
Insights into Canopy Escape Ratio from Canopy Structures: Correlations Uncovered through Sentinel-2 and Field Observation


Abstract
:1. Introduction
2. Study Area and Data
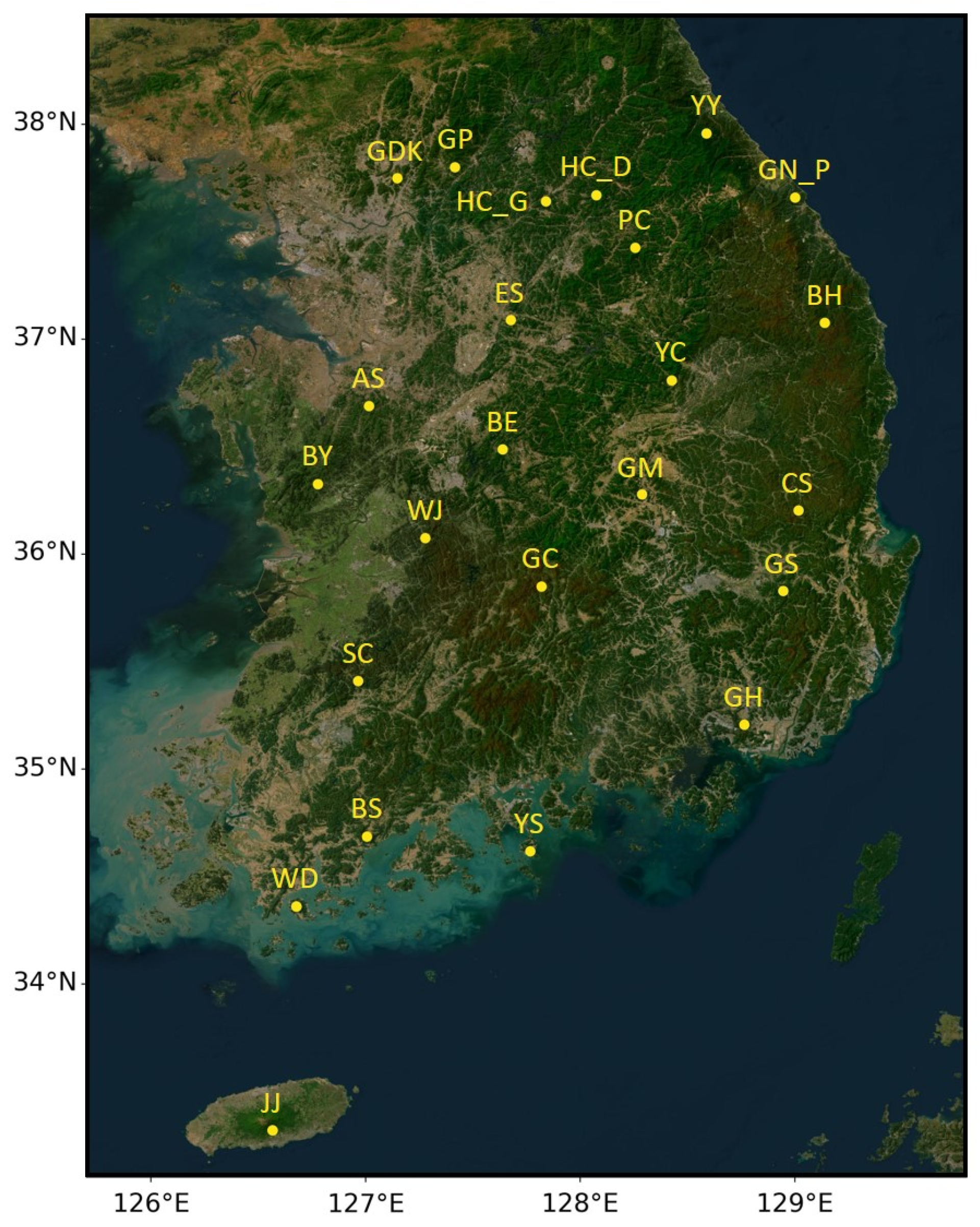
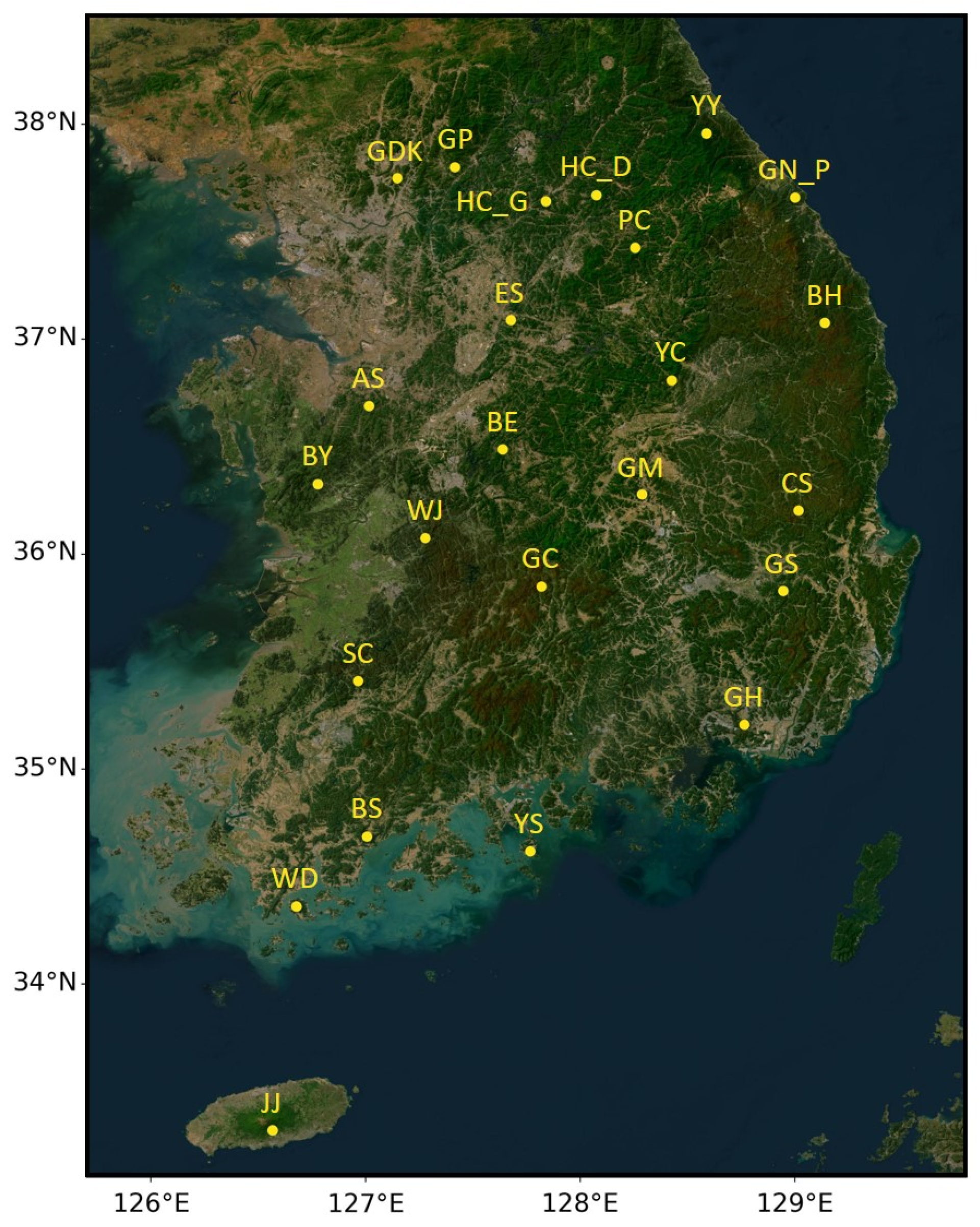
2.1. Study Sites with Ground Observation Network of LAI

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Name | Abbreviations | Longitude | Latitude | Köppen Climate Classification | Forest Type | Observation Period |
|---|---|---|---|---|---|---|
| Asan | AS | 127.0165 | 36.6893 | Dwa | MF | 23.08.27–23.11.24 23.11.29–23.11.30 |
| Boeun | BE | 127.6389 | 36.4875 | Dwa | DBF | 23.11.11–23.11.30 |
| Bonghwa | BH | 129.1394 | 37.0758 | Dfb | DBF | 23.06.18–23.09.30 23.11.07–23.11.30 |
| Boseong | BS | 127.0067 | 34.6854 | Cwa | DBF | 23.04.30–23.08.01 23.11.10–23.11.24 23.11.29–23.11.30 |
| Buyeo | BY | 126.7785 | 36.3257 | Dwa | MF | 23.08.27–23.11.24 |
| Cheongsong | CS | 129.0172 | 36.2025 | Dwa | MF | 23.03.11–23.08.02 23.08–03-23.10.12 23.11.08–23.11.24 23.11.29–23.11.30 |
| Eumseong | ES | 127.6764 | 37.0882 | Dwa | DBF | 23.08.02–23.11.24 23.11.29–23.11.30 |
| Gangneung_P | GN_P | 129.0010 | 37.6589 | Dfa | MF | 23.06.01–23.07.27 23.08.11–23.08.16 23.11.30 |
| Gapyeong | GP | 127.4168 | 37.8005 | Dwa | DBF | 23.09.23–23.11.30 |
| Geochang | GC | 127.8192 | 35.8495 | Dwa | DBF | 23.02.26–23.10.01 23.11.09–23.11.30 |
| Gimhae | GH | 128.7647 | 35.2053 | Cwa | DBF | 23.06.19–23.11.24 23.11.29–23.11.30 |
| Gumi | GM | 128.2876 | 36.2780 | Dwa | MF | 23.03.12–23.08.05 23.08.25–23.08.27 23.11.09–23.11.15 |
| Gwangneung | GDK | 127.1487 | 37.7488 | Dwa | DBF | 21.12.17–23.03.24 |
| Gyeongsan | GS | 128.9451 | 35.8286 | Dwa | ENF | 23.03.12–23.08.30 23.11.09–23.11.30 |
| Hongcheon_D | HC_D | 128.0748 | 37.6695 | Dwa | DBF | 23.08.08–23.11.24 23.11.29–23.11.30 |
| Hongcheon_G | HC_G | 127.8407 | 37.6410 | Dwa | DBF | 23.08.08–23.11.24 23.11.29–23.11.30 |
| Jeju | JJ1 | 126.5677 | 33.3179 | Cfa | MF | 22.08.26–23.07.25 23.09.05–23.11.24 23.11.29–23.11.30 |
| JJ2 | 126.5676 | 33.3178 | Cfa | MF | 22.08.26–23.07.25 23.09.05–23.11.24 23.11.29–23.11.30 | |
| JJ3 | 126.5675 | 33.3178 | Cfa | EBF | 22.08.26–23.07.25 | |
| Pyeongchang | PC | 128.2559 | 37.4258 | Dwb | DNF | 23.06.01–23.07.07 23.07.15–23.07.20 23.07.29–23.08.03 23.09.09–23.09.17 |
| Sunchang | SC | 126.9664 | 35.4098 | Dfa | DBF | 23.05.27–23.11.24 23.11.29–23.11.30 |
| Wando | WD1 | 126.6779 | 34.3594 | Cfa | EBF | 22.11.01–23.06.15 23.08.25–23.08.27 23.11.11–23.11.16 |
| WD2 | 126.6776 | 34.3594 | Cfa | EBF | 22.11.01–23.06.15 23.08.25–23.08.27 23.11.11–23.11.24 23.11.29–23.11.30 | |
| WD3 | 126.6778 | 34.3593 | Cfa | EBF | 23.05.01–23.07.02 23.08.25–23.09.04 23.11.11–23.11.24 23.11.29–23.11.30 | |
| WD4 | 126.6777 | 34.3592 | Cfa | EBF | 22.11.01–23.07.02 23.08.25–23.09.04 23.11.11–23.11.24 23.11.29–23.11.30 | |
| WD5 | 126.6779 | 34.3593 | Cfa | EBF | 22.11.01–23.05.04 23.11.11–23.11.24 23.11.29–23.11.30 | |
| Wanju | WJ | 127.2778 | 36.0748 | Dwa | DBF | 23.02.25–23.05.14 23.05.22–23.05.26 23.06.05 23.08.26–23.11.24 23.11.29–23.11.30 |
| YangYang | YY | 128.5888 | 37.9569 | Dfb | DBF | 23.09.22–23.11.24 23.11.29–23.11.30 |
| Yecheon | YC | 128.4259 | 36.8083 | Dwa | DBF | 23.03.13–23.11.24 23.11.29–23.11.30 |
| Yeosu | YS | 127.7676 | 34.6162 | Cwa | MF | 23.06.04–23.11.24 23.11.29–23.11.30 |
2.2. DEM
2.3. Sentinel-2 Vegetation Indices
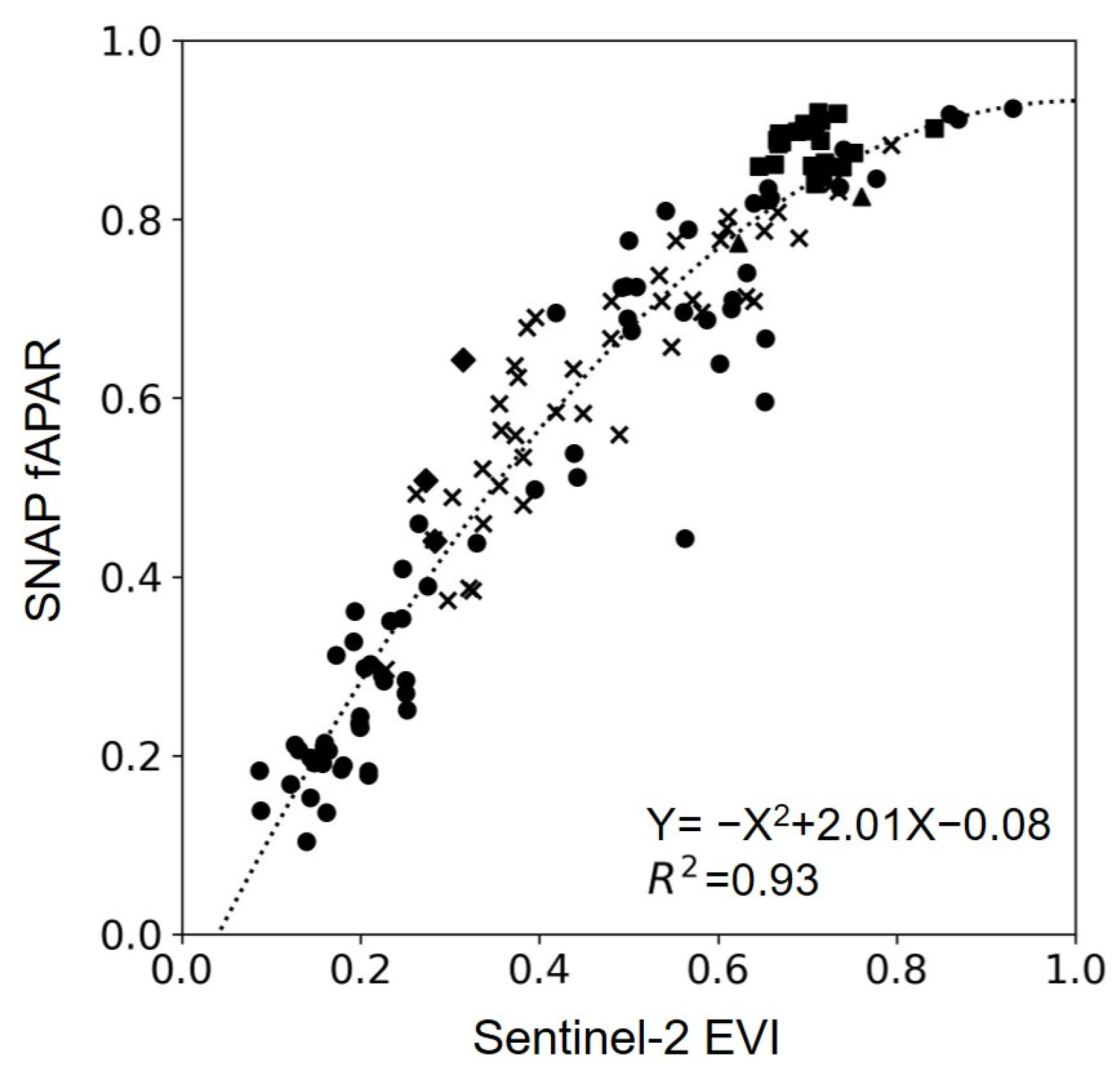
2.4. SNAP LAI and fAPAR
2.5. Tower-Based Multispectral Reflectance
3. Methods
3.1. fesc Calculation
3.2. Comparison between Canopy Structure and fesc
4. Results
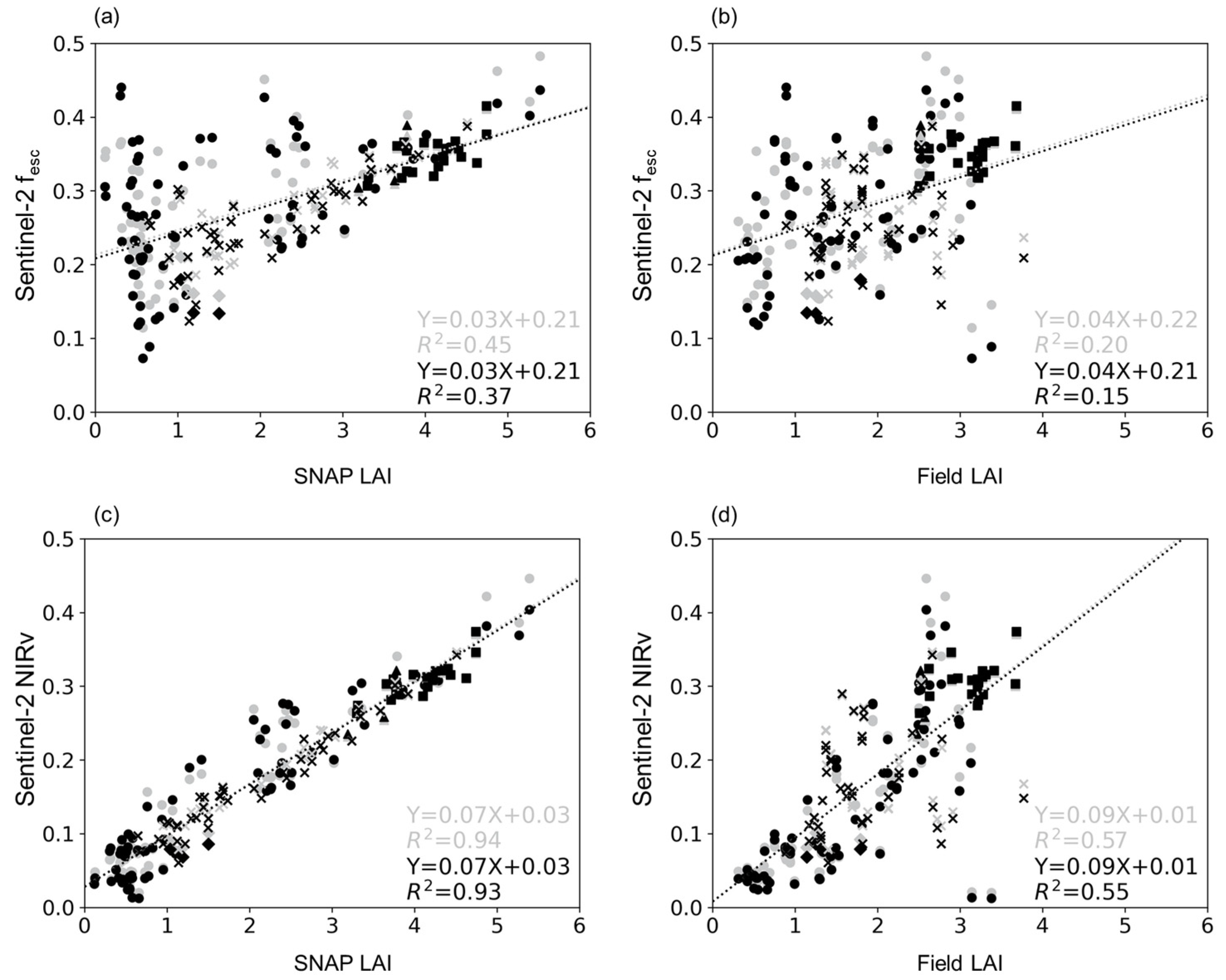
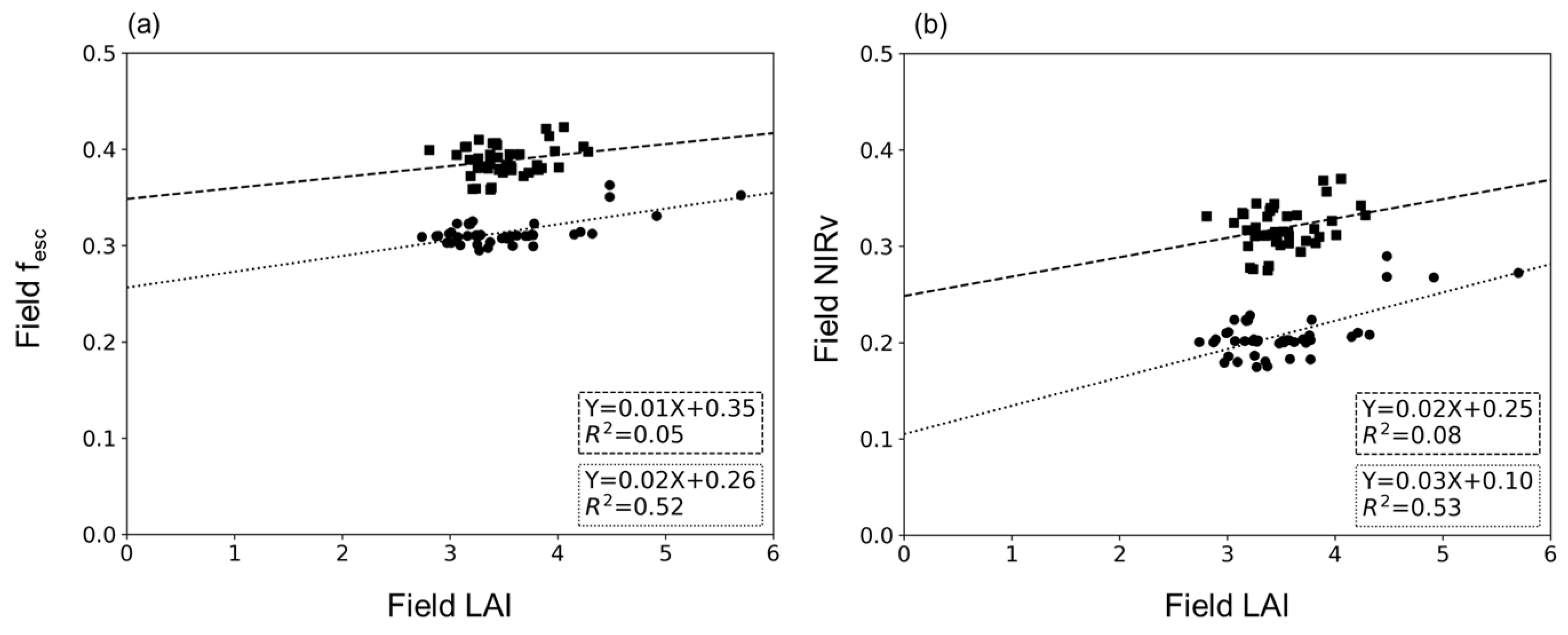
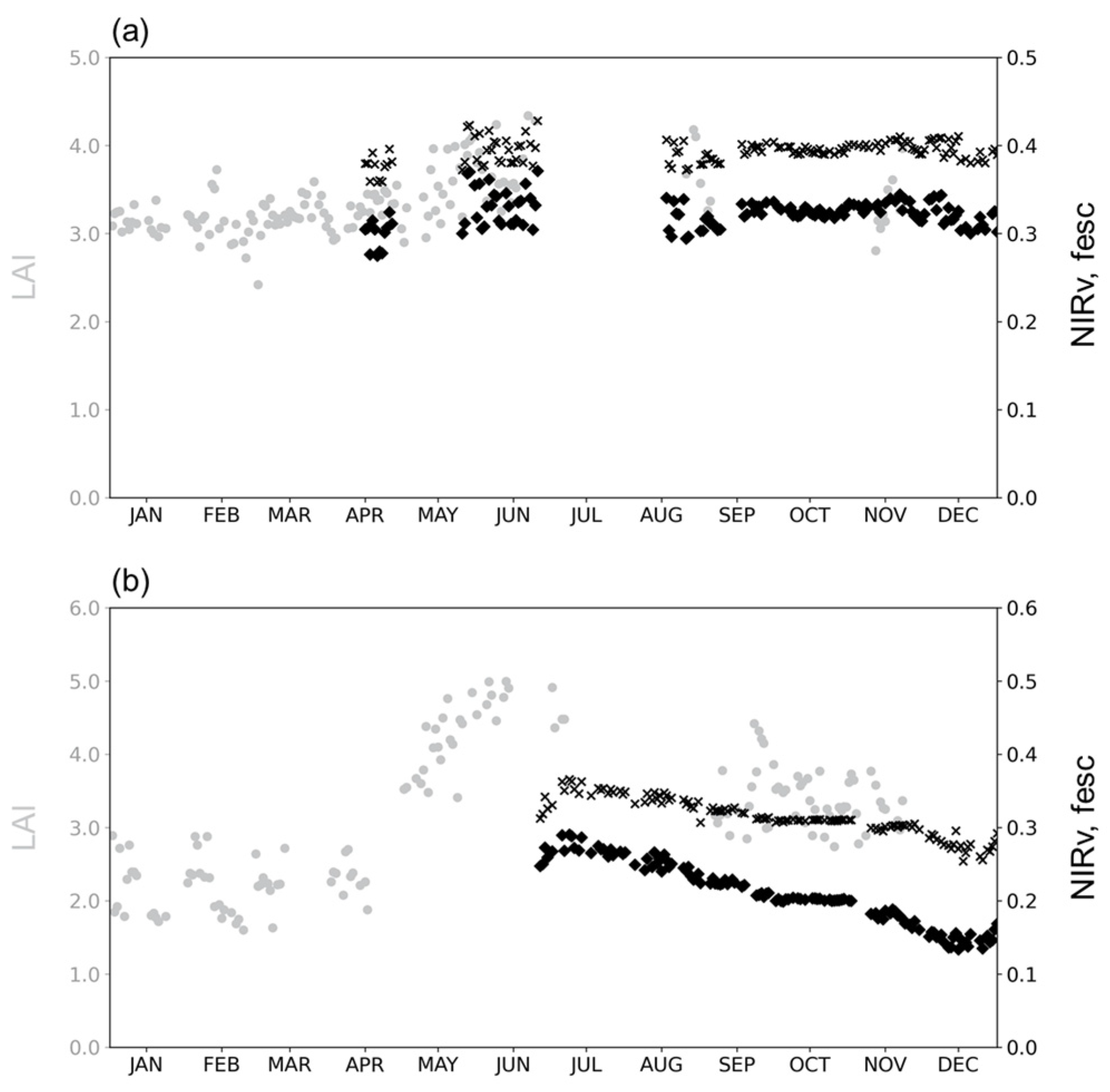
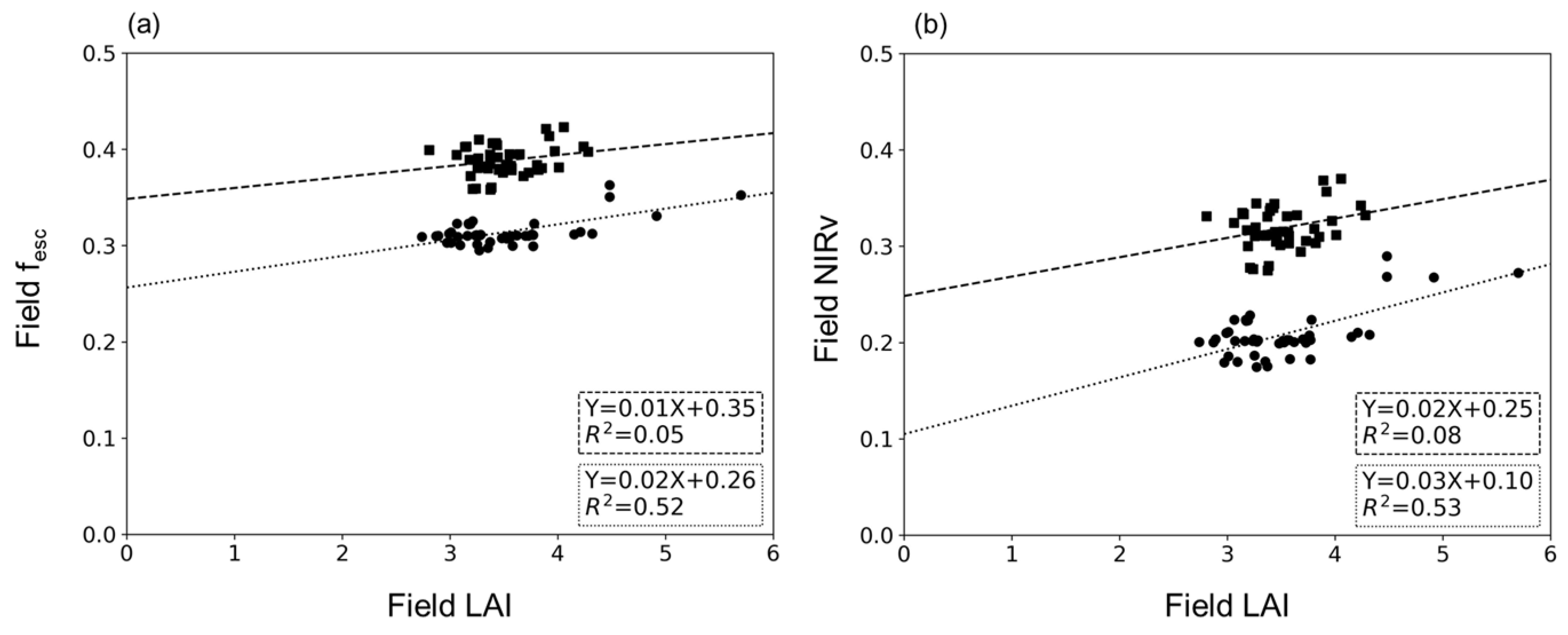
4.1. Relationship between LAI and fesc
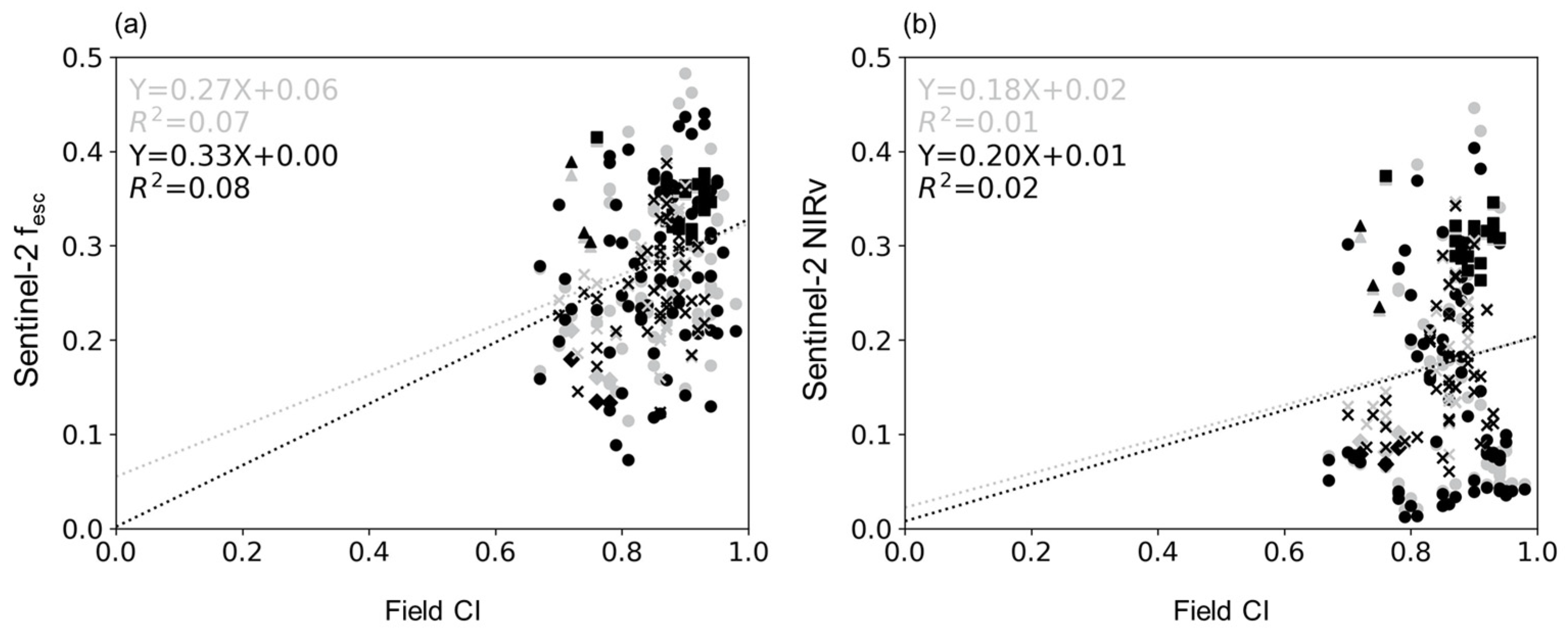
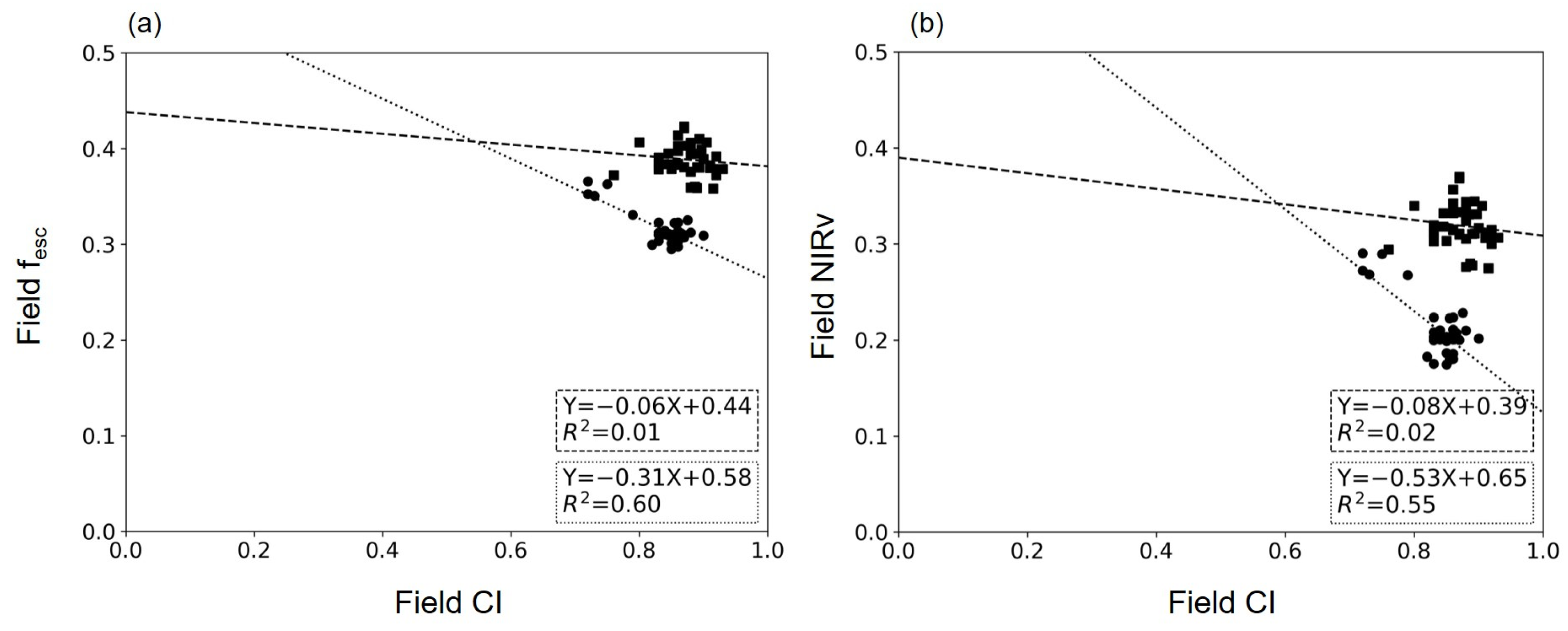
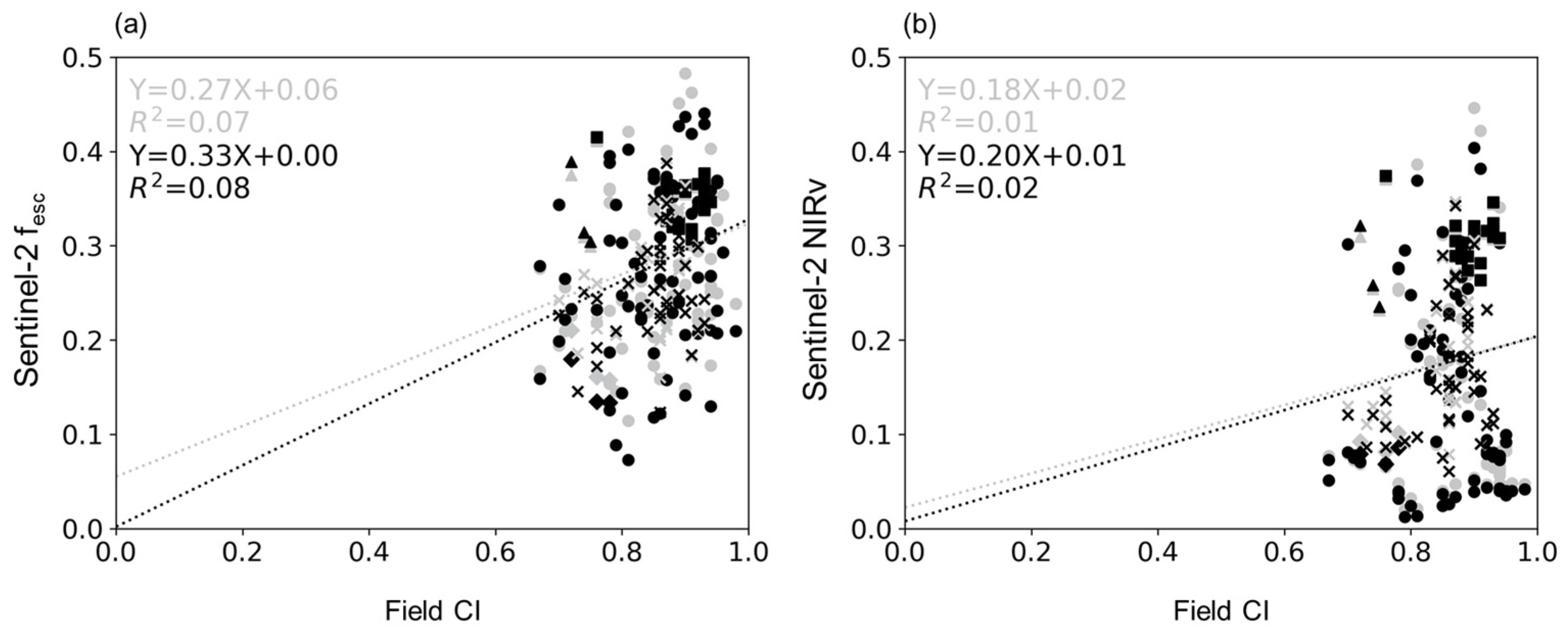
4.2. Relationship between CI and fesc
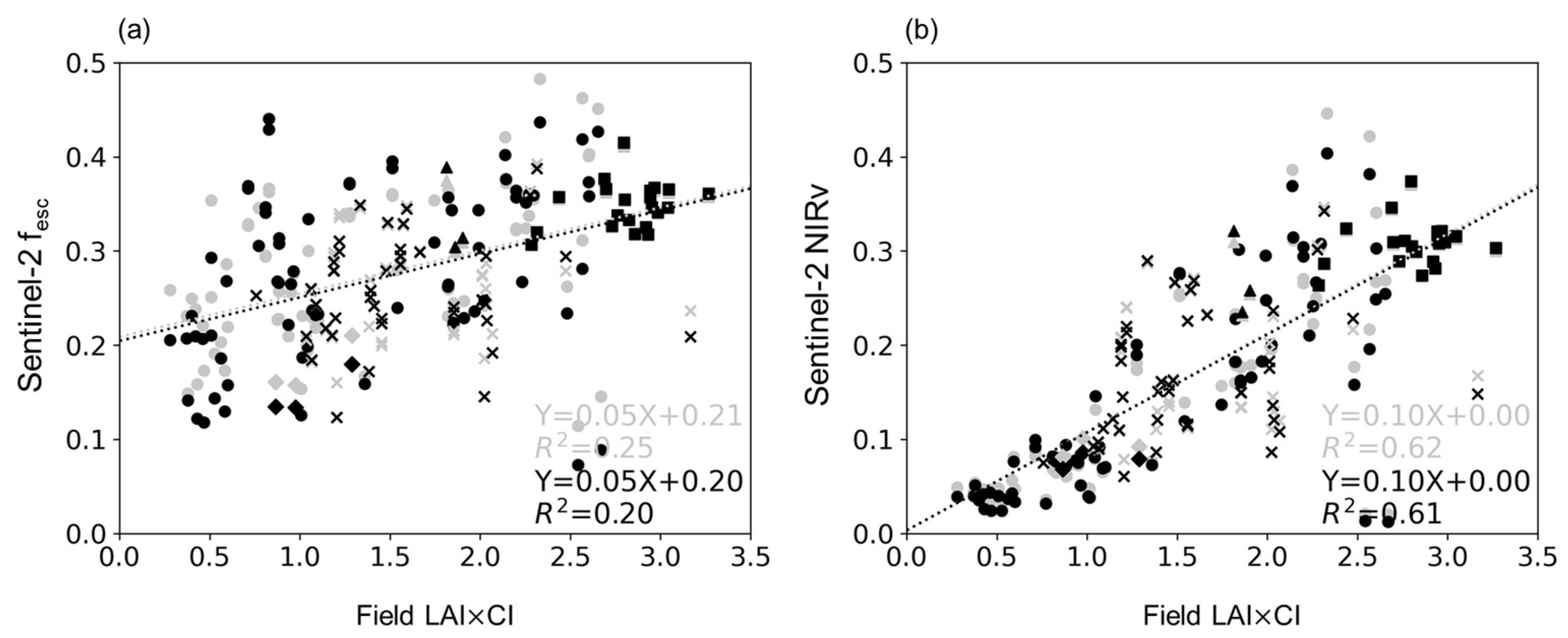
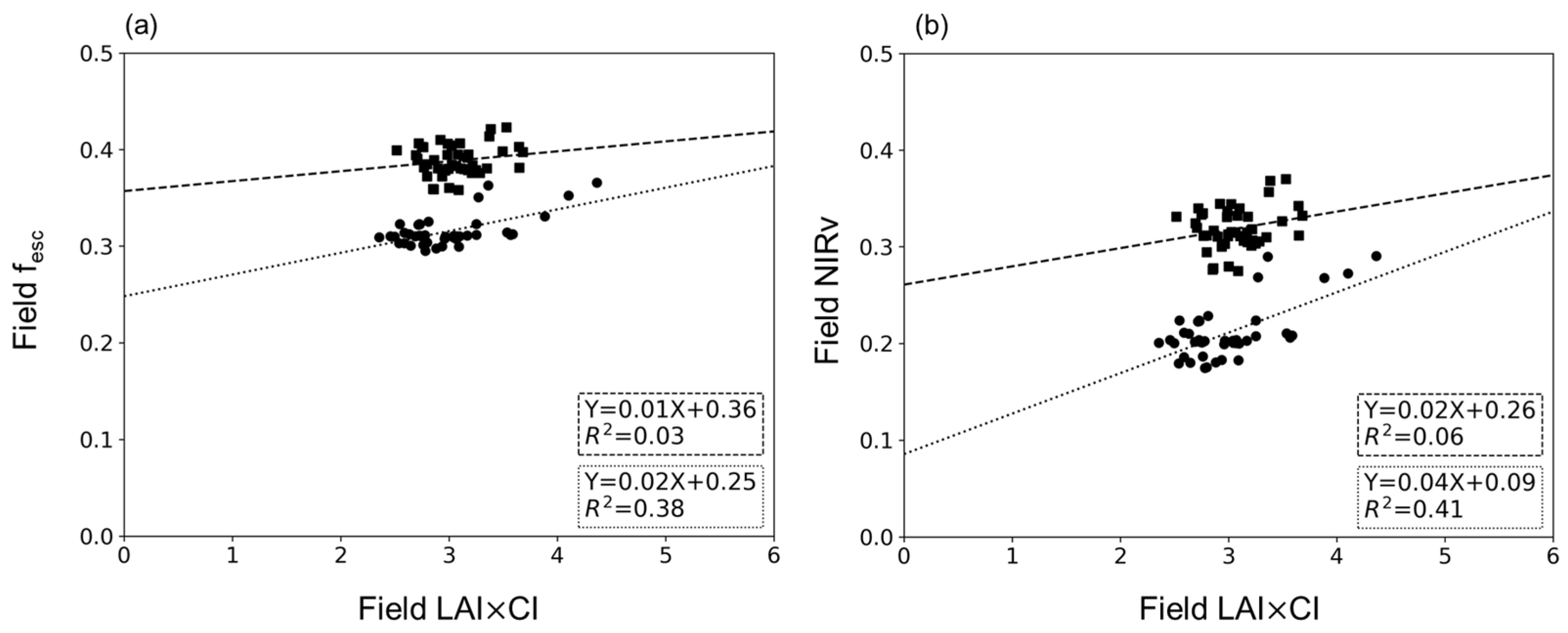
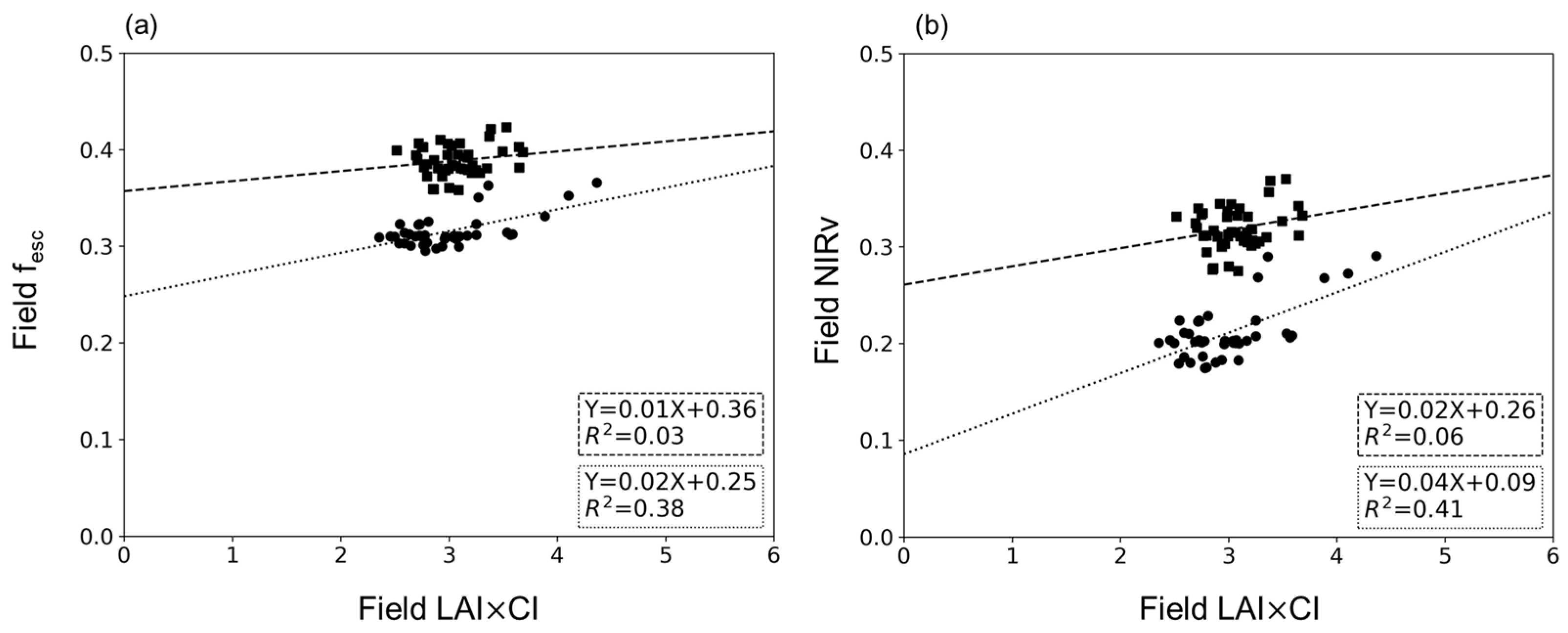
4.3. Relationship between LAI×CI and fesc
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Badgley, G.; Anderegg, L.D.L.; Berry, J.A.; Field, C.B. Terrestrial Gross Primary Production: Using NIRV to Scale from Site to Globe. Glob. Change Biol. 2019, 25, 3731–3740. [Google Scholar] [CrossRef]
- Zeng, Y.; Badgley, G.; Dechant, B.; Ryu, Y.; Chen, M.; Berry, J.A. A Practical Approach for Estimating the Escape Ratio of Near-Infrared Solar-Induced Chlorophyll Fluorescence. Remote Sens. Environ. 2019, 232, 111209. [Google Scholar] [CrossRef]
- Zhu, W.; Xie, Z.; Zhao, C.; Zheng, Z.; Qiao, K.; Peng, D.; Fu, Y.H. Remote Sensing of Terrestrial Gross Primary Productivity: A Review of Advances in Theoretical Foundation, Key Parameters and Methods. GISci. Remote Sens. 2024, 61, 2318846. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, Y.; Ju, W.; Qiu, B.; Zhang, Z. Tracking the Seasonal and Inter-Annual Variations of Global Gross Primary Production during Last Four Decades Using Satellite near-Infrared Reflectance Data. Sci. Total Environ. 2021, 755, 142569. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Guan, K.; Jiang, C.; Peng, B.; Kimm, H.; Chen, M.; Yang, X.; Wang, S.; Suyker, A.E.; Bernacchi, C.J.; et al. Radiance-Based NIRv as a Proxy for GPP of Corn and Soybean. Environ. Res. Lett. 2020, 15, 34009. [Google Scholar] [CrossRef]
- Dechant, B.; Ryu, Y.; Badgley, G.; Köhler, P.; Rascher, U.; Migliavacca, M.; Zhang, Y.; Tagliabue, G.; Guan, K.; Rossini, M.; et al. NIRVP: A Robust Structural Proxy for Sun-Induced Chlorophyll Fluorescence and Photosynthesis across Scales. Remote Sens. Environ. 2022, 268, 112763. [Google Scholar] [CrossRef]
- Jin, J.; Hou, W.; Ma, X.; Wang, H.; Xie, Q.; Wang, W.; Zhu, Q.; Fang, X.; Zhou, F.; Liu, Y.; et al. Improved Estimation of Gross Primary Production with NIRvP by Incorporating a Phenophase Scheme for Temperate Deciduous Forest Ecosystems. For. Ecol. Manage 2024, 556, 121742. [Google Scholar] [CrossRef]
- Huang, X.; Xiao, J.; Ma, M. Evaluating the Performance of Satellite-Derived Vegetation Indices for Estimating Gross Primary Productivity Using FLUXNET Observations across the Globe. Remote Sens. 2019, 11, 1823. [Google Scholar] [CrossRef]
- Yang, R.; Wang, J.; Zeng, N.; Sitch, S.; Tang, W.; McGrath, M.J.; Cai, Q.; Liu, D.; Lombardozzi, D.; Tian, H.; et al. Divergent Historical GPP Trends among State-of-the-Art Multi-Model Simulations and Satellite-Based Products. Earth Syst. Dyn. 2022, 13, 833–849. [Google Scholar] [CrossRef]
- McMahon, D.E.; Jackson, R.B. Management Intensification Maintains Wood Production over Multiple Harvests in Tropical Eucalyptus Plantations. Ecol. Appl. 2019, 29, e01879. [Google Scholar] [CrossRef]
- Tang, L.; Zhang, S.; Zhang, J.; Liu, Y.; Bai, Y. Estimating Evapotranspiration Based on the Satellite-Retrieved near-Infrared Reflectance of Vegetation (NIRv) over Croplands. GISci. Remote Sens. 2021, 58, 889–913. [Google Scholar] [CrossRef]
- Merrick, T.; Pau, S.; Detto, M.; Broadbent, E.N.; Bohlman, S.A.; Still, C.J.; Almeyda Zambrano, A.M. Unveiling Spatial and Temporal Heterogeneity of a Tropical Forest Canopy Using High-Resolution NIRv, FCVI, and NIRvrad from UAS Observations. Biogeosciences 2021, 18, 6077–6091. [Google Scholar] [CrossRef]
- Baldocchi, D.D.; Ryu, Y.; Dechant, B.; Eichelmann, E.; Hemes, K.; Ma, S.; Sanchez, C.R.; Shortt, R.; Szutu, D.; Valach, A.; et al. Outgoing Near-Infrared Radiation From Vegetation Scales With Canopy Photosynthesis Across a Spectrum of Function, Structure, Physiological Capacity, and Weather. J. Geophys. Res. Biogeosci. 2020, 125, e2019JG005534. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, R.; Yin, G.; Wang, C.; Liu, G.; Verger, A.; Descals, A.; Filella, I.; Peñuelas, J. Divergent Performances of Vegetation Indices in Extracting Photosynthetic Phenology for Northern Deciduous Broadleaf Forests. IEEE Geosci. Remote Sens. Lett. 2022, 19, 1–5. [Google Scholar] [CrossRef]
- Ersi, C.; Bayaer, T.; Bao, Y.; Bao, Y.; Yong, M.; Lai, Q.; Zhang, X.; Zhang, Y. Comparison of Phenological Parameters Extracted from SIF, NDVI and NIRv Data on the Mongolian Plateau. Remote Sens. 2022, 15, 187. [Google Scholar] [CrossRef]
- Chen, R.; Yin, G.; Liu, G.; Yang, Y.; Wang, C.; Xie, Q.; Zhao, W.; Verger, A. Correction of Illumination Effects on Seasonal Divergent NIRv Photosynthetic Phenology. Agric. For. Meteorol. 2023, 339, 109542. [Google Scholar] [CrossRef]
- Zhang, J.; Xiao, J.; Tong, X.; Zhang, J.; Meng, P.; Li, J.; Liu, P.; Yu, P. NIRv and SIF Better Estimate Phenology than NDVI and EVI: Effects of Spring and Autumn Phenology on Ecosystem Production of Planted Forests. Agric. For. Meteorol. 2022, 315, 108819. [Google Scholar] [CrossRef]
- Qiu, R.; Li, X.; Han, G.; Xiao, J.; Ma, X.; Gong, W. Monitoring Drought Impacts on Crop Productivity of the US Midwest with Solar-Induced Fluorescence: GOSIF Outperforms GOME-2 SIF and MODIS NDVI, EVI, and NIRv. Agric. For. Meteorol. 2022, 323, 109038. [Google Scholar] [CrossRef]
- Zeng, Y.; Chen, M.; Hao, D.; Damm, A.; Badgley, G.; Rascher, U.; Johnson, J.E.; Dechant, B.; Siegmann, B.; Ryu, Y.; et al. Combining Near-Infrared Radiance of Vegetation and Fluorescence Spectroscopy to Detect Effects of Abiotic Changes and Stresses. Remote Sens. Environ. 2022, 270, 112856. [Google Scholar] [CrossRef]
- Gu, H.; Yin, G.; Yang, Y.; Verger, A.; Descals, A.; Filella, I.; Zeng, Y.; Hao, D.; Xie, Q.; Li, X.; et al. Satellite-Detected Contrasting Responses of Canopy Structure and Leaf Physiology to Drought. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2023, 16, 2427–2436. [Google Scholar] [CrossRef]
- Chen, J.; Liu, X.; Du, S.; Ma, Y.; Liu, L. Effects of Drought on the Relationship between Photosynthesis and Chlorophyll Fluorescence for Maize. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2021, 14, 11148–11161. [Google Scholar] [CrossRef]
- Sakai, Y.; Kobayashi, H.; Kato, T. FLiES-SIF Version 1.0: Three-Dimensional Radiative Transfer Model for Estimating Solar Induced Fluorescence. Geosci. Model Dev. 2020, 13, 4041–4066. [Google Scholar] [CrossRef]
- Magney, T.S.; Barnes, M.L.; Yang, X. On the Covariation of Chlorophyll Fluorescence and Photosynthesis across Scales. Geophys. Res. Lett. 2020, 47, e2020GL091098. [Google Scholar] [CrossRef]
- Maguire, A.J.; Eitel, J.U.H.; Magney, T.S.; Frankenberg, C.; Köhler, P.; Orcutt, E.L.; Parazoo, N.C.; Pavlick, R.; Pierrat, Z.A. Spatial Covariation between Solar-Induced Fluorescence and Vegetation Indices from Arctic-Boreal Landscapes. Environ. Res. Lett. 2021, 16, 95002. [Google Scholar] [CrossRef]
- Liu, W.; Luo, S.; Lu, X.; Atherton, J.; Gastellu-Etchegorry, J.-P. Simulation-Based Evaluation of the Estimation Methods of Far-Red Solar-Induced Chlorophyll Fluorescence Escape Probability in Discontinuous Forest Canopies. Remote Sens. 2020, 12, 3962. [Google Scholar] [CrossRef]
- Liu, X.; Guanter, L.; Liu, L.; Damm, A.; Malenovský, Z.; Rascher, U.; Peng, D.; Du, S.; Gastellu-Etchegorry, J.-P. Downscaling of Solar-Induced Chlorophyll Fluorescence from Canopy Level to Photosystem Level Using a Random Forest Model. Remote Sens. Environ. 2019, 231, 110772. [Google Scholar] [CrossRef]
- Yang, P.; van der Tol, C. Linking Canopy Scattering of Far-Red Sun-Induced Chlorophyll Fluorescence with Reflectance. Remote Sens. Environ. 2018, 209, 456–467. [Google Scholar] [CrossRef]
- Filella, I.; Descals, A.; Balzarolo, M.; Yin, G.; Verger, A.; Fang, H.; Peñuelas, J. Photosynthetically Active Radiation and Foliage Clumping Improve Satellite-Based NIRv Estimates of Gross Primary Production. Remote Sens. 2023, 15, 2207. [Google Scholar] [CrossRef]
- Lee, J.; Cha, S.; Lim, J.; Chun, J.; Jang, K. Practical LAI Estimation with DHP Images in Complex Forest Structure with Rugged Terrain. Forests 2023, 14, 2047. [Google Scholar] [CrossRef]
- Chianucci, F.; Macek, M. HemispheR: An R Package for Fisheye Canopy Image Analysis. Agric. For. Meteorol. 2023, 336, 109470. [Google Scholar] [CrossRef]
- Chianucci, F.; Zou, J.; Leng, P.; Zhuang, Y.; Ferrara, C. A New Method to Estimate Clumping Index Integrating Gap Fraction Averaging with the Analysis of Gap Size Distribution. Can. J. For. Res. 2019, 49, 471–479. [Google Scholar] [CrossRef]
- Beck, H.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Lutsko, N.J.; Dufour, A.; Zeng, Z.; Jiang, X.; van Dijk, A.I.J.M.; Miralles, D.G. High-Resolution (1 Km) Köppen-Geiger Maps for 1901–2099 Based on Constrained CMIP6 Projections. Sci. Data 2023, 10, 724. [Google Scholar] [CrossRef] [PubMed]
- Badgley, G.; Field, C.B.; Berry, J.A. Canopy Near-Infrared Reflectance and Terrestrial Photosynthesis. Sci. Adv. 2017, 3, e1602244. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Z.; Chen, S.; Yin, T.; Gastellu-Etchegorry, J.-P. Globally Quantitative Analysis of the Impact of Atmosphere and Spectral Response Function on 2-Band Enhanced Vegetation Index (EVI2) over Sentinel-2 and Landsat-8. ISPRS J. Photogramm. Remote Sens. 2023, 205, 206–226. [Google Scholar] [CrossRef]
- Chen, R.; Yin, G.; Zhao, W.; Xu, B.; Zeng, Y.; Liu, G.; Verger, A. TCNIRv: Topographically Corrected Near-Infrared Reflectance of Vegetation for Tracking Gross Primary Production Over Mountainous Areas. IEEE Trans. Geosci. Remote Sens. 2022, 60, 1–10. [Google Scholar] [CrossRef]
- Weiss, M.; Baret, F.; Jay, S. S2ToolBox Level 2 Products: LAI, FAPAR, FCOVER; Research for Agriculture, Food and Environment (INRAE): Avignon, France, 2016. [Google Scholar]
- Ma, X.; Huete, A.; Yu, Q.; Restrepo-Coupe, N.; Beringer, J.; Hutley, L.B.; Kanniah, K.D.; Cleverly, J.; Eamus, D. Parameterization of an Ecosystem Light-Use-Efficiency Model for Predicting Savanna GPP Using MODIS EVI. Remote Sens. Environ. 2014, 154, 253–271. [Google Scholar] [CrossRef]
- Liang, S.; Ma, W.; Sui, X.; Wang, M.; Li, H. An Assessment of Relations between Vegetation Green FPAR and Vegetation Indices through a Radiative Transfer Model. Plants 2023, 12, 1927. [Google Scholar] [CrossRef] [PubMed]
- Regaieg, O.; Yin, T.; Malenovský, Z.; Cook, B.D.; Morton, D.C.; Gastellu-Etchegorry, J.P. Assessing Impacts of Canopy 3D Structure on Chlorophyll Fluorescence Radiance and Radiative Budget of Deciduous Forest Stands Using DART. Remote Sens. Environ. 2021, 265, 112673. [Google Scholar] [CrossRef]
- Qi, M.; Liu, X.; Du, S.; Guan, L.; Chen, R.; Liu, L. Improving the Estimation of Canopy Fluorescence Escape Probability in the Near-Infrared Band by Accounting for Soil Reflectance. Remote Sens. 2023, 15, 4361. [Google Scholar] [CrossRef]
- Lu, X.; Liu, Z.; Zhao, F.; Tang, J. Comparison of Total Emitted Solar-Induced Chlorophyll Fluorescence (SIF) and Top-of-Canopy (TOC) SIF in Estimating Photosynthesis. Remote Sens. Environ. 2020, 251, 112083. [Google Scholar] [CrossRef]
- Brown, L.A.; Meier, C.; Morris, H.; Pastor-Guzman, J.; Bai, G.; Lerebourg, C.; Gobron, N.; Lanconelli, C.; Clerici, M.; Dash, J. Evaluation of Global Leaf Area Index and Fraction of Absorbed Photosynthetically Active Radiation Products over North America Using Copernicus Ground Based Observations for Validation Data. Remote Sens. Environ. 2020, 247, 111935. [Google Scholar] [CrossRef]
- Huang, D.; Knyazikhin, Y.; Dickinson, R.E.; Rautiainen, M.; Stenberg, P.; Disney, M.; Lewis, P.; Cescatti, A.; Tian, Y.; Verhoef, W.; et al. Canopy Spectral Invariants for Remote Sensing and Model Applications. Remote Sens. Environ. 2007, 106, 106–122. [Google Scholar] [CrossRef]
- Zeng, Y.; Badgley, G.; Chen, M.; Li, J.; Anderegg, L.D.L.; Kornfeld, A.; Liu, Q.; Xu, B.; Yang, B.; Yan, K.; et al. A Radiative Transfer Model for Solar Induced Fluorescence Using Spectral Invariants Theory. Remote Sens. Environ. 2020, 240, 111678. [Google Scholar] [CrossRef]
- Smolander, S.; Stenberg, P. Simple Parameterizations of the Radiation Budget of Uniform Broadleaved and Coniferous Canopies. Remote Sens. Environ. 2005, 94, 355–363. [Google Scholar] [CrossRef]
- Roy, D.P.; Li, J.; Zhang, H.K.; Yan, L.; Huang, H.; Li, Z. Examination of Sentinel-2A Multi-Spectral Instrument (MSI) Reflectance Anisotropy and the Suitability of a General Method to Normalize MSI Reflectance to Nadir BRDF Adjusted Reflectance. Remote Sens. Environ. 2017, 199, 25–38. [Google Scholar] [CrossRef]
- Galvão, L.S.; Arlanche Petri, C.; Dalagnol, R. Coupled Effects of Solar Illumination and Phenology on Vegetation Index Determination: An Analysis over the Amazonian Forests Using the SuperDove Satellite Constellation. GISci. Remote Sens. 2024, 61, 2290354. [Google Scholar] [CrossRef]
- Putzenlechner, B.; Castro, S.; Kiese, R.; Ludwig, R.; Marzahn, P.; Sharp, I.; Sanchez-Azofeifa, A. Validation of Sentinel-2 FAPAR Products Using Ground Observations across Three Forest Ecosystems. Remote Sens. Environ. 2019, 232, 111310. [Google Scholar] [CrossRef]
- Estévez, J.; Vicent, J.; Rivera-Caicedo, J.P.; Morcillo-Pallarés, P.; Vuolo, F.; Sabater, N.; Camps-Valls, G.; Moreno, J.; Verrelst, J. Gaussian Processes Retrieval of LAI from Sentinel-2 Top-of-Atmosphere Radiance Data. ISPRS J. Photogramm. Remote Sens. 2020, 167, 289–304. [Google Scholar] [CrossRef]





| Band | SD-600 | Sentinel-2 | ||
|---|---|---|---|---|
| Central Wavelength (nm) | Bandwidth (nm) | Central Wavelength (nm) | Bandwidth (nm) | |
| Blue | 475 | 30 | 492.4 | 66 |
| Green | 550 | 35 | 559.8 | 36 |
| Red | 640 | 50 | 664.6 | 31 |
| Red Edge | 690 | 55 | 705 | 15 |
| NIR | 855 | 54 | 832.8 | 106 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.; Im, J.; Lim, J.; Kim, K. Insights into Canopy Escape Ratio from Canopy Structures: Correlations Uncovered through Sentinel-2 and Field Observation. Forests 2024, 15, 665. https://doi.org/10.3390/f15040665
Lee J, Im J, Lim J, Kim K. Insights into Canopy Escape Ratio from Canopy Structures: Correlations Uncovered through Sentinel-2 and Field Observation. Forests. 2024; 15(4):665. https://doi.org/10.3390/f15040665
Chicago/Turabian StyleLee, Junghee, Jungho Im, Joongbin Lim, and Kyungmin Kim. 2024. "Insights into Canopy Escape Ratio from Canopy Structures: Correlations Uncovered through Sentinel-2 and Field Observation" Forests 15, no. 4: 665. https://doi.org/10.3390/f15040665
APA StyleLee, J., Im, J., Lim, J., & Kim, K. (2024). Insights into Canopy Escape Ratio from Canopy Structures: Correlations Uncovered through Sentinel-2 and Field Observation. Forests, 15(4), 665. https://doi.org/10.3390/f15040665