
Effects of Flooding and Shade on Survival, Growth, and Leaf Gas Exchange of Bottomland Tree Species across the Great Lakes Region (USA)
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Data Analysis
3. Results
3.1. Survival
3.2. Patterns of Response
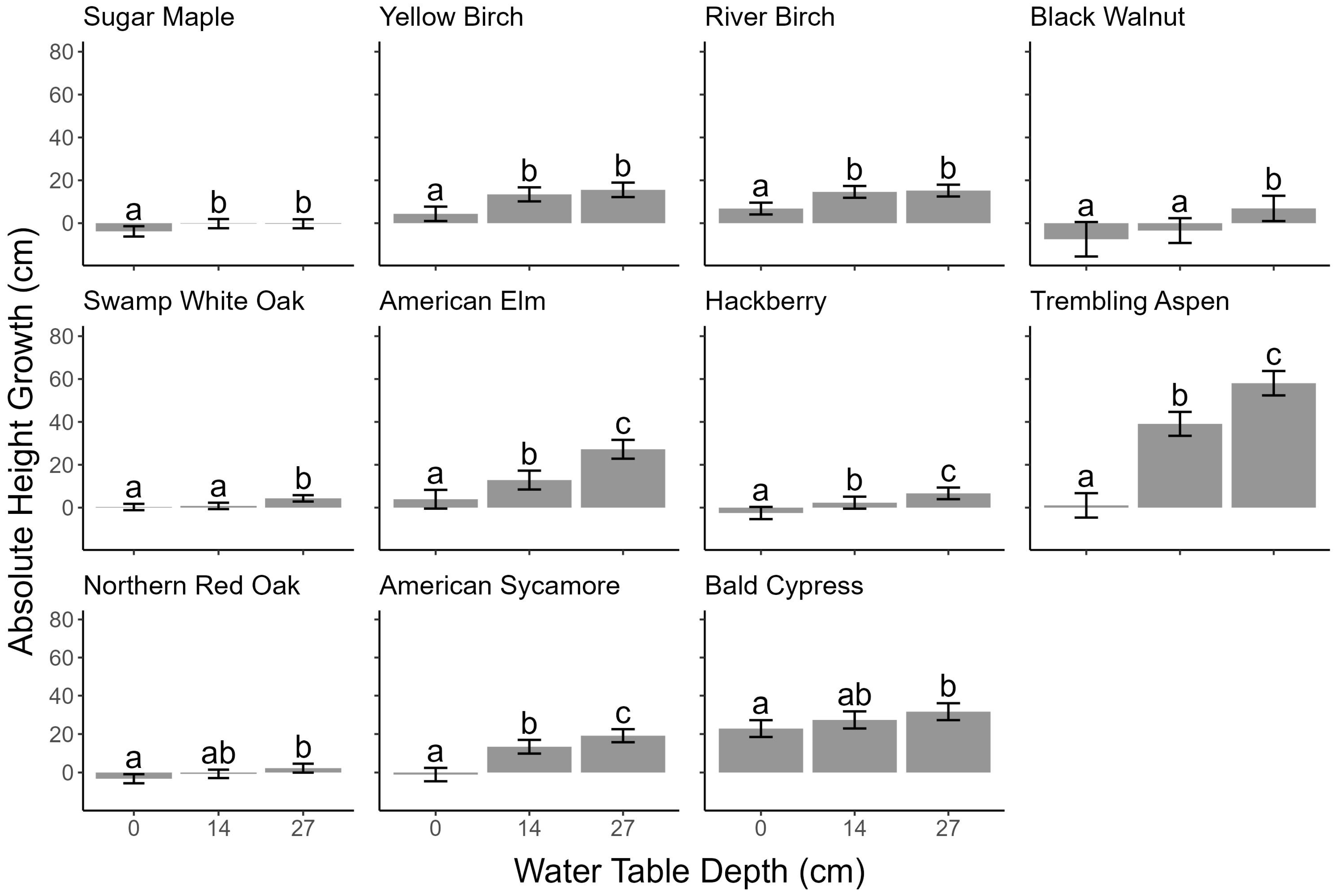
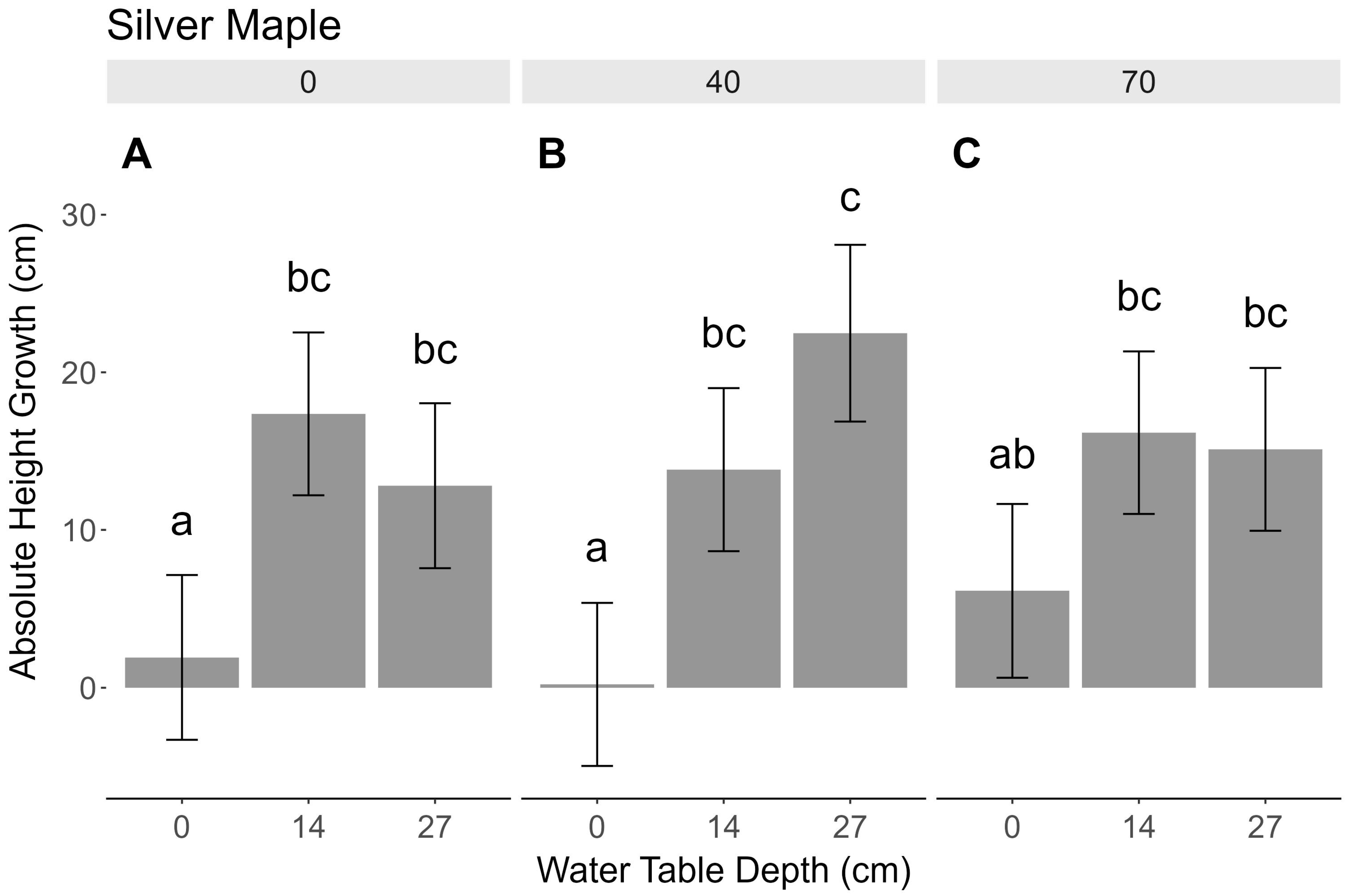
3.3. Height Growth
3.4. Basal Diameter Growth
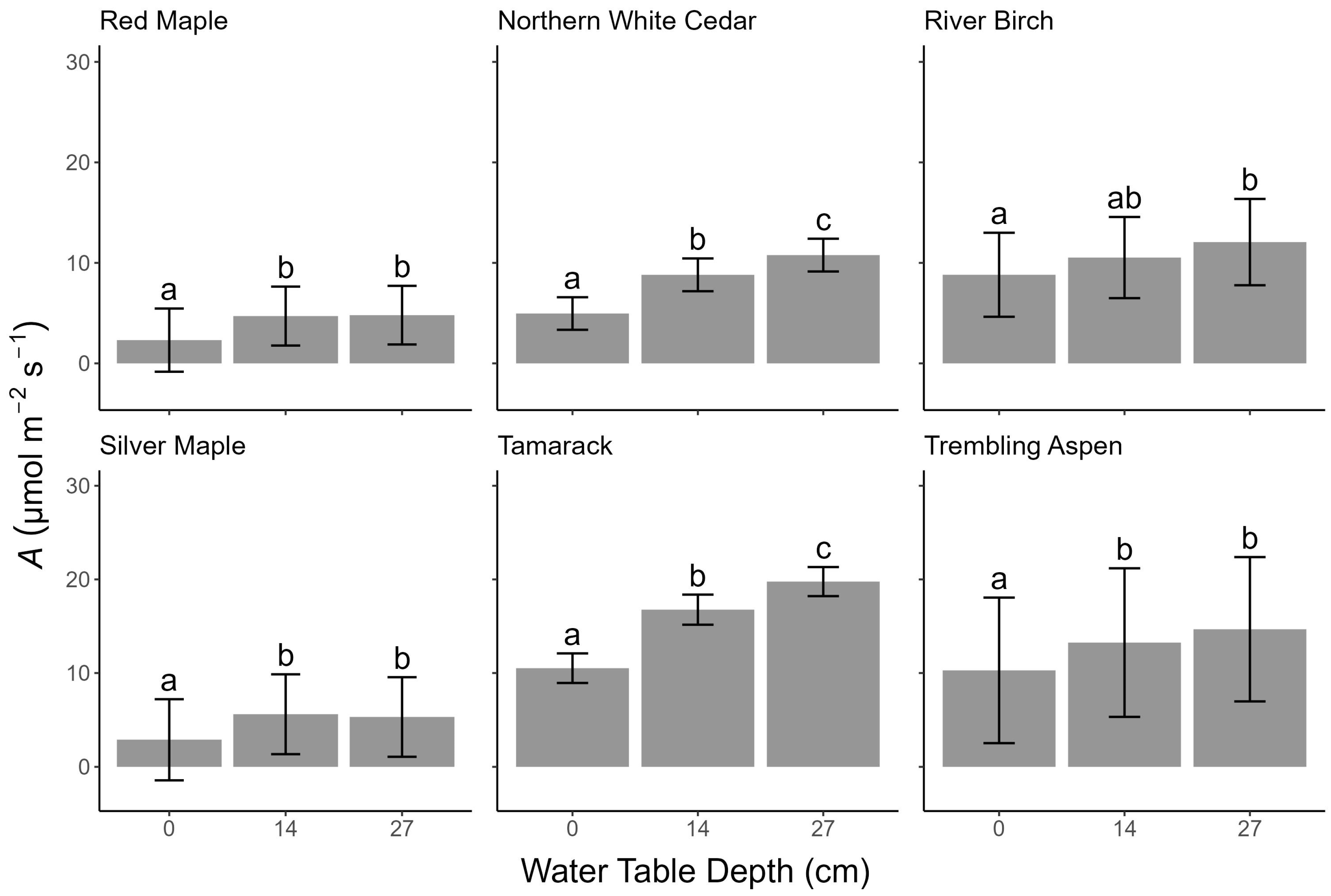
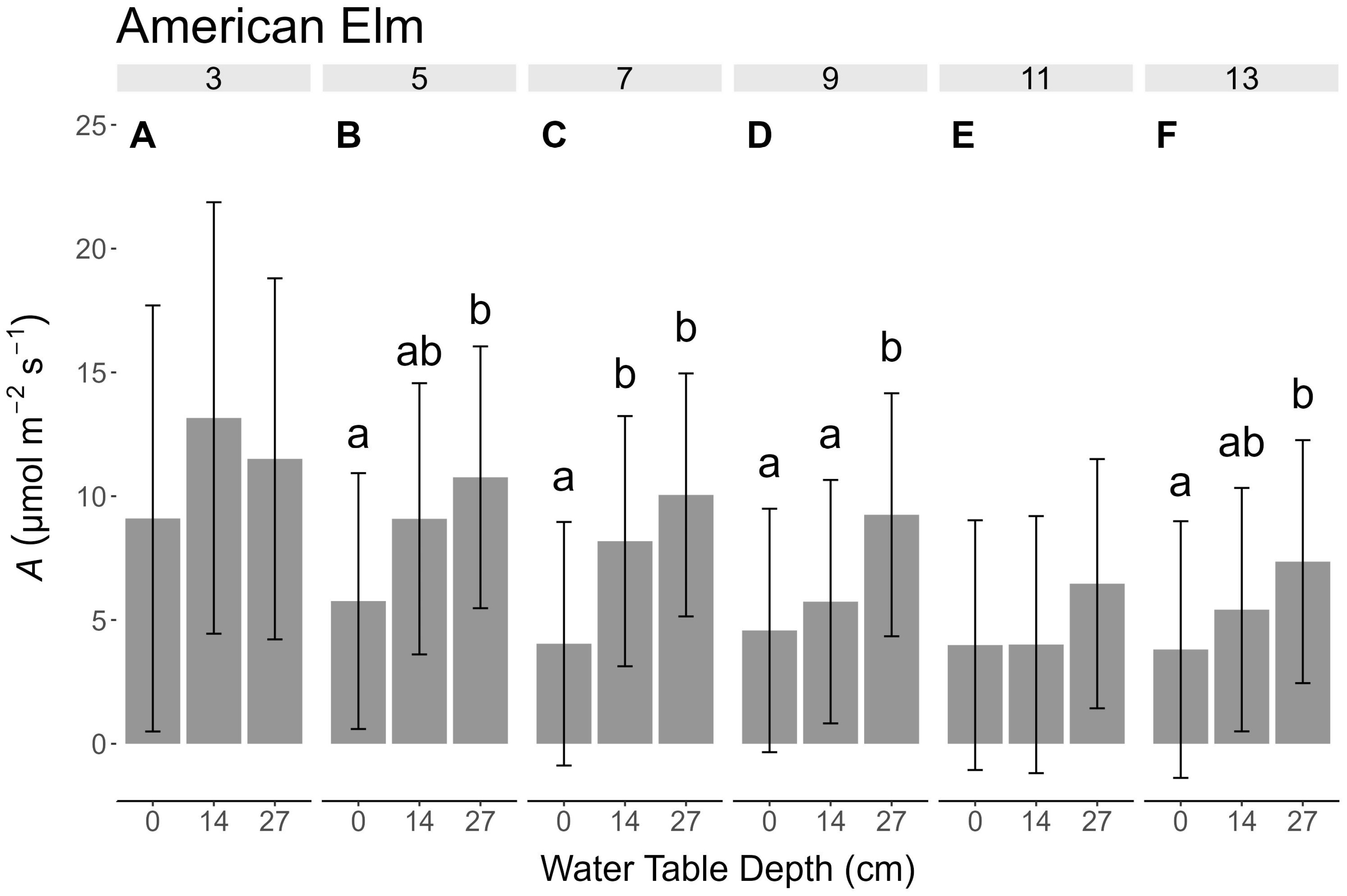
3.5. Photosynthesis
3.6. Stomatal Conductance
3.7. Transpiration
4. Discussion
4.1. Water Table Depth
4.2. Transpiration
4.3. Light Reduction
4.4. Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Knapp, A.K.; Beier, C.; Briske, D.D.; Classen, A.T.; Luo, Y.; Reichstein, M.; Smith, M.D.; Smith, S.D.; Bell, J.E.; Fay, P.A.; et al. Consequences of more extreme precipitation regimes for terrestrial ecosystems. Bioscience 2008, 58, 811–821. [Google Scholar] [CrossRef]
- Niu, S.; Luo, Y.; Li, D.; Cao, S.; Xia, J.; Li, J.; Smith, M.D. Plant growth and mortality under climatic extremes: An overview. Environ. Exp. Bot. 2014, 98, 13–19. [Google Scholar] [CrossRef]
- Tebaldi, C.; Hayhoe, K.; Arblaster, J.M.; Meehl, G.A. Going to the extremes: An intercomparison of model-simulated historical and future changes in extreme events. Clim. Chang. 2006, 79, 185–211. [Google Scholar] [CrossRef]
- Easterling, D.R.; Meehl, G.A.; Parmesan, C.; Changnon, S.A.; Karl, T.R.; Mearns, L.O. Climate extremes: Observations, modeling, and impacts. Science 2000, 289, 2068–2074. [Google Scholar] [CrossRef]
- Gutschick, V.P.; BassiriRad, H. Extreme events as shaping physiology, ecology, and evolution of plants: Toward a unified definition and evaluation of their consequences. New Phytol. 2003, 160, 21–42. [Google Scholar] [CrossRef]
- Stanturf, J.A.; Gardiner, E.S.; Shepard, J.P.; Schweitzer, C.J.; Portwood, C.J.; Dorri, L.C., Jr. Restoration of bottomland hardwood forests across a treatment intensity gradient. For. Ecol. Manag. 2009, 257, 1803–1814. [Google Scholar] [CrossRef]
- Campbell, D.; Keddy, P.A.; Broussard, M.; McFalls-Smith, T.B. Small changes in flooding have large consequences: Experimental data from ten wetland plants. Wetlands 2016, 36, 457–466. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Hernandez, M.E. Landscape and climate change threats to wetlands of North and Central America. Aquat. Sci. 2013, 75, 133–149. [Google Scholar] [CrossRef]
- Kingsford, R.T.; Basset, A.; Jackson, L. Wetlands: Conservation’s poor cousins. Aquat. Conserv. Mar. Freshw. Ecosyst. 2016, 26, 892–916. [Google Scholar] [CrossRef]
- Casanova, M.T.; Brock, M.A. How do depth, duration and frequency of flooding influence the establishment of wetland plant communities? Plant Ecol. 2000, 147, 237–250. [Google Scholar] [CrossRef]
- Vervuren, P.J.A.; Blom CW, P.M.; De Kroon, H. Extreme flooding events on the Rhine and the survival and distribution of riparian plant species. J. Ecol. 2003, 91, 135–146. [Google Scholar] [CrossRef]
- Harms, W.R. Some effects of soil type and water regime on growth of tupelo seedlings. Ecology 1973, 54, 188–193. [Google Scholar] [CrossRef]
- Landhäusser, S.M.; Silins, U.; Lieffers, V.J.; Liu, W. Response of Populus tremuloides, Populus balsamifera, Betula papyrifera and Picea glauca seedlings to low soil temperature and water-logged soil conditions. Scand. J. For. Res. 2003, 18, 391–400. [Google Scholar] [CrossRef]
- Islam, M.A.; Macdonald, S.E. Ecophysiological adaptations of black spruce (Picea mariana) and tamarack (Larix laricina) seedlings to flooding. Trees 2004, 18, 35–42. [Google Scholar] [CrossRef]
- Kozlowski, T.T. Responses of woody plants to flooding and salinity. Tree Physiol. 1997, 17, 490. [Google Scholar] [CrossRef]
- Walsh, M.P.; Van Sambeek, J.W.; Coggeshall, M.V. Variation in flood tolerance of container-grown seedlings of swamp white oak, bur oak, and white oak. In Proceedings of the 16th Central Hardwood Forest Conference, West Lafayette, IN, USA, 8–9 April 2008; General Technical Report, NRS-P-24; Jacobs, D.F., Michler, C.H., Eds.; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2008; pp. 446–456. [Google Scholar]
- Kreuzwieser, J.; Rennenberg, H. Molecular and physiological responses of trees to waterlogging stress. Plant Cell Environ. 2014, 37, 2245–2259. [Google Scholar] [CrossRef] [PubMed]
- Kimmins, J.P. Forest ecology. In Fishes and Forestry: Worldwide Watershed Interactions and Management; Blackwell Publishing: Oxford, UK, 2004; pp. 17–43. [Google Scholar]
- Loach, K. Shade Tolerance in Tree Seedlings: I. Leaf Photosynthesis and Respiration in Plants Raised Under Artificial Shade. New Phytol. 1967, 66, 607–621. [Google Scholar] [CrossRef]
- Mielke, M.S.; Schaffer, B. Leaf gas exchange, chlorophyll fluorescence and pigment indexes of Eugenia uniflora L. in response to changes in light intensity and soil flooding. Tree Physiol. 2010, 30, 45–55. [Google Scholar] [CrossRef]
- Valladares, F.; Niinemets, Ü. Shade tolerance, a key plant feature of complex nature and consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef]
- Lavinsky, A.O.; De Souza Sant’Ana, C.; Mielke, M.S.; De Almeida, A.A.F.; Gomes, F.P.; França, S.; Da Costa Silva, D. Effects of light availability and soil flooding on growth and photosynthetic characteristics of Genipa americana L. seedlings. New For. 2007, 34, 41–50. [Google Scholar] [CrossRef]
- Hopkins, W.; Hüner, N. Introduction to Plant Physiology, 4th ed.; Wiley: Hoboken, NJ, USA, 2009. [Google Scholar]
- Mielke, M.S.; Schaffer, B. Effects of soil flooding and changes in light intensity on photosynthesis of Eugenia uniflora L. seedlings. Acta Physiol. Plant. 2011, 33, 1661–1668. [Google Scholar] [CrossRef]
- Wang, X.; Liu, T.; Li, C.; Chen, H. Effects of soil flooding on photosynthesis and growth of Zea mays L. seedlings under different light intensities. Afr. J. Biotechnol. 2012, 11, 7676–7685. [Google Scholar]
- Wagner, P.A.; Dreyer, E. Interactive effects of waterlogging and irradiance on the photosynthetic performance of seedlings from three oak species displaying different sensitivities (Quercus robur, Q. petraea and Q. rubra). Ann. For. Sci. 1997, 54, 409–429. [Google Scholar] [CrossRef]
- Porporato, A.; Daly, E.; Rodriguez-Iturbe, I. Soil water balance and ecosystem response to climate change. Am. Nat. 2004, 164, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Kreyling, J.; Wenigmann, M.; Beierkuhnlein, C.; Jentsch, A. Effects of extreme weather events on plant productivity and tissue die-back are modified by community composition. Ecosystems 2008, 11, 752–763. [Google Scholar] [CrossRef]
- Byun, K.; Chiu, C.M.; Hamlet, A.F. Effects of 21st century climate change on seasonal flow regimes and hydrologic extremes over the Midwest and Great Lakes region of the US. Sci. Total Environ. 2019, 650, 1261–1277. [Google Scholar] [CrossRef] [PubMed]
- Granger, J.J.; Zobel, J.M.; Buckley, D.S. Differential Impacts of Emerald Ash Borer (Agrilus planipennis Fairmaire) on Forest Communities Containing Native Ash (Fraxinus spp.) Species in Eastern North America. For. Sci. 2020, 66, 38–48. [Google Scholar] [CrossRef]
- Stanturf, J.A.; Schoenholtz, S.H.; Schweitzer, C.J.; Shepard, J.P. Achieving restoration success: Myths in bottomland hardwood forests. Restor. Ecol. 2001, 9, 189–200. [Google Scholar] [CrossRef]
- Stanturf, J.A.; Conner, W.H.; Gardiner, E.S.; Schweitzer, C.J.; Ezell, A.W. Recognizing and overcoming difficult site conditions for afforestation of bottomland hardwoods. Ecol. Restor. 2004, 22, 183–193. [Google Scholar] [CrossRef]
- Pedlar, J.H.; McKenney, D.W.; Aubin, I.; Beardmore, T.; Beaulieu, J.; Iverson, L.; O’Neill, G.A.; Winder, R.S.; Ste-Marie, C. Placing forestry in the assisted migration debate. BioScience 2012, 62, 835–842. [Google Scholar] [CrossRef]
- Keller, G.J.; Slesak, R.A.; Bronson, D.; Windmuller-Campione, M.A.; McGraw, A. Effects of flood duration on seedling survival and growth of potential replacement species in black ash (Fraxinus nigra Marshall) forests threatened by emerald ash borer. Trees For. People 2023, 11, 100367. [Google Scholar] [CrossRef]
- Bermudez, R.; Stefanski, A.; Montgomery, R.A.; Reich, P.B. Short-and long-term responses of photosynthetic capacity to temperature in four boreal tree species in a free-air warming and rainfall manipulation experiment. Tree Physiol. 2021, 41, 89–102. [Google Scholar] [CrossRef]
- Kramer, P.J.; Kozlowski, T.T. Physiology of Woody Plants; Academic Press: Orlando, FL, USA, 1979. [Google Scholar]
- Hupperts, S.F.; Webster, C.R.; Froese, R.E.; Dickinson, Y.L. Seedling and sapling recruitment following novel silvicultural treatments in Great Lakes northern hardwoods. For. Ecol. Manag. 2020, 462, 117983. [Google Scholar] [CrossRef]
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. Emmeans: Estimated Marginal Means, aka Least-Squares Means; R Package Version 1; R Foundation for Statistical Computing: Vienna, Austira, 2019. [Google Scholar]
- Looney, C.E.; D’Amato, A.W.; Palik, B.J.; Slesak, R.A. Canopy treatment influences growth of replacement tree species in Fraxinus nigra forests threatened by the emerald ash borer in Minnesota, USA. Can. J. For. Res. 2017, 47, 183–192. [Google Scholar] [CrossRef]
- Luo, D.; Ganesh, S.; Koolaard, J.; Luo, M.D. Package ‘Predictmeans’. 2018. Available online: https://CRAN.R-project.org/package=predictmeans (accessed on 15 December 2021).
- Bronson, D.R.; Gower, S.T.; Tanner Mm Linder, S.; Van Herk, I. Response of soil surface CO2 flux in a boreal forest to ecosystem warming. Glob. Chang. Biol. 2008, 14, 856–867. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression; Sage Publications: Thousand Oaks, CA, USA, 2018. [Google Scholar]
- Middleton, B.A.; Souter, N.J. Functional integrity of freshwater forested wetlands, hydrologic alteration, and climate change. Ecosyst. Health Sustain. 2016, 2, e01200. [Google Scholar] [CrossRef]
- Kabrick, J.M.; Dey, D.C.; Van Sambeek, J.W.; Coggeshall, M.V.; Jacobs, D.F. Quantifying flooding effects on hardwood seedling survival and growth for bottomland restoration. New For. 2012, 43, 695–710. [Google Scholar] [CrossRef]
- Frye, J.; Grosse, W. Growth Responses to Flooding and Recovery of Deciduous Trees. Z. Naturforschung C 1992, 47, 683–689. [Google Scholar] [CrossRef]
- Burns, R.; Honkala, B. Silvics of North America; United States Forest Service: Washington, DC, USA, 1990. [Google Scholar]
- Iverson, L.R.; Prasad, A.M.; Matthews, S.N.; Peters, M. Estimating potential habitat for 134 eastern US tree species under six climate scenarios. For. Ecol. Manag. 2008, 254, 390–406. [Google Scholar] [CrossRef]
- Hagerman, S.; Kozak, R. isentangling the social complexities of assisted migration through deliberative methods. J. Ecol. 2021, 109, 2309–2316. [Google Scholar] [CrossRef]
- Tang, Z.C.; Kozlowski, T.T. Ethylene production and morphological adaptation of woody plants to flooding. Can. J. Bot. 1984, 62, 1659–1664. [Google Scholar] [CrossRef]
- Barnes, B.V.; Wagner, W.H. Michigan Trees, Revised and Updated: A Guide to the Trees of the Great Lakes Region; University of Michigan Press: Ann Arbor, MI, USA, 2004. [Google Scholar]
- Peterson, D.; Bazzaz, F. Photosynthetic and growth responses of silver maple (Acer saccharinum L.) seedlings to flooding. Am. Midl. Nat. 1984, 112, 261–272. [Google Scholar] [CrossRef]
- Guyon, L.; Battaglia, L. Ecological characteristics of floodplain forest reference sites in the Upper Mississippi River System. For. Ecol. Manag. 2018, 427, 208–216. [Google Scholar] [CrossRef]
- Boulfroy, E.; Forget, E.; Hofmeyer, P.V.; Kenefic, L.S.; Larouche, C.; Lessard, G.; Lussier, J.; Pinto, F.; Ruel, J.; Weiskittel, A. Silvicultural Guide for Northern White-Cedar (Eastern White Cedar); Gen. Tech. Rep. NRS-98; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2012; 74p. [Google Scholar]
- Lieffers, V.; Rothwell, R. Effects of depth of water table and substrate temperature on root and top growth of Picea mariana and Larix laricina seedlings. Can. J. For. Res. 1986, 16, 1201–1206. [Google Scholar] [CrossRef]
- Gilbert, A.M. Silvical Characteristics of Yellow Birch (Betula alleghaniensis); Station Paper NE-134; U.S. Department of Agriculture, Forest Service, Northeastern Forest Experiment Station: Upper Darby, PA, USA, 1960; 18p. [Google Scholar]
- Heinselman, M.L. Silvical Characteristics of Black Spruce (Picea mariana) (No. 45); Lake States Forest Experiment Station, Forest Service, US Department of Agriculture: Washington, DC, USA, 1957. [Google Scholar]
- Iverson, L.; Knight, K.S.; Prasad, A.; Herms, D.A.; Matthews, S.; Peters, M.; Smith, A.; Hartzler, D.M.; Long, R.; Almendinger, J. Potential Species Replacements for Black Ash (Fraxinus nigra) at the Confluence of Two Threats: Emerald Ash Borer and a Changing Climate. Ecosystems 2016, 19, 248–270. [Google Scholar] [CrossRef]
- Coggeshall, M.V.; Van Sambeek, J.W.; Schlarbaum, S.E. Genotypic variation in flood tolerance of black walnut and three southern bottomland oaks. In Proceedings of the 15th Central Hardwood Forest Conference, Knoxville, TN, USA, 27 February 27–1 March 2006; Buckley, D.S., Clatterbuck, W.K., Eds.; e-Gen. Tech. Rep. SRS-101; U.S. Department of Agriculture, Forest Service, Southern Research Station: Washington, DC, USA, 2007; pp. 629–637. [Google Scholar]
- Shifley, P.; Rogers, S.; Dey, R.; Kabrick, D.; Johnson, J.; Rogers, R. The Ecology and Silviculture of Oaks, 3rd ed.; CABI: Wallingford, UK; Boston, MA, USA, 2019. [Google Scholar]
- Hosner, J.F.; Leaf, A.L. The effect of soil saturation upon the dry weight, ash content, and nutrient absorption of various bottomland tree seedlings. Soil Sci. Soc. Am. J. 1962, 26, 401–404. [Google Scholar] [CrossRef]
- Flower, C.E.; Lynch, D.J.; Knight, K.S.; Gonzalez-Meler, M.A. Biotic and abiotic drivers of sap flux in mature green ash trees (Fraxinus pennsylvanica) experiencing varying levels of emerald ash borer (Agrilus planipennis) infestation. Forests 2018, 9, 301. [Google Scholar] [CrossRef]
- Parker, J. The effects of flooding on the transpiration and survival of some southeastern forest tree species. Plant Physiol. 1950, 25, 453. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Polanco, M.; Señorans, J.; Zwiazek, J.J. Role of adventitious roots in water relations of tamarack (Larix laricina) seedlings exposed to flooding. BMC Plant Biol. 2012, 12, 1–9. [Google Scholar] [CrossRef]
- Foote, K.C.; Schaedle, M. Physiological characteristics of photosynthesis and respiration in stems of Populus tremuloides Michx. Plant Physiol. 1976, 58, 91–94. [Google Scholar] [CrossRef]
- Gardiner, E.S.; Krauss, K.W. Photosynthetic light response of flooded cherrybark oak (Quercus pagoda) seedlings grown in two light regimes. Tree Physiol. 2001, 21, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Palik, B.J.; D’Amato, A.W.; Slesak, R.A.; Kastendick, D.; Looney, C.; Kragthorpe, J. Eighth-year survival and growth of planted replacement tree species in black ash (Fraxinus nigra) wetlands threatened by emerald ash borer in Minnesota, USA. For. Ecol. Manag. 2021, 484, 118958. [Google Scholar] [CrossRef]
- Van den Driessche, R. Importance of current photosynthate to new root growth in planted conifer seedlings. Can. J. For. Res. 1987, 17, 776–782. [Google Scholar] [CrossRef]
- Burdett, A. Physiological processes in plantation establishment and the development of specifications for forest planting stock. Can. J. For. Res. 1990, 20, 415–427. [Google Scholar] [CrossRef]
- Noland, T.; Mohammed, G.; Scott, M. The dependance of root growth potential on light level, photosynthetic rate and root starch content in jack pine seedlings. New For. 1997, 13, 105–119. [Google Scholar] [CrossRef]
- Grossnickle, S. Importance of root growth in overcoming planting stress. New For. 2005, 30, 273–294. [Google Scholar] [CrossRef]
- Jones, R.H.; Sharitz, R.R. Survival and growth of woody plant seedlings in the understorey of floodplain forests in South Carolina. J. Ecol. 1998, 86, 574–587. [Google Scholar] [CrossRef]
- Diamond, J.M. Ecology: Laboratory, field and natural experiments. Nature 1983, 304, 586–587. [Google Scholar] [CrossRef]
- Carpenter, S.R. Microcosm experiments have limited relevance for community and ecosystem ecology. Ecology 1996, 77, 677–680. [Google Scholar] [CrossRef]
- Miao, G.; Noormets, A.; Domec, J.C.; Trettin, C.C.; McNulty, S.G.; Sun, G.; King, J.S. The effect of water table fluctuation on soil respiration in a lower coastal plain forested wetland in the southeastern US. J. Geophys. Res. Biogeosci. 2013, 118, 1748–1762. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Common Name | Stock Type | Wetland Indicator Status | Shade Tolerance |
|---|---|---|---|---|
| Acer rubrum L. | red maple | 2-0 | Fac. | Intermediate |
| Acer saccharinum L. | silver maple | 2-0 | Fac./Fac. Wetland | Intermediate |
| Acer saccharum Marshall | sugar maple | 2-0 | Fac. Upland/Obligate Upland | Tolerant |
| Betula nigra L. | river birch | 2-0 | Fac. Wetland | Intolerant |
| Betula alleghaniensis Britton | yellow birch | 2-0 | Fac./Fac. Upland | Intermediate |
| Carya cordiformis (Wangenh.) K. Koch | bitternut hickory | 2-1 | Fac./Fac. Upland | Intolerant |
| Celtis occidentalis L. | hackberry | 2-0 | Fac. | Tolerant |
| Juglans nigra L. | black walnut | 2-0 | Fac. Upland/Obligate Upland | Intolerant |
| Larix laricina (Du Roi) K. Koch | tamarack | 2-0 | Fac. Wetland | Intolerant |
| Picea glauca (Moench) Voss | white spruce | 2-0 | Fac./Fac. Upland | Intermediate |
| Picea mariana (Mill.) Britton, Sterns & Poggenb. | black spruce | 2-0 | Fac. Wetland | Tolerant |
| Pinus resinosa Aiton | red pine | 3-0 | Fac. Upland | Intolerant |
| Pinus strobus L. | eastern white pine | 2-0 | Fac. Upland | Intermediate |
| Platanus occidentalis L. | American sycamore | 1-0 | Fac./Fac. Wetland | Intermediate |
| Populus tremuloides Michx. | trembling aspen | 2-0 | Fac. | Intolerant |
| Quercus alba L. | white oak | 2-0 | Fac. Upland | Intermediate |
| Quercus bicolor Willd. | swamp white oak | 2-0 | Fac. Wetland | Intermediate |
| Quercus macrocarpa Michx. | bur oak | 2-0 | Fac. | Intermediate |
| Quercus rubra L. | northern red oak | 2-0 | Fac. | Intermediate |
| Taxodium distichum (L.) Rich. | bald cypress | 1-0 | Obligate Wetland | Intermediate |
| Thuja occidentalis L. | northern white cedar | 3-0 | Fac. Wetland | Intermediate |
| Tilia americana L. | basswood | 2-0 | Fac. | Tolerant |
| Ulmus americana L. | American elm | 2-0 | Fac./Fac. Wetland | Intermediate |
| Pattern | Height Growth (cm) | Basal Diameter Growth (mm) | Photosynthesis (A, μmol m−2 s−1) | Stomatal Conductance (gs, mol m−2 s−1) | Transpiration (E, mmol m−2 s−1) |
|---|---|---|---|---|---|
| 1 Inconclusive results | bitternut hickory bur oak eastern white pine red pine white oak | bitternut hickory black walnut eastern white pine northern red oak sugar maple | red maple | northern red oak | |
| 2 Greatest response value at 27 cm depth | bald cypress black walnut northern red oak swamp white oak | northern white cedar red pine | river birch | swamp white oak | northern white cedar swamp white oak |
| 3 Greatest response values at 27 and 14 cm depths | American elm American sycamore hackberry river birch sugar maple trembling aspen yellow birch | American elm black spruce hackberry silver maple tamarack trembling aspen yellow birch | northern white cedar red maple silver maple tamarack trembling aspen | bur oak hackberry tamarack trembling aspen | American elm American sycamore hackberry river birch silver maple tamarack trembling aspen |
| 4 High-response values across all depths | bald cypress river birch | American elm American sycamore bald cypress river birch | bald cypress |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keller, G.J.; Bronson, D.; Slesak, R.A.; Windmuller-Campione, M.A. Effects of Flooding and Shade on Survival, Growth, and Leaf Gas Exchange of Bottomland Tree Species across the Great Lakes Region (USA). Forests 2024, 15, 530. https://doi.org/10.3390/f15030530
Keller GJ, Bronson D, Slesak RA, Windmuller-Campione MA. Effects of Flooding and Shade on Survival, Growth, and Leaf Gas Exchange of Bottomland Tree Species across the Great Lakes Region (USA). Forests. 2024; 15(3):530. https://doi.org/10.3390/f15030530
Chicago/Turabian StyleKeller, Gwendolen J., Dustin Bronson, Robert A. Slesak, and Marcella A. Windmuller-Campione. 2024. "Effects of Flooding and Shade on Survival, Growth, and Leaf Gas Exchange of Bottomland Tree Species across the Great Lakes Region (USA)" Forests 15, no. 3: 530. https://doi.org/10.3390/f15030530