
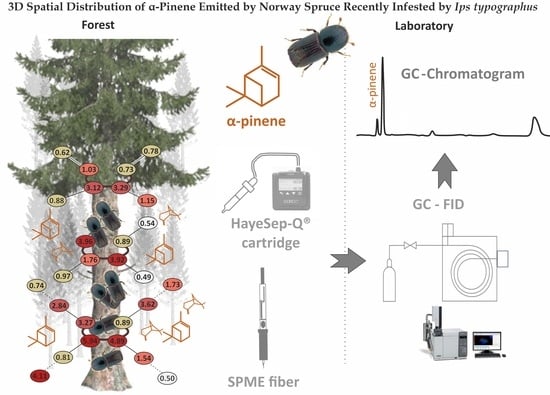
Pilot Study of 3D Spatial Distribution of α-Pinene Emitted by Norway Spruce (L.) Karst Recently Infested by Ips typographus (L. 1758) (Coleoptera: Scolytinae)
 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Optimization of VOC Collection from Simulatively Infested Spruce Logs in a Laboratory
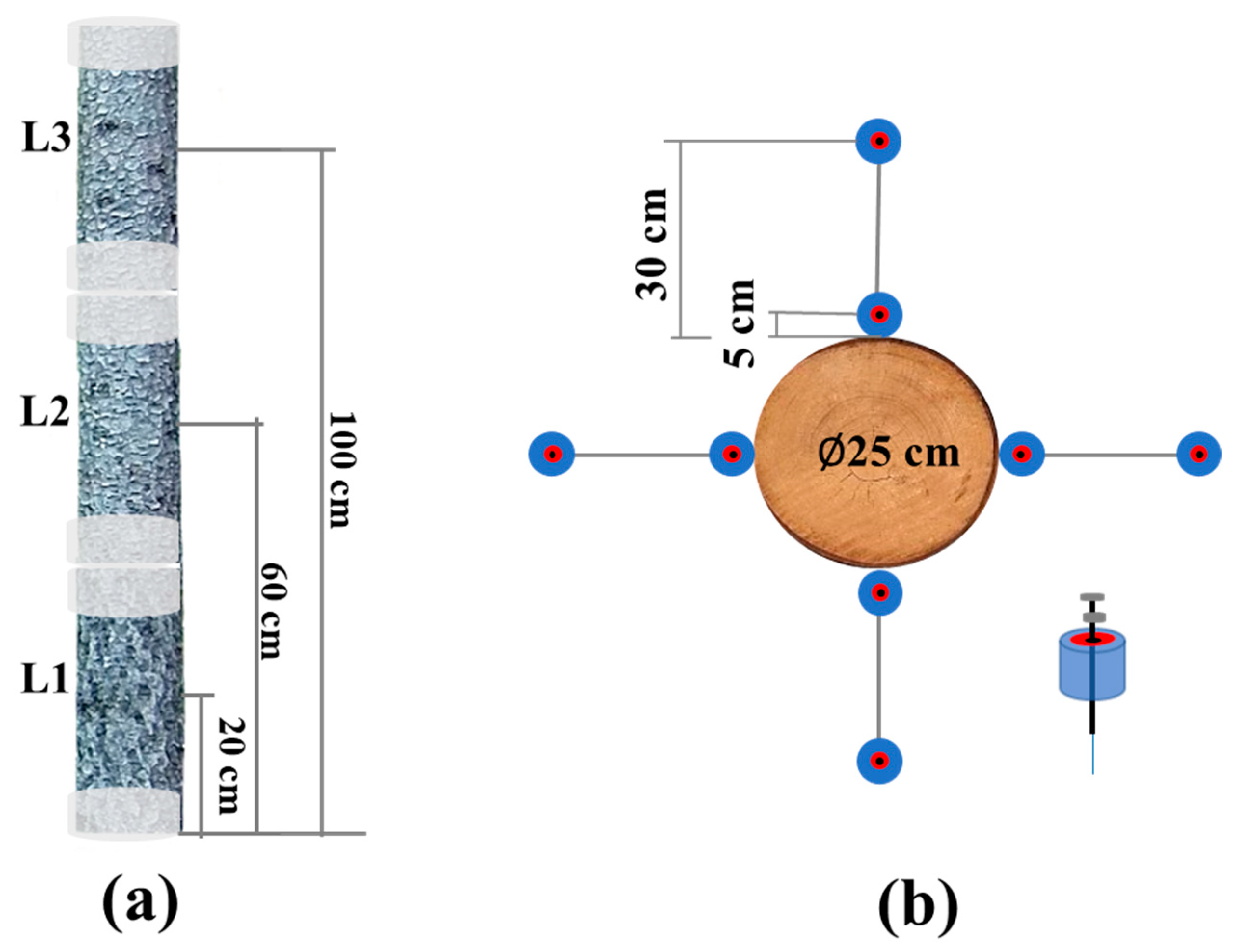
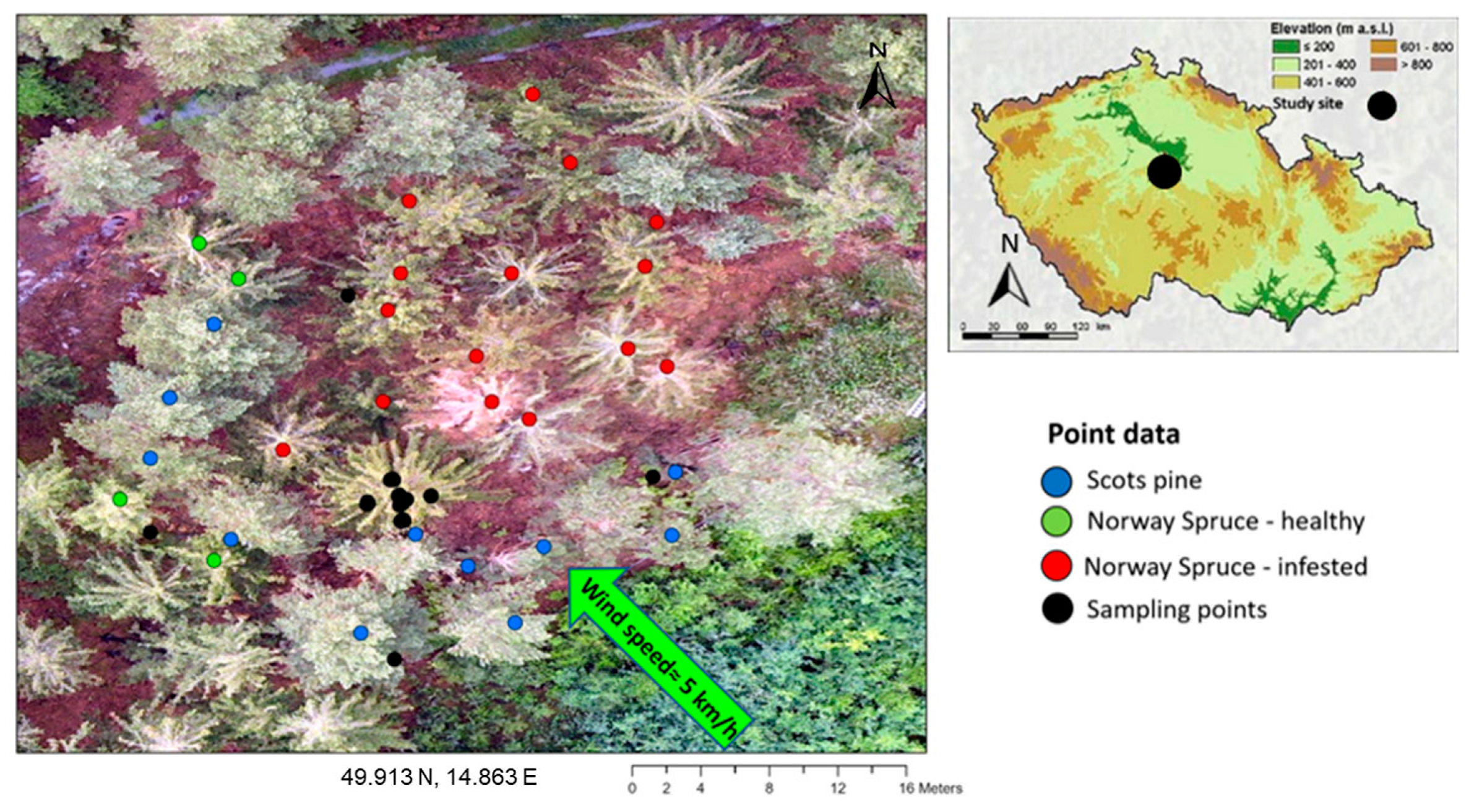
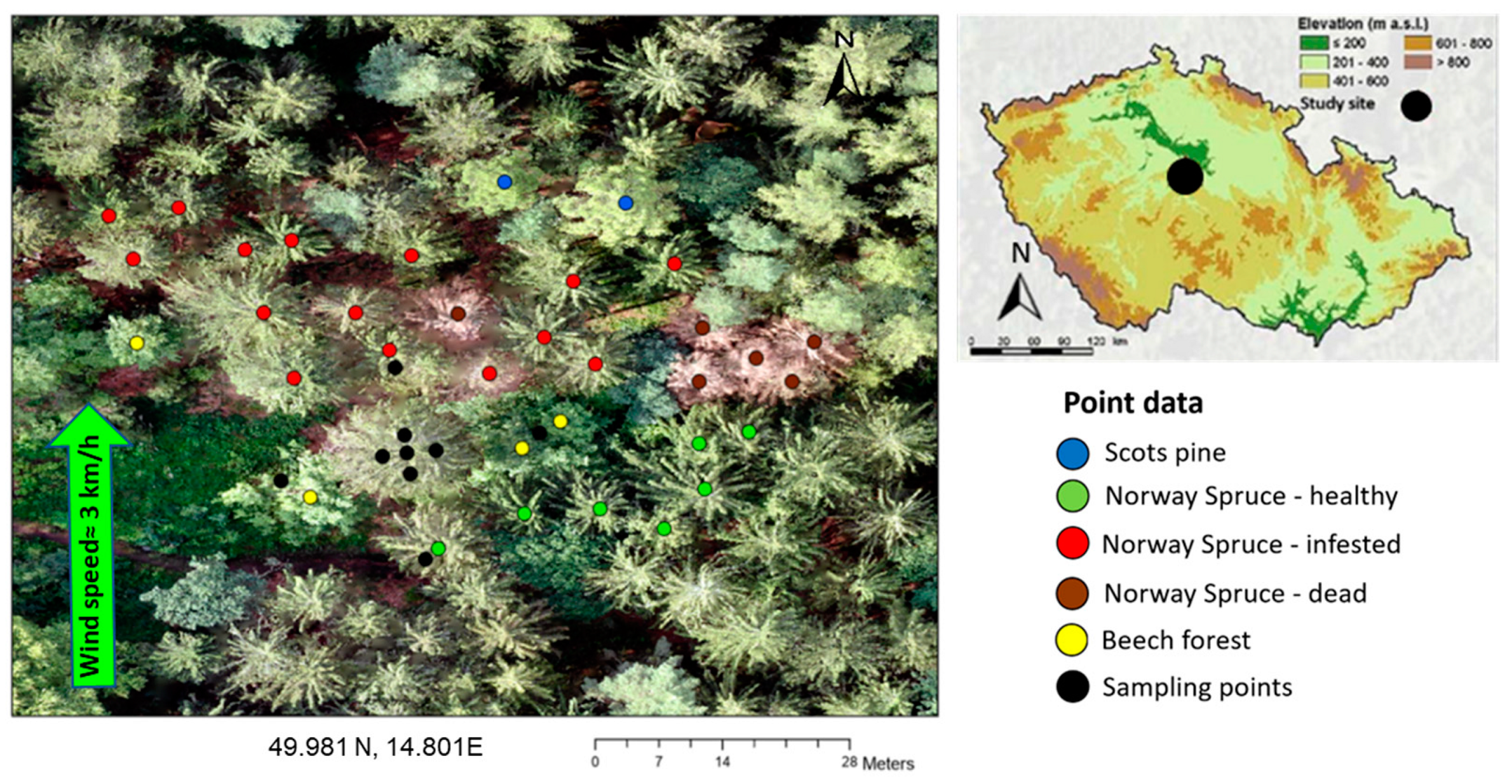
2.2. Field Collection of Distributed VOCs Emitted by the Ips typographus Naturally Infested Picea abies
2.3. Chemical Analyses of SMPE Fiber and Cartridges via Gas Chromatography-Flame Ionization Detection (GC-FID)
2.4. Determination of Relative Quantities of α-Pinene via SPME and Absolute Quantities Sorbed to Cartridges
2.5. Statistical Analyses
3. Results
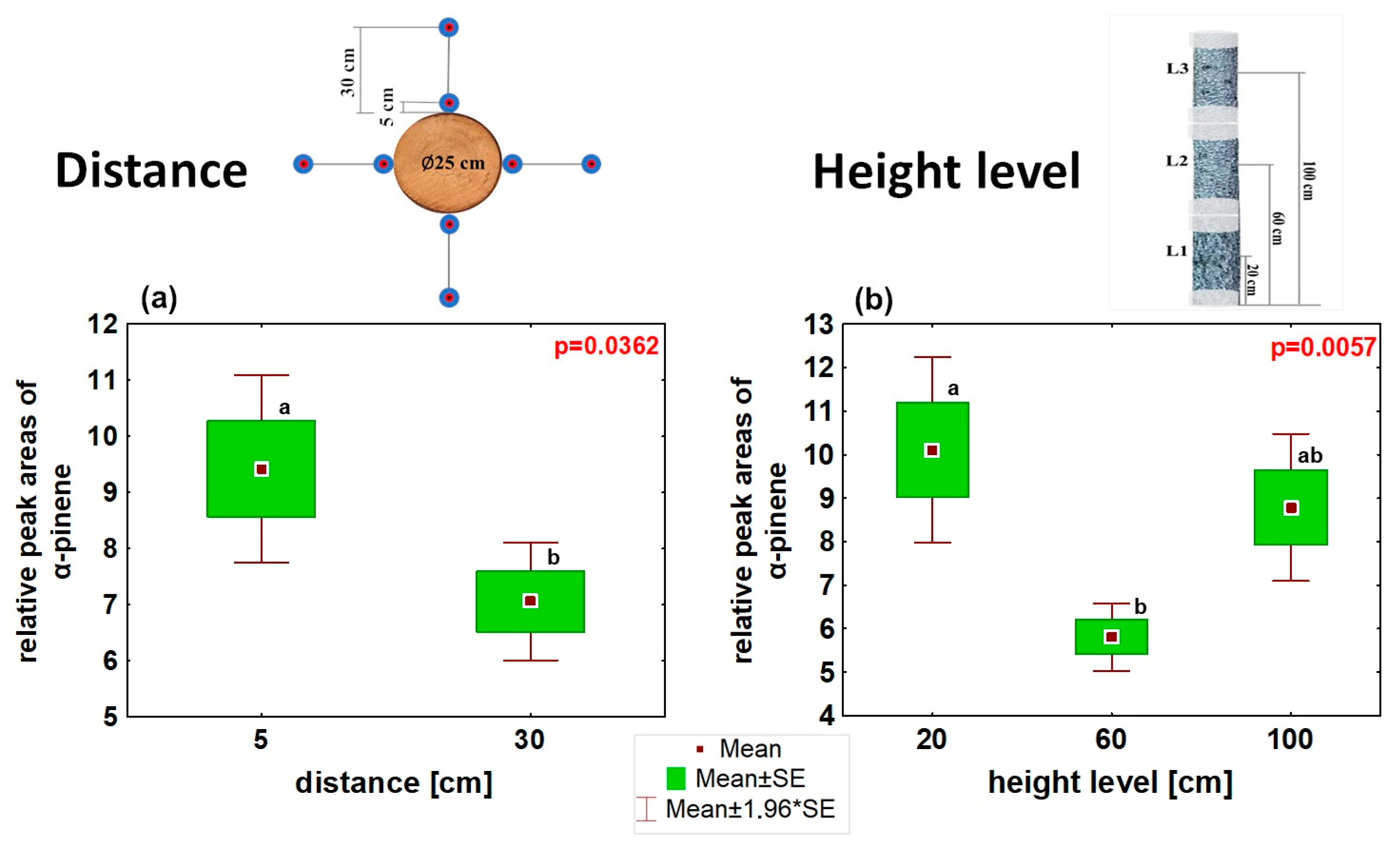
3.1. Optimization of VOC Collection to SPME in a Laboratory
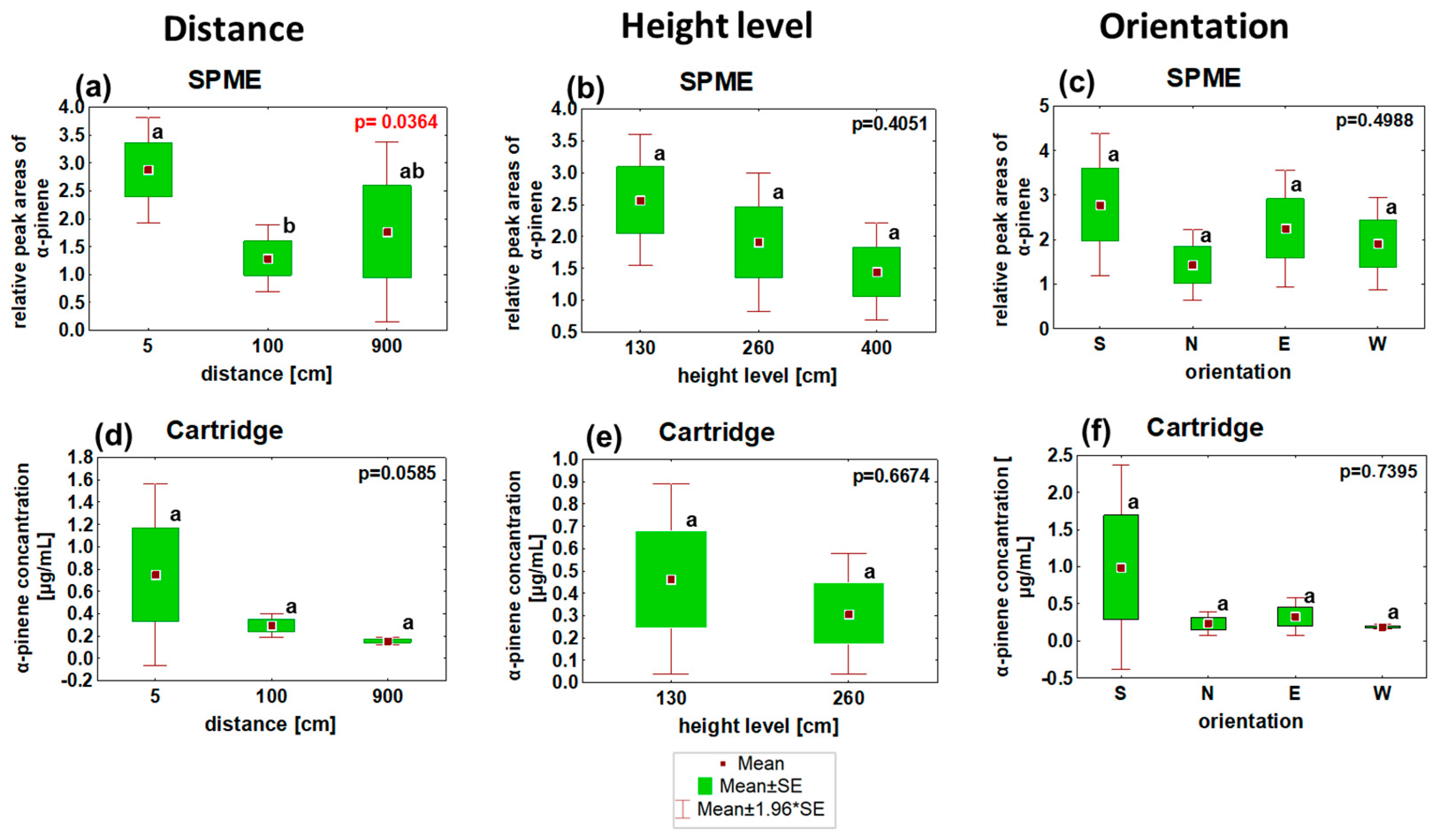
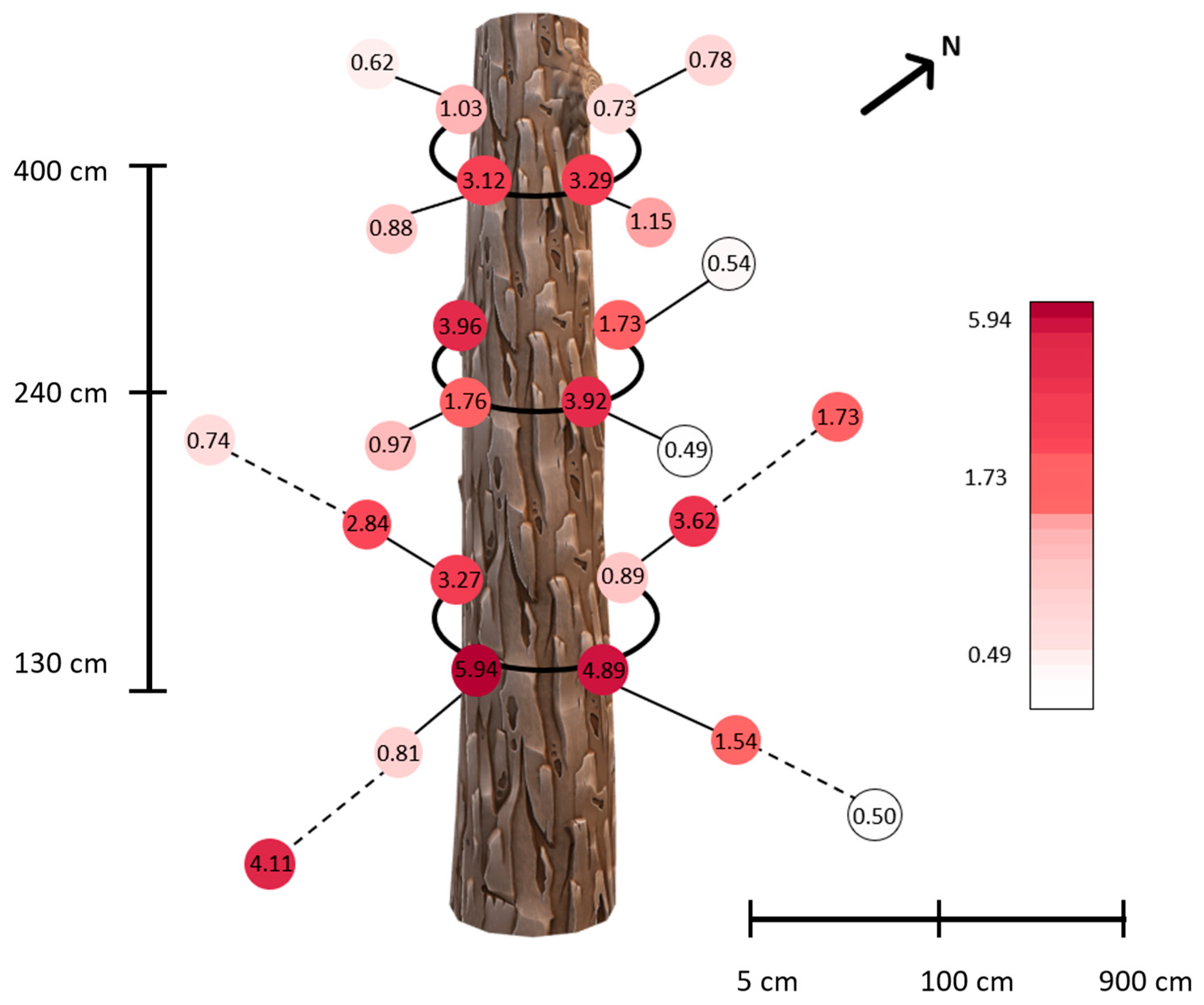
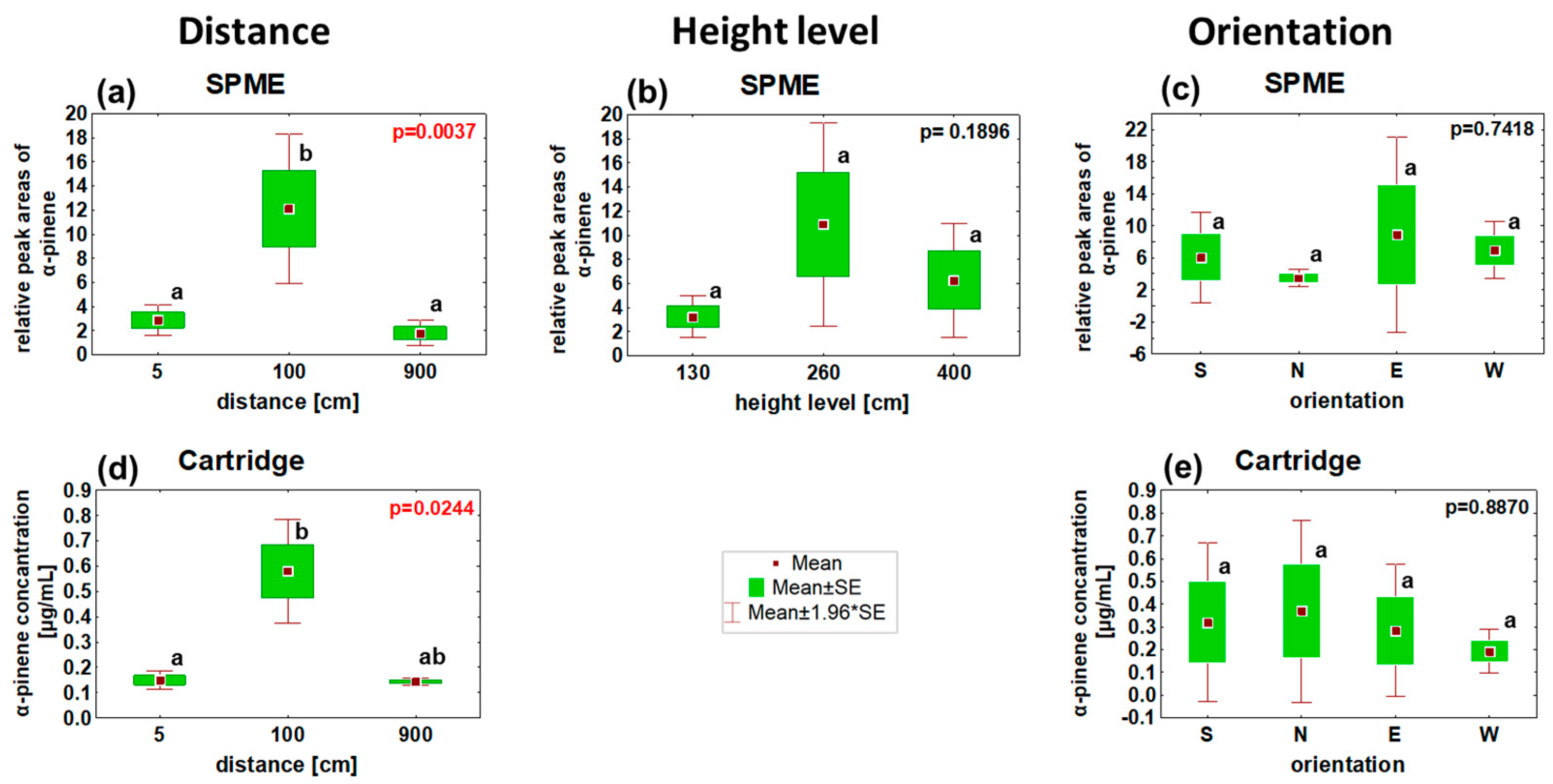
3.2. α-Pinene Spatial Distribution around Norway Spruce Infested by Ips typographus for Two Weeks
4. Discussion
Limitation of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Biedermann, P.H.W.; Müller, J.; Grégoire, J.C.; Gruppe, A.; Hagge, J.; Hammerbacher, A.; Hofstetter, R.W.; Kandasamy, D.; Kolarik, M.; Kostovcik, M.; et al. Bark Beetle Population Dynamics in the Anthropocene: Challenges and Solutions. Trends Ecol. Evol. 2019, 34, 914–924. [Google Scholar] [CrossRef]
- Hlásny, T.; König, L.; Krokene, P.; Lindner, M.; Montagné-Huck, C.; Müller, J.; Qin, H.; Raffa, K.F.; Schelhaas, M.J.; Svoboda, M.; et al. Bark Beetle Outbreaks in Europe: State of Knowledge and Ways Forward for Management. Curr. For. Rep. 2021, 7, 138–165. [Google Scholar] [CrossRef]
- Hlásny, T.; Zimová, S.; Merganičová, K.; Štěpánek, P.; Modlinger, R.; Turčáni, M. Devastating Outbreak of Bark Beetles in the Czech Republic: Drivers, Impacts, and Management Implications. For. Ecol. Manag. 2021, 490, 119075. [Google Scholar] [CrossRef]
- Hlásny, T.; Barka, I.; Merganičová, K.; Křístek, Š.; Modlinger, R.; Turčáni, M.; Marušák, R. A New Framework for Prognosing Forest Resources under Intensified Disturbance Impacts: Case of the Czech Republic. For. Ecol. Manag. 2022, 523, 120483. [Google Scholar] [CrossRef]
- Kautz, M.; Schopf, R.; Ohser, J. The “Sun-Effect”: Microclimatic Alterations Predispose Forest Edges to Bark Beetle Infestations. Eur. J. For. Res. 2013, 132, 453–465. [Google Scholar] [CrossRef]
- Huo, L.; Persson, H.J.; Lindberg, E. Remote Sensing of Environment Early Detection of Forest Stress from European Spruce Bark Beetle Attack, and a New Vegetation Index: Normalized Distance Red & SWIR (NDRS). Remote Sens. Environ. 2021, 255, 112240. [Google Scholar] [CrossRef]
- Van Der Velde, I.R.; Van Der Werf, G.R.; Houweling, S.; Maasakkers, J.D.; Borsdorff, T.; Landgraf, J.; Tol, P.; Van Kempen, T.A.; Van Hees, R.; Hoogeveen, R.; et al. Vast CO2 Release from Australian Fires in 2019–2020 Constrained by Satellite. Nature 2021, 597, 366–369. [Google Scholar] [CrossRef]
- Stříbrská, B.; Hradecký, J.; Čepl, J.; Modlinger, R.; Tomášková, I.; Jirošová, A. Physiological and Biochemical Indicators in Norway Spruces Freshly Infested by Ips typographus: Potential for Early Detection Methods. Front. For. Glob. Chang. 2023, 6, 1197229. [Google Scholar] [CrossRef]
- Hüttnerová, T.; Paczkowski, S.; Neubert, T.; Jirošová, A.; Surový, P. Comparison of Individual Sensors in the Electronic Nose for Stress Detection in Forest Stands. Sensors 2023, 23, 2001. [Google Scholar] [CrossRef]
- Paczkowski, S.; Datta, P.; Irion, H.; Paczkowska, M.; Habert, T.; Pelz, S.; Jaeger, D. Evaluation of Early Bark Beetle Infestation Localization by Drone-Based Monoterpene Detection. Forests 2021, 12, 228. [Google Scholar] [CrossRef]
- Vošvrdová, N.; Johansson, A.; Turčáni, M.; Jakuš, R.; Tyšer, D.; Schlyter, F.; Modlinger, R. Dogs Trained to Recognise a Bark Beetle Pheromone Locate Recently Attacked Spruces Better than Human Experts. For. Ecol. Manag. 2023, 528, 120626. [Google Scholar] [CrossRef]
- Johansson, A.; Birgersson, G.; Schlyter, F. Using Synthetic Semiochemicals to Train Canines to Detect Bark Beetle–Infested Trees. Ann. For. Sci. 2019, 76, 1–10. [Google Scholar] [CrossRef]
- Boone, C.K.; Aukema, B.H.; Bohlmann, J.; Carroll, A.L.; Raffa, K.F. Efficacy of Tree Defense Physiology Varies with Bark Beetle Population Density: A Basis for Positive Feedback in Eruptive Species. Can. J. For. Res. 2011, 41, 1174–1188. [Google Scholar] [CrossRef]
- Franceschi, V.R.; Krokene, P.; Christiansen, E.; Krekling, T. Anatomical and Chemical Defenses of Conifer Bark against Bark Beetles and Other Pests. New Phytol. 2005, 167, 353–376. [Google Scholar] [CrossRef] [PubMed]
- Borg-Karlson, A.K.; Lindström, M.; Norin, T.; Persson, M.; Valterová, I. Enantiomeric Composition of Monoterpene Hydrocarbons in Different Tissues of Norway Spruce, Picea Ab/Es (L) Karst. A Multi-Dimensional Gas Chromatography Study. Acta Chem. Scand. 1993, 47, 138–144. [Google Scholar] [CrossRef]
- Netherer, S.; Kandasamy, D.; Jirosová, A.; Kalinová, B.; Schebeck, M.; Schlyter, F. Interactions among Norway Spruce, the Bark Beetle Ips typographus and Its Fungal Symbionts in Times of Drought. J. Pest Sci. 2021, 94, 591–614. [Google Scholar] [CrossRef] [PubMed]
- Stříbrská, B.; Hradecký, J.; Čepl, J.; Tomášková, I.; Jakuš, R.; Modlinger, R.; Netherer, S.; Jirošová, A. Forest Margins Provide Favourable Microclimatic Niches to Swarming Bark Beetles, but Norway Spruce Trees Were Not Attacked by Ips typographus Shortly after Edge Creation in a Field Experiment. For. Ecol. Manag. 2022, 506, 119950. [Google Scholar] [CrossRef]
- Lerdau, M.; Litvak, M.; Palmer, P.; Monson, R. Controls over Monoterpene Emissions from Boreal Forest Conifers. Tree Physiol. 1997, 17, 563–569. [Google Scholar] [CrossRef]
- Juráň, S.; Pallozzi, E.; Guidolotti, G.; Fares, S.; Šigut, L.; Calfapietra, C.; Alivernini, A.; Savi, F.; Večeřová, K.; Křůmal, K.; et al. Fluxes of Biogenic Volatile Organic Compounds above Temperate Norway Spruce Forest of the Czech Republic. Agric. For. Meteorol. 2017, 232, 500–513. [Google Scholar] [CrossRef]
- Baier, P.; Bader, R.; Rosner, S. Monoterpene Content and Monoterpene Emission of Norway Spruce (Pices abies L. Karst.) Bark in Relation to Primary Attraction of Bark Beetles (Col. Scolytidae). In Physiology and Genetics of Tree-Phytophage Interactions; Lieutier, F., Mattson, W.J., Wagner, M.R., Eds.; Les Colloques de l’INRA: Gujan, France, 1999; Volume 90, pp. 249–259. [Google Scholar]
- Marešová, J.; Majdák, A.; Jakuš, R.; Hradecký, J.; Kalinová, B.; Blaženec, M. The Short-Term Effect of Sudden Gap Creation on Tree Temperature and Volatile Composition Profiles in a Norway Spruce Stand. Trees 2020, 34, 1397–1409. [Google Scholar] [CrossRef]
- Amin, H.; Atkins, P.T.; Russo, R.S.; Brown, A.W.; Sive, B.; Hallar, A.G.; Hu, K.E. Effect of Bark Beetle Infestation on Secondary Organic Aerosol Precursor Emissions. Environ. Sci. Technol. 2012, 46, 5696–5703. [Google Scholar] [CrossRef] [PubMed]
- Joensuu, J.; Altimir, N.; Hakola, H.; Rostás, M.; Raivonen, M.; Vestenius, M. Role of Needle Surface Waxes in Dynamic Exchange of Mono- and Sesquiterpenes. Atmos. Chem. Phys. 2016, 16, 7813–7823. [Google Scholar] [CrossRef]
- Kivimäenpää, M.; Magsarjav, N.; Ghimire, R.; Markkanen, J.; Heijari, J.; Vuorinen, M.; Holopainen, J.K. Influence of Tree Provenance on Biogenic VOC Emissions of Scots Pine (Pinus sylvestris) Stumps. Atmos. Environ. 2012, 60, 477–485. [Google Scholar] [CrossRef]
- Cataldo, F.; Ursini, O.; Lilla, E.; Angelini, G.; Cataldo, F.; Ursini, O.; Lilla, E.; Angelini, G. Ozone: Science & Engineering Turpentine Oil Studied by Chirooptical Methods; Some Ozonolysis of α-Pinene, β-Pinene, d-and l-Turpentine Oil Studied by Chirooptical Methods; Some Implications on the Atmospheric Chemistry of Biogenic Volatile Organic Compo. Ozone Sci. Eng. 2010, 9512, 274–285. [Google Scholar] [CrossRef]
- Yu, H.; Holopainen, J.K.; Kivimäenpää, M.; Virtanen, A.; Blande, J.D. Potential of Climate Change and Herbivory to Affect the Release and Atmospheric Reactions of BVOCs from Boreal and Subarctic Forests. Molecules 2021, 26, 2283. [Google Scholar] [CrossRef] [PubMed]
- Jaakkola, E.; Gärtner, A.; Jönsson, A.M.; Olsson, K.L.P.; Holst, T. Spruce Bark Beetle (Ips typographus) Infestation Cause up to 700 Times Higher Bark BVOC Emission Rates from Norway Spruce (Picea abies). Biogeosci. Discuss 2022, 3, 1–32. [Google Scholar] [CrossRef]
- Ghimire, R.P.; Kivimäenpää, M.; Blomqvist, M.; Holopainen, T.; Lyytikäinen-Saarenmaa, P.; Holopainen, J.K. Effect of Bark Beetle (Ips typographus L.) Attack on Bark VOC Emissions of Norway Spruce (Picea abies Karst.) Trees. Atmos. Environ. 2016, 126, 145–152. [Google Scholar] [CrossRef]
- Raffa, K.F.; Andersson, M.N.; Schlyter, F. Host Selection by Bark Beetles: Playing the Odds in a High-Stakes Game. In Advances in Insect Physiology; Tittiger, C., Blomquist, G.J., Eds.; Academic Press: London, UK, 2016; Volume 50, pp. 1–74. ISBN 9780128027233. [Google Scholar]
- Erbilgin, N.; Powell, J.S.; Raffa, K.F. Effect of Varying Monoterpene Concentrations on the Response of Ips Pini (Coleoptera: Scolytidae) to Its Aggregation Pheromone: Implications for Pest Management and Ecology of Bark Beetles. Agric. For. Entomol. 2003, 5, 269–274. [Google Scholar] [CrossRef]
- Bohbot, J.D.; Vernick, S. He Emergence of Insect Odorant Receptor-Based Biosensors. Biosensors 2020, 10, 26. [Google Scholar] [CrossRef]
- Lehmanski, L.M.; Kandasamy, D.; Andersson, M.N.; Netherer, S.; Alves, E.G.; Huang, J.; Hartmann, H. Viewpoints Addressing a Century-Old Hypothesis – Do Pioneer Beetles of Ips typographus Use Volatile Cues to Find Suitable Host Trees? New Phytol. 2023, 238, 1762–1770. [Google Scholar] [CrossRef]
- Andersson, M.N.; Larsson, M.C.; Schlyter, F. Specificity and Redundancy in the Olfactory System of the Bark Beetle Ips typographus: Single-Cell Responses to Ecologically Relevant Odors. J. Insect Physiol. 2009, 55, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Tholl, D.; Boland, W.; Hansel, A.; Loreto, F.; Ro, U.S.R. Practical Approaches to Plant Volatile Analysis. Plant J. 2006, 45, 540–560. [Google Scholar] [CrossRef] [PubMed]
- Tholl, D.; Weinhold, A.; Röse, U.S.R. Practical Approaches to Plant Volatile Collection and Analysis. In Biology of Plant Volatiles; CRC Press: Boca Raton, FL, USA, 2020; p. 24. [Google Scholar]
- Malik, T.G.; Sahu, L.K.; Gupta, M.; Mir, B.A.; Gajbhiye, T.; Dubey, R.; McCormick, A.C.; Pandey, S.K. Environmental Factors Affecting Monoterpene Emissions from Terrestrial Vegetation. Plants 2023, 12, 3146. [Google Scholar] [CrossRef] [PubMed]
- Perreca, E.; Gershenzon, J.; Eberl, F. Tree Volatiles. In Biology of Plant Volatiles; CRC Press: Boca Raton, FL, USA, 2020; p. 15. ISBN 9780429455612. [Google Scholar]
- Křůmal, K.; Mikuška, P.; Večeřová, K.; Urban, O.; Pallozzi, E.; Večeřa, Z. Wet Effluent Diffusion Denuder: The Tool for Determination of Monoterpenes in Forest. Talanta 2016, 153, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Amin, H.S.; Russo, R.S.; Sive, B.; Hoebeke, E.R.; Dodson, C.; Mccubbin, I.B.; Hallar, A.G.; Huff, K.E. Monoterpene Emissions from Bark Beetle Infested Engelmann Spruce Trees. Atmos. Environ. 2013, 72, 130–133. [Google Scholar] [CrossRef]
- Birgersson, G.; Bergström, G. Volatiles Released from Individual Spruce Bark Beetle Entrance Holes Quantitative Variations during the First Week of Attack. J. Chem. Ecol. 1989, 15, 2465–2483. [Google Scholar] [CrossRef] [PubMed]
- Giunta, A.D.; Runyon, J.B.; Jenkins, M.J.; Teich, M. Volatile and Within-Needle Terpene Changes to Douglas-Fir Trees Associated with Douglas-Fir Beetle (Coleoptera: Curculionidae) Attack. Environ. Entomol. 2016, 45, 920–929. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Kim, K.; Jung, I.; Lee, S.; Kim, J. Seasonal Variations and Emission Fluxes of Monoterpene Emitted from Coniferous Trees in East Asia: Focused on Pinus Rigida and Pinus Koraiensis. J. Atmos. Chem. 2015, 72, 27–41. [Google Scholar] [CrossRef]
- Komenda, M.; Koppmann, R.; Wald, H. Monoterpene Emissions from Scots Pine (Pinus sylvestris): Field Studies of Emission Rate Variabilities. J. Geophys. Res. Atmos. 2002, 107, ACH-1. [Google Scholar] [CrossRef]
- Janson, R.; Serves, C. De Acetone and Monoterpene Emissions from the Boreal Forest in Northern Europe. Atmos. Environ. 2001, 35, 4629–4637. [Google Scholar] [CrossRef]
- Raveane, L.; Tisato, F.; Isak, I.; Traldi, P. Analyses of BioVOCs Variation Related to Vegetation Predominance in the Natural Park of Ampezzo Dolomites, UNESCO World Heritage Area of Dolomites. J. For. Res. 2013, 24, 439–448. [Google Scholar] [CrossRef]
- Graus, M.; Hansel, A.; Wisthaler, A.; Lindinger, C.; Forkel, R.; Hauff, K.; Steinbrecher, R. A Relaxed-Eddy-Accumulation Method for the Measurement of Isoprenoid Canopy-Fluxes Using an Online Gas-Chromatographic Technique and PTR-MS Simultaneously. Atmos. Environ. 2006, 40, 43–54. [Google Scholar] [CrossRef]
- Pallozzi, E.; Guidolotti, G.; Ciccioli, P.; Brilli, F.; Feil, S.; Calfapietra, C. Does the Novel Fast-GC Coupled with PTR-TOF-MS Allow a Significant Advancement in Detecting VOC Emissions from Plants? Agric. For. Meteorol. 2016, 216, 232–240. [Google Scholar] [CrossRef]
- Ruuskanen, T.M.; Kolari, P.; Bäck, J.; Kulmala, M.; Rinne, J.; Hakola, H.; Taipale, R.; Raivonen, M.; Altimir, N.; Hari, P. On-Line Field Measurements of Monoterpene Emissions from Scots Pine by Proton-Transfer-Reaction Mass Spectrometry. Boreal. Environ. Res. 2005, 10, 553–567. [Google Scholar]
- Bouvier-Brown, N.C.; Goldstein, A.H.; Gilman, J.B.; Kuster, W.C.; De Gouw, J.A. In-Situ Ambient Quantification of Monoterpenes, Sesquiterpenes, and Related Oxygenated Compounds during BEARPEX 2007: Implications for Gas-and Particle-Phase Chemistry. Atmos. Chem. Phys. 2009, 9, 5505–5518. [Google Scholar] [CrossRef]
- Faiola, C.; Taipale, D. Impact of Insect Herbivory on Plant Stress Volatile Emissions from Trees: A Synthesis of Quantitative Measurements and Recommendations for Future Research. Atmos. Environ. X 2020, 5, 100060. [Google Scholar] [CrossRef]
- Sousa, M.; Birgersson, G.; Karlsson Green, K.; Pollet, M.; Becher, P.G. Odors Attracting the Long-Legged Predator Medetera Signaticornis Loew to Ips typographus L. Infested Norway Spruce Trees. J. Chem. Ecol. 2023, 49, 451–464. [Google Scholar] [CrossRef]
- Bergström, R.; Hallquist, M.; Simpson, D.; Wildt, J.; Mentel, T.F. Biotic Stress: A Significant Contributor to Organic Aerosol in Europe? Atmos. Chem. Phys. 2014, 14, 13643–13660. [Google Scholar] [CrossRef]
- Park, J.H.; Goldstein, A.H.; Timkovsky, J.; Fares, S.; Weber, R.; Karlik, J.; Holzinger, R. Eddy Covariance Emission and Deposition Flux Measurements Using Proton Transfer Reaction–Time of Flight–Mass Spectrometry (PTR-TOF-MS): Comparison with PTR-MS Measured Vertical Gradients and Fluxes. Atmos. Chem. Phys. 2013, 13, 1439–1456. [Google Scholar] [CrossRef]
- Hakola, H.; Taipale, D.; Praplan, A.; Schallhart, S.; Thomas, S.; Tykkä, T.; Helin, A.; Bäck, J.; Hellén, H. Emissions of Volatile Organic Compounds from Norway Spruce and Potential Atmospheric Impacts. Front. For. Glob. Chang. 2023, 6, 1116414. [Google Scholar] [CrossRef]
- NJ Health Alpha-PINENE. Available online: https://nj.gov/health/eoh/rtkweb/documents/fs/0052.pdf (accessed on 11 December 2023).
- Baker, J.; Arey, J.; Atkinson, R. Kinetics of the Gas-Phase Reactions of OH Radicals, NO3 Radicals and O3 with Three C 7-Carbonyls Formed from the Atmospheric Reactions of Myrcene, Ocimene and Terpinolene. J. Atmos. Chem. 2004, 48, 241–260. [Google Scholar] [CrossRef]
- Guenther, A. Biological and Chemical Diversity of Biogenic Volatile Organic Emissions into the Atmosphere. Int. Sch. Res. Not. 2013, 786290. [Google Scholar] [CrossRef]
- Duan, C.; Li, J.; Zhang, Y.; Ding, K.; Geng, X. Portable Instruments for On-Site Analysis of Environmental Samples. Trends Anal. Chem. 2022, 154, 116653. [Google Scholar] [CrossRef]
- Hummel, H.E.; Miller, T.A. Techniques in Pheromone Research; Springer Science & Business Media: New York, NY, USA, 2012; ISBN 978-1-4612-9743-7. [Google Scholar]
- McQuate, S. The Smellicopter Is an Obstacle-Avoiding Drone That Uses a Live Moth Antenna to Seek out Smells. Available online: https://www.washington.edu/news/2020/12/07/smellicopter-avoids-obstacles-uses-live-moth-antenna-to-smell/ (accessed on 11 December 2023).
- Pajares, G. Overview and Current Status of Remote Sensing Applications Based on Unmanned Aerial Vehicles (UAVs). Photogramm. Eng. Remote. Sens. 2015, 81, 281–330. [Google Scholar] [CrossRef]
- Smigaj, M.; Gaulton, R.; Suárez, J.C.; Barr, S.L. Forest Ecology and Management Canopy Temperature from an Unmanned Aerial Vehicle as an Indicator of Tree Stress Associated with Red Band Needle Blight Severity. For. Ecol. Manag. 2019, 433, 699–708. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Species/Stress Occasions | Sampling Specification | Compound (Unit) | Technical Parameters (Sorbent; Amount; Flow Rate) | Time of Sorption | Analytical Method | Source |
|---|---|---|---|---|---|---|
| Picea abies/intact forest | Stem (not specify) | Individual monoterpenes | Tenax TA, (35/60) | 30 min | GC-MS | [38] |
| 5 m above the ground; stainless steel TD tubes | α-pinene 3.07 ± 0.25 ppbv | 200 mg; 200 mL/min | ||||
| Picea abies/attacked trees | Stem | Individual monoterpenes | Tenax-TA a Carbopack-B, (60/80) | 60 min | GC-MS | [28] |
| 1.3 m above the ground; surrounded PET (25 × 38 cm) encloser | α-pinene 62.8 ± 23.6 μg h−1 m−2 bark area | 100/100 mg; 200 mL/min | ||||
| Picea engelmannii/attacked trees | Stem | Individual monoterpenes | Porapak Q | 120 min | GC-MS | [39] |
| 0.5 to 1.5 m above the ground; the trunk by dynamic sampling <1 cm from stem (sorbent trap) | α-pinene 8.5 ± 2.1 ng L−1 | 110 mg; 400 mL/min | ||||
| Picea abies/attacked trees on forest edge | Stem 3 h | Verbenone (ng/3 h): α-pinene (μg/3 h) | Porapak Q, (80/100) | 180 min | GC-MS | [40] |
| 1–2 m above the ground; sanitized T gllas tube | 0.6 (ng/3 h): (μg/3 h) | 70 mg; 20 mL/min | ||||
| Pseudotsuga menziesii/attacked trees | Branch | Individual VOCs | HayeSep-Q | 30 min | GC-MS | [41] |
| 1.5 m above the ground; Teflon bag (50 × 75 cm) | α-pinene 813.9 ± 482.29 ng h−1 g−1 fresh weight | 30 mg; 500 mL/min | ||||
| Pinus rigida and Pinus koraiensis/intact forest | Branch | Total monoterpenes emission (μgC gdw−1 h−1) | Tenax TA, (60/80) and Carbotrap, (20/40) | 15–60 min | GC-MS GC-FID | [42] |
| 20 L Tedlar bag | Pinus rigida 0.9 μgC gdw−1 h−1 Pinus koraiensis 0.4 μgC gdw−1 h−1 | 200 mg; 100–200 mL/min | ||||
| Pinus sylvestris/intact forest | Branch | Individual monoterpenes | Tenax TA, (60/80) and Carbotrap, (20/40) | 60 min | GC-MS GC-FID | [43] |
| canopy height; (FEP) copolymer foil (50 μm thickness) mounted in cylindrical frames | α-pinene 917 ± 58 ng h−1 g−1 (April) α-pinene 75 ± 12 ng h−1 g−1 (July) | 50–100 mg; 100 mL/min | ||||
| Pinus sylvestris and Picea abies/intact forest | Branch | Acetone and α-pinene (ng gdw−1 h−1) | Tenax TA | 12–40 min | GC-MS GC-FID | [44] |
| 18 L all-Teflon chamber made of 0.05 mm transparent FEP-Teflon film enclosing a 20–30 cm branch segment | α-pinene 80 ng gdw−1 h−1 Monoterpene emission 900 ± 640 ng C gdw−1 h−1 | 200 mg; 100 mL/min | ||||
| Picea abies/attacked trees | Stem | Individual VOCs (μg m−2 h −1) | Tenax TA and Carbograph 1TD | 30 min | GC-MS | [27] |
| 1.3 m above the ground; tree trunk chamber connected with PTFE tubing | α-pinene 911.14 μg m−2 h−1 | 200 mL/min | ||||
| Picea abies/stress from sun irradiation | Stem | Individual monoterpenes | SPME | 60 min (from 1 to 2 p.m.) | GC-MS | [17] |
| 3.5 m above the ground; aluminum chamber | sum of eight main MT 8.5 log10 VOC | |||||
| Picea abies/attacked trees | Stem | Individual monoterpenes | SPME | 60 min (from 1 to 2 p.m.) | GC-MS | [8] |
| 3.5 m above the ground; aluminum chamber | α-pinene 9.5 log10 sum peak area | |||||
| Lerosa NP forest/conifer forest; 6 different plots | open air | Individual VOCs | SPME | 300 min (from 10 a.m. to 3 p.m.) | GC-MS | [45] |
| 894 abundance relative to hexanal (%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stříbrská, B.; Moliterno, A.A.C.; Hüttnerová, T.; Leiner, M.; Surový, P.; Jirošová, A. Pilot Study of 3D Spatial Distribution of α-Pinene Emitted by Norway Spruce (L.) Karst Recently Infested by Ips typographus (L. 1758) (Coleoptera: Scolytinae). Forests 2024, 15, 10. https://doi.org/10.3390/f15010010
Stříbrská B, Moliterno AAC, Hüttnerová T, Leiner M, Surový P, Jirošová A. Pilot Study of 3D Spatial Distribution of α-Pinene Emitted by Norway Spruce (L.) Karst Recently Infested by Ips typographus (L. 1758) (Coleoptera: Scolytinae). Forests. 2024; 15(1):10. https://doi.org/10.3390/f15010010
Chicago/Turabian StyleStříbrská, Barbora, Antonioni Acacio Campos Moliterno, Tereza Hüttnerová, Martin Leiner, Peter Surový, and Anna Jirošová. 2024. "Pilot Study of 3D Spatial Distribution of α-Pinene Emitted by Norway Spruce (L.) Karst Recently Infested by Ips typographus (L. 1758) (Coleoptera: Scolytinae)" Forests 15, no. 1: 10. https://doi.org/10.3390/f15010010