
Quantifying Forest Cover Loss as a Response to Drought and Dieback of Norway Spruce and Evaluating Sensitivity of Various Vegetation Indices Using Remote Sensing

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection
2.3. Data Processing
2.4. Forest Cover Loss Analysis
2.5. Evaluation of VI Sensitivity in Detecting and Predicting Drought Effects in Norway Spruce Forests

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Vegetation Indices | Abrev. | Formula | Reference |
|---|---|---|---|---|
| Water VIs | Moisture Stress index | MSI | [107] | |
| Normalized Difference Moisture Index | NDMI | [39] | ||
| Disease Water Stress Index | DSWI | [58] | ||
| Normalised Multi-band Drought Index | NMDI | [55] | ||
| Typical VIs | Normalized Difference Vegetation Index | NDVI | [108] | |
| Enhanced Vegetation Index | EVI | [109] | ||
| Soil-Adjusted Vegetation Index | SAVI | [110] | ||
| Transformed Vegetation Index | TVI | [46] | ||
| TC components | Tasseled Cap Greeness (Landsat 8) | TCG | BLUE ∗ (−0.2941) + GREEN ∗ (−0.243) + RED ∗ (−0.5424) + NIR ∗ 0.7276 + SWIR1 ∗ 0.0713 + SWIR2 ∗ (−0.1608) + | [103] |
| Tasseled Cap Wetness (Landsat 8) | TCW | BLUE ∗ 0.1511 + GREEN ∗ 0.1973 + RED ∗ 0.3283 + NIR ∗ 0.3407 + SWIR1 ∗ (−0.7117) + SWIR2 ∗ (−0.4559) | [103] |
3. Results
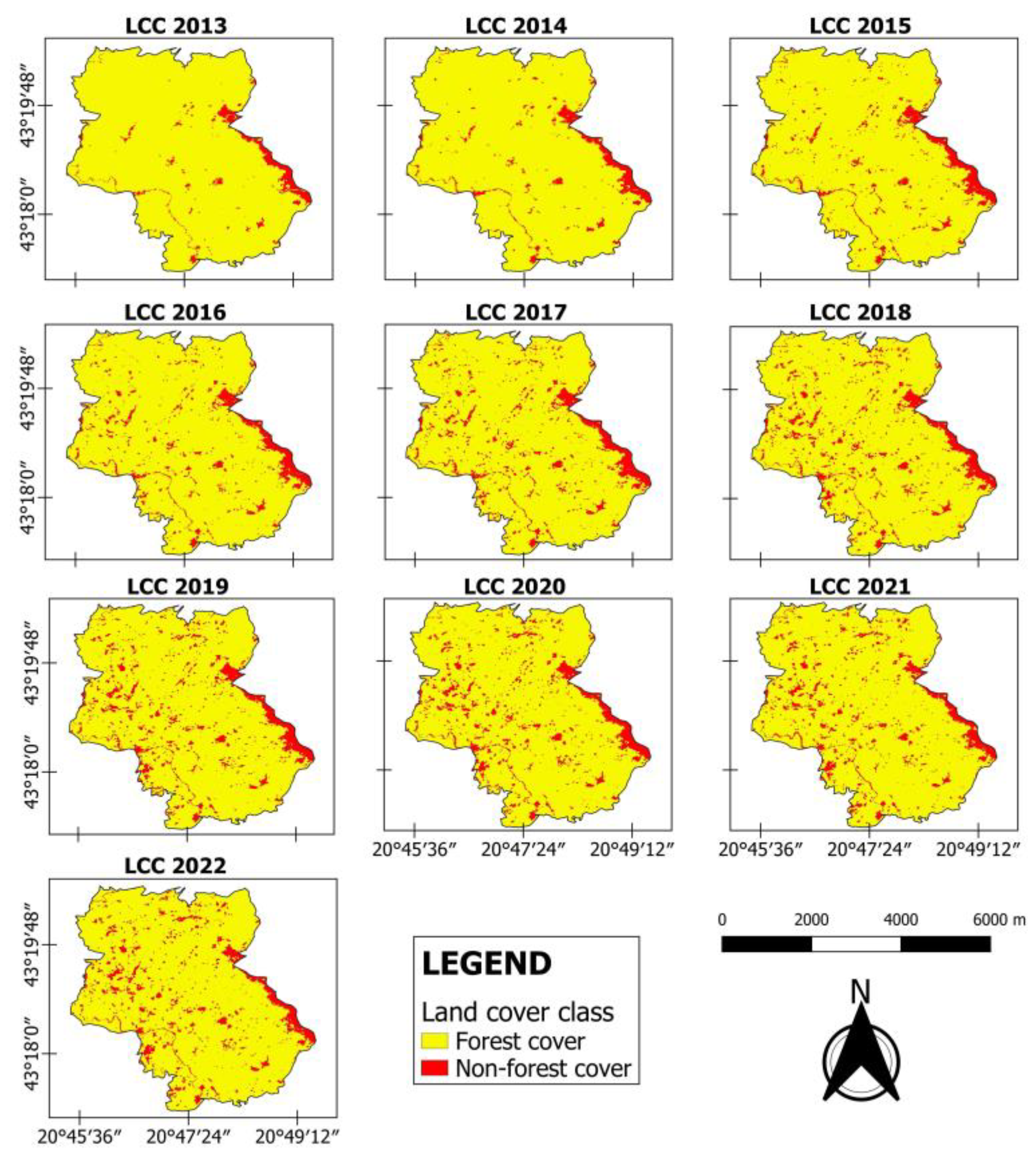
3.1. Forest Cover Loss
3.2. Evaluation of VI Sensitivity in Detecting Responses to Drought and Predicting the Dieback of Norway Spruce
4. Discussion
4.1. Forest Cover Loss
4.2. Evaluation of VI Sensitivity in Detecting Responses to Drought and Predicting the Dieback of Norway Spruce
4.3. Implication for Conservation of Norway Spruce Stands in the Kopaonik NP
4.4. Methodological Limitations of the Used Methodology in Detecting Responses to Drought and Predicting the Dieback of Norway Spruce
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

References
- FAO. Forest Extentand Changes. In Global Forest Resources Assessment 2020: Main Report; FAO: Italy, Rome, 2020; pp. 15–19. Available online: https://www.fao.org/3/ca9825en/ca9825en.pdf (accessed on 12 March 2024). [CrossRef]
- Dudík, R.; Palátová, P.; Jarský, V. Restoration of Declining Spruce Stands in the Czech Republic: A Bioeconomic View on Use of Silver Birch in Case of Small Forest Owners. Austrian J. For. Sci. 2021, 4, 375–394. [Google Scholar]
- Klavina, D.; Menkis, A.; Gaitnieks, T.; Velmala, S.; Lazdins, A.; Rajala, T.; Pennanen, T. Analysis of Norway spruce dieback phenomenon in Latvia—A belowground perspective. Scand. J. For. Res. 2015, 31, 156–165. [Google Scholar] [CrossRef]
- Piedallu, C.; Dallery, D.; Bresson, C.; Legay, M.; Gégout, J.; Pierrat, R. Spatial vulnerability assessment of silver fir and Norway spruce dieback driven by climate warming. Landsc. Ecol. 2023, 38, 341–361. [Google Scholar] [CrossRef]
- Boczoń, A.; Kowalska, A.; Ksepko, M.; Sokołowski, K. Climate Warming and Drought in the Bialowieza Forest from 1950–2015 and Their Impact on the Dieback of Norway Spruce Stands. Water 2018, 10, 1502. [Google Scholar] [CrossRef]
- Rosner, S.; Luss, S.; Světlík, J.; Andreassen, K.; Børja, I.; Dalsgaard, L.; Evans, R.; Tveito, O.E.; Solberg, S. Chronology of hydraulic vulnerability in trunk wood of conifer trees with and without symptoms of top dieback. J. Plant Hydraul. 2016, 3, e001. [Google Scholar] [CrossRef]
- Matić, S. The impact of site changes and management methods on dieback of common spruce (Picea abies Karst.) in Croatia. Croat. J. For. Eng. 2011, 32, 7–16. [Google Scholar]
- Popa, A.; van der Maaten, E.; Popa, I.; van der Maaten-Theunissen, M. Early warning signals indicate climate change-induced stress in Norway spruce in the Eastern Carpathians. Sci. Total Environ. 2024, 912, 169167. [Google Scholar] [CrossRef]
- Caudullo, G.; Tinner, W.; de Rigo, D. Picea abies in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; European Union: Luxembourg, 2016; pp. 114–116. [Google Scholar]
- Jovanović, B. Picea abies Karst. In Dendrology, 6th ed.; Uskoković, Đ., Ed.; Univerzitetska Štampa: Belgrade, Serbia, 2000; pp. 84–89. [Google Scholar]
- Banković, S.; Medarević, M.; Pantić, D.; Petrović, N. Forests by Stand Categories, 1st ed.; National Forest Inventory of the Republic of Serbia, Ministry of Agriculture, Tomter, S., Vasiljević, A., Eds.; Forestry and Water Management of the Republic of Serbia, Forest Directorate: Belgrade, Serbia, 2009; pp. 54–55. [Google Scholar]
- Matović, B. Optimal Condition in Spruce-Fir Forests—Goals and Management Problems at Mountain Zlatar. Master’s Thesis, University of Belgrade, Faculty of Forestry, Belgrade, Serbia, 2005. (In Serbian). [Google Scholar]
- Kolesnikov, B.P. Forest Vegetation in the South-Eastern Part of the Vychegda Basin; Nauka Publ.: Leningrad, Soviet Union, 1985. [Google Scholar]
- Eckstein, D.; Krause, C.; Bauch, J. Dendroecological investigations of spruce trees (Picea abies (L.) Karst.) of different damage and canopy classes. Holzforschung 1989, 43, 411–417. [Google Scholar] [CrossRef]
- Spiecker, H. Growth variation and environmental stresses: Long-term observations on permanent research plots in South-western Germany. Water Air Soil Pollut. 1991, 54, 247–256. [Google Scholar] [CrossRef]
- Kahle, H.P.; Spiecker, H. Adaptability of radial growth of Norway spruce to climate variations: Results of a site specific dendroecological study in the high elevations of the Black Forest (Germany). Radiocarbon 1996, 9999, 785–801. [Google Scholar]
- Mäkinen, H.; Nöjd, P.; Kahle, H.P.; Neumann, U.; Tveite, B.; Mielikäinen, K.; Röhle, H.; Spiecker, H. Radial growth variation of Norway spruce (Picea abies (L.) Karst.) across latitudinal and altitudinal gradients in central and northern Europe. Forest Ecol. Manage. 2002, 171, 243–259. [Google Scholar] [CrossRef]
- Andreassen, K.; Solberg, S.; Tveito, O.E.; Lystad, S.F. Regional differences in climatic responses of Norway spruce (Picea abies (L.) Karst) growth in Norway. Forest Ecol. Manag. 2006, 222, 211–221. [Google Scholar] [CrossRef]
- Pichler, P.; Oberhuber, W. Radial growth response of coniferous forest trees in an inner Alpine environment to the heat-wave in 2003. Forest Ecol. Manag. 2007, 242, 688–699. [Google Scholar] [CrossRef]
- Lebourgeois, L. Climatic signal in annual growth variation of silver fir (Abies alba Mill.) and spruce (Picea abies Karst.) from the French Permanent Plot Network (RENECOFOR). Ann. For. Sci. 2007, 64, 333–343. [Google Scholar] [CrossRef]
- Reichstein, M.; Ciais, P.; Papale, D.; Valentini, R.; Running, S.; Viovy, N.; Cramer, W.; Granier, A.; Ogée, J.; Allard, V.; et al. Reduction of ecosystem productivity and respiration during the European summer 2003 climate anomaly: A joint flux tower, remote sensing and modelling analysis. Glob. Chang. Biol. 2007, 13, 634–651. [Google Scholar] [CrossRef]
- Van der Maaten-Theunissen, M.; Kahle, H.P.; van der Maaten, E. Drought sensitivity of Norway spruce is higher than that of silver fir along an altitudinal gradient in southwestern Germany. Annal. For. Sci. 2013, 70, 185–193. [Google Scholar] [CrossRef]
- Pretzsch, H.; Grams, T.; Häberle, K.H.; Pritsch, K.; Bauerle, T.; Rötzer, T. Growth and mortality of Norway spruce and European beech in monospecific and mixed-species stands under natural episodic and experimentally extended drought. Results of the KROOF throughfall exclusion experiment. Trees 2020, 34, 957–970. [Google Scholar] [CrossRef]
- Begović, K.; Rydval, M.; Mikac, S.; Čupić, S.; Svobodova, K.; Mikoláš, M.; Kozák, D.; Kameniar, O.; Frankovič, M.; Pavlin, J.; et al. Climate-growth relationships of Norway Spruce and silver fir in primary forests of the Croatian Dinaric mountains. Agric. For. Meteorol. 2020, 288–289, 108000. [Google Scholar] [CrossRef]
- Stangler, D.F.; Miller, T.W.; Honer, H.; Larysch, E.; Puhlmann, H.; Seifert, T.; Kahle, H.-P. Multivariate drought stress response of Norway spruce, silver fir and Douglas fir along elevational gradients in Southwestern Germany. Front. Ecol. Evol. 2022, 10, 907492. [Google Scholar] [CrossRef]
- D’Andrea, G.; Šimunek, V.; Pericolo, O.; Vacek, Z.; Vacek, S.; Corleto, R.; Olejár, L.; Ripullone, F. Growth Response of Norway Spruce (Picea abies [L.] Karst.) in Central Bohemia (Czech Republic) to Climate Change. Forests 2023, 14, 1215. [Google Scholar] [CrossRef]
- Bouriaud, O.; Popa, I. Comparative dendroclimatic study of Scots pine, Norway spruce, and silver fir in the Vrancea Range, Eastern Carpathian. Trees 2009, 23, 95–106. [Google Scholar] [CrossRef]
- Kesić, L.; Matović, B.; Stojnić, S.; Stjepanović, S.; Stojanović, D. Climate Change as a Factor Reducing the Growth of Trees in the Pure Norway Spruce Stand (Picea abies (L.) H. Karst.) in the National Park “Kopaonik”. Topola/Poplar 2016, 197–198, 25–34. [Google Scholar]
- Levanič, T.; Gričar, J.; Gagen, M.; Jalkanen, R.; Loader, N.J.; McCarroll, D.; Oven, P.; Robertson, I. The climate sensitivity of Norway spruce [Picea abies (L.) Karst.] in the southeastern European Alps. Trees 2009, 23, 169–180. [Google Scholar] [CrossRef]
- Karadžić, D.; Milanović, S.; Golubović Ćurguz, V. Climatic conditions that preceded the massive dieback of Norway spruce stands at Mt. Golija. In Causes of Spruce (Picea abies Karst.) Dieback in the Area of the Nature Park “Golija”; University of Belgrade, Faculty of Forestry: Belgrade, Serbia, 2017; pp. 15–17. (In Serbian) [Google Scholar]
- Matović, B.; Stojanović, D.; Kesić, L.; Stjepanović, S. Impact of Climate on Growth and Vitality of Norway Spruce at Kopaonik Mountain. Topola/Poplar 2018, 201–202, 99–116. (In Serbian) [Google Scholar]
- Vemić, A.; Milenković, I. Contribution to the Knowledge of Top Dying of Norway Spruce in The Forests of Serbia and Montenegro. Forestry/Šumarstvo 2021, 1–2, 189–199. [Google Scholar]
- Stojanović, B.D.; Orlović, S.; Zlatković, M.; Kostić, S.; Vasić, V.; Miletić, B.; Kesić, L.; Matović, B.; Božanić, D.; Pavlović, L.; et al. Climate change within Serbian forests: Current state and future perspectives. Topola/Poplar 2021, 208, 39–56. [Google Scholar] [CrossRef]
- Zahirović, K.; Dautbašić, M.; Mujezinović, O. Dieback of Spruce Stands Caused by Bark Beetles in Central Bosnia. Our Forests/Naše Šume 2014, 36–37, 4–13. [Google Scholar]
- Dautbašić, M.; Bjelić, M.; Mujezinović, O. Forest Decline in the Area on Zenica—Doboj Canton. Our Forests/Naše šume 2014, 38–39, 5–15. [Google Scholar]
- Ballian, D.; Božić, G. Biochemical Variability of Spruce (Picea abies Karst) in Bosnia and Herzegovina; UŠIT FBiH: Sarajevo, Bosnia and Herzegovina; Silva Slovenica: Ljubljana, Slovenia, 2018; p. 14. [Google Scholar]
- Miletić, B.; Orlović, S.; Lalić, B.; Đurđević, V.; Vujadinović Mandić, M.; Vuković, A.; Gutalj, M.; Stjepanović, S.; Matović, B.; Stojanović, D.B. The potential impact of climate change on the distribution of key tree species in Serbia under RCP 4.5 and RCP 8.5 scenarios. Austrian J. For. Sci. 2021, 138, 183–208. [Google Scholar]
- Assal, T.J.; Anderson, P.J.; Sibold, J. Spatial and temporal trends of drought effects in a heterogeneous semi-arid forest ecosystem. For. Ecol. Manag. 2016, 365, 137–151. [Google Scholar] [CrossRef]
- Hais, M.; Neudertová Hellebrandová, K.; Šrámek, V. Potential of Landsat spectral indices in regard to the detection of forest health changes due to drought effects. J. For. Sci. 2018, 65, 70–78. [Google Scholar] [CrossRef]
- Páscoa, P.; Gouveia, C.M.; Russo, A.C.; Bojariu, R.; Vicente-Serrano, S.M.; Trigo, R.M. Drought Impacts on Vegetation in Southeastern Europe. Remote Sens. 2020, 12, 2156. [Google Scholar] [CrossRef]
- Avetisyan, D.; Borisova, D.; Velizarova, E. Integrated Evaluation of Vegetation Drought Stress through Satellite Remote Sensing. Forests 2021, 12, 974. [Google Scholar] [CrossRef]
- Abdollahnejad, A.; Panagiotidis, D.; Surový, P.; Modlinger, R. Investigating the Correlation between Multisource Remote Sensing Data for Predicting Potential Spread of Ips typographus L. Spots in Healthy Trees. Remote Sens. 2021, 13, 4953. [Google Scholar] [CrossRef]
- DeRose, R.J.; Long, J.N.; Ramsey, R.D. Combining dendrochronological data and the disturbance index to assess Engelmann spruce mortality caused by a spruce beetle outbreak in southern Utah, USA. Remote Sens. Environ. 2011, 115, 2342–2349. [Google Scholar] [CrossRef]
- Lausch, A.; Heurich, M.; Gordalla, D.; Dobner, H.-J.; Gwillym-Margianto, S.; Salbach, C. Forecasting potential bark beetle outbreaks based on spruce forest vitality using hyperspectral remote-sensing techniques at different scales. For. Ecol. Manag. 2013, 308, 76–89. [Google Scholar] [CrossRef]
- Havašová, M.; Bucha, T.; Ferenčík, J.; Jakuš, R. Applicability of a vegetation indices-based method to map bark beetle outbreaks in the High Tatra Mountains. Ann. For. Res. 2015, 58, 295–310. [Google Scholar] [CrossRef]
- Stych, P.; Lastovicka, J.; Hladky, R.; Paluba, D. Evaluation of the Influence of Disturbances on Forest Vegetation Using the Time Series of Landsat Data: A Comparison Study of the Low Tatras and Sumava National Parks. ISPRS Int. J. Geo-Inf. 2019, 8, 71. [Google Scholar] [CrossRef]
- Dalponte, M.; Solano-Correa, Y.T.; Frizzera, L.; Gianelle, D. Mapping a European Spruce Bark Beetle Outbreak Using Sentinel-2 Remote Sensing Data. Remote Sens. 2022, 14, 3135. [Google Scholar] [CrossRef]
- Mandl, L.; Lang, S. Uncovering Early Traces of Bark Beetle Induced Forest Stress via Semantically Enriched Sentinel-2 Data and Spectral Indices. PFG 2023, 91, 211–231. [Google Scholar] [CrossRef]
- Jin, S.; Sader, S.A. Comparison of time series tasseled cap wetness and the normalized difference moisture index in detecting forest disturbances. Remote Sens. Environ. 2005, 94, 364–372. [Google Scholar] [CrossRef]
- Nath, B.; Acharjee, S. Forest Cover Change Detection using Normalized Difference Vegetation Index (NDVI). A Study of Reingkhyongkine Lake’s Adjoining Areas, Rangamati, Bangladesh. Indian Cartogr. 2013, 33, 348–353. [Google Scholar]
- Zhang, K.; Thapa, B.; Ross, M.; Gann, D. Remote sensing of seasonal changes and disturbances in mangrove forest: A case study from South Florida. Ecosphere 2016, 7, e01366. [Google Scholar] [CrossRef]
- Schultz, M.; Clevers, J.G.P.W.; Carter, S.; Verbesselt, J.; Avitabile, V.; Quang, H.V.; Herold, M. Performance of vegetation indices from Landsat time series in deforestation monitoring. Int. J. Appl. Earth Obs. Geoinf. 2016, 52, 318–327. [Google Scholar] [CrossRef]
- Le, T.S.; Harper, R.; Dell, B. Application of Remote Sensing in Detecting and Monitoring Water Stress in Forests. Remote Sens. 2023, 15, 3360. [Google Scholar] [CrossRef]
- Bright, B.C.; Hudak, A.T.; Meddens, A.J.H.; Egan, J.M.; Jorgensen, C.L. Mapping Multiple Insect Outbreaks across Large Regions Annually Using Landsat Time Series Data. Remote Sens. 2020, 12, 1655. [Google Scholar] [CrossRef]
- Candotti, A.; De Giglio, M.; Dubbini, M.; Tomelleri, E. A Sentinel-2 Based Multi-Temporal Monitoring Framework for Wind and Bark Beetle Detection and Damage Mapping. Remote Sens. 2022, 14, 6105. [Google Scholar] [CrossRef]
- Fernandez-Carrillo, A.; Patočka, Z.; Dobrovolný, L.; Franco-Nieto, A.; Revilla-Romero, B. Monitoring Bark Beetle Forest Damage in Central Europe. A Remote Sensing Approach Validated with Field Data. Remote Sens. 2020, 12, 3634. [Google Scholar] [CrossRef]
- Basak, D.; Bose, A.; Roy, S.; Chowdhury, I.R. Chapter 17—Understanding the forest cover dynamics and its health status using GIS-based analytical hierarchy process: A study from Alipurduar district, West Bengal, India. In Water, Land, and Forest Susceptibility and Sustainability; Chatterjee, U., Pradhan, B., Kumar, S., Saha, S., Zakwan, M., Fath, B.D., Fiscus, D., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2023; Volume 1, pp. 475–508. [Google Scholar] [CrossRef]
- Bochenek, Z.; Ziolkowski, D.; Bartold, M.; Orlowska, K.; Ochtyra, A. Monitoring forest biodiversity and the impact of climate on forest environment using high-resolution satellite images. Eur. J. Remote Sens. 2017, 51, 166–181. [Google Scholar] [CrossRef]
- Meng, J.; Li, S.; Wang, W.; Liu, Q.; Xie, S.; Ma, W. Mapping Forest Health Using Spectral and Textural Information Extracted from SPOT-5 Satellite Images. Remote Sens. 2016, 8, 719. [Google Scholar] [CrossRef]
- Forsythe, K.; McCartney, G. Investigating Forest Disturbance Using Landsat Data in the Nagagamisis Central Plateau, Ontario, Canada. ISPRS Int. J. Geo-Inf. 2014, 3, 254–273. [Google Scholar] [CrossRef]
- Jovanović, D.; Govedarica, M.; Đorđević, I.; Pajić, V. Object based image analysis in forestry change detection. In Proceedings of the IEEE 8th International Symposium on Intelligent Systems and Informatics, Subotica, Serbia, 10–11 September 2010; pp. 231–236. [Google Scholar] [CrossRef]
- Jovanović, D.; Govedarica, M.; Sabo, F.; Bugarinović, Ž.; Novović, O.; Beker, T.; Lauter, M. Land cover change detection by using remote sensing: A case study of Zlatibor (Serbia). Geogr. Pannonica 2015, 19, 162–173. [Google Scholar] [CrossRef]
- Valjarević, A.; Djekić, T.; Stevanović, V.; Ivanović, R.; Jandziković, B. GIS numerical and remote sensing analyses of forest changes in the Toplica region for the period of 1953–2013. Appl. Geogr. 2018, 92, 131–139. [Google Scholar] [CrossRef]
- Milanović, M.M.; Micić, T.; Lukić, T.; Nenadović, S.S.; Basarin, B.; Filipović, D.J.; Tomić, M.; Samardžić, I.; Srdić, Z.; Nikolić, G.; et al. Application of Landsat-Derived NDVI in Monitoring and Assessment of Vegetation Cover Changes in Central Serbia. Carpathian J. Earth Environ. Sci. 2018, 14, 119–129. [Google Scholar] [CrossRef]
- Kovačević, J.; Cvijetinović, Ž.; Lakušić, D.; Kuzmanović, N.; Šinžar-Sekulić, J.; Mitrović, M.; Stančić, N.; Brodić, N.; Mihajlović, D. Spatio-Temporal Classification Framework for Mapping Woody Vegetation from Multi-Temporal Sentinel-2 Imagery. Remote Sens. 2020, 12, 2845. [Google Scholar] [CrossRef]
- Jovanović, D.; Gavrilović, M.; Borisov, M.; Govedarica, M. Deforestation Monitoring with Sentinel 1 and Sentinel 2 Images—The Case Study of Fruska Gora (Serbia). Šumarski List. 2021, 3–4, 127–135. [Google Scholar] [CrossRef]
- Potić, I.; Srdić, Z.; Vakanjac, B.; Bakrač, S.; Ðordević, D.; Banković, R.; Jovanović, J.M. Improving Forest Detection Using Machine Learning and Remote Sensing: A Case Study in Southeastern Serbia. Appl. Sci. 2023, 13, 8289. [Google Scholar] [CrossRef]
- Jovanović, M.M.; Milanović, M.M. Remote Sensing and Forest Conservation: Challenges of Illegal Logging in Kursumlija Municipality (Serbia). In Forest Ecology and Conservation; Chakravarty, S., Shukla, G., Eds.; IntechOpen: Rijeka, Croatia, 2017; pp. 99–118. [Google Scholar] [CrossRef]
- Jovanović, M.M.; Milanović, M.M.; Vračarević, R.B. Chapter 1—Comparing NDVI and Corine Land Cover as Tools for Improving National Forest Inventory Updates and Preventing Illegal Logging in Serbia. In Vegetation; Sebata, A., Ed.; IntechOpen: London, UK, 2017; pp. 1–22. [Google Scholar] [CrossRef]
- Potić, I.; Ćurčić, N.; Potić, M.; Radovanović, M.; Tretiakova, T. Remote sensing role in environmental stress analysis: East Serbia wildfires case study (2007–2017). J. Geogr. Inst. “Jovan Cvijić” SASA 2017, 67, 249–264. [Google Scholar] [CrossRef]
- Brovkina, O.; Stojanović, M.; Milanović, S.; Latypov, I.; Marković, N.; Cienciala, E. Monitoring of post-fire forest scars in Serbia based on satellite Sentinel-2 data. Geomat. Nat. Hazards Risk 2020, 11, 2315–2339. [Google Scholar] [CrossRef]
- Jovanović, M.M.; Milanović, M.M. Normalized Difference Vegetation Index (NDVI) as the Basis for Local Forest Management. Example of the Municipality of Topola, Serbia. Pol. J. Environ. Stud. 2015, 24, 529–535. [Google Scholar]
- Nonić, D.; Šumarac, P.; Ranković, N.; Đorđević, I.; Nedeljković, J. Sustainable Management of the National Park Kopaonik—Opportunities and Challenges; Special edition; Bulletin of the Faculty of Forestry, University of Belgrade: Belgrade, Serbia, 2023; pp. 59–80, (In Serbian). [Google Scholar] [CrossRef]
- Horwath Consulting Zagreb (HCS). Master Plan for the Kopaonik Tourist Destination (Final Report); Ordering Party: Ministry of Economy and Regional Development of the Republic of Serbia: Belgrade, Serbia, 2009; pp. 14–20. (In Serbian) [Google Scholar]
- Đorđević, N.; Lakićević, N.; Milićević, S. Benchmarking analysis of tourism in national parks Tara and Kopaonik. Ekon. Teor. I Praksa 2018, 3, 52–70. (In Serbian) [Google Scholar] [CrossRef]
- Ostojić, D.; Krsteski, B.; Dinić, A.; Perković, A. Vegetation Characteristics of Forest Ecosystems on “Kopaonik” National Park with the Reference to the Forests with the Protection Regime Level I. Šumarstvo/Forestry 2018, 3–4, 179–194. (In Serbian) [Google Scholar]
- MEP (Ministry of Environmental Protection); PE (Public Enterprise) National Park ‘’Kopaonik”. National Park Kopaonik Management Program for 2021, p. 12. 2020. Available online: https://npkopaonik.rs/wp-content/uploads/2021/08/Program-upravljanja-2021..pdf (accessed on 15 January 2024). (In Serbian).
- Republic Hydrometeorological Service of the Republic of Serbia (RHMS). Climatology: 30 Years Averages (1991–2020). 2024. Available online: https://www.hidmet.gov.rs/latin/meteorologija/klimatologija_srednjaci.php (accessed on 13 March 2024). (In Serbian)
- Republic Hydrometeorological Service of the Republic of Serbia (RHMS). Climatology: 30 Years Averages (1961–1990). 2024. Available online: http://www.hidmet.gov.rs/eng/meteorologija/klimatologija_srednjaci.php (accessed on 13 March 2024).
- Congedo, L. Semi-Automatic Classification Plugin: A Python tool for the download and processing of remote sensing images in QGIS. J. Open Source Softw. 2021, 6, 3172. [Google Scholar] [CrossRef]
- Chavez, P.S. Image-Based Atmospheric Corrections—Revisited and Improved Photogrammetric Engineering and Remote Sensing, [Falls Church, Va.]. Am. Soc. Photogramm. 1996, 62, 1025–1036. [Google Scholar]
- Roy, D.P.; Kovalskyy, V.; Zhang, H.K.; Vermote, E.F.; Yan, L.; Kumar, S.S.; Egorov, A. Characterization of Landsat-7 to Landsat-8 reflective wavelength and normalized difference vegetation index continuity. Remote Sens. Environ. 2016, 185, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.; Tateishi, R.; Hoan, N. Satellite Image Pansharpening Using a Hybrid Approach for Object-Based Image Analysis. ISPRS Int. J. Geo-Inf. 2012, 1, 228–241. [Google Scholar] [CrossRef]
- Rahaman, K.; Hassan, Q.; Ahmed, M. Pan-Sharpening of Landsat-8 Images and Its Application in Calculating Vegetation Greenness and Canopy Water Contents. ISPRS Int. J. Geo-Inf. 2017, 6, 168. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 2 November 2023).
- Hijmans, R. Raster: Geographic Data Analysis and Modeling. R Package Version 3.6-26. 2023. Available online: https://CRAN.R-project.org/package=raster (accessed on 8 January 2024).
- Richards, A.J.; Jia, X. Remote Sensing Digital Image Analysis, 4th ed.; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar] [CrossRef]
- Nagamani, K.; Jayakumar, K.; Suresh, Y.; Sriganesh, J. Study on Error Matrix Analysis of Classified Remote Sensed Data for Pondicherry Coast. J. Adv. Res. GeoSci. Rem. Sens. 2015, 2, 3–4. [Google Scholar]
- Cochran, W.G. Sampling Techniques, 3rd ed.; John Wiley & Sons: New York, NY, USA, 1977. [Google Scholar]
- Olofsson, P.; Foody, G.M.; Stehman, S.V.; Woodcock, C.E. Making better use of accuracy data in land change studies: Estimating accuracy and area and quantifying uncertaintyusing stratified estimation. Remote Sens. Environ. 2013, 129, 122–131. [Google Scholar] [CrossRef]
- Pebesma, E.; Bivand, R. Introduction to sf and stars. In Spatial Data Science: With Applications in R, 1st ed.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2023. [Google Scholar] [CrossRef]
- Neuwirth, E. RColorBrewer: ColorBrewer Palettes. R Package Version 1.1-3. 2022. Available online: https://CRAN.R-project.org/package=RColorBrewer (accessed on 8 January 2024).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; Available online: https://ggplot2.tidyverse.org (accessed on 8 January 2024).
- Aphalo, P. ggpmisc: Miscellaneous Extensions to ‘ggplot2’. R Package Version 0.5.5. 2023. Available online: https://CRAN.R-project.org/package=ggpmisc (accessed on 8 January 2024).
- Pedersen, T. patchwork: The Composer of Plots. R Package Version 1.2.0. 2024. Available online: https://CRAN.R-project.org/package=patchwork (accessed on 8 January 2024).
- Iannone, R.; Cheng, J.; Schloerke, B.; Hughes, E.; Lauer, A.; Seo, J. gt: Easily Create Presentation-Ready Display Tables. R Package Version 0.10. 2023. Available online: https://CRAN.R-project.org/package=gt (accessed on 8 January 2024).
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.M.; Tucker, C.J.; Stenseth, N.C. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends Ecol. Evol. 2005, 20, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Tuominen, J.; Lipping, T.; Kuosmanen, V.; Haapane, R. Remote Sensing of Forest Health. In Geoscience and Remote Sensing, 1st ed.; Ho, P., Ed.; InTech: London, UK, 2009; pp. 29–52. [Google Scholar] [CrossRef]
- Lambert, J.; Drenou, C.; Denux, J.-P.; Balent, G.; Cheret, V. Monitoring forest decline through remote sensing time series analysis. GIScience Remote Sens. 2013, 50, 437–457. [Google Scholar] [CrossRef]
- Wang, L.; Luo, Y.Q.; Huang, H.G.; Shi, J.; Keliövaara, K.; Teng, W.X.; Qi, G.X. Reflectance features of water stressed Larix gmelinii needles. For. Stud. China 2009, 11, 28–33. [Google Scholar] [CrossRef]
- Wang, L.; Qu, J.J. NMDI: A normalized multi-band drought index for monitoring soil and vegetation moisture with satellite remote sensing. Geophys. Res. Lett. 2007, 34, L20405. [Google Scholar] [CrossRef]
- Baig, M.H.A.; Zhang, L.; Shuai, T.; Tong, Q. Derivation of a tasselled cap transformation based on Landsat 8 at-satellite reflectance. Remote Sens. Lett. 2014, 5, 423–431. [Google Scholar] [CrossRef]
- Lastovicka, J.; Svec, P.; Paluba, D.; Kobliuk, N.; Svoboda, J.; Hladky, R.; Stych, P. Sentinel-2 Data in an Evaluation of the Impact of the Disturbances on Forest Vegetation. Remote Sens. 2020, 12, 1914. [Google Scholar] [CrossRef]
- Healey, S.; Cohen, W.; Zhiqiang, Y.; Krankina, O. Comparison of Tasseled Cap-based Landsat data structures for use in forest disturbance detection. Remote Sens. Environ. 2005, 97, 301–310. [Google Scholar] [CrossRef]
- Wickham, H.; Bryan, J. readxl: Read Excel Files. R Package Version 1.4.3. 2023. Available online: https://CRAN.R-project.org/package=readxl (accessed on 8 January 2024).
- Welikhe, P.; Quansah, J.E.; Fall, S.; McElhenney, W. Estimation of Soil Moisture Percentage Using LANDSAT-based Moisture Stress Index. J. Remote Sens. GIS 2017, 6, 1–5. [Google Scholar] [CrossRef]
- Jiang, Z.; Huete, A.R.; Chen, J.; Chen, Y.; Li, J.; Yan, G.; Zhang, X. Analysis of NDVI and scaled difference vegetation index retrievals of vegetation fraction. Remote Sens. Environ. 2006, 101, 366–378. [Google Scholar] [CrossRef]
- Fraga, H.; Amraoui, M.; Malheiro, A.C.; Moutinho-Pereira, J.; Eiras-Dias, J.; Silvestre, J.; Santos, J.A. Examining the relationship between the Enhanced Vegetation Index and grapevine phenology. Eur. J. Remote Sens. 2014, 47, 753–771. [Google Scholar] [CrossRef]
- Mancino, G.; Ferrara, A.; Padula, A.; Nolè, A. Cross-Comparison between Landsat 8 (OLI) and Landsat 7 (ETM+) Derived Vegetation Indices in a Mediterranean Environment. Remote Sens. 2020, 12, 291. [Google Scholar] [CrossRef]
- Cohen, J. The t Test for Means. In Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Lawrence Erlbaum Associates: Hillsdale, NJ, USA, 1988; pp. 19–27. [Google Scholar]
- Navarro, D.J. Learning Statistics with R: A Tutorial for Psychology Students and Other Beginners; Version 0.6; University of New South Wales: Sydney, Australia, 2015; Available online: https://learningstatisticswithr.com (accessed on 8 January 2024).
- Sawilowsky, S. New effect size rules of thumb. J. Mod. Appl. Stat. Methods 2009, 8, 467–474. [Google Scholar] [CrossRef]
- KC, A.; Wagle, N.; Acharya, T.D. Spatiotemporal Analysis of Land Cover and the Effects on Ecosystem Service Values in Rupandehi, Nepal from 2005 to 2020. ISPRS Int. J. Geo-Inf. 2021, 10, 635. [Google Scholar] [CrossRef]
- Marini, L.; Økland, B.; Jönsson, A.M.; Bentz, B.; Carroll, A.; Forster, B.; Grégoire, J.C.; Hurling, R.; Nageleisen, L.M.; Netherer, S.; et al. Climate drivers of bark beetle outbreak dynamics in Norway spruce forests. Ecography 2017, 40, 1426–1435. [Google Scholar] [CrossRef]
- Hlásny, T.; Zimová, S.; Merganičová, K.; Štěpánek, P.; Modlinger, R.; Turčáni, M. Devastating outbreak of bark beetles in the Czech Republic: Drivers, impacts, and management implications. For. Ecol. Manag. 2021, 490, 119075. [Google Scholar] [CrossRef]
- Seidl, R.; Müller, J.; Hothorn, T.; Bässler, C.; Heurich, M.; Kautz, M. Small beetle, large-scale drivers: How regional and landscape factors affect outbreaks of the European spruce bark beetle. J. Appl. Ecol. 2015, 53, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Netherer, S.; Lehmanski, L.; Bachlehner, A.; Rosner, S.; Savi, T.; Schmidt, A.; Huang, J.; Paiva, M.R.; Mateus, E.; Hartmann, H.; et al. Drought increases Norway spruce susceptibility to the Eurasian spruce bark beetle and its associated fungi. New Phytol. 2024, 242, 1000–1017. [Google Scholar] [CrossRef] [PubMed]
- Moreno, A.; Maselli, F.; Chiesi, M.; Genesio, L.; Vaccari, F.; Seufert, G.; Gilabert, M.A. Monitoring water stress in Mediterranean semi-natural vegetation with satellite and meteorological data. Int. J. Appl. Earth Obs. Geoinf. 2014, 26, 246–255. [Google Scholar] [CrossRef]
- Dotzler, S.; Hill, J.; Buddenbaum, H.; Stoffels, J. The Potential of EnMAP and Sentinel-2 Data for Detecting Drought Stress Phenomena in Deciduous Forest Communities. Remote Sens. 2015, 7, 14227–14258. [Google Scholar] [CrossRef]
- Rehschuh, R.; Mette, T.; Menzel, A.; Buras, A. Soil properties affect the drought susceptibility of Norway spruce. Dendrochronologia 2017, 45, 81–89. [Google Scholar] [CrossRef]
- Kiseleva, V.; Korotkov, S.; Stonozhenko, L.; Naidenova, E. Structure and regeneration of spruce forests as affected by forest management practices in the Moscow Region. IOP Conf. Ser. Earth Environ. Sci. 2019, 226, 012042. [Google Scholar] [CrossRef]
- Chernenkova, T.; Kotlov, I.; Belyaeva, N.; Suslova, E.; Morozova, O.; Pesterova, O.; Arkhipova, M. Role of Silviculture in the Formation of Norway Spruce Forests along the Southern Edge of Their Range in the Central Russian Plain. Forests 2020, 11, 778. [Google Scholar] [CrossRef]
- Neuner, S.; Albrecht, A.; Cullmann, D.; Engels, F.; Griess, V.C.; Hahn, W.A.; Hanewinkel, M.; Härtl, F.; Kölling, C.; Staupendahl, K.; et al. Survival of Norway spruce remains higher in mixed stands under a dryer and warmer climate. Glob. Change Biol. 2014, 21, 935–946. [Google Scholar] [CrossRef] [PubMed]
- Tanovski, V.; Matović, B.; Kesić, L.; Stojanović, D.B. A review of the influence of climate change on coniferous forests in the Balkan peninsula. Topola/Poplar 2022, 210, 41–64. [Google Scholar] [CrossRef]





| Sensor | Date Acquired | Path/Row |
|---|---|---|
| Landsat 7 | 22 August 2009 | 186/030 |
| Landsat 7 | 21 July 2009 | 186/030 |
| Landsat 7 | 19 June 2009 | 186/030 |
| Landsat 7 | 21 August 2011 | 185/030 |
| Landsat 7 | 27 July 2011 | 186/030 |
| Landsat 7 | 12 August 2011 | 186/030 |
| Landsat 7 | 28 August 2011 | 186/030 |
| Landsat 7 | 23 August 2012 | 185/030 |
| Landsat 7 | 29 July 2012 | 186/030 |
| Landsat 7 | 14 August 2012 | 186/030 |
| Landsat 7 | 30 August 2012 | 186/030 |
| Landsat 8 | 01 July 2013 | 185/030 |
| Landsat 8 | 25 August 2013 | 186/030 |
| Landsat 8 | 28 August 2014 | 186/030 |
| Landsat 8 | 09 August 2013 | 186/030 |
| Landsat 8 | 12 August 2014 | 186/030 |
| Sensor | Date Acquired | Path/Row |
|---|---|---|
| Landsat 8 | 09 August 2013 | 186/030 |
| Landsat 8 | 12 August 2014 | 186/030 |
| Sentinel 2 | 31 August 2015 | 136 * |
| Sentinel 2 | 15 August 2016 | 136 * |
| Sentinel 2 | 11 July 2017 | 136 * |
| Sentinel 2 | 21 July 2018 | 136 * |
| Sentinel 2 | 01 July 2019 | 136 * |
| Sentinel 2 | 29 August 2020 | 136 * |
| Sentinel 2 | 25 July 2021 | 136 * |
| Sentinel 2 | 20 July 2022 | 136 * |
| Year | MEAN (ha) | MIN (ha) | MAX (ha) | SD (ha) |
|---|---|---|---|---|
| 2013 | / | / | / | / |
| 2014 | 0.05 | 0.02 | 0.74 | 0.07 |
| 2015 | 0.07 | 0.01 | 0.73 | 0.10 |
| 2016 | 0.07 | 0.01 | 1.60 | 0.15 |
| 2017 | 0.08 | 0.01 | 1.90 | 0.16 |
| 2018 | 0.12 | 0.01 | 3.87 | 0.28 |
| 2019 | 0.11 | 0.01 | 4.60 | 0.29 |
| 2020 | 0.12 | 0.01 | 4.74 | 0.32 |
| 2021 | 0.14 | 0.01 | 5.49 | 0.35 |
| 2022 | 0.13 | 0.01 | 5.72 | 0.37 |
| Year | OA | PA/F | PA/N | UA/F | UA/N |
|---|---|---|---|---|---|
| 2013 | 97.9 | 99.84 | 69.28 | 97.97 | 96.61 |
| 2014 | 97.79 | 99.56 | 73.47 | 98.1 | 92.41 |
| 2015 | 95.97 | 99.37 | 66.14 | 96.27 | 92.28 |
| 2016 | 96.78 | 99.61 | 72.22 | 96.9 | 95.47 |
| 2017 | 92.49 | 99.17 | 56.04 | 92.49 | 92.49 |
| 2018 | 94.1 | 99.01 | 68 | 95.29 | 91.27 |
| 2019 | 95.61 | 99.12 | 72.28 | 95.96 | 92.52 |
| 2020 | 91.83 | 99.61 | 55.63 | 91.26 | 96.88 |
| 2021 | 92.7 | 99.41 | 56.82 | 92.49 | 94.76 |
| 2022 | 96.63 | 99.43 | 76.3 | 96.82 | 94.88 |
| Year | LCC | Forest | Non-Forest | Total (Wi) | Am,i [pixel] | Am,i [ha] |
|---|---|---|---|---|---|---|
| 2013 | Forest | 0.935 | 0.019 | 0.954 | 101,142 | 2276 |
| Non-forest | 0.002 | 0.044 | 0.046 | 4804 | 108 | |
| Total | 0.937 | 0.063 | 1 | 105,946 | 2384 | |
| 2014 | Forest | 0.928 | 0.018 | 0.946 | 100,211 | 2255 |
| Non-forest | 0.004 | 0.050 | 0.054 | 5710 | 128 | |
| Total | 0.932 | 0.068 | 1 | 105,921 | 2383 | |
| 2015 | Forest | 0.892 | 0.034 | 0.926 | 220,937 | 2209 |
| Non-forest | 0.006 | 0.068 | 0.074 | 17,466 | 175 | |
| Total | 0.898 | 0.102 | 1 | 238,403 | 2384 | |
| 2016 | Forest | 0.893 | 0.029 | 0.922 | 219,819 | 2198 |
| Non-forest | 0.004 | 0.074 | 0.078 | 18,584 | 186 | |
| Total | 0.897 | 0.103 | 1 | 238,403 | 2384 | |
| 2017 | Forest | 0.838 | 0.068 | 0.906 | 216,032 | 2160 |
| Non-forest | 0.007 | 0.087 | 0.094 | 22,371 | 224 | |
| Total | 0.845 | 0.155 | 1 | 238,403 | 2384 | |
| 2018 | Forest | 0.859 | 0.042 | 0.901 | 214,862 | 2149 |
| Non-forest | 0.009 | 0.09 | 0.099 | 23,541 | 235 | |
| Total | 0.868 | 0.132 | 1 | 238,403 | 2384 | |
| 2019 | Forest | 0.862 | 0.035 | 0.897 | 214,007 | 2140 |
| Non-forest | 0.008 | 0.095 | 0.103 | 24,366 | 244 | |
| Total | 0.87 | 0.130 | 1 | 238,373 | 2384 | |
| 2020 | Forest | 0.82 | 0.079 | 0.899 | 214,178 | 2142 |
| Non-forest | 0.003 | 0.098 | 0.101 | 24,225 | 242 | |
| Total | 0.823 | 0.177 | 1 | 238,403 | 2384 | |
| 2021 | Forest | 0.838 | 0.068 | 0.906 | 215,885 | 2159 |
| Non-forest | 0.005 | 0.090 | 0.095 | 22,518 | 225 | |
| Total | 0.843 | 0.157 | 1 | 238,403 | 2384 | |
| 2022 | Forest | 0.874 | 0.029 | 0.903 | 215,167 | 2152 |
| Non-forest | 0.005 | 0.093 | 0.098 | 23,236 | 232 | |
| Total | 0.879 | 0.122 | 1 | 238,403 | 2384 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miletić, B.R.; Matović, B.; Orlović, S.; Gutalj, M.; Đorem, T.; Marinković, G.; Simović, S.; Dugalić, M.; Stojanović, D.B. Quantifying Forest Cover Loss as a Response to Drought and Dieback of Norway Spruce and Evaluating Sensitivity of Various Vegetation Indices Using Remote Sensing. Forests 2024, 15, 662. https://doi.org/10.3390/f15040662
Miletić BR, Matović B, Orlović S, Gutalj M, Đorem T, Marinković G, Simović S, Dugalić M, Stojanović DB. Quantifying Forest Cover Loss as a Response to Drought and Dieback of Norway Spruce and Evaluating Sensitivity of Various Vegetation Indices Using Remote Sensing. Forests. 2024; 15(4):662. https://doi.org/10.3390/f15040662
Chicago/Turabian StyleMiletić, Boban R., Bratislav Matović, Saša Orlović, Marko Gutalj, Todor Đorem, Goran Marinković, Srđan Simović, Mirko Dugalić, and Dejan B. Stojanović. 2024. "Quantifying Forest Cover Loss as a Response to Drought and Dieback of Norway Spruce and Evaluating Sensitivity of Various Vegetation Indices Using Remote Sensing" Forests 15, no. 4: 662. https://doi.org/10.3390/f15040662
APA StyleMiletić, B. R., Matović, B., Orlović, S., Gutalj, M., Đorem, T., Marinković, G., Simović, S., Dugalić, M., & Stojanović, D. B. (2024). Quantifying Forest Cover Loss as a Response to Drought and Dieback of Norway Spruce and Evaluating Sensitivity of Various Vegetation Indices Using Remote Sensing. Forests, 15(4), 662. https://doi.org/10.3390/f15040662