

The Water Dynamics of Norway Spruce Stands Growing in Two Alpine Catchments in Austria



Abstract
:1. Introduction
- (i)
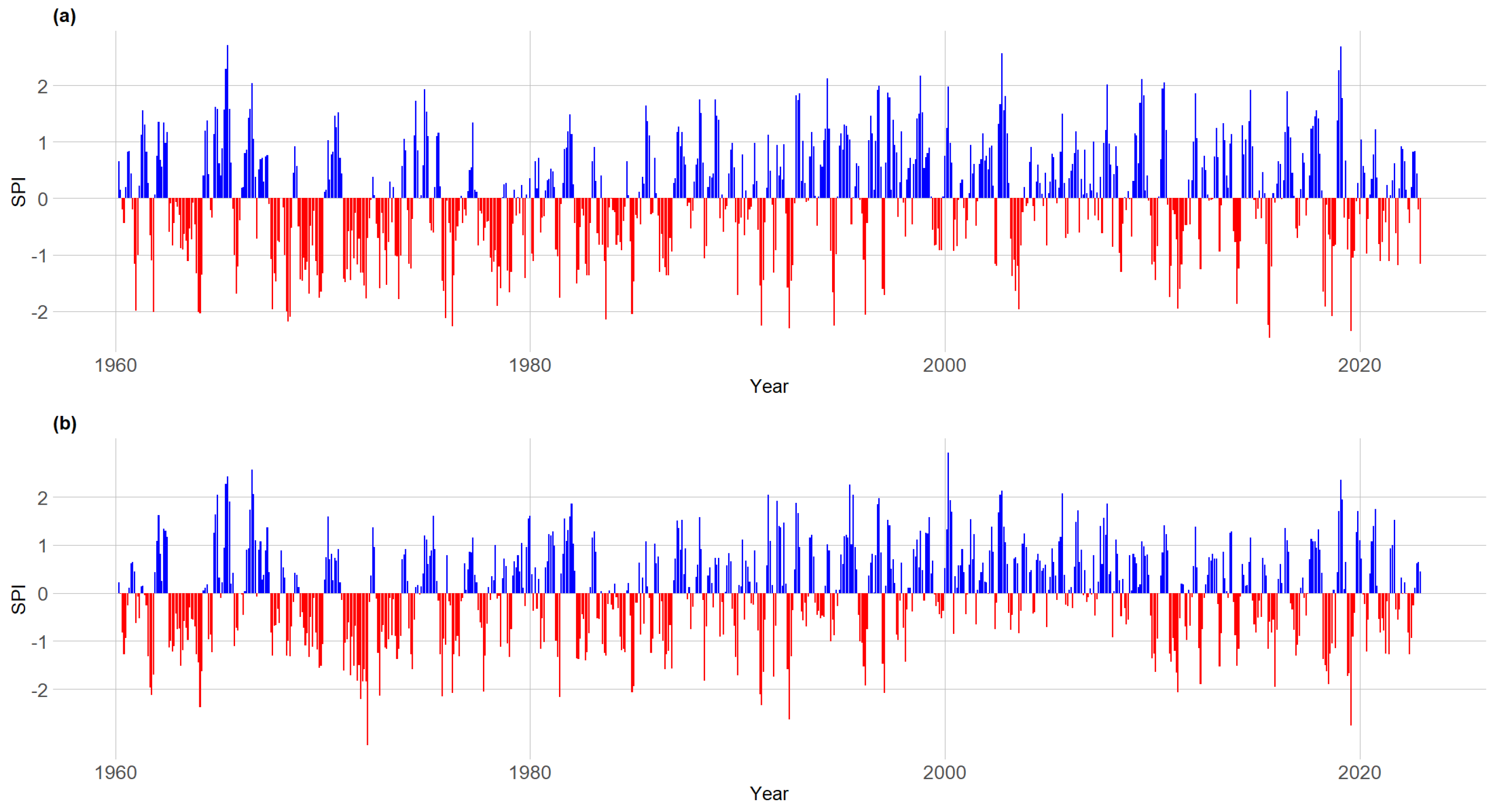
- Seasonal precipitation pattern and snow accumulation from 1960 to 2022.
- (ii)
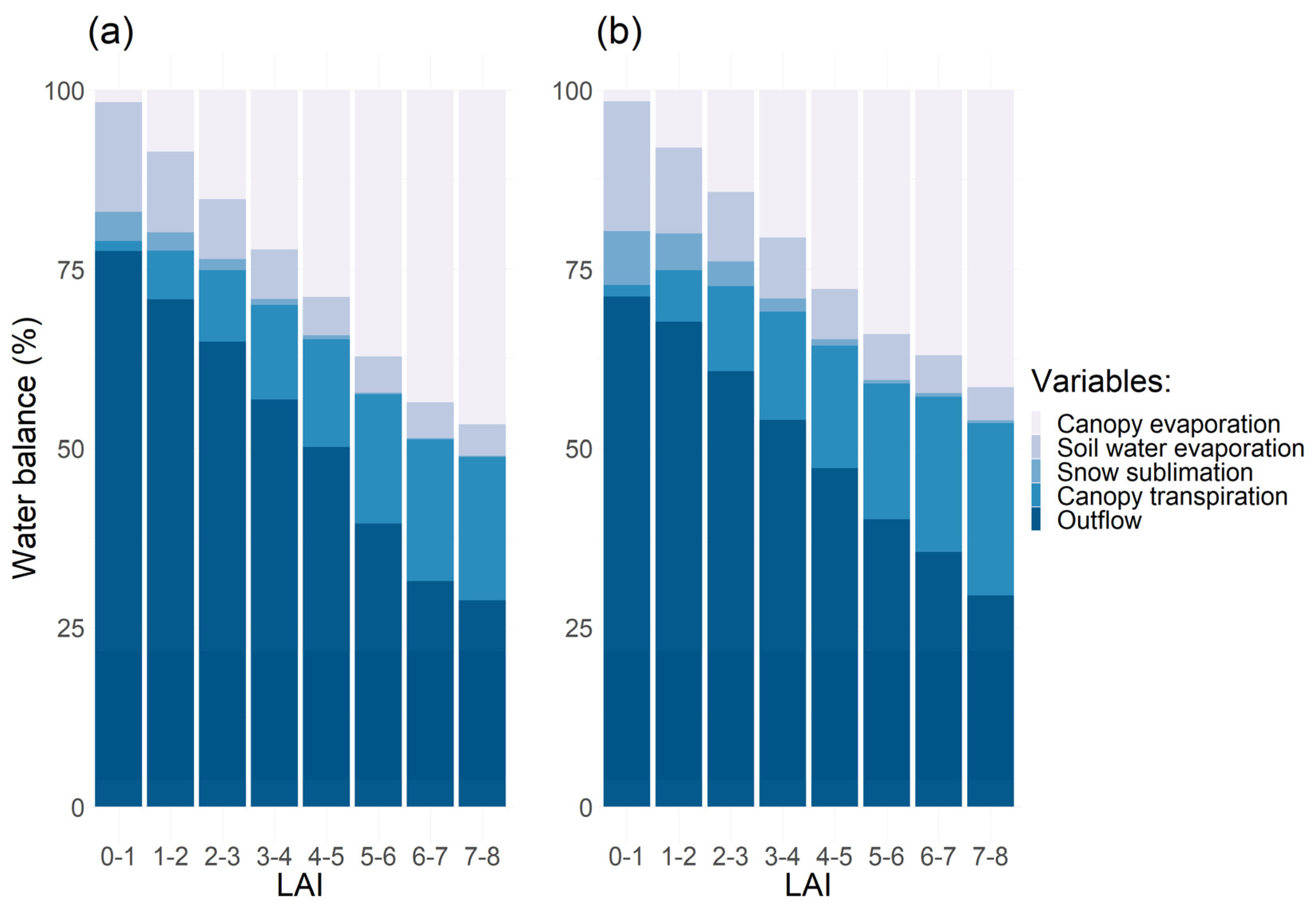
- Evapotranspiration, transpiration, and water use efficiency by leaf area index.
- (iii)
- The water balance of the Norway spruce stands within each catchment.
2. Materials and Methods
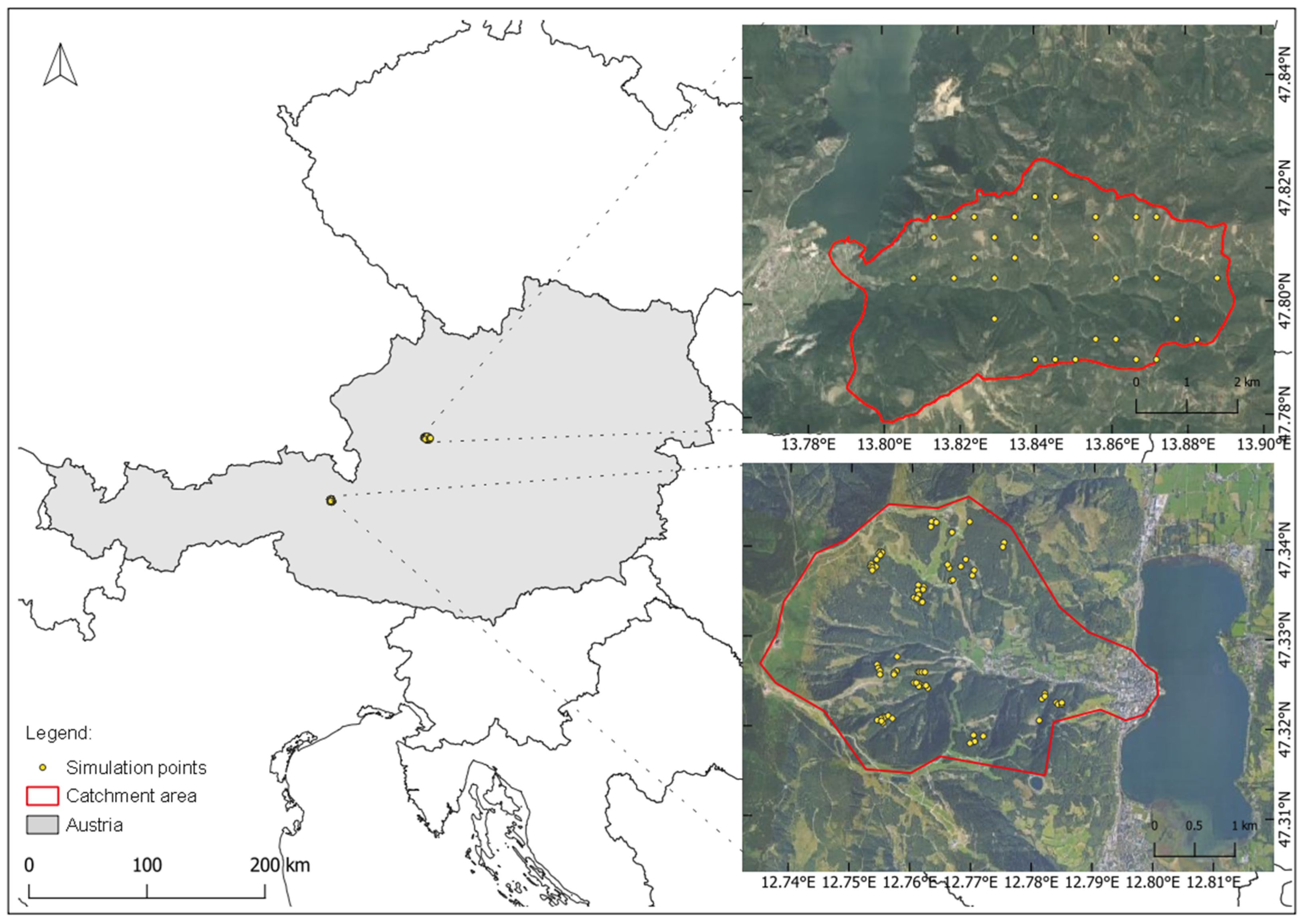
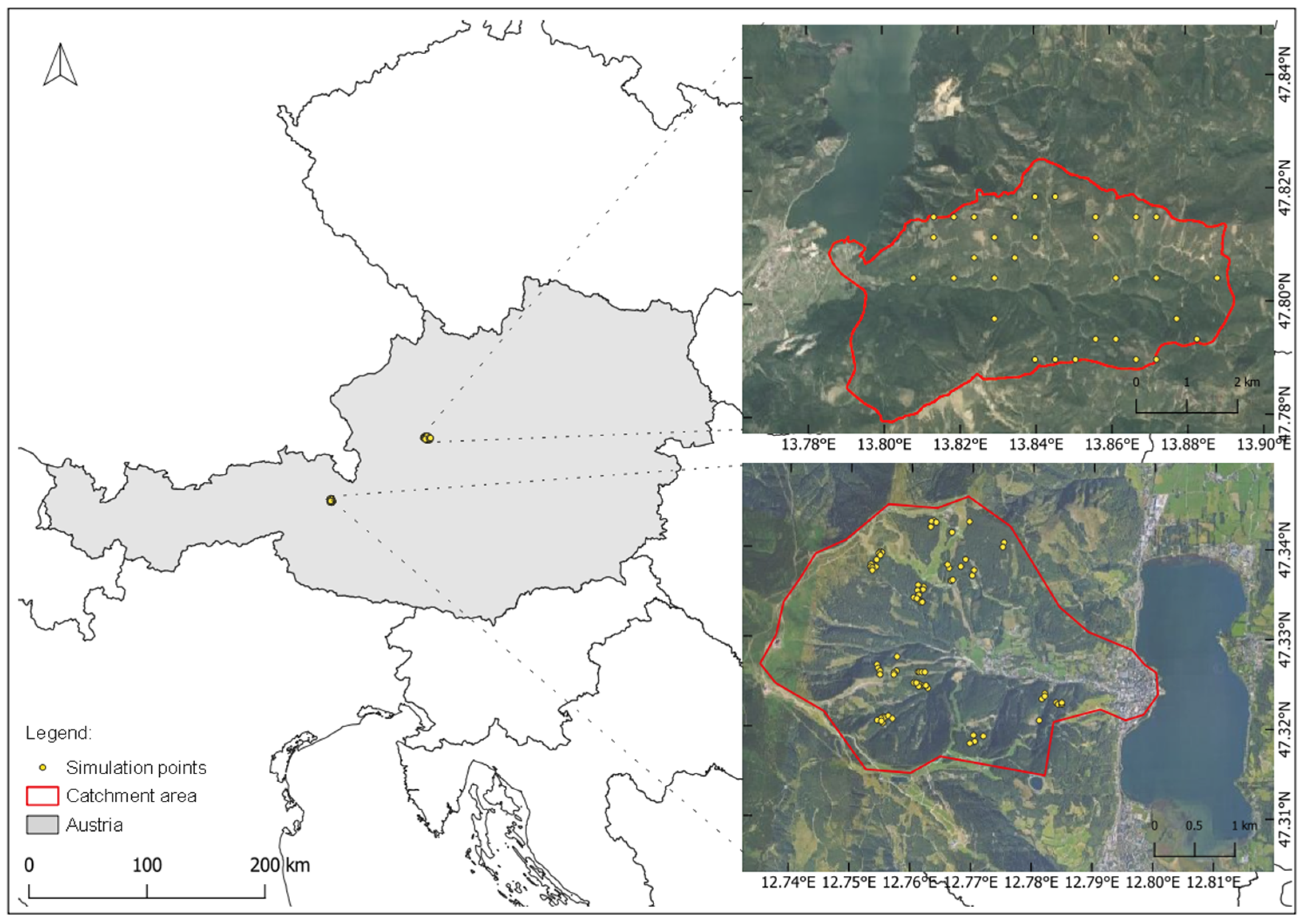
2.1. Study Area
2.2. Forest Data
2.3. Biome-BGC Ecosystem Modeling Description and Usage
2.3.1. Model Description
2.3.2. Simulation Procedure
2.3.3. Validation Procedure
3. Results
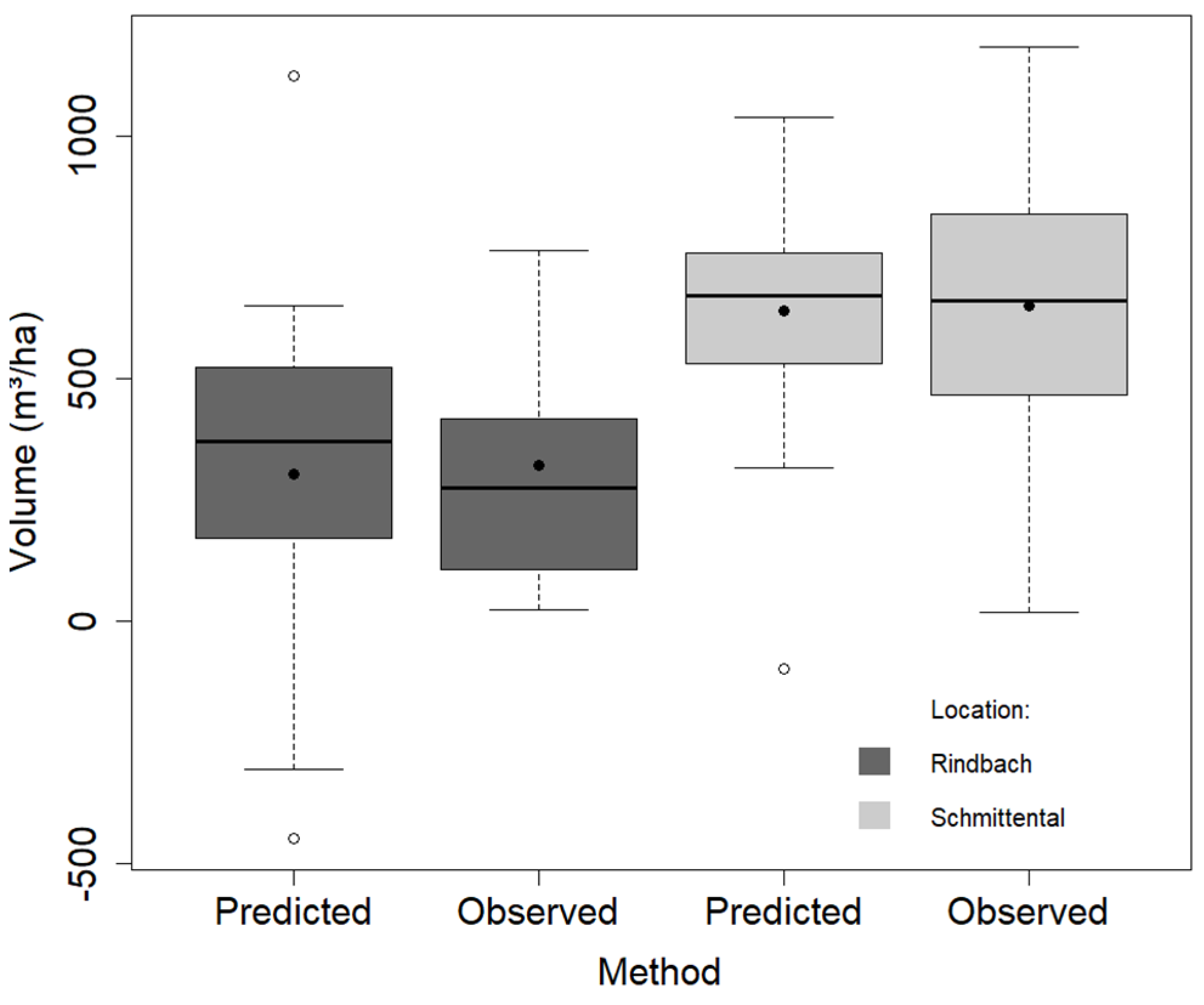
3.1. Model Validation
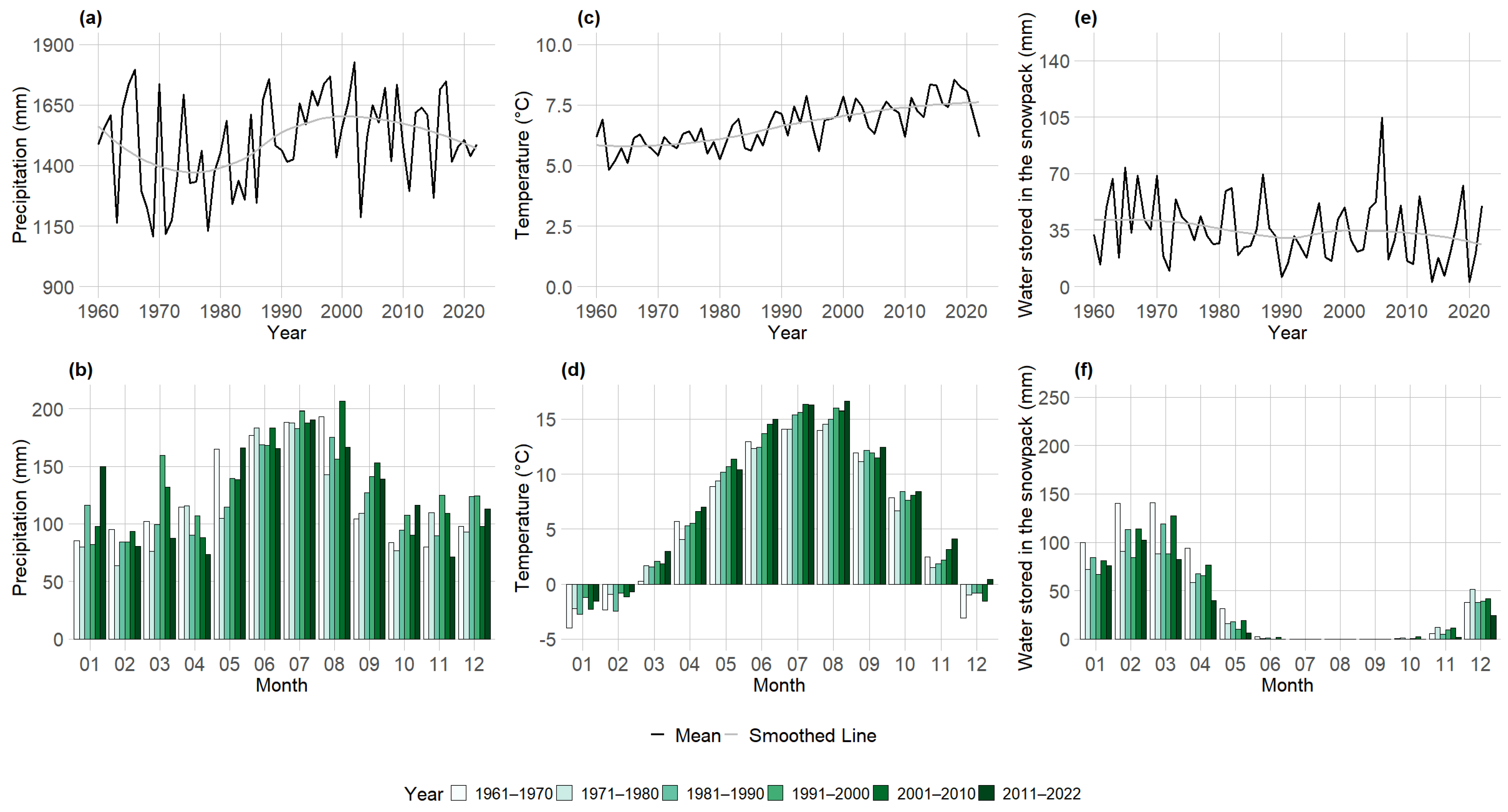
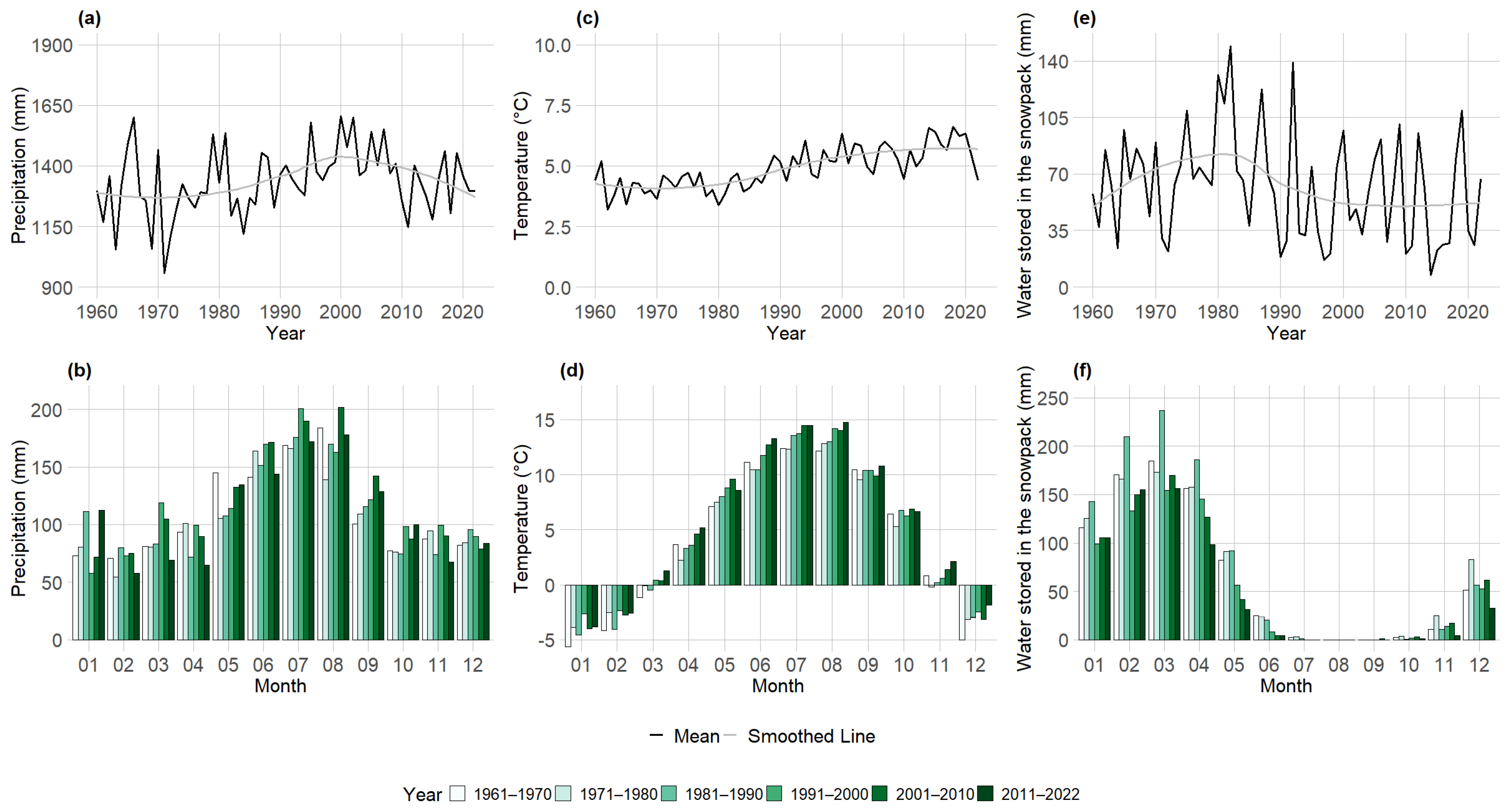
3.2. Seasonal Climate Pattern
3.3. Plant Water Use and Leaf Area Index
3.4. Impacts of Plant Water Use
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Myhre, G.; Alterskjær, K.; Stjern, C.W.; Hodnebrog, Ø.; Marelle, L.; Samset, B.H.; Sillmann, J.; Schaller, N.; Fischer, E.; Schulz, M.; et al. Frequency of Extreme Precipitation Increases Extensively with Event Rareness under Global Warming. Sci. Rep. 2019, 9, 16063. [Google Scholar] [CrossRef]
- Schmid, U.; Frehner, M.; Glatthorn, J.; Bugmann, H. ProForM: A Simulation Model for the Management of Mountain Protection Forests. Ecol. Model. 2023, 478, 110297. [Google Scholar] [CrossRef]
- Price, M.F.; Gratzler, G.; Duguma, L.A.; Kohler, T.; Maselli, D.; Romeo, R. Mountain Forests in a Changing World—Realizing Values, Addressing Challenges; FAO with the support of the Swiss Agency for Development and Cooperation, SDC; Food and Agriculture Organization of the United Nations: Roma, Italy, 2011. [Google Scholar]
- Pötzelsberger, E.; Hasenauer, H. Forest–Water Dynamics within a Mountainous Catchment in Austria. Nat. Hazards 2015, 77, 625–644. [Google Scholar] [CrossRef]
- Scheidl, C.; Heiser, M.; Kamper, S.; Thaler, T.; Klebinder, K.; Nagl, F.; Lechner, V.; Markart, G.; Rammer, W.; Seidl, R. The Influence of Climate Change and Canopy Disturbances on Landslide Susceptibility in Headwater Catchments. Sci. Total Environ. 2020, 742, 140588. [Google Scholar] [CrossRef] [PubMed]
- Caudullo, G.; Tinner, W.; de Rigo, D. Picea Abies in Europe: Distribution, Habitat, Usage and Threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publication Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Rehschuh, R.; Mette, T.; Menzel, A.; Buras, A. Soil Properties Affect the Drought Susceptibility of Norway Spruce. Dendrochronologia 2017, 45, 81–89. [Google Scholar] [CrossRef]
- Cavalli, J.P.; Reichert, J.M.; Rodrigues, M.F.; de Araújo, E.F. Composition and Functional Soil Properties of Arenosols and Acrisols: Effects on Eucalyptus Growth and Productivity. Soil Tillage Res. 2020, 196, 104439. [Google Scholar] [CrossRef]
- Hümann, M.; Schüler, G.; Müller, C.; Schneider, R.; Johst, M.; Caspari, T. Identification of Runoff Processes—The Impact of Different Forest Types and Soil Properties on Runoff Formation and Floods. J. Hydrol. 2011, 409, 637–649. [Google Scholar] [CrossRef]
- Sun, G.; Wei, X.; Hao, L.; Sanchis, M.G.; Hou, Y.; Yousefpour, R.; Zhang, Z. Forest Hydrology Modeling Tools for Watershed Management: A Review. For. Ecol. Manag. 2023, 530, 120755. [Google Scholar] [CrossRef]
- Arnold, J.G.; Srinivasan, R.; Muttiah, R.S.; Williams, J.R. Large Area Hydrologic Modeling and Assessment Part I: Model Development. J. Am. Water Resour. Assoc. 1998, 34, 73–89. [Google Scholar] [CrossRef]
- Šimůnek, J.; Šejna, M.; Saito, H.; Sakai, M.; van Genuchten, M.T. The HYDRUS-1D Software Package for Simulating the One-Dimensional Movement of Water, Heat, and Multiple Solutes in Variably-Saturated Media; University of California Riverside: Riverside, CA, USA, 2008. [Google Scholar]
- Thornton, P.E.; Law, B.E.; Gholz, H.L.; Clark, K.L.; Falge, E.; Ellsworth, D.S.; Goldstein, A.H.; Monson, R.K.; Hollinger, D.; Falk, M.; et al. Modeling and Measuring the Effects of Disturbance History and Climate on Carbon and Water Budgets in Evergreen Needleleaf Forests. Agric. For. Meteorol. 2002, 113, 185–222. [Google Scholar] [CrossRef]
- White, M.A.; Thornton, P.E.; Running, S.W.; Nemani, R.R. Parameterization and Sensitivity Analysis of the BIOME-BGC Terrestrial Ecosystem Model: Net Primary Production Controls. Earth Interact. 2000, 4, 1–84. [Google Scholar] [CrossRef]
- Pietsch, S.A.; Hasenauer, H.; Thornton, P.E. BGC-Model Parameters for Tree Species Growing in Central European Forests. For. Ecol. Manag. 2005, 211, 264–295. [Google Scholar] [CrossRef]
- Rupp, C.; Linner, M.; Mandl, G.W. Geologische Karte von Oberösterreich; Geologische Bundesanstalt: Vienna, Austria, 2011. [Google Scholar]
- Federal Research and Training Center for Forests, Natural Hazards and Landscape (BFW). “eBOD”—Digital Soil Map of Austria; BFW: Vienna, Austria, 2023. [Google Scholar]
- Umweltbundesamt GmbH. BORIS—Bodeninformationssystem; Umweltbundesamt GmbH: Dessau-Roßlau, Germany, 2023. [Google Scholar]
- Gabler, K.; Schadauer, K. Methods of the Austrian Forest Inventory 2000/0—Origins, Approaches, Design, Sampling, Data Models, Evaluation and Calculation of Standard Error; BFW: Vienna, Austria, 2008. [Google Scholar]
- Bitterlich, W. Die Winkelzählprobe. Allg. Forst Holzwirtsch. 1948, 59, 4–5. [Google Scholar] [CrossRef]
- Pietsch, S.A.; Hasenauer, H. Evaluating the Self-Initialization Procedure for Large-Scale Ecosystem Models. Glob. Change Biol. 2006, 12, 1658–1669. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration—Guidelines for Computing Crop Water Requirements—FAO Irrigation and Drainage Paper 56; FAO—Food and Agriculture Organization of the United Nations: Roma, Italy, 1998. [Google Scholar]
- Clapp, R.B.; Hornberger, G.M. Empirical Equations for Some Soil Hydraulic Properties. Water Resour. Res. 1978, 14, 601–604. [Google Scholar] [CrossRef]
- Cosby, B.J.; Hornberger, G.M.; Clapp, R.B.; Ginn, T.R. A Statistical Exploration of the Relationships of Soil Moisture Characteristics to the Physical Properties of Soils. Water Resour. Res. 1984, 20, 682–690. [Google Scholar] [CrossRef]
- Saxton, K.E.; Rawls, W.J.; Romberger, J.S.; Papendick, R.I. Estimating Generalized Soil-Water Characteristics from Texture. Soil Sci. Soc. Am. J. 1986, 50, 1032–1036. [Google Scholar] [CrossRef]
- Ballabio, C.; Panagos, P.; Monatanarella, L. Mapping Topsoil Physical Properties at European Scale Using the LUCAS Database. Geoderma 2016, 261, 110–123. [Google Scholar] [CrossRef]
- Hiederer, R. Mapping Soil Typologies—Spatial Decision Support Applied to the European Soil Database; European Commission: Luxembourg, 2013. [Google Scholar] [CrossRef]
- Lamarque, J.F.; Dentener, F.; McConnell, J.; Ro, C.U.; Shaw, M.; Vet, R.; Bergmann, D.; Cameron-Smith, P.; Dalsoren, S.; Doherty, R.; et al. Multi-Model Mean Nitrogen and Sulfur Deposition from the Atmospheric Chemistry and Climate Model Intercomparison Project (ACCMIP): Evaluation of Historical and Projected Future Changes. Atmos. Chem. Phys. 2013, 13, 7997–8018. [Google Scholar] [CrossRef]
- Tian, H.; Yang, J.; Lu, C.; Xu, R.; Canadell, J.G.; Jackson, R.B.; Arneth, A.; Chang, J.; Chen, G.; Ciais, P.; et al. The Global N2O Model Intercomparison Project. Bull. Am. Meteorol. Soc. 2018, 99, 1231–1251. [Google Scholar] [CrossRef]
- Enting, I.G.; Wigley, T.M.L.; Heimann, M. Future Emissions and Concentrations of Carbon Dioxide: Key Ocean/Atmosphere/Land Analyses; CSIRO: Brisbane City, QLD, Australia, 1994. [Google Scholar]
- Thornton, P.E.; Running, S.W.; White, M.A. Generating Surfaces of Daily Meteorological Variables over Large Regions of Complex Terrain. J. Hydrol. 1997, 190, 214–251. [Google Scholar] [CrossRef]
- Edwards, D.C.; McKee, T.B. Characteristics of 20th Century Drought in the United States at Multiple Time Scales; Climatology Report Number 97-2; Colorado State University: Fort Collins, CO, USA, 1997. [Google Scholar]
- Bella, I.E. A New Competition Model for Individual Trees. For. Sci. 1971, 17, 364–372. [Google Scholar]
- Hasper, T.B.; Wallin, G.; Lamba, S.; Hall, M.; Jaramillo, F.; Laudon, H.; Linder, S.; Medhurst, J.L.; Räntfors, M.; Sigurdsson, B.D.; et al. Water Use by Swedish Boreal Forests in a Changing Climate. Funct. Ecol. 2016, 30, 690–699. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand Characteristics | Rindbach N = 31 Plots | Schmittental N = 20 Plots | ||||||
|---|---|---|---|---|---|---|---|---|
| Min. | Max. | Mean | sd | Min. | Max. | Mean | sd | |
| Elevation (m) | 466 | 1379 | 1027 | 200.4 | 924 | 1690 | 1370 | 212.1 |
| Stand age (years) | 12 | 177 | 66 | 42 | 27 | 235 | 84 | 41 |
| DBH (cm) | 14.9 | 69.8 | 30.7 | 13.9 | 11.3 | 54.7 | 33.7 | 10.7 |
| Tree height (m) | 4.3 | 33.5 | 16.6 | 6.0 | 6.7 | 41.5 | 27.8 | 8.0 |
| Volume (m3/ha) | 21.8 | 766 | 304 | 218.7 | 16.8 | 1184.9 | 640 | 277.5 |
| BA (m2/ha) | 5.0 | 86.0 | 41.7 | 25.3 | 4.0 | 88.0 | 57.4 | 16.9 |
| N/ha | 20.8 | 2704.3 | 1005 | 784.1 | 85.5 | 7915.3 | 1296 | 1332.8 |
| CCF (%) | 13.0 | 481.3 | 204 | 139.4 | 29.4 | 1073.3 | 349 | 161.9 |
| Location | Intercept | Regression Coefficients | Standard Error of Estimates | Correlation Coefficient | F-Value | α |
|---|---|---|---|---|---|---|
| Rindbach | 190.63 | 0.35 | 191.5 | 0.26 | 9.54 | <0.01 |
| Schmittental | 130.91 | 0.80 | 233.04 | 0.27 | 28.11 | <0.01 |
| Variable (mm Year−1) | Minimum | Maximum | Mean | sd |
|---|---|---|---|---|
| Rindbach | ||||
| Canopy evaporation (mm/year) | 5.0 | 971.2 | 501.5 | 214.1 |
| Soil water evaporation (mm/year) | 18.6 | 313.8 | 96.2 | 50.9 |
| Snow sublimation (mm/year) | 0.0 | 171.4 | 10.7 | 19.4 |
| Transpiration (mm/year) | 4.5 | 361.8 | 243.5 | 87.7 |
| Outflow (mm/year) | 42.5 | 1874.3 | 645.6 | 286.2 |
| Schmittental | ||||
| Canopy evaporation (mm/year) | 4.3 | 873.1 | 402.5 | 161.2 |
| Soil water evaporation (mm/year) | 16.9 | 307.3 | 96.3 | 49.5 |
| Snow sublimation (mm/year) | 0.2 | 264.9 | 17.1 | 28.9 |
| Transpiration (mm/year) | 4.2 | 394.8 | 239.7 | 84.5 |
| Outflow (mm/year) | 113.6 | 1480.5 | 582.0 | 207.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Bastos, F.; Hasenauer, H. The Water Dynamics of Norway Spruce Stands Growing in Two Alpine Catchments in Austria. Forests 2024, 15, 35. https://doi.org/10.3390/f15010035
de Bastos F, Hasenauer H. The Water Dynamics of Norway Spruce Stands Growing in Two Alpine Catchments in Austria. Forests. 2024; 15(1):35. https://doi.org/10.3390/f15010035
Chicago/Turabian Stylede Bastos, Franciele, and Hubert Hasenauer. 2024. "The Water Dynamics of Norway Spruce Stands Growing in Two Alpine Catchments in Austria" Forests 15, no. 1: 35. https://doi.org/10.3390/f15010035
APA Stylede Bastos, F., & Hasenauer, H. (2024). The Water Dynamics of Norway Spruce Stands Growing in Two Alpine Catchments in Austria. Forests, 15(1), 35. https://doi.org/10.3390/f15010035