
Responses of the Crown Shape of Larix kaempferi Plantations to Site Index in Subtropical Areas of China
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area Description
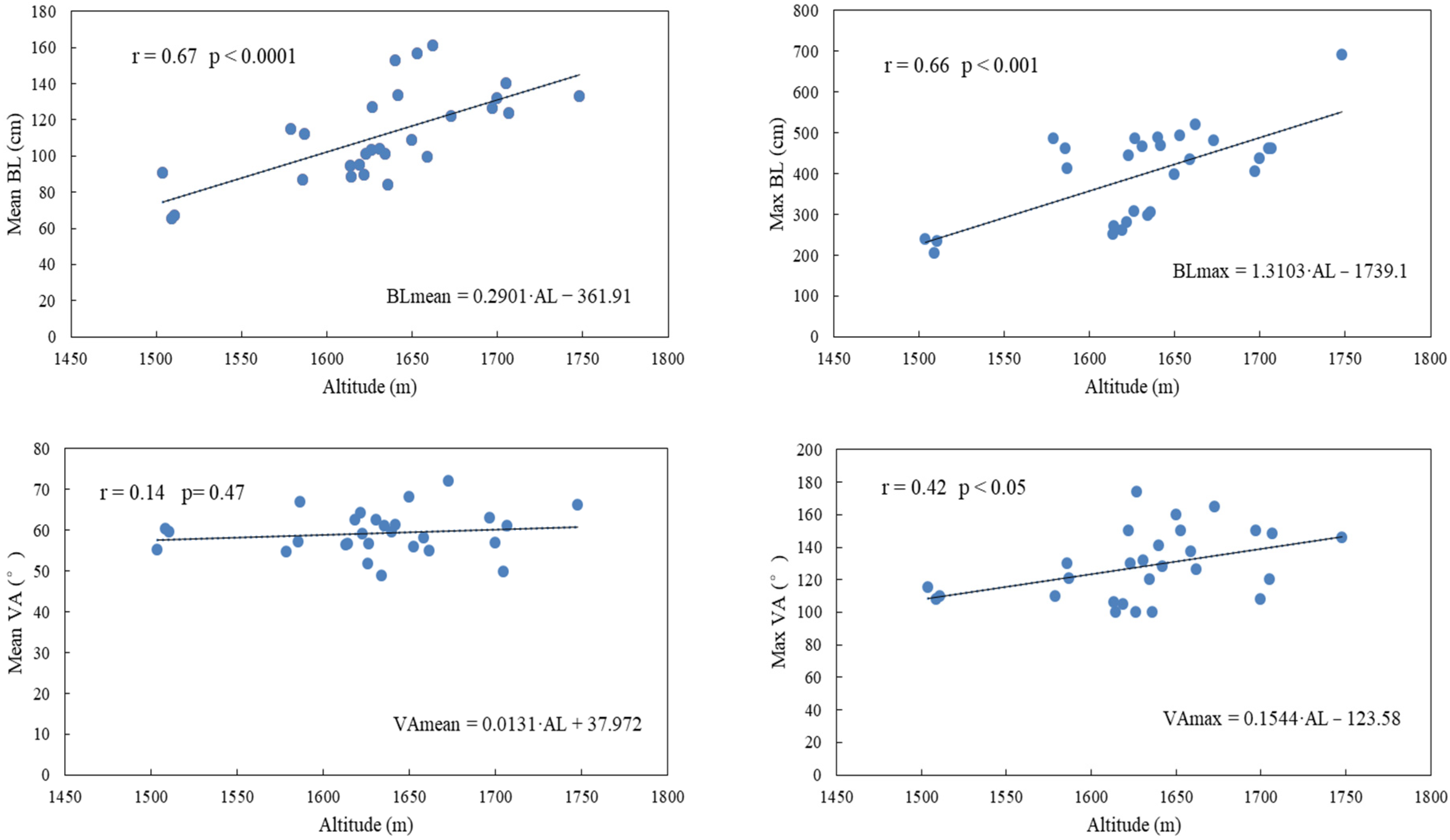
2.2. Data Collection
2.3. Basic Crown Shape Model for L. kaempferi
2.4. Inclusion of Site Index
3. Results
3.1. Crown Shape Model Development by Incorporating Site Index
3.2. Nonlinear Mixed-Effect Crown Shape Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chmura, D.J.; Tjoelker, M.J. Leaf traits in relation to crown development, light interception and growth of elite families of loblolly and slash pine. Tree Physiol. 2008, 28, 729–742. [Google Scholar] [CrossRef] [PubMed]
- Alcorn, P.J.; Forrester, D.I.; Smith, R.G.B.; Thomas, D.S.; James, R.N.; Nicotra, A.B.; Bauhus, J. Crown structure and vertical foliage distribution in 4-year-old plantation-grown Eucalyptus pilularis and Eucalyptus cloeziana. Trees 2013, 27, 555–566. [Google Scholar] [CrossRef]
- Condés, S.; Aguirre, A.; del Río, M. Crown plasticity of five pine species in response to competition along an aridity gradient. For. Ecol. Manag. 2020, 473, 118302. [Google Scholar] [CrossRef]
- Barbeito, I.; Collet, C.; Ningre, F. Crown responses to neighbor density and species identity in a young mixed deciduous stand. Trees 2014, 28, 1751–1765. [Google Scholar] [CrossRef]
- Mário, D.J.; Marcos, F.N.; Julio, E.A. Influence of crown thinning on radial growth pattern of Pinus taeda in southern Brazil. New For. 2018, 50, 437–454. [Google Scholar] [CrossRef]
- Horn, H.S. The Adaptive Geometry of Trees; Princeton University Press: Princeton, NJ, USA, 1971. [Google Scholar]
- Ishii, H.; Clement, J.P.; Shaw, D.C. Branch growth and crown form in old coastal Douglas-fir. For. Ecol. Manag. 2000, 131, 81–91. [Google Scholar] [CrossRef]
- Trincado, G.; Burkhart, H.E. A framework for modeling the dynamics of first order branches and spatial distribution of knots in loblolly pine trees. Can. J. For. Res. 2009, 39, 566–579. [Google Scholar] [CrossRef]
- Burkhart, H.E.; Tomé, M. Modeling Forest Trees and Stands; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; New York, NY, USA; London, UK, 2012; pp. 25–26. [Google Scholar]
- Schröter, M.; Härdtle, W.; Oheimb, G.V. Crown plasticity and neighborhood interactions of European beech (Fagussylvatica L.) in an old-growth forest. Eur. J. For. Res. 2012, 131, 787–798. [Google Scholar] [CrossRef]
- Bergès, L.; Nepveu, G.; Franc, A. Effects of ecological factors on radial growth and wood density components of Sessile oak (Quercus petraea Liebl.) in northern France. For. Ecol. Manag. 2008, 255, 567–579. [Google Scholar] [CrossRef]
- Macdonald, E.; Hubert, J. Effect of crown class and habitat type on climate–growth relationships of ponderosa pine and Douglas-fir. Forestry 2002, 75, 107–138. [Google Scholar] [CrossRef]
- Cattaneo, N.; Bravo-Oviedo, A.; Bravo, F. Analysis of tree interactions in a mixed Mediterranean pine stand using competition indices. Eur. J. For. Res. 2018, 137, 109–120. [Google Scholar] [CrossRef]
- Fichtner, A.; Sturm, K.; Rickert, C.; von Oheimb, G.; Härdtle, W. Crown size-growth relationships of European beech (Fagus sylvatica L.) are driven by the interplay of disturbance intensity and inter-specific competition. For. Ecol. Manag. 2013, 302, 178–184. [Google Scholar] [CrossRef]
- Linnell Nemec, A.F.; Parish, R.; Goudie, J.W. Modelling number, vertical distribution, and size of live branches on coniferous tree species in British Columbia. Can. J. For. Res. 2012, 42, 1072–1090. [Google Scholar] [CrossRef]
- Harrington, T.B.; Harrington, C.A.; DeBell, D.S. Effects of planting spacing and site quality on 25-year growth and mortality relationships of Douglas-fir (Pseudotsuga menziesii var. menziesii). For. Ecol. Manag. 2009, 258, 18–25. [Google Scholar] [CrossRef]
- Attocchi, G.; Skovsgaard, J.P. Crown radius of pedunculate oak (Quercus robur L.) depending on stem size, stand density and site productivity. Scand. J. For. Res. 2015, 30, 289–303. [Google Scholar] [CrossRef]
- Paulo, M.J.; Stein, A.; Tomé, M.A. A spatial statistical analysis of cork oak competition in two Portuguese silvopastoral systems. Can. J. For. Res. 2002, 32, 1893–1903. [Google Scholar] [CrossRef]
- Longuetaud, F.; Piboule, A.; Wernsdörfer, H.; Collet, C. Crown plasticity reduces inter-tree competition in a mixed broadleaved forest. Eur. J. For. Res. 2013, 132, 621–634. [Google Scholar] [CrossRef]
- Marron, N.; Priault, P.; Gana, C.; Gérant, D.; Epron, D. Prevalence of interspecific competition in a mixed poplar/black locust plantation under adverse climate conditions. Ann. For. Sci. 2018, 75, 23. [Google Scholar] [CrossRef]
- Brisson, J. Neighborhood competition and crown asymmetry in Acer saccharum. Can. J. For. Res. 2001, 31, 2151–2159. [Google Scholar] [CrossRef]
- Valentine, H.T.; Mäkelä, A.; Green, E.J. Models relating stem growth to crown length dynamics: Application to loblolly pine and Norway spruce. Trees 2012, 26, 469–478. [Google Scholar] [CrossRef]
- Sumida, A.; Miyaura, T.; Torii, H. Relationships of tree height and diameter at breast height revisited: Analyses of stem growth using 20-year data of an even-aged Chamaecyparis obtusa stand. Tree Physiol. 2013, 33, 106–118. [Google Scholar] [CrossRef]
- Kaiser, E.H. The Influence of Stand Structure and Topography on Growth, Leaf Area and Efficiency of Young Douglas-Fir in the Oregon Coast Range; Oregon State University: Corvallis, OR, USA, 1986. [Google Scholar]
- Umeki, K. Modeling the relationship between the asymmetry in crown display and local environment. Ecol. Model. 1995, 82, 11–20. [Google Scholar] [CrossRef]
- Uemura, A.; Harayama, H.; Koike, N. Coordination of crown structure, leaf plasticity and carbon gain within the crowns of three winter-deciduous mature trees. Tree Physiol. 2006, 26, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Gavrikov, V.L.; Grabarnik, P.Y.; Stoyan, D. Trunk–top relations in a Siberian pine forest. Biom. J. 1993, 35, 487–498. [Google Scholar] [CrossRef]
- Skatter, S.; Kucera, B. The cause of the prevalent directions of the spiral grain patterns in conifers. Trees 1998, 12, 265–273. [Google Scholar] [CrossRef]
- Takenaka, A. Shoot growth responses to light microenvironment and correlative inhibition in tree seedlings under a forest canopy. Tree Physiol. 2000, 20, 987–991. [Google Scholar] [CrossRef]
- Rudnicki, M.; Lieffers, V.J.; Silins, U. Stand structure governs the crown collisions of lodgepole pine. Can. J. For. Res. 2003, 33, 1238–1244. [Google Scholar] [CrossRef]
- Ancelin, P.; Courbaud, B.; Fourcaud, T. Development of an individual tree-based mechanical model to predict wind damage within forest stands. For. Ecol. Manag. 2004, 203, 101–121. [Google Scholar] [CrossRef]
- Lang, A.C.; Hardtle, W.; Bruelheide, H.; Geißler, C.; Nadrowski, K.; Schuldt, A.; Yu, M.; von Oheimb, G. Tree morphology responds to neighbourhood competition and slope in species-rich forests of subtropical China. For. Ecol. Manag. 2010, 260, 1708–1715. [Google Scholar] [CrossRef]
- Vovides, A.G.; Berger, U.; Grueters, U.; Guevara, R.; Pommerening, A.; Lara-Domínguez, A.L.; López-Portillo, J. Change in drivers of mangrove crown displacement along a salinity stress gradient. Funct. Ecol. 2018, 32, 2753–2765. [Google Scholar] [CrossRef]
- Cocozza, C.; Palombo, C.; Tognetti, R.; La Porta, N.; Anichini, M.; Giovannelli, A.; Emiliani, G. Monitoring intra-annual dynamics of wood formation with microcores and dendrometers in Picea abies at two different altitudes. Tree Physiol. 2016, 36, 832–846. [Google Scholar] [CrossRef] [PubMed]
- Danby, R.K.; Hik, D.S. Responses of white spruce (Picea glauca) to experimental warming at a subarctic alpine treeline. Global Chang. Biol. 2007, 13, 437–451. [Google Scholar] [CrossRef]
- Larsary, M.K.; Pourbabaei, H.; Sanaei, A.; Salehi, A.; Yousefpour, R.; Ali, A. Tree-size dimension inequality shapes aboveground carbon stock across temperate forest strata along environmental gradients. For. Ecol. Manag. 2021, 496, 119482. [Google Scholar] [CrossRef]
- Kübler, D.; Hildebrandt, P.; Günter, S. Effects of silvicultural treatments and topography on individual tree growth in a tropical mountain forest in Ecuador. For. Ecol. Manag. 2020, 457, 117726. [Google Scholar] [CrossRef]
- Dickmann, D.I.; Michael, D.A.; Isebrands, J.G.; Westin, S. Effects of leaf display on light interception and apparent photosynthesis in two contrasting Populus cultivars during their second growing season. Tree Physiol. 1991, 7, 7–20. [Google Scholar] [CrossRef]
- Wykoff, W. A basal area increment model for individual conifers in the northern Rocky Mountains. For. Sci. 1990, 36, 1077–1104. [Google Scholar]
- Weiskittel, A.R.; Crookston, N.L.; Radtke, P.J. Linking climate, gross primary productivity, and site index across forests of the western United States. Can. J. For. Res. 2011, 41, 1710–1721. [Google Scholar] [CrossRef]
- Bravo-Oviedo, A.; Tomé, M.; Bravo, F.; Montero, G.; del Río, M. Dominant height growth equations including site attributes in the generalized algebraic difference approach. Can. J. For. Res. 2008, 38, 2348–2358. [Google Scholar] [CrossRef]
- Shelbume, V.B.; Hedden, R.L. Effect of stem height, dominance class, and site quality on sapwood permeability in loblolly pine, (Pinus tuedu L.). For. Ecol. Manag. 1996, 83, 163–169. [Google Scholar] [CrossRef]
- Hann, D.W. An adjustable predictor of crown profile for stand-grown Douglas-fir trees. For. Sci. 1999, 45, 217–225. [Google Scholar]
- Crecente-Campo, F.; Marshall, P.; Lemay, V.; Diéguez-Aranda, U. A crown profile model for Pinus radiata D. Don in northwestern Spain. For. Ecol. Manag. 2009, 257, 2370–2379. [Google Scholar] [CrossRef]
- Ferrarese, J.; Affleck, D.; Seielstad, C. Conifer crown profile models from terrestrial laser scanning. Silva Fenn. 2015, 49, 1. [Google Scholar] [CrossRef]
- Gao, H.; Chen, D.; Sun, X.; Zhang, S. Modelling the responses of crown profiles to competition in terms of different tree classes and directions in two coniferous tree species in northeastern China. Eur. J. For. Res. 2021, 140, 159–174. [Google Scholar] [CrossRef]
- Baldwin, V.C.; Jr Peterson, K.D. Predicting the crown shape of loblolly pine trees. Can. J. For. Res. 1997, 77, 407–415. [Google Scholar] [CrossRef]
- Dong, C.; Wu, B.; Wang, C.; Guo, Y.; Han, Y. Study on crown profile models for Chinese fir (Cunninghamia lanceolata) in Fujian Province and its visualization simulation. Scand. J. For. Res. 2016, 31, 302–313. [Google Scholar] [CrossRef]
- Seidel, D.; Leuschner, C.; Müller, A.; Krause, B. Crown plasticity in mixed forests—Quantifying asymmetry as a measure of competition using terrestrial laser scanning. For. Ecol. Manag. 2011, 261, 2123–2132. [Google Scholar] [CrossRef]
- Sievnen, R.; Raumonen, P.; Perttunen, J.; Nikinmaa, E.; Kaitaniemi, P. A study of crown development mechanisms using a shoot-based tree model and segmented terrestrial laser scanning data. Ann. Bot. 2018, 3, 3. [Google Scholar] [CrossRef]
- Mendez-Toribio, M.; Meave, J.A.; Zermeno-Hernandez, I.; Ibarra-Manriquez, G. Effects of slope aspect and topographic position on environmental variables, disturbance regime and tree community attributes in a seasonal tropical dry forest. J. Veg. Sci. 2016, 27, 1094–1103. [Google Scholar] [CrossRef]
- Getzin, S.; Wiegand, K. Asymmetric tree growth at the stand level: Random crown patterns and the response to slope. For. Ecol. Manag. 2007, 242, 165–174. [Google Scholar] [CrossRef]
- Valinger, E. Effects of wind sway on stem form and crown development of Scots pine (Pinus sylvestris L.). Aust. For. 1992, 55, 15–21. [Google Scholar] [CrossRef]
- Metz, J.; Seidel, D.; Schall, P.; Scheffer, D.; Schulze, E.D.; Ammer, C. Crown modeling by terrestrial laser scanning as an approach to assess the effect of aboveground intra- and interspecific competition on tree growth. For. Ecol. Manag. 2013, 310, 275–288. [Google Scholar] [CrossRef]
- Shen, J.; Chen, D.; Hong, Y.; Sun, X.; Zhang, S. Prediction on potential distribution areas of Larix kaempferi in China based on MaxEnt model. J. Plant Resour. Environ. 2019, 28, 19–25, (In Chinese with English abstract). [Google Scholar]
- Gao, H.; Bi, H.; Li, F. Modelling conifer crown profiles as nonlinear conditional quantiles: An example with planted Korean pine in northeast China. For. Ecol. Manag. 2017, 398, 101–115. [Google Scholar] [CrossRef]
- Ma, Y.; Ai, X.; Song, E. Compiling the site index table of Japanese larch in Changlinggang forest farm. J. Hubei Inst. Natl. 2001, 19, 24–27, (In Chinese with English abstract). [Google Scholar]
- Wang, C.; Zhang, Z.; Zhang, J. The effect of terrain factors on rice production: A case study in Hunan Province. J. Geogr. Sci. 2019, 29, 287–305. [Google Scholar] [CrossRef]
- Antos, J.A.; Parish, R.; Nigh, G.D. Effects of neighbours on crown length of Abies lasiocarpa and Picea engelmannii in two old-growth stands in British Columbia. Can. J. For. Res. 2010, 40, 638–647. [Google Scholar] [CrossRef]
- Medhurst, J.; Ottenschlaeger, M.; Wood, M.; Harwood, C.; Beadle, C.; Valencia, J.C. Stem eccentricity, crown dry mass distribution, and longitudinal growth strain of plantation-grown Eucalyptus nitens after thinning. Can. J. For. Res. 2011, 41, 2209–2218. [Google Scholar] [CrossRef]
- Davies, O.; Pommerening, A. The contribution of structural indices to the modelling of Sitka spruce (Picea sitchensis) and birch (Betula spp.) crowns. For. Ecol. Manag. 2008, 256, 68–77. [Google Scholar] [CrossRef]
- Kahriman, A.; Şahin, A.; Sönmez, T.; Yavuz, M. A novel approach to selecting a competition index: The effect of competition on individual tree diameter growth of Calabrian pine. Can. J. For. Res. 2018, 48, 1217–1226. [Google Scholar] [CrossRef]
- Jullien, D.; Widmann, R.; Loup, C.; Thibaut, B. Relationship between tree morphology and growth stress in mature European beech stands. Ann. For. Sci. 2013, 70, 133–142. [Google Scholar] [CrossRef]
- Seitz, S.; Goebes, P.; Kühn, P.; Schmidt, K.; Song, Z.; Scholten, T. Effects of soil fertility and topography on tree growth in subtropical forest ecosystems. Geophys. Res. Abstr. 2016, 18, EPSC2016-1536. [Google Scholar]
- Letts, M.G.; Nakonechny, K.N.; Gaalen, K.E.V.; Smith, C.M. Physiological acclimation of Pinus flexilis to drought stress on contrasting slope aspects in Waterton Lakes National Park, Alberta, Canada. Can. J. For. Res. 2009, 39, 629–641. [Google Scholar] [CrossRef]
- Pittman, S.D.; Turnblom, E.C. A study of self-thinning using coupled allometric equations: Implications for coastal Douglas-fir stand dynamics. Can. J. For. Res. 2003, 33, 1661–1669. [Google Scholar] [CrossRef]
- Hein, S.; Dhote, J.F. Effect of species composition, stand density and site index on the basal area increment of oak trees (Quercus sp.) in mixed stands with beech (Fagus sylvatica L.) in northern France. Ann. For. Sci. 2016, 63, 457–467. [Google Scholar] [CrossRef]

{kind=link}
| Variables | Mean | Minimum | Maximum | Standard Deviation | Q1 | Q3 |
|---|---|---|---|---|---|---|
| Age (year) | 26 | 7 | 44 | 9.2 | 18 | 34 |
| Dg (cm) | 16.7 | 4.2 | 28.0 | 7.4 | 8.9 | 23.2 |
| Hdom (m) | 20.7 | 6.5 | 32.5 | 4.5 | 20.2 | 22.2 |
| CW (m) | 1.7 | 1.1 | 2.3 | 0.3 | 1.6 | 1.9 |
| Density (trees·ha−1) | 927 | 283 | 1444 | 417 | 492 | 1392 |
| Site index | 18.6 | 12 | 24 | 3.0 | 16 | 22 |
| Components | Variables | Mean | Minimum | Maximum | Standard Deviation | Q1 | Q3 |
|---|---|---|---|---|---|---|---|
| Tree (n = 78) | DBH (cm) | 19.0 | 3.8 | 35.7 | 8.7 | 11.2 | 26.8 |
| HT (m) | 18.2 | 4.7 | 33.2 | 8.3 | 10.5 | 23.9 | |
| HD | 0.96 | 0.73 | 1.27 | 0.11 | 0.89 | 1.04 | |
| CR | 0.58 | 0.24 | 1.74 | 0.27 | 0.36 | 0.79 | |
| Largest branch (n = 1147) | BD (mm) | 17.41 | 1.35 | 73.74 | 12.34 | 7.14 | 25.01 |
| BL (cm) | 174.7 | 1.0 | 690.0 | 128.0 | 60.0 | 255.6 | |
| BC (cm) | 162.3 | 1.0 | 635.0 | 118.1 | 60.0 | 239.5 | |
| VA (°) | 58.4 | 10.0 | 101.0 | 13.2 | 50.0 | 68.0 |
| Model (4) | Estimate | SD | t Value | p Value | Model (6) | Estimate | SD | t Value | p Value |
|---|---|---|---|---|---|---|---|---|---|
| b1 | 0.3193 | 0.0316 | 10.10 | <0.001 | b1 | 0.3350 | 0.0330 | 10.15 | <0.001 |
| b2 | 0.5989 | 0.0330 | 18.14 | <0.001 | b2 | 0.5803 | 0.0332 | 17.49 | <0.001 |
| b3 | 0.6824 | 0.0260 | 26.28 | <0.001 | b30 | 0.4950 | 0.0686 | 7.21 | <0.001 |
| b4 | 0.1845 | 0.0541 | 3.41 | <0.001 | b31 | 0.0135 | 0.0044 | 3.07 | 0.002 |
| b5 | 0.3115 | 0.0152 | 20.54 | <0.001 | b4 | 0.2798 | 0.0629 | 4.45 | <0.001 |
| b5 | 0.3176 | 0.0155 | 20.54 | <0.001 | |||||
| R2adj | 0.67 | R2adj | 0.68 | ||||||
| MSE | 0.2704 | MSE | 0.2682 | ||||||
| AIC | 1762 | AIC | 1754 | ||||||
| BIC | 1792 | BIC | 1789 | ||||||
| MSER | - | MSER | 0.93% | ||||||
| DF | 1142 | DF | 1141 |
| Parameter | Estimate | SD | t Value | p Value |
|---|---|---|---|---|
| b1 | 0.3532 | 0.0415 | 8.50 | <0.001 |
| b2 | 0.5704 | 0.0406 | 14.06 | <0.001 |
| b30 | 0.5170 | 0.0922 | 5.61 | <0.001 |
| b31 | 0.0077 | 0.0059 | 1.30 | 0.1951 |
| b4 | 0.1652 | 0.0742 | 2.23 | 0.0261 |
| b5 | 0.3095 | 0.0147 | 20.99 | <0.001 |
| R2adj | 0.72 | |||
| MSE | 0.2324 | |||
| AIC | 1671 | |||
| BIC | 1712 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, H.; Chen, D.; Sun, X.; Zhang, S. Responses of the Crown Shape of Larix kaempferi Plantations to Site Index in Subtropical Areas of China. Forests 2023, 14, 2181. https://doi.org/10.3390/f14112181
Gao H, Chen D, Sun X, Zhang S. Responses of the Crown Shape of Larix kaempferi Plantations to Site Index in Subtropical Areas of China. Forests. 2023; 14(11):2181. https://doi.org/10.3390/f14112181
Chicago/Turabian StyleGao, Huilin, Dongsheng Chen, Xiaomei Sun, and Shougong Zhang. 2023. "Responses of the Crown Shape of Larix kaempferi Plantations to Site Index in Subtropical Areas of China" Forests 14, no. 11: 2181. https://doi.org/10.3390/f14112181