
Cloning, Bioinformatics Analysis and Physiological Function of the Pine Wood Nematode Bxadh2 Gene
Abstract
:1. Introduction
2. Materials and Methods
2.1. PWN Culture
2.2. Cloning of the PWN Bxadh2 Gene Coding Region
2.3. PWN Bxadh2 Gene Bioinformatics Analysis
2.4. PWN Bxadh2 Gene siRNA Synthesis
2.5. Functional Verification of the Bxadh2 Gene after PWN RNAi
2.5.1. Detection of Interference Efficiency
2.5.2. Determination of PWN Viability after RNA Interference
2.5.3. Determination of PWN Fecundity after RNA Interference
2.6. Data Statistics and Analysis
3. Results and Analysis
3.1. Cloning of the PWN Bxadh2 Gene
3.2. PWN Bxadh2 Gene Bioinformatics Analysis
3.2.1. Bxadh2 Physicochemical Properties
3.2.2. Bxadh2 Hydrophilic Analysis
3.2.3. Bxadh2 Transmembrane Domain Prediction
3.2.4. Bxadh2 Signal Peptide Prediction
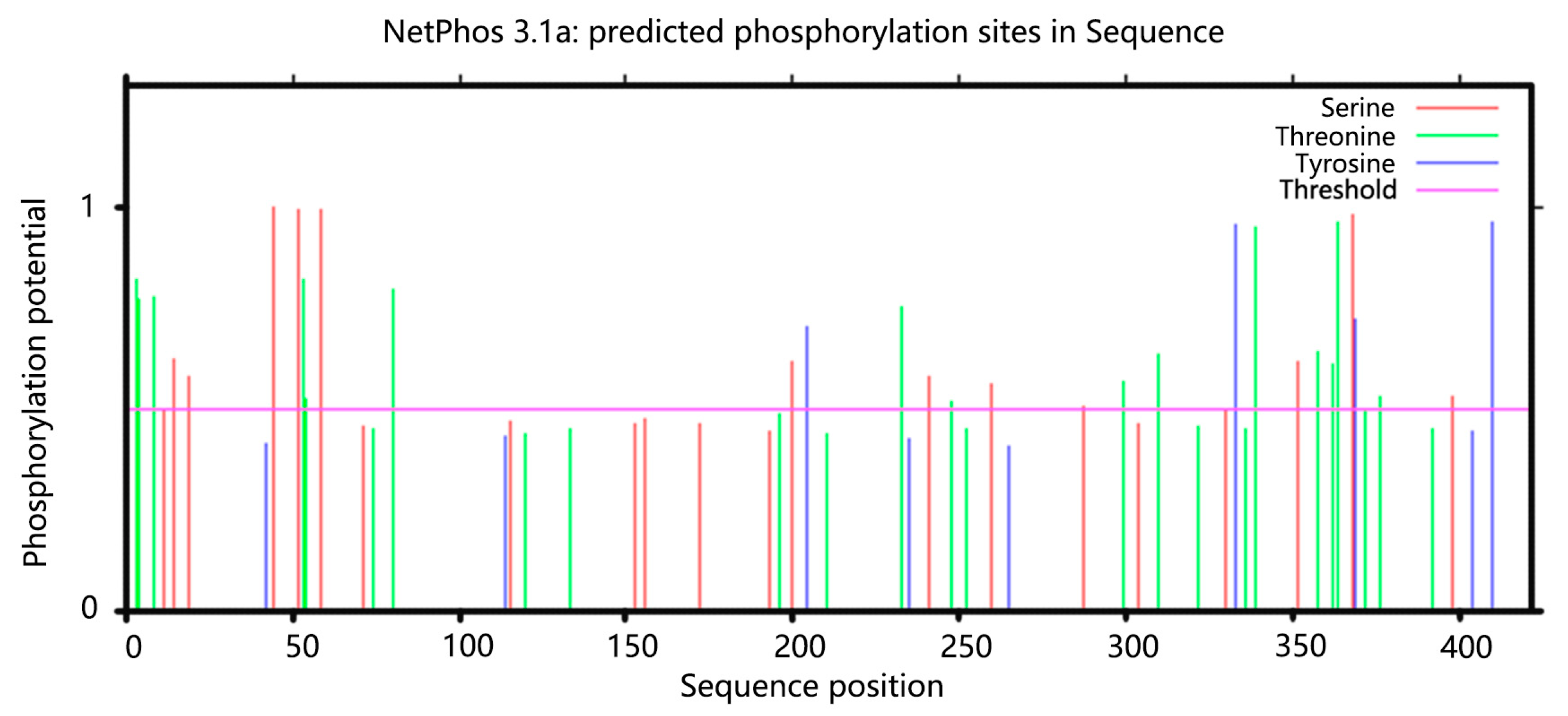
3.2.5. Bxadh2 Phosphorylation Site Prediction
3.2.6. Bxadh2 Secondary and Tertiary Structure Predictions
3.3. PWN Bxadh2 Gene Function Studies
3.3.1. PWN Bxadh2 Gene Interference Efficiency
3.3.2. Changes in PWN Activity after RNA Interference
3.3.3. PWN Fecundity Changes Following RNA Interference
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhu, L.H.; Ye, J.R.; Negi, S.; Xu, X.-L.; Wang, Z.-L.; Ji, J.-Y. Pathogenicity of Aseptic Bursaphelenchus xylophilus. PLoS ONE 2012, 7, e38095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, F.; Moreira, C.; Fonseca, L.; van Asch, B.; Mota, M.; Abrantes, I.; Amorim, A. New Insights into the Phylogeny and Worldwide Dispersion of Two Closely Related Nematode Species, Bursaphelenchus xylophilus and Bursaphelenchus mucronatus. PLoS ONE 2013, 8, e56288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallez, S.; Castagnone, C.; Espada, M.; Vieira, P.; Eisenback, J.D.; Mota, M.; Guillemaud, T.; Castagnone-Sereno, P. First Insights into the Genetic Diversity of the Pine Wood Nematode in Its Native Area Using New Polymorphic Microsatellite Loci. PLoS ONE 2013, 8, e59165. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi, T.; Cotton, J.A.; Dalzell, J.J.; Hasegawa, K.; Kanzaki, N.; McVeigh, P.; Takanashi, T.; Tsai, I.J.; Assefa, S.A.; Cock, P.J.A. Genomic insights into the origin of parasitism in the emerging plant pathogen Bursaphelenchus xylophilus. PLoS Pathog. 2011, 7, e1002219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosaka, H.; Aikawa, T.; Ogura, N.; Tabata, K.; Kiyohara, T. Pine wilt disease caused by the pine wood nematode: The induced resistance of pine trees by the avirulent isolates of nematode. Eur. J. Plant Pathol. 2001, 107, 667–675. [Google Scholar] [CrossRef]
- Kawai, M.; Shoda-Kagaya, E.; Maehara, T.; Zhou, Z.; Lian, C.; Iwata, R.; Yamane, A.; Hogetsu, T. Genetic structure of pine sawyer Monochamus alternatus (Coleoptera: Cerambycidae) populations in northeast Asia: Consequences of the spread of pine wilt disease. Environ. Entomol. 2006, 35, 569–579. [Google Scholar] [CrossRef]
- Jianren, Y. Analysis of the epidemic status, control techniques and countermeasures of pine wood nematode disease in China. For. Sci. 2019, 55, 1–10. [Google Scholar]
- Abelleira, A.; Picoaga, A.; Mansilla, J.P.; Aguin, O. Detection of Bursaphelenchus Xylophilus, Causal Agent of Pine Wilt Disease on Pinus pinaster in Northwestern Spain. Plant Dis. 2011, 95, 776–780. [Google Scholar] [CrossRef]
- Inacio, M.L.; Nobrega, F.; Vieira, P.; Bonifácio, L.; Naves, P.; Sousa, E.; Mota, M. First detection of Bursaphelenchus xylophilus associated with Pinus nigra in Portugal and in Europe. For. Pathol. 2015, 45, 235–238. [Google Scholar] [CrossRef]
- Zhao, L.L.; Wei, W.; Kang, L.; Sun, J.H. Chemotaxis of the pine wood nematode, Bursaphelenchus xylophilus, to volatiles associated with host pine, Pinus massoniana, and its vector Monochamus alternatus. J. Chem. Ecol. 2007, 33, 1207–1216. [Google Scholar] [CrossRef]
- Harmana, L.; Krusbergl, R.; Nicklew, R. Pine Wood Nematode, Bursaphelenchus xylophilus, Associated with Red Pine, Pinus resinosa, in Western Maryland. J. Nematol. 1986, 18, 575–580. [Google Scholar]
- Wang, M.; Wang, D.D.; Zhang, X.; Wang, X.; Liu, W.; Hou, X.; Huang, X.; Xie, B.; Cheng, X. Double-stranded RNA-mediated interference of dumpy genes in Bursaphelenchus xylophilus by feeding on filamentous fungal transformants. Int. J. Parasitol. 2016, 46, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.Y. The spread of pine wood nematode disease in China is serious. China For. Sci. Technol. 1992, 3, 6–7. [Google Scholar]
- Zhao, H.; Chen, C.; Liu, J.; Liu, P.; Liu, Q.; Jian, H. Aseptic Bursaphelenchus xylophilus does not reduce the mortality of young pine tree. For. Pathol. 2013, 43, 444–454. [Google Scholar] [CrossRef]
- Qiu, X.W.; Wu, X.Q.; Huang, L.; Tian, M.; Ye, J. Specifically Expressed Genes of the Nematode Bursaphelenchus xylophilus Involved with Early Interactions with Pine Trees. PLoS ONE 2013, 8, e78063. [Google Scholar] [CrossRef] [Green Version]
- Dang, Q.L.; Son, S.W.; Cheon, H.M.; Choi, G.J.; Choi, Y.H.; Jang, K.S.; Lim, C.H.; Kim, J.-C. Pyochelin isolated from Burkholderia arboris KRICT1 carried by pine wood nematodes exhibits phytotoxicity in pine callus. Nematology 2011, 13, 521–528. [Google Scholar] [CrossRef]
- Oku, H. Role of phytotoxins in pine wilt diseases. J. Nematol. 1988, 20, 245–251. [Google Scholar]
- Oku, H.S.T.; Kurozumi, S. Participation of toxin in wilting of Japanese pines caused by a nematode. Naturwissenschaften 1979, 66, 210. [Google Scholar] [CrossRef]
- Mamiya, Y. Inoculation of the first year pine (Pinus densiflora) seedlings with Bursaphelenchus lignicolus and the histopathology of diseased seedlings. J. Jpn. For. Soc. 1980, 62, 176–183. [Google Scholar]
- Wang, X. Study on the Pathogenesis of CYP450 Gene in Pine Wood Nematode; Chinese Academy of Forestry: Beijing, China, 2016; pp. 9–10. [Google Scholar]
- Schulerm, A.; Werck-Reichhart, D. Functional genomics of P450s. Annu. Rev. Plant Biol. 2003, 54, 629–667. [Google Scholar] [CrossRef]
- Arlt, V.M.; Stiborova, M.; Henderson, C.J.; Thiemann, M.; Frei, E.; Aimová, D.; Singh, R.; da Costa, G.G.; Schmitz, O.J.; Farmer, P.B.; et al. Metabolic activation of benzo a pyrene in vitro by hepatic cytochrome P450 contrasts with detoxification in vivo: Experiments with hepatic cytochrome P450 reductase null mice. Carcinogenesis 2008, 29, 656–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laing, R.; Bartley, D.J.; Morrison, A.A.; Rezansoff, A.; Martinelli, A.; Laing, S.T.; Gilleard, J.S. The cytochrome P450 family in the parasitic nematode Haemonchus contortus. Int. J. Parasitol. 2015, 45, 243–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.X.; Yan, Z.M.; Tan, J.J. Identification of the endophytic bacterium GD2 in Pinus sylvestris and its effect on the occurrence of pine nematode disease. Phytosanitary 2021, 5, 35. [Google Scholar] [CrossRef]
- Chen, Y.X.; Zhao, X.J.; Tan, J.J. Effects of the endophytic bacterium GD2 on the transcriptome of pine wood nematode (Pinus sylvestris) during host invasion. J. Huazhong Agric. Univ. 2021, 40, 9. [Google Scholar]
- Shen, D.P.; Zhang, X.Y.; Feng, M.T.; Jianren, Y.; Xiuwen, Q. Construction of cytochrome cyp-13A11 gene RNA interference vector and its function analysis in Pine Wood Nematode. J. Beijing For. Univ. 2021, 43, 96–102. [Google Scholar]
- Qiu, X.W.; Wu, X.Q.; Huang, L.; Ye, J.J. Cloning of Bxpel2 gene from pine wood nematode and construction of RNA interference vector. For. Res. 2016, 29, 283–288. [Google Scholar]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Xu, X.L.; Wu, X.Q.; Ye, J.R.; Huang, L. Molecular Characterization and Functional Analysis of Three Pathogenesis-Related Cytochrome P450 Genes from Bursaphelenchus xylophilus (Tylenchida: Aphelenchoidoidea). Int. J. Mol. Sci. 2015, 16, 5216–5234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, L.N.; Wu, X.Q.; Ye, J.R.; Xue, Q. Identification of Autophagy in the Pine Wood Nematode Bursaphelenchus xylophilus and the Molecular Characterization and Functional Analysis of Two Novel Autophagy-Related Genes, BxATG1 and BxATG8. Int. J. Mol. Sci. 2016, 17, 279. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.D.; Chen, G.H.; Pan, L.M. Analysis of pine nematode ncr gene function using RNAi. J. Plant Pathol. 2018, 48, 518–526. [Google Scholar]
- Wang, S.X. Captive Rearing and RNA Interference of Pine Nematodes; Zhejiang Forestry College: Hangzhou, China, 2007. [Google Scholar]
- Viglierchio, D.R.; Schmitt, R.V. On the methodology of nematode extraction from field samples: Baermann funnel modifications. J. Nematol. 1983, 15, 438–444. [Google Scholar] [PubMed]
- Mamiya, Y. Pathology of the Pine Wilt Disease Caused by Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 1983, 21, 201–220. [Google Scholar] [CrossRef]
- Futai, K. Pine Wood Nematode, Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 2013, 51, 61–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benenati, G.; Penkov, S.; Muller-Reichert, T.; Entchev, E.V.; Kurzchalia, T.V. Two cytochrome P450s in Caenorhabditis elegans are essential for the organization of eggshell, correct execution of meiosis and the polarization of embryo. Mech. Dev. 2009, 126, 382–393. [Google Scholar] [CrossRef]
- Chapple, C. Molecular-genetic Analysis of Plant Cytochrome P450-Dependent Monooxtgenases. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 311–343. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.T.; Shen, D.P.; Ye, J.R.; Chen, F.; Zhang, X.; Qiu, X. RNA Interference and Function of cyp-33D3 Gene in Pine wood Nematode. Res. For. Sci. 2020, 33, 32–38. [Google Scholar]
- Wang, D.D. Cloning and Functional Study of Dormance-Related Genes daf-3, daf-9, daf-12 and daf-16 in Pine Wood Nematodes; Jilin University: Changchun, China, 2012. [Google Scholar]
- Xu, C.J.; Li, C.Y.T.; Kong, A.N.T. Induction of phase I, II and III drug metabolism/transport by xenobiotics. Arch. Pharm. Res. 2005, 28, 249–268. [Google Scholar] [CrossRef]
- Yan, X.; Cheng, X.Y.; Wang, Y.S.; Luo, J.; Mao, Z.-C.; Ferris, V.R.; Xie, B.-Y. Comparative transcriptomics of two pathogenic pine wood nematodes yields insights into parasitic adaptation to life on pine hosts. Gene 2012, 505, 81–90. [Google Scholar] [CrossRef]
- Guengerichf, P. Cytochrome P450 and chemical toxicology. Chem. Res. Toxicol. 2008, 21, 70–83. [Google Scholar] [CrossRef]
- Niu, J.H.; Jian, H.; Xu, J.M.; Guo, Y.D.; Liu, Q. RNAi technology extends its reach: Engineering plant resistance against harmful eukaryotes. Afr. J. Biotechnol. 2010, 9, 7573–7582. [Google Scholar]
- Wang, X.R.; Cheng, X.; Li, Y.D.; Zhang, J.-A.; Zhang, Z.-F.; Wu, H.-R. Cloning arginine kinase gene and its RNAi in Bursaphelenchus xylophilus causing pine wilt disease. Eur. J. Plant Pathol. 2012, 134, 521–532. [Google Scholar] [CrossRef]
- Kang, J.S.; Koh, Y.H.; Moon, Y.S.; Lee, S.H. Molecular properties of a venom allergen-like protein suggest a parasitic function in the pine wood nematode Bursaphelenchus xylophilus. Int. J. Parasitol. 2012, 42, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.W.; Yang, L.L.; Ye, J.R.; Wang, W.; Zhao, T.; Hu, H.; Zhou, G. Silencing of cyp-33C9 Gene Affects the Reproduction and Pathogenicity of the Pine Wood Nematode, Bursaphelenchus xylophilus. Int. J. Mol. Sci. 2019, 20, 4520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawazu, K.; Zhang, H.; Kanzaki, H. Accumulation of benzoic acid in suspension cultured cells of Pinus thunbergii Parl. in response to phenylacetic acid administration. Biosci. Biotechnol. Biochem. 1996, 60, 1410–1412. [Google Scholar] [CrossRef]
- Ma, H.B. Cloning and RNA Interference of the Cellulose Lineage and Its cDNA Gene of the Pine Wood Nematode; China Academy of Forestry Science: Beijing, China, 2008. [Google Scholar]
- Wang, L.S. Physiological Function of Alcohol Dehydrogenase Gene in Pinus sylvestris; Qingdao University: Qingdao, China, 2019. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward Primers | Reverse Primer |
|---|---|
| AAGGCCATGGCTGATATCGAAGAGGACCAAGATACGGA | GAATTCGGATCCGATATCTCACTTCCACAAG-TCAAGGAC |
| Bxadh2 (5′-3′) | Negative Control (5′-3′) | |
|---|---|---|
| Oligo—1 | GATCACTAATACGACTCACATAGGGGCCCAACTGTGACAACGTTTT | GATCACTAATACGACTCAC-TATAGGGGGGATGTCTCACATCTTGTTT |
| Oligo—2 | CTAGTGATTATGCTGAGTGATATCCCCGGGTTGACACTGTTGCAAAA | CTAGTGATTATGCTGAGTGATATCCCCCCTACAGAGTGTAGAACAAA |
| Oligo—3 | AAGCCCAACTGTGACAACGTTCCCTATAGTGAGTCGTATTAGTGATC | AAGGGATGTCTCACATCTTGTCCCTATAGTGAGTCGTATTAGTGATC |
| Oligo—4 | TTCGGGTTGACACTGTTGC-AAGGGATATCACTCAGCATAATCACTAG | TTCCCTACAGAGTGTAGAACAGGGATATCACTCAGCATAATCACTAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, L.; Hu, J.; Chen, Y.; Tan, J.; Ye, J. Cloning, Bioinformatics Analysis and Physiological Function of the Pine Wood Nematode Bxadh2 Gene. Forests 2023, 14, 1283. https://doi.org/10.3390/f14071283
Shen L, Hu J, Chen Y, Tan J, Ye J. Cloning, Bioinformatics Analysis and Physiological Function of the Pine Wood Nematode Bxadh2 Gene. Forests. 2023; 14(7):1283. https://doi.org/10.3390/f14071283
Chicago/Turabian StyleShen, Luyang, Jiafeng Hu, Yangxue Chen, Jiajin Tan, and Jianren Ye. 2023. "Cloning, Bioinformatics Analysis and Physiological Function of the Pine Wood Nematode Bxadh2 Gene" Forests 14, no. 7: 1283. https://doi.org/10.3390/f14071283
APA StyleShen, L., Hu, J., Chen, Y., Tan, J., & Ye, J. (2023). Cloning, Bioinformatics Analysis and Physiological Function of the Pine Wood Nematode Bxadh2 Gene. Forests, 14(7), 1283. https://doi.org/10.3390/f14071283