
Dynamic Shifts in Soil Fungal Functional Group Characteristics across Distinct Vegetation Types during Ecological Restoration in Degraded Red Soil Regions


 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Overview of the Experimental Site
2.2. Soil Sample Collection and Processing
2.3. Determination of Soil Physiochemical Properties
2.4. Soil DNA Extraction, PCR Amplification, and Sequencing
2.5. OTU Clustering and Annotation
2.6. Statistical Analysis
3. Results
3.1. Soil Physical and Chemical Properties of Different Vegetation Types
3.2. Sequencing Results of Soil Fungi
3.3. Composition of Soil Fungal Community
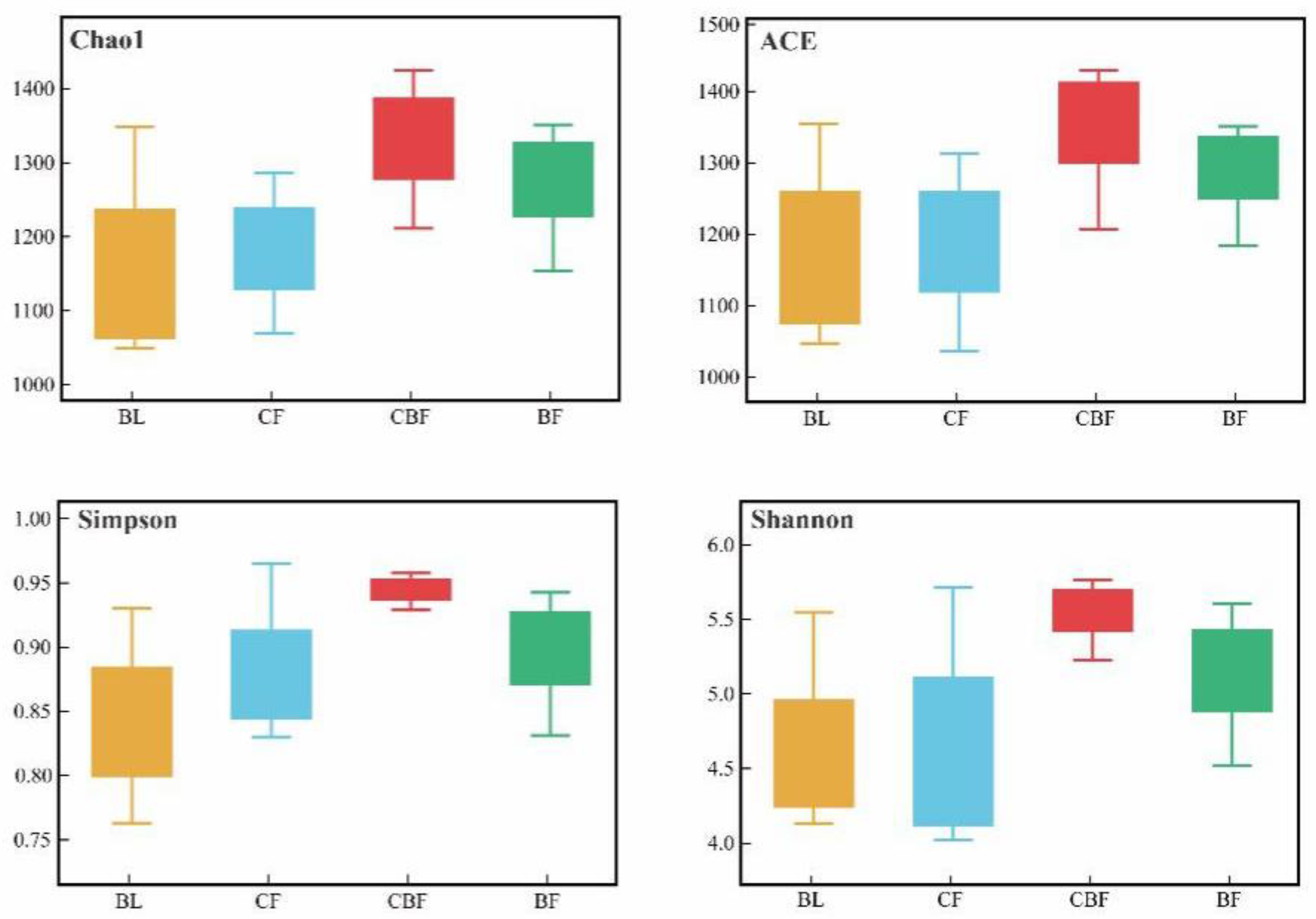
3.4. Alpha Diversity of Soil Fungi
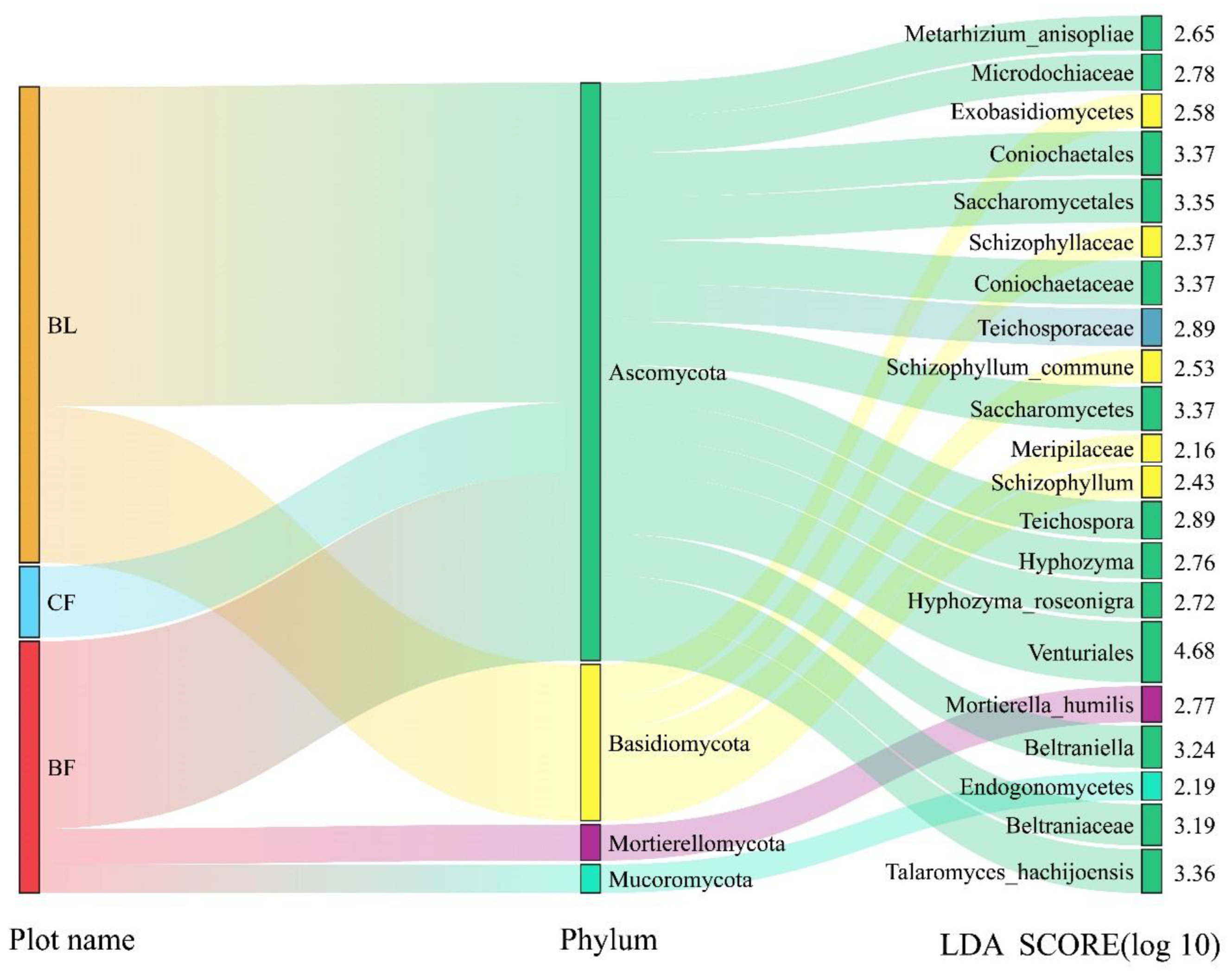
3.5. Analysis of Beta Diversity and Indicator Species of Soil Fungi
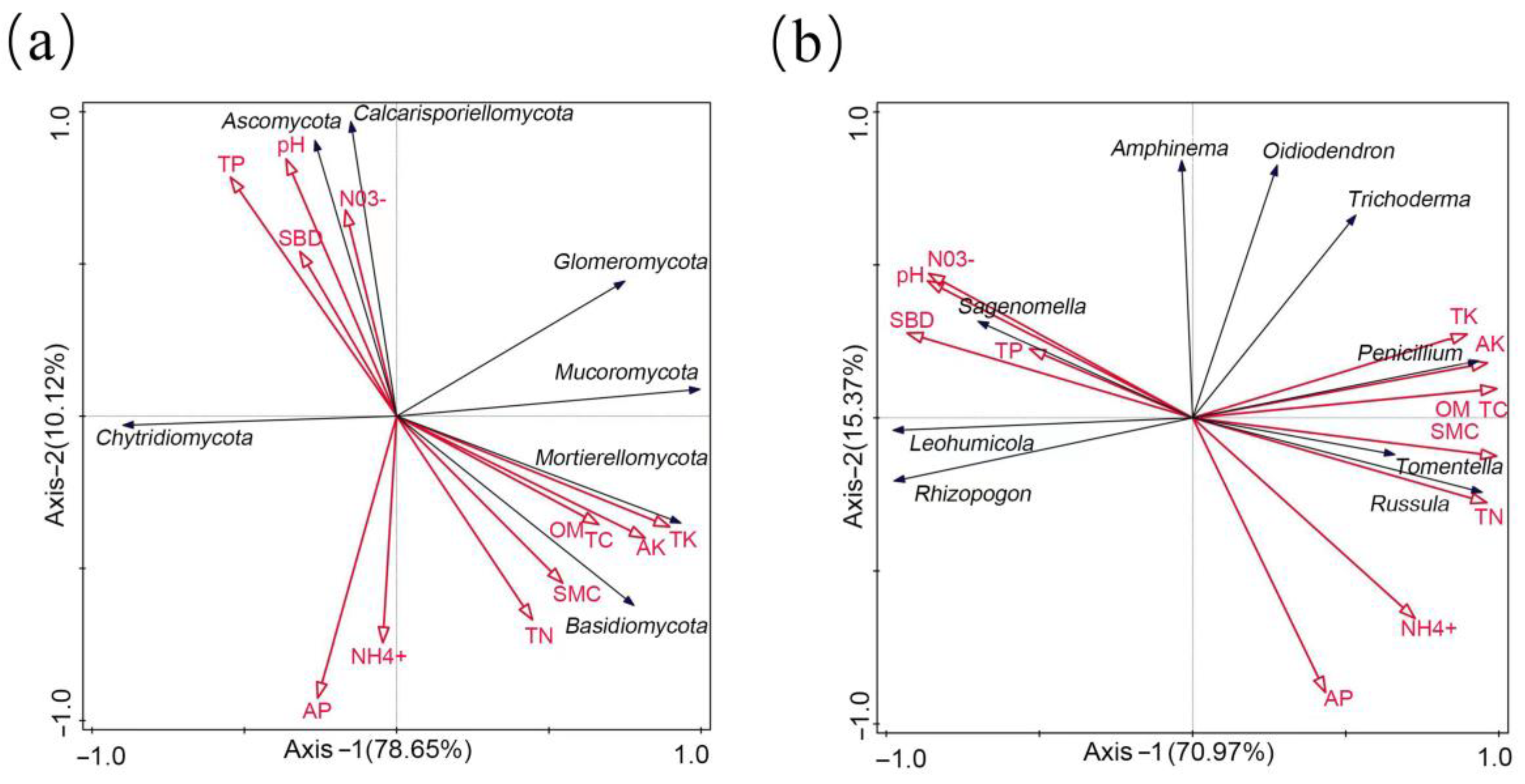
3.6. Correlation Analysis between Soil Fungi and Soil Properties
3.7. Notes on the Functions of Soil Fungal Communities
3.8. Model Analysis Driving Changes in Soil Fungal Communities
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Qin, X.; Qiu, Q.; Yu, R.G.; Yao, Y.F.; Li, H.Q.; Shao, M.G.; Wei, X.R. Soil and water conservation measures reduce erosion but result in carbon and nitrogen accumulation of red soil in Southern China. Agric. Ecosyst. Environ. 2023, 346, 108346. [Google Scholar] [CrossRef]
- Gao, J.; Shi, C.Q.; Yang, J.Y.; Yue, H.; Liu, Y.; Chen, B.Z. Analysis of spatiotemporal heterogeneity and influencing factors of soil erosion in a typical erosion zone of the southern red soil region, China. Ecol. Indic. 2023, 154, 110590. [Google Scholar] [CrossRef]
- Chen, S.; Zha, X.; Bai, Y.H.; Wang, L. Evaluation of soil erosion vulnerability on the basis of exposure, sensitivity, and adaptive capacity: A case study in the Zhuxi watershed, Changting, Fujian Province, Southern China. Catena 2019, 177, 57–69. [Google Scholar] [CrossRef]
- Hou, X.L.; Han, H.; Tigabu, M.; Cai, L.P.; Meng, F.R.; Liu, A.Q.; Ma, X.Q. Changes in soil physico-chemical properties following vegetation restoration mediate bacterial community composition and diversity in Changting, China. Ecol. Eng. 2019, 38, 171–179. [Google Scholar] [CrossRef]
- Feng, T.J.; Wei, T.X.; Saskia, D.K.; Zhang, J.J.; Bi, H.X.; Wang, R.S.; Ping, W. Long-term effects of vegetation restoration on hydrological regulation functions and the implications to afforestation on the Loess Plateau. Agric. For. Meteorol. 2023, 330, 109313. [Google Scholar] [CrossRef]
- Schwartz, N.B.; Medvigy, D.; Tijerin, J.; Pérez-Aviles, D.; Rivera-Polanco, D.; Pereira, D.; German, V.G.; Leland, W.; Du, D.; Arnold, L.; et al. Intra-annual variation in microclimatic conditions in relation to vegetation type and structure in two tropical dry forests undergoing secondary succession. Forest Ecol. Manag. 2022, 511, 120132. [Google Scholar] [CrossRef]
- Wen, H.; Ni, S.M.; Wang, J.G.; Cai, C.F. Changes of soil quality induced by different vegetation restoration in the collapsing gully erosion areas of southern China. Int. Soil Water Cons. 2021, 9, 195–206. [Google Scholar] [CrossRef]
- Xia, J.B.; Ren, R.R.; Chen, Y.P.; Sun, J.; Zhao, X.M.; Zhang, S.Y. Multifractal characteristics of soil particle distribution under different vegetation types in the Yellow River Delta chenier of China. Geoderma 2020, 368, 114311. [Google Scholar] [CrossRef]
- Guan, H.L.; Fan, J.W.; Lu, X.L. Soil specific enzyme stoichiometry reflects nitrogen limitation of microorganisms under different types of vegetation restoration in the karst areas. Appl. Soil Ecol. 2022, 169, 104253. [Google Scholar] [CrossRef]
- Yan, J.K.; Lou, L.; Bai, W.H.; Zhang, S.Q.; Zhang, N.N. Phosphorus deficiency is the main limiting factor for re-vegetation and soil microorganisms in Mu Us Sandy Land, Northwest China. Sci. Total Environ. 2023, 900, 165770. [Google Scholar] [CrossRef]
- Buee, M.; Reich, M.; Murat, C.; Morin, E.; Nilsson, R.H.; Uroz, S.; Martin, F. Pyrosequencing analyses of forest soils reveal an unexpectedly high fungal diversity. New Phytol. 2009, 184, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ostermann, A.; Karunarathna, S.C.; Xu, J.C.; Hyde, K.D.; Mortimer, P.E. The importance of plot size and the number of sampling seasons on capturing macrofungal species richness. Fungal Biol. 2018, 122, 692–700. [Google Scholar] [CrossRef] [PubMed]
- Blagodatskaya, E.; Kuzyakov, Y. Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure: Critical review. Biol. Fertil. Soils 2008, 45, 115–131. [Google Scholar] [CrossRef]
- White, C.; Tardif, J.C.; Adkins, A.; Staniforth, R. Functional diversity of microbial communities in the mixed boreal plain forest of central Canada. Soil Biol. Biochem. 2005, 37, 1359–1372. [Google Scholar] [CrossRef]
- Saetre, P.; Baê, E. Spatial variation and patterns of soil microbial community structure in a mixed spruce-birch stand. Soil Biol. Biochem. 2000, 32, 909–917. [Google Scholar] [CrossRef]
- Nagati, M.; Roy, M.; Manzi, S.; Richard, F.; Desrochers, A.; Gardes, M.; Bergeron, Y. Impact of local forest composition on soil fungal communities in a mixed boreal forest. Plant Soil 2018, 432, 345–357. [Google Scholar] [CrossRef]
- Liu, W.W.; Yu, L.F.; Yan, L.B.; Liu, N.; Zhao, Q. Composition of soil fungi communities in different stages of vegetation restoration in Karst rocky desertification area. Ecol. Environ. Sci. 2019, 28, 669–675. [Google Scholar] [CrossRef]
- Wang, J.C.; Rhodes, G.; Huang, Q.W.; Shen, Q.R. Plant growth stages and fertilization regimes drive soil fungal community compositions in a wheat-rice rotation system. Biol. Fertil. Soils 2018, 54, 731–742. [Google Scholar] [CrossRef]
- Marín, C.; Godoy, R.; Valenzuela, E.; Schloter, M.; Wubet, T.; Boy, J.; Gschwendtner, S. Functional land-use change effects on soil fungal communities in Chilean temperate rainforests. J. Soil Sci. Plant Nutr. 2017, 17, 985–1002. [Google Scholar] [CrossRef]
- Liu, L.L.; Zhou, G.Y.; Dang, P.; Chen, J.; Shang, H.; Qiu, L.Q.; Zhu, N.H. Differences of soil fungal community structure under three afforestation modes in rocky desertification region of Western Hunan Province. Acta Ecol. Sin. 2022, 42, 4150–4159. [Google Scholar] [CrossRef]
- Li, P.F.; Zhang, X.C.; Hao, M.D.; Cui, Y.X.; Zhang, Y.J.; Zhu, S.L. Effects of vegetation restoration on soil physicochemical properties, enzyme activities, and fungal community of reconstructed soil in a mining area on Loess Plateau. Bull. Soil Water Conserv. 2019, 39, 1–7. [Google Scholar] [CrossRef]
- Zeng, X.H.; Zhang, W.J.; Cao, J.S.; Liu, X.P.; Shen, H.T.; Zhao, X. Changes in soil organic carbon, nitrogen, phosphorus, and bulk density after afforestation of the Beijing-Tianjin Sandstorm Source Control program in China. Catena 2014, 118, 186–194. [Google Scholar] [CrossRef]
- Lu, R.K. Analytical Methods of Soil and Agricultural Chemistry; China Agricultural Science and Technology Press: Beijing, China, 1999; pp. 107–150. [Google Scholar]
- He, X.X.; Sheng, M.Y.; Wang, L.J.; Zhang, S.L.; Luo, N. Effects on soil organic carbon accumulation and mineralization of long-term vegetation restoration in Southwest China karst. Ecol. Indic. 2022, 145, 109622. [Google Scholar] [CrossRef]
- Zhang, H.F.; Wang, L.L.; Liu, H.M.; Zhao, J.N.; Li, G.; Wang, H.; Lai, X.; Li, J.; Xiu, W.M.; Yang, D.L. Nitrogen deposition combined with elevated precipitation is conducive to maintaining the stability of the soil fungal diversity on the Stipa baicalensis steppe. Soil Biol. Biochem. 2018, 117, 135–138. [Google Scholar] [CrossRef]
- Sun, Y.J.; He, C.X.; Xu, G.P.; Mo, L.; Gu, D.X.; Yao, Y.F.; Huang, Y.Q. Soil microbial properties under different vegetation types in Shiwan Mountain, Guangxi Province. Chin. J. Ecol. 2015, 34, 352–359. [Google Scholar] [CrossRef]
- Qiao, S.S.; Zhou, Y.N.; Chai, B.F.; Jia, T.; Li, C. Characteristics of fungi community structure and genetic diversity of forests in Guandi Mountains. Environ. Sci. 2017, 38, 2502–2512. [Google Scholar] [CrossRef]
- Tang, J.; Mo, Y.H.; Zhang, J.Y.; Zhang, R.D. Influence of biological aggregating agents associated with microbial population on soil aggregate stability. Appl. Soil Ecol. 2011, 47, 153–159. [Google Scholar] [CrossRef]
- Větrovský, T.; Kolaříková, Z.; Lepinay, C.; Hollá, S.A.; Davison, J.; Fleyberková, A.; Gromyko, A.; Jelínková, B.; Kolařík, M.; Krüger, M.; et al. GlobalAMFungi: A global database of arbuscular mycorrhizal fungal occurrences from high-throughput sequencing metabarcoding studies. New Phytol. 2023, 240, 2151–2163. [Google Scholar] [CrossRef]
- Macías-Rodríguez, L.; Contreras-Cornejo, H.A.; Adame-Garnica, S.G.; Del-Val, E.; Larsen, J. The interactions of Trichoderma at multiple trophic levels: Inter-kingdom communication. Microbiol. Res. 2020, 240, 126552. [Google Scholar] [CrossRef]
- Wu, D.; Zhang, M.M.; Peng, M.; Sui, X.; Li, W.; Sun, G.Y. Variations in soil functional fungal community structure associated with pure and mixed plantations in typical temperate forests of China. Front. Microbiol. 2019, 10, 1636. [Google Scholar] [CrossRef]
- She, W.W.; Bai, Y.X.; Zhang, Y.Q.; Qin, S.G.; Feng, W.; Sun, Y.F.; Zheng, J.; Wu, B. Resource availability drives responses of soil microbial communities to short-term precipitation and nitrogen addition in a desert shrubland. Front. Microbiol. 2018, 9, 186. [Google Scholar] [CrossRef] [PubMed]
- Li, D.H.; Li, X.W.; Su, Y.; Li, X.Z.; Yin, H.F.; Li, X.J.; Guo, M.J.; He, Y.X. Forest gaps influence fungal community assembly in a weeping cypress forest. Appl. Microbiol. Biot. 2019, 103, 3215–3224. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.H.; Xu, X.L.; Li, Z.W.; Liu, M.X.; Xu, C.H.; Zhang, R.F.; Luo, W. Effects of vegetation restoration on soil quality in degraded karst landscapes of southwest China. Sci. Total Environ. 2019, 650, 2657–2665. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.Q.; Chen, C.R.; Xu, Z.Z.; Liu, Y.Q. Effects of single and mixed species forest ecosystems on diversity and function of soil microbial community in subtropical China. J. Soil. Sediments. 2012, 12, 228–240. [Google Scholar] [CrossRef]
- Weber, C.; Lockhart, J.; Charaska, E.; Aho, K.; Lohse, K.A. Bacterial composition of soils in ponderosa pine and mixed conifer forests exposed to different wildfire burn severity. Soil Biol. Biochem. 2014, 69, 242–250. [Google Scholar] [CrossRef]
- Stephanie, N.; Hawkes, C.V. Tree species, spatial heterogeneity, and seasonality drive soil fungal abundance, richness, and composition in Neotropical rainforests. Environ. Microbiol. 2016, 18, 4662–4673. [Google Scholar] [CrossRef]
- Courty, P.E.; Buee, M.; Diedhiou, A.G.; Frey-Klett, P.; Le Tacon, F.; Rineau, F.; Turpault, M.P.; Uroz, S.; Garbaye, J. The role of ectomycorrhizal communities in forest ecosystem processes: New perspectives and emerging concepts. Soil Biol. Biochem. 2010, 42, 679–698. [Google Scholar] [CrossRef]
- Ceci, A.; Pinzari, F.; Russo, F.; Persiani, A.M.; Gadd, G.M. Roles of saprotrophic fungi in biodegradation or transformation of organic and inorganic pollutants in co-contaminated sites. Appl. Microbiol. Biotechnol. 2019, 103, 53–68. [Google Scholar] [CrossRef]
- Detheridge, A.P.; Comont, D.; Callaghan, T.M.; Bussell, J.; Brand, G.; Gwynn-Jones, D.; Scullion, J.; Griffith, G.W. Vegetation and edaphic factors influence rapid establishment of distinct fungal communities on former coal-spoil sites. Fungal Ecol. 2018, 33, 92–103. [Google Scholar] [CrossRef]
- Becquer, A.; Garcia, K.; Amenc, L.; Rivard, C.; Dore, J.; Trives-Segura, C.; Szponarski, W.; Russet, S.; Bazeza, Y.; Lassalle-Kaiser, B.; et al. The Hebeloma cylindrosporum HcPT2 Pi transporter plays a key role in ectomycorrhizal symbiosis. New Phytol. 2018, 220, 1185–1199. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Song, Z.W.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cover * | BL | CF | CBF | BF |
|---|---|---|---|---|
| pH | 4.78 ± 0.37 a | 4.84 ± 0.14 a | 4.70 ± 0.12 a | 4.66 ± 0.09 a |
| TC g·kg−1 | 1.83 ± 0.22 d | 5.06 ± 0.50 c | 6.30 ± 0.48 b | 9.66 ± 1.80 a |
| TN g·kg−1 | 0.22 ± 0.03 c | 0.26 ± 0.01 c | 0.32 ± 0.03 b | 0.58 ± 0.09 a |
| TP g·kg−1 | 0.16 ± 0.01 ab | 0.17 ± 0.02 a | 0.15 ± 0.01 b | 0.15 ± 0.01 b |
| TK g·kg−1 | 3.32 ± 0.17 b | 4.89 ± 0.50 b | 9.95 ± 1.90 a | 10.41 ± 2.43 a |
| SOM g·kg−1 | 3.16 ± 0.20 d | 8.72 ± 0.58 c | 10.87 ± 1.10 b | 16.65 ± 2.07 a |
| NH4+ mg·kg−1 | 13.22 ± 1.74 c | 15.67 ± 1.25 b | 13.19 ± 0.83 c | 22.55 ± 1.89 a |
| NO3− mg·kg−1 | 3.59 ± 0.24 b | 4.65 ± 0.09 a | 4.29 ± 0.18 a | 3.67 ± 0.82 b |
| AK mg·kg−1 | 10.48 ± 0.59 d | 12.67 ± 0.94 c | 16.81 ± 1.46 b | 18.41 ± 1.05 a |
| AP mg·kg−1 | 6.21 ± 0.37 b | 6.85 ± 0.05 ab | 6.83 ± 0.05 ab | 7.45 ± 1.22 a |
| SBD g·cm−3 | 1.52 ± 0.10 a | 1.43 ± 0.17 ab | 1.43 ± 0.08 ab | 1.32 ± 0.08 b |
| SMC % | 15.30 ± 1.31 c | 16.91 ± 0.34 bc | 18.01 ± 1.02 b | 21.92 ± 1.47 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, X.; Yu, J.; Han, X.; Zhai, L.; Farooq, T.H.; Li, Q.; Liu, L. Dynamic Shifts in Soil Fungal Functional Group Characteristics across Distinct Vegetation Types during Ecological Restoration in Degraded Red Soil Regions. Forests 2024, 15, 89. https://doi.org/10.3390/f15010089
Hou X, Yu J, Han X, Zhai L, Farooq TH, Li Q, Liu L. Dynamic Shifts in Soil Fungal Functional Group Characteristics across Distinct Vegetation Types during Ecological Restoration in Degraded Red Soil Regions. Forests. 2024; 15(1):89. https://doi.org/10.3390/f15010089
Chicago/Turabian StyleHou, Xiaolong, Junbao Yu, Xuejie Han, Lin Zhai, Taimoor Hassan Farooq, Qiyan Li, and Linghua Liu. 2024. "Dynamic Shifts in Soil Fungal Functional Group Characteristics across Distinct Vegetation Types during Ecological Restoration in Degraded Red Soil Regions" Forests 15, no. 1: 89. https://doi.org/10.3390/f15010089
APA StyleHou, X., Yu, J., Han, X., Zhai, L., Farooq, T. H., Li, Q., & Liu, L. (2024). Dynamic Shifts in Soil Fungal Functional Group Characteristics across Distinct Vegetation Types during Ecological Restoration in Degraded Red Soil Regions. Forests, 15(1), 89. https://doi.org/10.3390/f15010089