
Functional Validation and Promoter DNA Methylation Analysis of the OfPAO Gene of Osmanthus fragrans ‘Yinbi Shuanghui’
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DNA and RNA Extraction, cDNA Preparation, and qRT-PCR Analysis
2.3. Transient Overexpression of OfPAO in Tobacco
2.4. Transformation of OfPAO in Tobacco
2.5. qRT-PCR Analysis of Transgenic Tobacco
2.6. Promoter Cloning Ligation Transformation Sequencing
2.7. Promoter Bisulfite Sequencing
2.8. Bioinformatic Analysis of Gene Sequences
3. Results
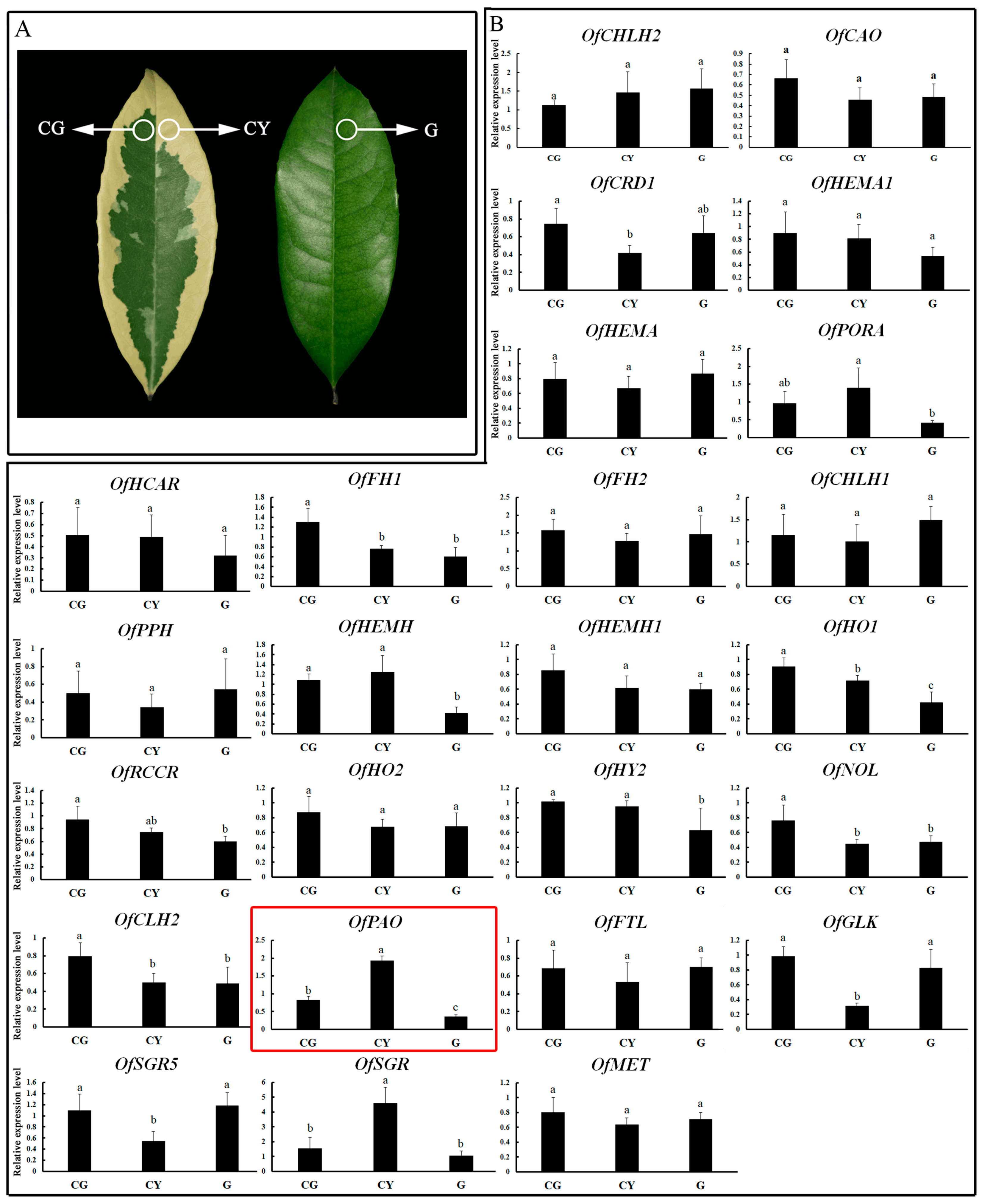
3.1. Expression Patterns of Leaf Color-Related Genes
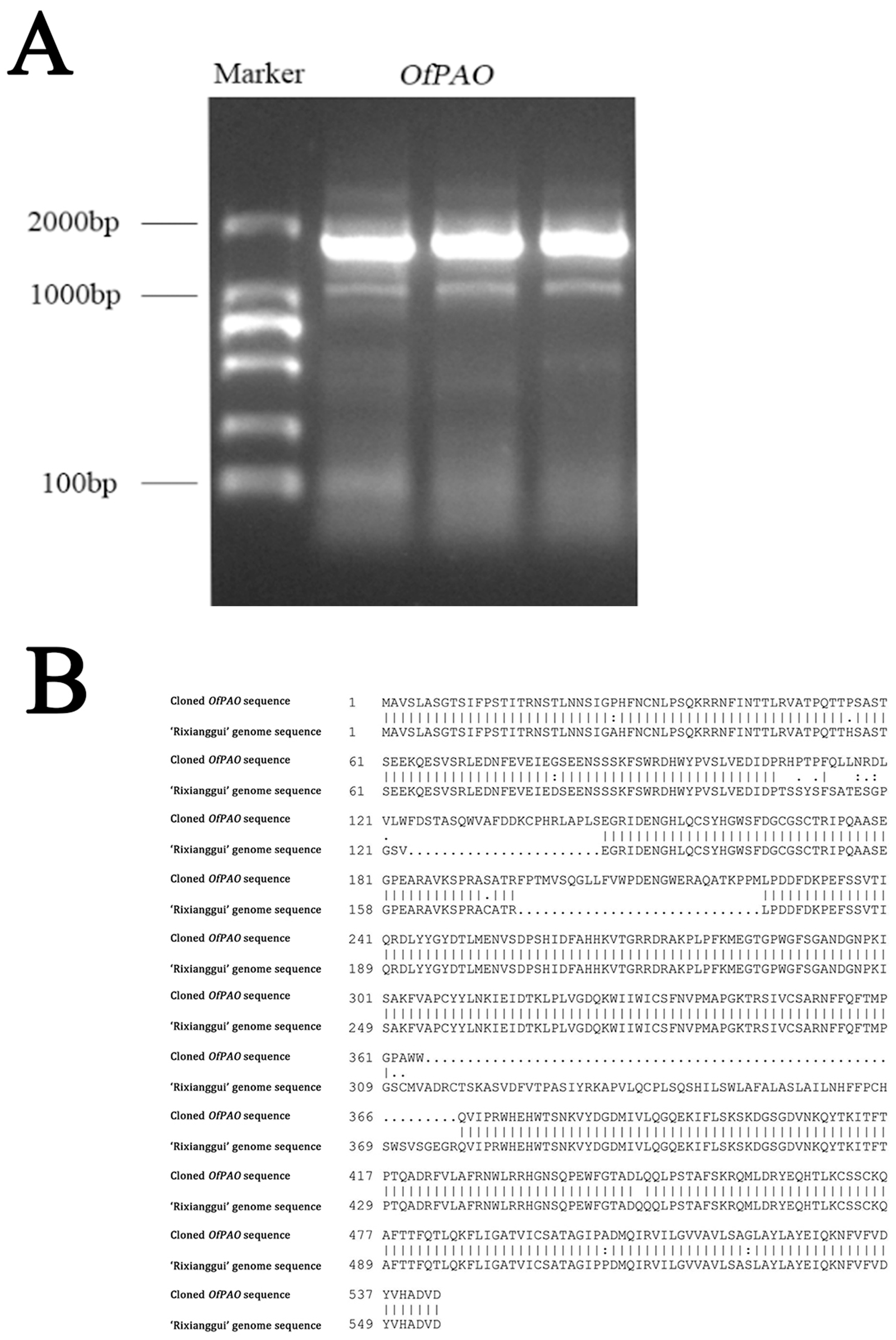
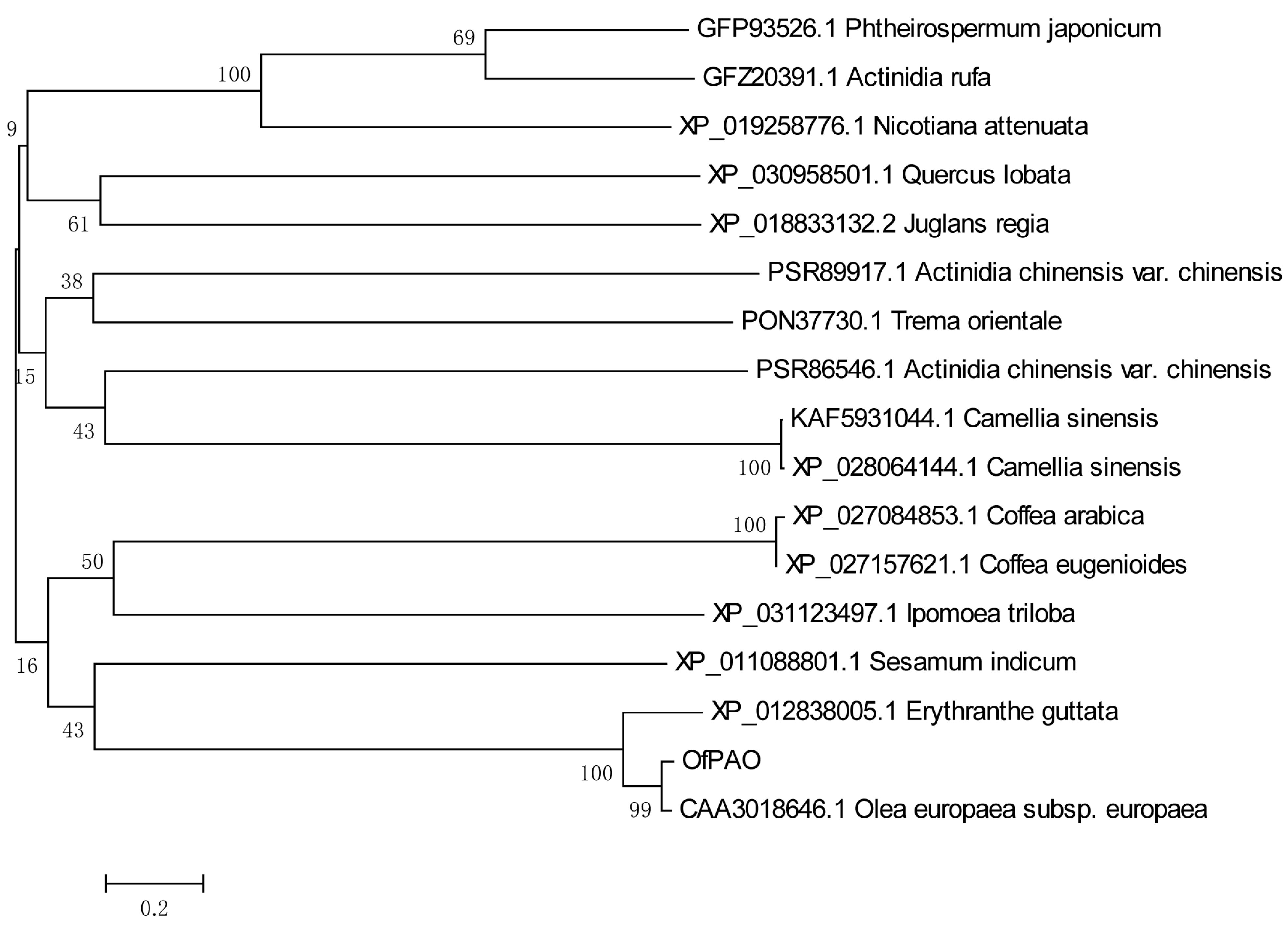
3.2. OfPAO Cloning and Sequence Analysis
3.3. Potential Regulatory Roles of OfPAO in Tobacco with Transient Expression
3.4. The Overexpression of OfPAO Promotes the Degradation of Chlorophyll in Tobacco
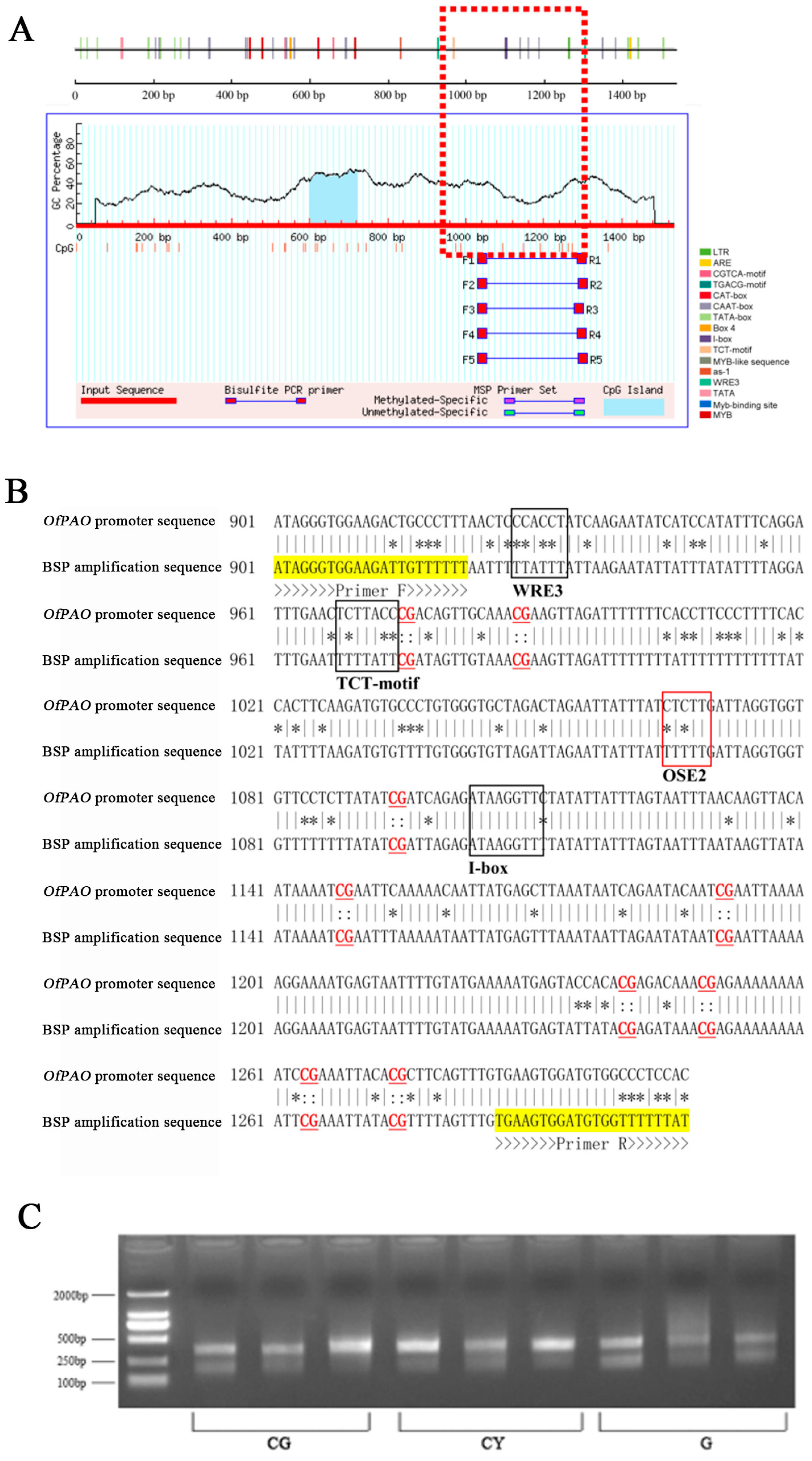
3.5. OfPAO Promoter Sequence Cis-Acting Element Prediction
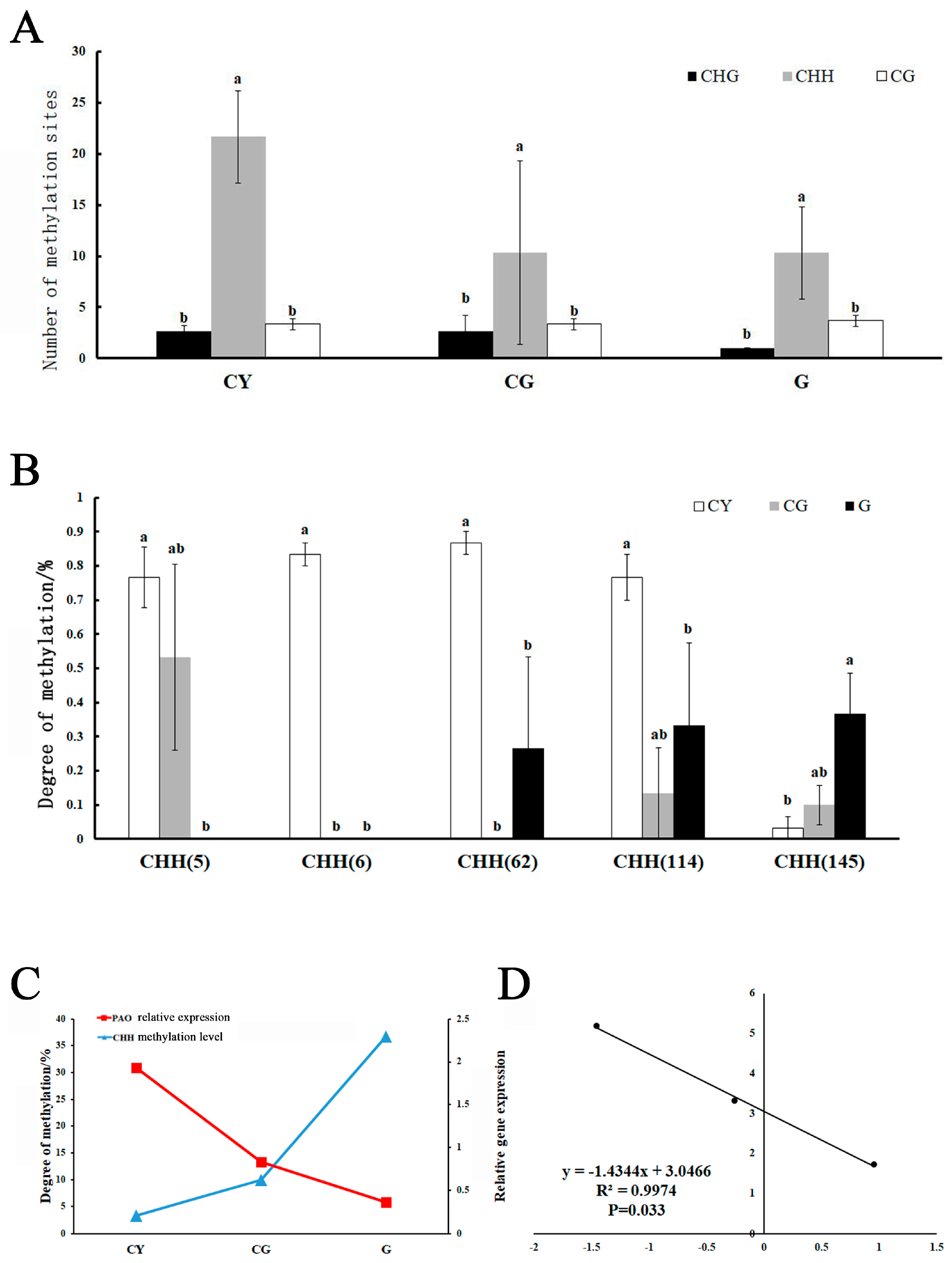
3.6. DNA Methylation CpG Island Prediction of the OfPAO Promoter
3.7. Relationship between the Methylation Degree and the Gene Expression Level of the OfPAO Promoter
4. Discussion
4.1. OfPAO Is a Key Gene for Leaf Color Formation
4.2. The Methylation of the OfPAO Promoter Is Involved in Regulating Its Expression
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fambrini, M. Characterization of a pigment-deficient mutant of sunflower (Helianthus annuus L.) with abnormal chloroplast biogenesis, reduced PS II activity and low endogenous level of abscisic acid. Plant Sci. 2004, 167, 79–89. [Google Scholar] [CrossRef]
- Parks, B.M.; Quail, P.H. Phytochrome-Deficient hy1 and hy2 Long Hypocotyl Mutants of Arabidopsis Are Defective in Phytochrome Chromophore Biosynthesis. Plant Cell 1991, 3, 1177–1186. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.-H.; Li, X.; Zhang, X.-X.; Zhang, H.; Zhao, X.-Y. Mutation Mechanism of Leaf Color in Plants: A Review. Forests 2020, 11, 851. [Google Scholar] [CrossRef]
- Chen, X.; Yang, X.; Xie, J.; Ding, W.; Li, Y.; Yue, Y.; Wang, L. Biochemical and Comparative Transcriptome Analyses Reveal Key Genes Involved in Major Metabolic Regulation Related to Colored Leaf Formation in Osmanthus fragrans ‘Yinbi Shuanghui’ during Development. Biomolecules 2020, 10, 549. [Google Scholar] [CrossRef] [PubMed]
- Singh, U.P.; Prithiviraj, B.; Sarma, B.K. Development of Erysiphe pisi (Powdery Mildew) on Normal and Albino Mutants of Pea (Pisum sativum L.). J. Phytopathol. 2000, 148, 591–595. [Google Scholar] [CrossRef]
- Pruzinská, A.; Tanner, G.; Aubry, S.; Anders, I.; Moser, S.; Müller, T.; Ongania, K.H.; Kräutler, B.; Youn, J.Y.; Liljegren, S.J.; et al. Chlorophyll breakdown in senescent Arabidopsis leaves. Characterization of chlorophyll catabolites and of chlorophyll catabolic enzymes involved in the degreening reaction. Plant Physiol. 2005, 139, 52–63. [Google Scholar] [CrossRef]
- Agrawal, G.K.; Yamazaki, M.; Kobayashi, M.; Hirochika, R.; Miyao, A.; Hirochika, H. Screening of the rice viviparous mutants generated by endogenous retrotransposon tos17 insertion. tagging of a zeaxanthin epoxidase gene and a novel ostatc gene. Plant Physiol. 2001, 125, 1248–1257. [Google Scholar] [CrossRef] [PubMed]
- Pruzinska, A.; Tanner, G.; Anders, I.; Roca, M.; Hoertensteiner, S. Chlorophyll breakdown: Pheophorbide a oxygenase is a Rieske-type iron–sulfur protein, encoded by the accelerated cell death 1 gene. Proc. Natl. Acad. Sci. USA 2003, 100, 15259–15264. [Google Scholar] [CrossRef]
- Xie, Q.; Liang, Y.; Zhang, J.; Zheng, H.; Dong, G.; Qian, Q.; Zuo, J. Involvement of a Putative Bipartite Transit Peptide in Targeting Rice Pheophorbide a Oxygenase into Chloroplasts for Chlorophyll Degradation during Leaf Senescence. J. Genet. Genom. 2016, 43, 145–154. [Google Scholar] [CrossRef]
- Becker, C.; Hagmann, J.; Müller, J.; Koenig, D.; Stegle, O.; Borgwardt, K.; Weigel, D. Spontaneous epigenetic variation in the Arabidopsis thaliana methylome. Nature 2011, 480, 245–249. [Google Scholar] [CrossRef]
- Finnegan, E.J.; Peacock, W.J.; Dennis, E.S. DNA methylation, a key regulator of plant development and other processes. Curr. Opin. Genet. Dev. 2000, 10, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.; Dubin, M.; Zhang, P.; Osborne, E.J.; Stegle, O.; Clark, R.M.; Nordborg, M. Limited Contribution of DNA Methylation Variation to Expression Regulation in Arabidopsis thaliana. PLoS Genet. 2016, 12, e1006141. [Google Scholar] [CrossRef] [PubMed]
- Grativol, C.; Hemerly, A.S.; Ferreira, P.C. Genetic and epigenetic regulation of stress responses in natural plant populations. Biochim. Biophys. Acta 2012, 1819, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Ji, W.; Yang, J.; Yang, L.; Chen, W.; Zhuang, Z. Comparison of global DNA methylation profiles in replicative versus premature senescence. Life Sci. 2008, 83, 475–480. [Google Scholar] [CrossRef]
- Wu, S.C.; Zhang, Y. Active DNA demethylation: Many roads lead to Rome. Nat. Rev. Mol. Cell Biol. 2010, 11, 607–620. [Google Scholar] [CrossRef]
- Zou, J.-J.; Cai, X.; Yang, J.; Zeng, X.; Liu, D.-X.; Huang, S.; Chen, X.; Yang, Q.-Y.; Wang, C.; Chen, H. DNA hypomethylation mediates flower opening and senescence in sweet osmanthus through auxin and ethylene responsive pathways. Postharvest Biol. Technol. 2023, 198, 112250. [Google Scholar] [CrossRef]
- Kuhlmann, M.; Jiang, H.; Catoni, M.; Johannes, F. Editorial: DNA Methylation in Plants Associated With Abiotic Stress. Front. Plant Sci. 2021, 12, 778004. [Google Scholar] [CrossRef]
- Huang, Q.; Hua, X.; Zhang, Q.; Pan, W.; Wang, Y.; Liu, G.; Wei, H.; Chen, Y.; Zhang, J. Identification and functional verification of salt tolerance hub genes in Salix matsudana based on QTL and transcriptome analysis. Environ. Exp. Bot. 2023, 215, 105470. [Google Scholar] [CrossRef]
- Jin, P.; Chao, K.; Li, J.; Wang, Z.; Cheng, P.; Li, Q.; Wang, B. Functional Verification of Two Genes Related to Stripe Rust Resistance in the Wheat-Leymus mollis Introgression Line M8664-3. Front. Plant Sci. 2021, 12, 754823. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, P.; Hu, G.; Xiao, J.; Zhang, A.; Luo, X.; Wu, J. Overexpressing rice lesion simulating disease 1-like gene (OsLOL1) in Gossypium hirsutum promotes somatic embryogenesis and plant regeneration. J. Cotton Res. 2020, 3, 19. [Google Scholar] [CrossRef]
- Zhang, T.; Li, Z.; Song, X.; Han, L.; Wang, L.; Zhang, J.; Long, Y.; Pei, X. Identification and Characterization of microRNAs in the Developing Seed of Linseed Flax (Linum usitatissimum L.). Int. J. Mol. Sci. 2020, 21, 2708. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.X.; Qi, L.W. Over-Expression of LaMIR166a Promotes Organs Development in Nicotiana benthamiana. Russ. J. Plant Physiol. 2019, 66, 718–724. [Google Scholar] [CrossRef]
- Zheng, B.; Li, H.; Zhang, J.; Cui, C.; Jiang, L. Construction of Overexpression Vector of CONSTANS Gene Plant in Brassica napus and Production of Transgenic Plants. Mol. Plant Breed. 2018, 9, 73–79. [Google Scholar] [CrossRef]
- Renckens, S.; De Greve, H.; Van Montagu, M.; Hernalsteens, J.P. Petunia plants escape from negative selection against a transgene by silencing the foreign DNA via methylation. Mol. Gen. Genet. 1992, 233, 53–64. [Google Scholar] [CrossRef]
- Sri, T.; Gupta, B.; Tyagi, S.; Singh, A. Homeologs of Brassica SOC1, a central regulator of flowering time, are differentially regulated due to partitioning of evolutionarily conserved transcription factor binding sites in promoters. Mol. Phylogenet. Evol. 2020, 147, 106777. [Google Scholar] [CrossRef]
- Rahman, M.A.; Ullah, H. Receptor for Activated C Kinase1B (RACK1B) Delays Salinity-Induced Senescence in Rice Leaves by Regulating Chlorophyll Degradation. Plants 2023, 12, 2385. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.Y.; Wang, L.L.; Lin, Y.L.; Li, X.; Liu, X.F.; Xu, Z.H.; Fu, B.L.; Wang, W.Q.; Allan, A.C.; Tu, M.Y.; et al. AcHZP45 is a repressor of chlorophyll biosynthesis and activator of chlorophyll degradation in kiwifruit. J. Exp. Bot. 2023, erad361. [Google Scholar] [CrossRef]
- Rong, H.; Tang, Y.; Zhang, H.; Wu, P.; Chen, Y.; Li, M.; Wu, G.; Jiang, H. The Stay-Green Rice like (SGRL) gene regulates chlorophyll degradation in rice. J. Plant Physiol. 2013, 170, 1367–1373. [Google Scholar] [CrossRef]
- Li, J.; Wu, K.; Li, L.; Ma, G.; Fang, L.; Zeng, S. Transcriptomic analysis reveals biosynthesis genes and transcription factors related to leaf anthocyanin biosynthesis in Aglaonema commutatum. BMC Genom. 2023, 24, 28. [Google Scholar] [CrossRef]
- Eckhardt, U.; Grimm, B.; Hörtensteiner, S. Recent advances in chlorophyll biosynthesis and breakdown in higher plants. Plant Mol. Biol. 2004, 56, 1–14. [Google Scholar] [CrossRef]
- Hu, X.; Jia, T.; Hörtensteiner, S.; Tanaka, A.; Tanaka, R. Subcellular localization of chlorophyllase2 reveals it is not involved in chlorophyll degradation during senescence in Arabidopsis thaliana. Plant Sci. 2020, 290, 110314. [Google Scholar] [CrossRef]
- Xue, Y.; Li, X.; Mao, M.; He, Y.; Owusu Adjei, M.; Zhou, X.; Hu, H.; Liu, J.; Li, X.; Ma, J. AbhemC encoding porphobilinogen deaminase plays an important role in chlorophyll biosynthesis and function in albino Ananas comosus var. bracteatus leaves. PeerJ 2021, 9, e11118. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Z.; Wang, P.; Wang, S.; Ma, L.; Li, L.; Yang, R.; Ma, Y.; Wang, Q. Comprehensive transcriptome analysis discovers novel candidate genes related to leaf color in a Lagerstroemia indica yellow leaf mutant. Genes Genom. 2015, 37, 851–863. [Google Scholar] [CrossRef]
- Hörtensteiner, S. Update on the biochemistry of chlorophyll breakdown. Plant Mol. Biol. 2013, 82, 505–517. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Q.; Fu, N.; Song, M.; Han, X.; Yang, Q.; Zhang, Y.; Tong, Z.; Zhang, J. Cyanidin-3-O-glucoside Contributes to Leaf Color Change by Regulating Two bHLH Transcription Factors in Phoebe bournei. Int. J. Mol. Sci. 2023, 24, 3829. [Google Scholar] [CrossRef]
- Mingyue, F.U.; Cheng, S.; Feng, X.U.; Chen, Z.; Wang, L. Advance in mechanism of plant leaf colour mutation. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12071. [Google Scholar]
- Amir-Shapira, D.; Goldschmidt, E.E.; Altman, A. Chlorophyll catabolism in senescing plant tissues: In vivo breakdown intermediates suggest different degradative pathways for Citrus fruit and parsley leaves. Proc. Natl. Acad. Sci. USA 1987, 84, 1901–1905. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Han, J.; Zhu, B.; Jia, H.; Yang, T.; Wang, R.; Deng, W.W.; Zhang, Z.Z. Significantly increased amino acid accumulation in a novel albino branch of the tea plant (Camellia sinensis). Planta 2019, 249, 363–376. [Google Scholar] [CrossRef] [PubMed]
- Li, C.F.; Xu, Y.X.; Ma, J.Q.; Jin, J.Q.; Huang, D.J.; Yao, M.Z.; Ma, C.L.; Chen, L. Biochemical and transcriptomic analyses reveal different metabolite biosynthesis profiles among three color and developmental stages in ‘Anji Baicha’ (Camellia sinensis). BMC Plant Biol. 2016, 16, 195. [Google Scholar] [CrossRef] [PubMed]
- Rosianskey, Y.; Dahan, Y.; Yadav, S.; Freiman, Z.E.; Milo-Cochavi, S.; Kerem, Z.; Eyal, Y.; Flaishman, M.A. Chlorophyll metabolism in pollinated vs. parthenocarpic fig fruits throughout development and ripening. Planta 2016, 244, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Chen, Z.; Sun, W.; Deng, T.; Chen, M. De novo Sequencing of the Leaf Transcriptome Reveals Complex Light-Responsive Regulatory Networks in Camellia sinensis cv. Baijiguan. Front. Plant Sci. 2016, 7, 332. [Google Scholar] [CrossRef]
- Gao, Y.; Hao, J.-L.; Wang, Z.; Song, K.-J.; Ye, J.-H.; Zheng, X.-Q.; Liang, Y.-R.; Lu, J.-L. DNA methylation levels in different tissues in tea plant via an optimized HPLC method. Hortic. Environ. Biotechnol. 2019, 60, 967–974. [Google Scholar] [CrossRef]
- Vilperte, V.; Agapito-Tenfen, S.Z.; Wikmark, O.G.; Nodari, R.O. Levels of DNA methylation and transcript accumulation in leaves of transgenic maize varieties. Environ. Sci. Eur. 2016, 28, 29. [Google Scholar] [CrossRef]
- Akimoto, K.; Katakami, H.; Kim, H.J.; Ogawa, E.; Sano, C.M.; Wada, Y.; Sano, H. Epigenetic inheritance in rice plants. Ann. Bot. 2007, 100, 205–217. [Google Scholar] [CrossRef]
- Xiao, K.; Chen, J.; He, Q.; Wang, Y.; Shen, H.; Sun, L. DNA methylation is involved in the regulation of pepper fruit ripening and interacts with phytohormones. J. Exp. Bot. 2020, 71, 1928–1942. [Google Scholar] [CrossRef]
- Telias, A.; Lin-Wang, K.; Stevenson, D.E.; Cooney, J.M.; Hellens, R.P.; Allan, A.C.; Hoover, E.E.; Bradeen, J.M. Apple skin patterning is associated with differential expression of MYB10. BMC Plant Biol. 2011, 11, 93. [Google Scholar] [CrossRef]
- Wang, Z.; Meng, D.; Wang, A.; Li, T.; Jiang, S.; Cong, P.; Li, T. The methylation of the PcMYB10 promoter is associated with green-skinned sport in Max Red Bartlett pear. Plant Physiol. 2013, 162, 885–896. [Google Scholar] [CrossRef]
- Liu, G.; Li, C.; Yu, H.; Tao, P.; Yuan, L.; Ye, J.; Chen, W.; Wang, Y.; Ge, P.; Zhang, J.; et al. GREEN STRIPE, encoding methylated TOMATO AGAMOUS-LIKE 1, regulates chloroplast development and Chl synthesis in fruit. New Phytol. 2020, 228, 302–317. [Google Scholar] [CrossRef]
- Cocciolone, S.M.; Cone, K.C. Pl-Bh, an anthocyanin regulatory gene of maize that leads to variegated pigmentation. Genetics 1993, 135, 575–588. [Google Scholar] [CrossRef]
- Wang, Q.M.; Wang, L.; Zhou, Y.; Cui, J.; Wang, Y.; Zhao, C. Leaf patterning of Clivia miniata var. variegata is associated with differential DNA methylation. Plant Cell Rep. 2016, 35, 167–184. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function | Number | Element Name | Sequence |
|---|---|---|---|
| Cis-acting elements involved in the cryogenic response | 1 | LTR | CCGAAA |
| Cis-acting regulatory elements essential for anaerobic induction | 1 | ARE | AAACCA |
| Cis-acting regulatory elements involved in MeJA reactivity | 3 | CGTCA-motif | CGTCA |
| TGACG-motif | TGACG | ||
| Cis-regulatory elements associated with meristematic tissue expression | 1 | CAT-box | GCCACT |
| Common cis-acting elements in promoter and enhancer regions | 16 | CAAT-box | CAAAT |
| CAAT-box | TGCCAAC | ||
| Core promoter elements around the transcription promoter (i.e., about −30) | 14 | TATA-box | TATA |
| TATA-box | ATATAT | ||
| TATA-box | ATATAA | ||
| TATA-box | ATTATA | ||
| Part of a conserved DNA module involved in light responsiveness | 1 | Box 4 | ATTAAT |
| Part of a light-responsive element | 2 | I-box | AGATAAGG |
| TCT-motif | TCTTAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; Zhou, Y.; Chen, X.; Wei, H.; Zheng, D.; Zhu, W.; Wang, L.; Yang, X. Functional Validation and Promoter DNA Methylation Analysis of the OfPAO Gene of Osmanthus fragrans ‘Yinbi Shuanghui’. Forests 2024, 15, 11. https://doi.org/10.3390/f15010011
Wang R, Zhou Y, Chen X, Wei H, Zheng D, Zhu W, Wang L, Yang X. Functional Validation and Promoter DNA Methylation Analysis of the OfPAO Gene of Osmanthus fragrans ‘Yinbi Shuanghui’. Forests. 2024; 15(1):11. https://doi.org/10.3390/f15010011
Chicago/Turabian StyleWang, Rui, Yixiao Zhou, Xuan Chen, Hao Wei, Dong Zheng, Wuwei Zhu, Lianggui Wang, and Xiulian Yang. 2024. "Functional Validation and Promoter DNA Methylation Analysis of the OfPAO Gene of Osmanthus fragrans ‘Yinbi Shuanghui’" Forests 15, no. 1: 11. https://doi.org/10.3390/f15010011