, E.; Bajguz, A.; Hayat, S. Specific roles of lipoxygenases in development and responses to stress in plants. Plants 2022, 11, 979. [Google Scholar] [CrossRef]
- Carvalhais, L.C.; Schenk, P.M.; Dennis, P.G. Jasmonic acid signalling and the plant holobiont. Curr. Opin. Microbiol. 2017, 37, 42–47. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Fernandes, Â.; Calhelha, R.C.; Rouphael, Y.; Petrović, J.; Soković, M.; Ferreira, I.C.F.R.; Barros, L. Antimicrobial properties, cytotoxic effects, and fatty acids composition of vegetable oils from purslane, linseed, luffa, and pumpkin seeds. Appl. Sci. 2021, 11, 5738. [Google Scholar] [CrossRef]
- Faizal, A.; Esyanti, R.R.; Aulianisa, E.N.; Iriawati; Santoso, E.; Turjaman, M. Formation of agarwood from Aquilaria malaccensis in response to inoculation of local strains of Fusarium solani. Trees 2017, 31, 189–197. [Google Scholar] [CrossRef]
- Cui, J.; Guo, S.; Fu, S.; Xiao, P.; Wang, M. Effects of inoculating fungi on agilawood formation in Aquilaria sinensis. Chin. Sci. Bull. 2013, 58, 3280–3287. [Google Scholar] [CrossRef] [Green Version]
- Miras-Moreno, B.; Sabater-Jara, A.B.; Pedreño, M.A.; Almagro, L. Bioactivity of phytosterols and their production in plant in vitro cultures. J. Agric. Food. Chem. 2016, 64, 7049–7058. [Google Scholar] [CrossRef]
- Skanda, S.; Vijayakumar, B.S. Antioxidant and antibacterial potential of crude extract of soil fungus Periconia sp. (SSS-8). Arab. J. Sci. Eng. 2022, 47, 6707–6714. [Google Scholar] [CrossRef]
- Castro, A.H.F.; Coimbra, M.C.; Crema, C.R.N.; de Oliveira Thomasi, R.M.; Cardoso, V.R. Elicitors as a biotechnological tool for in vitro production of bioactive phenolic compounds. In Exploring Plant Cells for the Production of Compounds of Interest; Malik, S., Ed.; Springer International Publishing: Cham, Switzerland, 2021; pp. 195–226. [Google Scholar]
- Tuladhar, P.; Sasidharan, S.; Saudagar, P. Role of phenols and polyphenols in plant defense response to biotic and abiotic stresses. In Biocontrol Agents and Secondary Metabolites; Jogaiah, S., Ed.; Woodhead Publishing: Sawston, UK, 2021; pp. 419–441. [Google Scholar]
- Jong, P.L.; Tsan, P.; Mohamed, R. Gas chromatography-mass spectrometry analysis of agarwood extracts from mature and juvenile Aquilaria malaccensis. Int. J. Agric. Biol. 2014, 16, 644–648. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Treatment | Specific Growth Rate (g/Day) | Doubling Time (Day) |
|---|---|---|
| GSL1 | 0.15 ± 0.047 | 4.98 ± 1.625 |
| GSL2 | 0.16 ± 0.048 | 4.56 ± 1.426 |
| MeJA | 0.16 ± 0.040 | 4.68 ± 1.308 |
| Control | 0.17 ± 0.044 | 4.31 ± 1.125 |
| Days Post Treatment | Treatment | |||
|---|---|---|---|---|
| GSL1 | GSL2 | MeJA | Control | |
| 5 | 0.59 ± 0.131 | 0.80 ± 0.069 | 0.59 ± 0.051 | 0.81 ± 0.108 |
| 10 | 0.69 ± 0.090 | 0.63 ± 0.092 | 0.51 ± 0.038 | 0.95 ± 0.178 |
| 15 | 0.83 ± 0.059 | 0.86 ± 0.044 | 0.72 ± 0.054 | 1.05 ± 0.185 |
| 20 | 0.92 ± 0.075 | 0.99 ± 0.298 | 0.75 ± 0.088 | 1.16 ± 0.366 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faizal, A.; Esyanti, R.R.; Utami, N.P.; Azar, A.W.P.; Hermawaty, D.; Setiyobudi, T.; Martin, A.F.; Hapsari, B.W.; Turjaman, M. Elicitation of Secondary Metabolites in Aquilaria malaccensis Lam. Callus Culture by Crude Mycelial Extract of Fusarium solani and Methyl Jasmonate. Forests 2023, 14, 48. https://doi.org/10.3390/f14010048
Faizal A, Esyanti RR, Utami NP, Azar AWP, Hermawaty D, Setiyobudi T, Martin AF, Hapsari BW, Turjaman M. Elicitation of Secondary Metabolites in Aquilaria malaccensis Lam. Callus Culture by Crude Mycelial Extract of Fusarium solani and Methyl Jasmonate. Forests. 2023; 14(1):48. https://doi.org/10.3390/f14010048
Chicago/Turabian StyleFaizal, Ahmad, Rizkita Rachmi Esyanti, Nadia Puji Utami, Alda Wydia Prihartini Azar, Dina Hermawaty, Titis Setiyobudi, Andri Fadillah Martin, Betalini Widhi Hapsari, and Maman Turjaman. 2023. "Elicitation of Secondary Metabolites in Aquilaria malaccensis Lam. Callus Culture by Crude Mycelial Extract of Fusarium solani and Methyl Jasmonate" Forests 14, no. 1: 48. https://doi.org/10.3390/f14010048